
Abstract
Despite the saline soils of Bangladesh having high potassium (K) availability, the acquisition of K by rice (Oryza sativa L.) is unsatisfactory. The study was designed to quantify the performance of rice yield components, macro-nutrient uptake, K+/Na+ ratio, and nutrient use efficiency under the influence of improved K fertilization in a salt-affected soil. The salt-tolerant Boro rice cultivar (BRRI-47) was tested with seven different K treatments (0, 20, 40, 60, 80, 100, and 120 kg K ha−1) with three replications followed by a complete randomized design (CRD). The results indicated that increasing K levels had a positive influence on most of the growth and yield parameters of rice except plant height, unfilled grain number, and 1000 grain weight. Application of 100 kg K ha−1 raised the grain yield by 49% over control and 19.13% over the present national recommended dose (40 kg K ha−1). Straw yield increased by 42% in treatment K120 and 36% in treatment K100 over control, respectively. The K+/Na+ ratio in plants was observed to be higher when K was applied at higher rates. Additionally, the different K doses improved the nutrient (N, P, K, S) uptake and decreased the internal N and K use efficiencies of rice. It can be summarized that 100 kg ha−1 K fertilization could be effective to improve rice production and nutrient uptake in coastal saline soil of Bangladesh while maintaining a high K+/Na+ ratio.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Coastal salinity, a limiting factor for crop cultivation in Bangladesh, becomes more apparent with serious concern for food security. At present, 53% of coastal area that constitutes around 1.06 million hectares of the country’s arable lands is affected by different levels of salinity (SRDI 2010). Regions under salinity are expanding over time as a result of rising sea level that increases with the global temperature. In Bangladesh, only over the past 40 years that a total of 0.223 million ha of new land is damaged by salinity (Hasan et al. 2019). Salinity problem adversely affects the plants growth and soil productivity. There are three types of direct effects of salinity on plants: osmotic stress, nutritional imbalances, and element ion toxicity (Isayenkov and Maathuis 2019; Zörb et al. 2019; Billah et al. 2021). Na+, Cl−, SO42−, and HCO3− are some common ions that are found in saline soil in a wide range of concentrations and proportions. The increasing influx of these ions into plant root cells with high osmotic stress prevents the uptake of other essential nutrients especially potassium, K+ (Arif et al. 2020; Makhlouf et al. 2022). Osmotic pressure and ionic toxicity from salinity can also disrupt a variety of physiological processes, including carbohydrate metabolism, respiration, and photosynthesis (Abbasi et al. 2016). Apart from that, in saline soil, the effectiveness of applied fertilizers is decreased by high salt concentration that results in poor plant development (Mohiti et al. 2011).
A number of chemical, biological, and genetic approaches have developed to counteract the harmful effects of salt stress. In the chain of stress-reduction mechanisms, mineral nutrients play a critical role to resist environmental stresses, including salinity problem (Mekdad et al. 2021b). Among the nutrients, potassium (K) is a macro-element with phyto-beneficial properties that organizes physio-biