
Abstract
Nitrogen (N) availability in soil regulates microbial communities and then affects the decomposition and formation of microbial residues carbon (C), which have great impacts on soil organic carbon (SOC) sequestration. However, Asia has not yet well-assessed patterns and determinants of microbial residues C in response to N input. Here, we performed a large-scale, systematic meta-analysis of the effects of N addition on microbial residues across cropland and forest ecosystems. A total of 31 publications regarding microbial residues were included in our database, with 235 observations. In croplands N addition significantly increased microbial residue and biomass, and total microbial residue contribution to SOC. The responses to N addition of microbial residues and biomass increased with N addition rate (0–800 kg N ha−1 year−1) and there was a bidirectional positive effect between these two. In forest ecosystems, bacterial residues increased by 14.7% in N addition rates of 0–50 kg N ha−1 year−1. Moreover, the negative correlation between the duration of N addition and microbial residues is highly significant, indicating that long-term N deposition could threaten SOC transformation and sequestration in forests. These results suggest that N addition effects on microbial residues are ecosystem-specific. This is related to the different main controlling factors affecting the microbial residues in cropland and forest ecosystems under N addition.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Over the past century, anthropogenic activities, such as fossil fuel combustion and intensive agriculture production, have substantially increased the amount of N deposited in natural ecosystems and directly supplied to managed land (Penuelas et al. 2013). Increased soil N availability can dramatically affect ecosystem processes (Treseder 2008; Yu et al. 2019), and a more detailed comprehension of the ecosystem response to N input helps to manage the potential of ecosystems to sequester C (Bragazza et al. 2012). Soil N availability can alter microbial activity, abundance, and community composition has been well-documented (Jiang et al. 2021; Treseder 2008). Therefore, N addition will affect the soil C turnover processes of microorganisms (Kätterer et al. 2012). Microbial-mediated C processes are the foundation for appreciating the dynamic mechanism of SOC since microbes play pivotal roles in regulating soil C cycling (Schimel and Schaeffer 2012; Liang et al. 2017). However, if the contribution of microorganisms to the formation of SOC is evaluated only in terms of microbial biomass, this contribution is seriously underestimated (Simpson et al. 2007; Liang et al. 2017).
Soil microbial residues are a major component of stable SOC, which play a central role in long-term soil C sequestration (Lehmann and Kleber 2015). Wang et al. (2021) based on the comprehensive analysis of the contribution of microbial residues to SOC in different ecosystems, concluded that the contribution of microbial residues was 35%–51%, indicating that the amount of organic matter transported by microorganisms to the soil through the form of cell residues was much higher than the traditional understanding. Amino sugars are components of microbial cell walls and are stabilized in the soil after cell death (Zhang and Amelung 1996). It can be used as an important marker of microbial residues to study the response of microbial residues to environmental changes (e.g., external N input). Specifically, muramic acid is a marker of bacterial residue (Joergensen 2018). Glucosamine was found to be the main component of chitin in fungal cell walls (Engelking et al. 2007). Bacterial and fungal residues can be used to assess the contribution of fungi and bacteria to SOC.
Given the recent boom in research on the effects of N addition on microbial residues, it allows us to compile Asian data to explore the response of soil microbial residues to different N addition regimes within various ecosystems, and further find out the fundamental factors controlling microbial-driven SOC sequestration process. Few meta-analysis, to date, is synthesized to investigate the responses and controls of microbial residues under N addition context across different ecosystems (Liao et al. 2020; Hu et al. 2022). However, these studies have focused on how the response of microbial residues to N addition depends on the N addition rate, type, duration of N addition, and ecosystem type with less attention to the relationship between microbial residues and microbial biomass under N addition. In this paper, we synthesized 235 observations across two ecosystem types characterized by different soil environments (i.e., SOC, TN, pH, C:N, and soil texture) and climate factors (Annual mean temperature and annual mean temperature) and quantitatively evaluated N addition effects on soil microbial residues and the difference in their contribution to SOC in forest and cropland ecosystems.
The aims of the study were to (a) identify the overall trends of N effects on soil microbial residues and microbial biomass across forest and cropland ecosystems, (b) reveal the relationship between microbial residues and microbial biomass in forest and cropland ecosystems under N addition, and (c) to characterize the biotic and abiotic factors controlling the response of microbial residues to N addition. To address these aims, we propose the following two scientific hypotheses: (a) due to differences in N utilization (croplands > forests) and physicochemical protection (less clay protection in forests), the accumulation effect of N addition on microbial residue C and microbial biomass differed between ecosystems, with the accumulation effect of microbial residue C and microbial biomass higher in croplands than in forests, (b) in addition differences in substrate availability (in vivo turnover of microbial is dominant in croplands, while in ex vivo modification in forests is dominant), the correlation between microbial residue C and microbial biomass were higher in croplands than in forests under N addition. It is expected to provide a more comprehensive and mechanistic understanding of N addition effects on microbial residue accumulation and soil organic matter formation.
2 Materials and Methods
2.1 Data Collection
We searched peer-reviewed publications regarding the effects of N addition/deposition on soil microbial residues through the Web of Science (http://apps.webofknowledge.com) and China National Knowledge Infrastructure (CNKI, http://www.cnki.net). “(fertilization OR nitrogen OR nutrient) AND (deposition OR input OR addition OR application OR enrichment) AND (fungal residues OR bacterial residues OR amino sugar OR microbial residues OR microbial necromass)” were keywords and terms that were used in literature retrieval. The literature search was limited to peer-reviewed papers published between June 2009 and October 2021. The retrieved literature was scrutinized according to the following criteria to obtain data meeting the requirements of meta-analysis: (i) the publication must include at least one of the target variables of amino sugars or microbial residues, such as glucosamine and muramic acid; (ii) the rate, nutrient type, and form of N addition were reported; (iii) experiment included at least one pair of treatments (control and N addition treatment), which can be extracted directly from publication for mean, standard deviation (SD), or standard error (SE), and sample size (n); (iv) all treatments and control must be established within the same time and space; (v) in order to effectively compare the results of different studies, we only included the amino sugar data determined by the method of Zhang and Amelung (1996). Microbial biomass data, such as microbial biomass C (MBC) measured by chloroform fumigation-extraction technique or microbial biomass measured by phospholipid fatty acid analysis (PLFA), were also recorded in selected publications to examine their relationships with microbial residue data. The microbial biomass data measured by these two methods are expressed by MBC and PLFA respectively. Based on the above criteria, a total of 31 publications regarding microbial residues were incorporated into our database with 235 observations. Of those, 113 were field observations and 122 were laboratory observations. 192 observations (81.7%) were from croplands and 43 (18.3%) from forests. It is worth noting that the low sample size (3 observations) of grassland ecosystems in the screened studies led to limitations in our results, which need to be addressed in future studies.
For better comparison, (1) we categorized the data into cropland ecosystem and forest ecosystem types; (2) the forest ecosystems were further binned by 50 kg N ha−1 year−1of N addition rate intervals, i.e., 0–50 kg N ha−1 year−1 and 50–100 kg N ha−1 year−1. In addition, SOC, soil potential of hydrogen (pH), soil total nitrogen (TN), C: N, clay, duration (duration of the N addition) and mean annual temperature (MAT), and mean annual precipitation (MAP) were also collected from cited papers.
In the process of data collection, if the data in the original papers are presented in the form of graphs, Engauge Digitizer software was used to obtain the data. Raw data presented in tabular form is obtained directly. If the SE were reported, then we used the formula \(\mathrm{SD}=\mathrm{SE}\sqrt{n}\) to calculate SD. Microbial residue C was estimated based on amino sugar data. Bacterial and fungal residue C were calculated using the following Eqs. (1) and (2), respectively (Appuhn and Joergensen 2006; Engelking et al. 2007).
45 is the conversion coefficient of muramic acid to bacterial residue C, 9 is the conversion coefficient of glucosamine to fungal residue C, 179 and 251 are the molecular weight of glucosamine and muramic acid, respectively. Total microbial residue C was estimated as the sum of fungal and bacterial residue C (Liang et al. 2019).
2.2 Data analysis
The statistical analysis was performed using the meta-analysis (Hedges et al. 1999) in OpenMEE. The response of microbial residues and microbial biomass to N addition was investigated quantitatively by meta-analysis. In each study, the N addition effect was estimated as the natural logarithm transformed (ln) response ratio (RR) with the following equation:
where \(\overline{{X }_{t}}\) and \(\overline{{X }_{c}}\) are the mean values of a concerned variable in N addition treatment and control, respectively. Subgroup meta-analysis was performed using OpenMEE software to calculate the “mean effect size” (E++) and 95% bootstrap confidence interval (CI). The iterations were 999. To account for any potential deviations, the restricted maximum likelihood (REML) method in the random-effects model was used to calculate the results. When the mean effect size of 95% CI value did not overlap with zero, the effect of each variable was significantly different. For a better explanation, the “mean effect size” (E++) is converted to a percentage change calculated according to the following formula: [exp (E++)-1] × 100%. The fail-safe N Rosenthal number and funnel plot suggested that there was no publication bias for the influences of N addition on variables (Table S1, P < 0.0001, Fig. S1). Overall, our results were robust (Fig. 1).

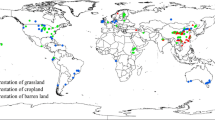
Locations of studies regarding the effects of nitrogen (N) addition on soil microbial residues included in this meta-analysis
Redundancy analysis (RDA) and regression analysis were conducted in the R program (version 3.1.1). The RDA analysis was handled in R with the “vegan” package. To determine the relationship between biotic and abiotic factors with RR of microbial residues, the “trendline” function in R was used for regression analysis.
3 Results
3.1 Microbial Residue C and Microbial Biomass
N addition significantly increased total, fungal, and bacterial residues C and total residue C/SOC but had no significant effect on the fungal/bacterial residue C. The effect of N addition on microbial residues C varies with ecosystem types. In cropland ecosystems, we found that the total, fungal, and bacterial residues C and total residue C/SOC increased significantly by 17.1%, 14%, 12.5%, and 8.4% respectively. In forest ecosystems, bacterial residue C increased significantly by 13.3% while fungal/bacterial residue C decreased significantly by 12.3% (Fig. 2).

Mean effect sizes and 95% confidence intervals (CIs) of total residue carbon (C), fungal residue C, bacterial residue C, fungal/bacterial residue C, and fungal residue C/soil organic C after nitrogen (N) addition in croplands and forests. The number in parentheses is the number of observations in each group and asterisks represent significant differences from zero (P < 0.05)
Total, fungal, and bacterial PLFAs were increased by 37.9%, 51.3%, and 51.3% respectively under N addition in cropland ecosystems. In forest ecosystems, total PLFA and bacterial PLFA were decreased by 9.5% and 9.2% under N addition, respectively, while fungal PLFA did not respond significantly. There were no significant differences in fungal/bacterial PLFA under N addition in forest and cropland ecosystems (Fig. 3).

Mean effect sizes and 95% confidence intervals (CIs) of total phospholipid fatty acid analysis (PLFA), fungal PLFA, bacterial PLFA, and fungal/bacterial PLFA after nitrogen (N) addition in croplands and forests. The number in parentheses is the number of observations in each group and asterisks represent significant differences from zero (P < 0.05)
In cropland ecosystems, total, fungal and bacterial residues C increased significantly with N addition rate. This also held true for MBC, fungal and bacterial PLFAs with an increased N addition rate (P < 0.01, Fig. 4a–f). Distinct response sizes of MBC and total residue C to N addition rates showed to an irregularly nonlinear relationship between RR of MBC/RR of total residue C and N addition rate. Specifically, RR of MBC/RR of total residue C decreased first with an increased N addition rate and then increased when N was added at a rate of 500–800 kg N ha−1 year−1 (P < 0.05, Fig. 5). Across all observations in forest ecosystems, only bacterial residues were observed a significant increase (14.7%) when N was added at a rate of 0–50 kg N ha−1 year−1. Total, fungal, and bacterial residues C under N addition at a rate of 0–100 kg N ha−1 year−1 were no significant differences compared to non-N addition in forest ecosystems (Fig. 6). Total, fungal and bacterial PLFAs were no significant response under N addition at a rate of 0–50 kg N ha−1 year−1, while total and bacterial PLFA decreased significantly by 18.3% and 17.9%, respectively, in forest ecosystems when N was added at rates of 50−100 kg N ha−1 year−1 (Fig. 6, P < 0.05).

Scatter plot with regression graph of response ratios (RRs) for total residue carbon (C, a), fungal residue C (b), bacterial residue C (c), microbial biomass C (MBC, d), fungal (e), and bacterial phospholipid fatty acid analysis (PLFA, f) versus nitrogen (N) addition rate (kg N ha−1 year−1) in cropland ecosystems

Scatter plot with regression graph of the ratio between response ratio (RR) of microbial biomass carbon (MBC) and the RR of total residual carbon (C) versus nitrogen (N) addition rate (kg N ha−1 year−1) in cropland ecosystems. The gray areas represent the 95% confidence interval of the fitting curve

Mean effect sizes and 95% confidence intervals (CIs) of total, fungal, and bacterial residues carbon (C) and total, fungal, and bacterial phospholipid fatty acid analysis (PLFAs) in response to nitrogen (N) addition with two N addition levels (0–50 and 50–100 kg N ha−1 year−.1) in forest ecosystems. The number in parentheses is the number of observations in each group and asterisks represent significant differences from zero (P < 0.05)
3.2 Controls of Microbial and Environmental Variables on Microbial Residues
The RR of MBC was positively correlated with that of total residual C in croplands (P < 0.0001, Fig. 7a). The RR of fungal PLFA was positively correlated with the RR of fungal residue C (P < 0.01, Fig. 7b). In cropland ecosystems, the RR of bacterial residue C increased with the increase of RR of bacterial PLFA (P < 0.001, Fig. 7c). In addition, we found that there is no significant relationship between microbial residues C and biomass in forest ecosystems after N addition (Fig. 7d–f).

Relationship between response ratio (RR) of total residue carbon (C, a), RR of fungal residue C (b), RR of bacterial residue C (c) and RR of microbial biomass C (MBC), RR of fungal phospholipid fatty acid analysis (PLFA), RR of bacterial PLFA in cropland ecosystems, and the relationship between RR of total residue C (d), RR of fungal residue C (e), RR of bacterial residue C (f), and RR of total PLFA, RR of fungal PLFA, and RR of bacterial PLFA in forest ecosystems
RDA showed the relative effects of living microorganisms, soil properties, climatic factors, and N addition regimes on the RRs of total, fungal, and bacterial residues C under N addition in cropland and forest ecosystems (Fig. 8). The first two RDA axes explained 62.20% (RDA1) and 1.91% (RDA2) of the total variation, respectively, in croplands. N addition effects on total, fungal and bacterial residues C of croplands were significantly and positively correlated with microbial biomass (MBC, fungal PLFA, and bacterial PLFA), soil properties (SOC and pH), significantly positively correlated with TN and MAP, and weakly and significantly positively with duration (P < 0.05), and it was not correlated with soil C:N, clay, and MAT (P > 0.05, Fig. 8a). About forests, the first two RDA axes explained 60.52% (RDA1) and 3.79% (RDA2) of the total variation, respectively. N addition effects on microbial residue C were not significantly correlated with microbial biomass (MBC, fungal PLFA, and bacterial PLFA), soil properties (SOC, TN, C: N, and pH), and climatic factor (MAT and MAP) (P > 0.05, Fig. 8b). Clay showed a weakly significant positive correlation with microbial residues C, and duration showed a strongly significant positive correlation with microbial residues C. (P < 0.05, Fig. 8b).

Redundancy analysis for multivariate effects of microbial biomass, soil properties, climate factors, and the duration on response ratios of microbial residue C in croplands (a) and forests (b). Blue arrows indicate RRs of microbial residue C and red arrows indicate geochemical variables. *** represents P < 0.001, ** represents P < 0.01, and * represents P < 0.05. RDA, redundancy analysis; RR, response ratio; MBC, microbial biomass C; PLFA, phospholipid fatty acid analysis; pH, potential of hydrogen; C:N, the carbon to nitrogen ratio; TN, total nitrogen; SOC, soil organic carbon; MAT, mean annual temperature; MAP, mean annual precipitation
4 Discussion
4.1 The Effects of N Addition on Microbial Residues and Biomass in Cropland and Forest Ecosystems
N addition increased the content of microbial residue C and microbial biomass in cropland ecosystems (Fig. 2). This may be because the availability of N fertilizer in cropland reduces the reuse of microbial residue C by microorganisms. A positive effect of N addition on microbial residue proportions in total SOC pools (Fig. 2) suggests that N-induced changes in SOC storage are highly coupled with the altered microbial anabolism. Microbial residue C and microbial biomass increased with N addition rate (Fig. 4), indicating a strengthened microbial anabolism under higher N availability. Moreover, N addition can increase the primary production, thus increasing the aboveground and root residues of plants in the soil, which is conducive to the increase of microbial biomass and the accumulation of microbial residue C (Zhang et al. 2021). These findings confirm that soil N availability is critical in microbial growth and byproduct formation, such as microbial cell wall components (Cotrufo et al. 2013). The positive effect of N enrichment on microbes may be due to a variety of mechanisms, which stimulates the increase of microbial residues. In the case of soil containing more abundant easily degradable C input, the supply of N must be increased to meet the increased N demand for microbial growth (Mills et al. 2008). When N is available at high rates, microbes may distribute more C to grow due to the lower metabolic expense of acquiring N (Spohn et al. 2016), consequently resulting in more microbial metabolites.
N addition had little effect on total and fungal residues C and significantly increased bacterial residue C in forest ecosystems (Fig. 2), while total PLFA and bacterial PLFA decreased significantly at higher N availability (Figs. 3, 6). It is worth noting that there was no significant relationship between microbial residue C and microbial biomass in forest ecosystems under N addition (Fig. 7d–f). Compared to croplands, forest soils, especially tropical/subtropical forests, are mostly enriched in N, and continuous N addition results in a decrease in pH (Li et al. 2015), which reduces microbial biomass but have less impact on microbial residues (Zhang et al. 2016). When N availability is high, microorganisms might allocate more C to growth due to lower metabolic costs of N acquisition (Manzoni et al. 2012). Thus, the iterative accumulation process of microbial proliferation-metabolism-death weeks results in the production of more microbial residues. According to the ecological stoichiometry theory, the growth of microorganisms needs an appropriate element stoichiometry ratio (such as C, N, and P). Bacterial C:N tends to be lower than that of fungi, and adequate N supply favors bacteria over fungi (Mooshammer et al. 2014). Secondly, N addition usually leads to a shift in microbial communities from fungal to bacterial dominance (Strickland and Rousk 2010), which promotes the accumulation of bacterial residual C. In addition, higher N addition can lead to soil acidification and depletion of cationic nutrients, resulting in a reduction in bacterial biomass (Rousk et al. 2009).
4.2 Biotic and Abiotic Factors Influencing Microbial Residues in Cropland and Forest Ecosystems
The generation and accumulation of microbial residues are closely related to the composition of microbial communities and their metabolic activities (Joergensen 2018). Our regression and RDA analyses showed that increased responses of MBC, fungal, and bacterial PLFAs to N addition favored an increase in total, fungal, and bacterial residues C in cropland ecosystems (Figs. 7, 8). However, this effect may not be unidirectional, and our findings indicate that the proliferation of living microorganisms at N addition rates < about 500 kg N ha−1 year−1 promotes the generation of microbial residues, and when N addition rates are 500–800 kg N ha−1 year−1, the increased microbial residues favors microbial growth (Fig. 5). This is because, with N addition, microorganisms can assimilate and use the substrate, continuously synthesize and build cellular components, and finally accumulate microbial residues in the soil during this continuous accumulation process of proliferation-metabolism-death turnover (Kallenbach et al. 2015; Liang et al. 2017). In addition, Jian et al. (2016) showed that adequate N supply appeared to sustain soil microbes to produce more extracellular enzymes associated with hydrolytic C-acquisition, resulting in an overall lower energy acquisition cost. This resulted in microbial residue C being more readily available for microbial decomposition, thus favoring microbial growth; N addition reduced oxidase activity and increased above-ground lignin litter (Jian et al. 2016), increasing the above-ground soil carbon pool and thus microbial activity and promoting the accumulation of microbial residues. It is worth noting that excessive exogenous N input may lead to part of the microbial residue being used as C sources, decomposed and exploited by microbes (Ding et al. 2015).
These environmental factors further influenced microbial residue accumulation mainly by regulating microbial biomass and composition as well as microbial C-use efficiency and turnover rate. (Ding et al. 2010). We found that MAP is the dominant climate factor in the dynamic changes of soil microbial residue accumulation under N addition. Increases in SOC and TN after N addition had a greater effect on microbial residue C (Fig. 8a). This indicates that the C and N pools in the soil directly affect the accumulation and transformation processes of fungal and bacterial residues C (Liang et al. 2019). In addition, the imbalance of soil C and N elements will result in the decomposition and mineralization of microbial residues (Lajeunesse 2011). This is an important mechanism for microbial adaptation to the stoichiometric imbalance between the substrate material and microbial demand (Buckeridge et al. 2020), and this mechanism leads to the reuse of microbial residues in croplands (Algora Gallardo et al. 2021). In addition, the long duration of N addition in cropland ecosystems favored the accumulation of microbial residual C, while the opposite was true in forest ecosystems (Fig. 8a, b). For forest ecosystems, the response of microbial residues to N addition had highly significant and significantly negative correlations with duration and clay, respectively. This suggests that long-term N deposition can negatively affect soil microbial residues in forest ecosystems.
5 Conclusions
In conclusion, our meta-analysis allowed us to analyze conflicting single-site findings as a whole and revealed the overall effect of N addition on microbial residues in croplands and forests. The results demonstrated that N addition in cropland ecosystems significantly increased microbial residue C and biomass and that they were positively correlated with the N addition rate. Further, microbial residue C and biomass under N addition had a bidirectional positive effect. N addition in forest ecosystems only significantly increased bacterial residue C. Notably, there was a significant negative correlation between microbial residue C and duration under N addition, indicating that long-term N deposition caused a significant decrease in microbial residue C in forests. The differential responses to N addition within different ecosystems highlight the necessity to consider microbial residue dynamics in process-based Earth System C models. Collectively, this study will improve our understanding and prediction of the response dynamics of soil microbial residue C to N addition, so as to cope with the changes in nutrient supply in croplands and the negative impacts of N deposition in forests in the future.
Data Availability
All data are provided either in the article or its supplementary materials.
References
Algora Gallardo C, Baldrian P, Lopez-Mondejar R (2021) Litter-inhabiting fungi show high level of specialization towards biopolymers composing plant and fungal biomass. Biol Fertil Soils 57:77–88. https://doi.org/10.1007/s00374-020-01507-3
Appuhn A, Joergensen RG (2006) Microbial colonisation of roots as a function of plant species. Soil Biol Biochem 38:1040–1051. https://doi.org/10.1016/j.soilbio.2005.09.002
Bragazza L, Buttler A, Habermacher J, Brancaleoni L, Gerdol R, Fritze H, Hanajik P, Laiho R, Johnson D (2012) High nitrogen deposition alters the decomposition of bog plant litter and reduces carbon accumulation. Glob Chang Biol 18:1163–1172. https://doi.org/10.1111/j.1365-2486.2011.02585.x
Buckeridge KM, La Rosa AF, Mason KE, Whitaker J, McNamara NP, Grant HK, Ostle NJ (2020) Sticky dead microbes: Rapid abiotic retention of microbial necromass in soil. Soil Biol Biochem 149:107929. https://doi.org/10.1016/j.soilbio.2020.107929
Cotrufo MF, Wallenstein MD, Boot CM, Denef K, Paul E (2013) The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob Chang Biol 19:988–995. https://doi.org/10.1111/gcb.12113
Ding X, Zhang X, He H, Xie H (2010) Dynamics of soil amino sugar pools during decomposition processes of corn residues as affected by inorganic N addition. J Soils Sediments 10:758–766. https://doi.org/10.1007/s11368-009-0132-7
Ding X, Liang C, Zhang B, Yuan Y, Han X (2015) Higher rates of manure application leads to greater accumulation of both fungal and bacterial residues in macroaggregates of a clay soil. Soil Biol Biochem 84:137–146. https://doi.org/10.1016/j.soilbio.2015.02.015
Engelking B, Flessa H, Joergensen RG (2007) Shifts in amino sugar and ergosterol contents after addition of sucrose and cellulose to soil. Soil Biol Biochem 39:2111–2118. https://doi.org/10.1016/j.soilbio.2007.03.020
Hedges LV, Gurevitch J, Curtis PS (1999) The meta-analysis of response ratios in experimental ecology. Ecol 80:1150–1156. https://doi.org/10.2307/177062
Hu J, Huang C, Zhou S, Liu X, Dijkstra FA (2022) Nitrogen addition increases microbial necromass in croplands and bacterial necromass in forests: A global meta-analysis. Soil Biol Biochem 165:108500. https://doi.org/10.1016/j.soilbio.2021.108500
Jian S, Li J, Chen J, Wang G, Mayes MA, Dzantor KE, Hui D, Luo Y (2016) Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol Biochem 101:32–43. https://doi.org/10.1016/j.soilbio.2016.07.003
Jiang J, Wang Y-P, Liu F, Du Y, Zhuang W, Chang Z, Yu M, Yan J (2021) Antagonistic and additive interactions dominate the responses of belowground carbon-cycling processes to nitrogen and phosphorus additions. Soil Biol Biochem 156:108216. https://doi.org/10.1016/j.soilbio.2021.108216
Joergensen RG (2018) Amino sugars as specific indices for fungal and bacterial residues in soil. Biol Fertil Soils 54:559–568. https://doi.org/10.1007/s00374-018-1288-3
Kallenbach CM, Grandy AS, Frey SD, Diefendorf AF (2015) Microbial physiology and necromass regulate agricultural soil carbon accumulation. Soil Biol Biochem 91:279–290. https://doi.org/10.1016/j.soilbio.2015.09.005
Kätterer T, Bolinder MA, Berglund K, Kirchmann H (2012) Strategies for carbon sequestration in agricultural soils in northern Europe. Acta Agric Scand A Anim Sci 62:181–198. https://doi.org/10.1080/09064702.2013.779316
Lajeunesse MJ (2011) On the meta-analysis of response ratios for studies with correlated and multi-group designs. Ecol 92:2049–2055. https://doi.org/10.1890/11-0423.1
Lehmann J, Kleber M (2015) The contentious nature of soil organic matter. Nat 528:60–68. https://doi.org/10.1038/nature16069
Li W, Jin C, Guan D, Wang Q, Wang A, Yuan F, Wu J (2015) The effects of simulated nitrogen deposition on plant root traits: A meta-analysis. Soil Biol Biochem 82:112–118. https://doi.org/10.1016/j.soilbio.2015.01.001
Liang C, Schimel JP, Jastrow JD (2017) The importance of anabolism in microbial control over soil carbon storage. Nat Microbiol 2:17105. https://doi.org/10.1038/nmicrobiol.2017.105
Liang C, Amelung W, Lehmann J, Kaestner M (2019) Quantitative assessment of microbial necromass contribution to soil organic matter. Glob Chang Biol 25:3578–3590. https://doi.org/10.1111/gcb.14781
Liao S, Tan S, Peng Y (2020) Increased microbial sequestration of soil organic carbon under nitrogen deposition over China’s terrestrial ecosystems. Ecol Process 9:52. https://doi.org/10.1186/s13717-020-00260-7
Manzoni S, Taylor P, Richter A, Porporato A, Agren GI (2012) Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol 196:79–91. https://doi.org/10.1111/j.1469-8137.2012.04225.x
Mills MM, Moore CM, Langlois R, Milne A, Achterberg E, Nachtigall K, Lochte K, Geider RJ, La Roche J (2008) Nitrogen and phosphorus co-limitation of bacterial productivity and growth in the oligotrophic subtropical North Atlantic. Limnol Oceanogr 53:824–834. https://doi.org/10.4319/lo.2008.53.2.0824
Mooshammer M, Wanek W, Zechmeister-Boltenstern S, Richter A (2014) Stoichiometric imbalances between terrestrial decomposer communities and their resources: mechanisms and implications of microbial adaptations to their resources. Front Microbiol 5:22. https://doi.org/10.3389/fmicb.2014.00022
Penuelas J, Poulter B, Sardans J, Ciais P, van der Velde M, Bopp L, Boucher O, Godderis Y, Hinsinger P, Llusia J, Nardin E, Vicca S, Obersteiner M, Janssens IA (2013) Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe. Nat Commun 4:2934. https://doi.org/10.1038/ncomms3934
Rousk J, Brookes PC, Bååth E (2009) Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl Environ Microbiol 75(6):1589–1596. https://doi.org/10.1128/AEM.02775-08
Schimel JP, Schaeffer SM (2012) Microbial control over carbon cycling in soil. Front Microbiol 3:348. https://doi.org/10.3389/fmicb.2012.00348
Simpson AJ, Simpson MJ, Smith E, Kelleher BP (2007) Microbially derived inputs to soil organic matter: are current estimates too low? Environ Sci Technol 41:8070–8076. https://doi.org/10.1021/es071217x
Spohn M, Poetsch EM, Eichorst SA, Woebken D, Wanek W, Richter A (2016) Soil microbial carbon use efficiency and biomass turnover in a long-term fertilization experiment in a temperate grassland. Soil Biol Biochem 97:168–175. https://doi.org/10.1016/j.soilbio.2016.03.008
Strickland MS, Rousk J (2010) Considering fungal: bacterial dominance in soils – Methods, controls, and ecosystem implications. Soil Biol Biochem 42:1385–1395. https://doi.org/10.1016/j.soilbio.2010.05.007
Treseder KK (2008) Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies. Ecol Lett 11:1111–1120. https://doi.org/10.1111/j.1461-0248.2008.01230.x
Wang B, An S, Liang C, Liu Y, Kuzyakov Y (2021) Microbial necromass as the source of soil organic carbon in global ecosystems. Soil Biol Biochem 162:108422. https://doi.org/10.1016/j.soilbio.2021.108422
Yu G, Jia Y, He N, Zhu J, Chen Z, Wang Q, Piao S, Liu X, He H, Guo X, Wen Z, Li P, Ding G, Goulding K (2019) Stabilization of atmospheric nitrogen deposition in China over the past decade. Nat Geosci 12:424. https://doi.org/10.1038/s41561-019-0352-4
Zhang X, Amelung W (1996) Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils. Soil Biol Biochem 28:1201–1206. https://doi.org/10.1016/0038-0717(96)00117-4
Zhang W, Cui Y, Lu X, Bai E, He HB, Xie H, Liang C, Zhang XD (2016) High nitrogen deposition decreases the contribution of fungal residues to soil carbon pools in a tropical forest ecosystem. Soil Biol Biochem 97:211–214. https://doi.org/10.1016/j.soilbio.2016.03.019
Zhang X, Jia J, Chen L, Chu H, He JS, Zhang Y, Feng X (2021) Aridity and NPP constrain contribution of microbial necromass to soil organic carbon in the Qinghai-Tibet alpine grasslands. Soil Biol Biochem 156:108213. https://doi.org/10.1016/j.soilbio.2021.108213
Funding
This work was financially supported by the National Natural Science Foundation of China (42077085).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Chen, Q., Ding, X. & Zhang, B. The Effects of N Addition on Soil Microbial Residues in Croplands and Forests: A Meta-analysis. J Soil Sci Plant Nutr 23, 1449–1458 (2023). https://doi.org/10.1007/s42729-023-01189-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-023-01189-4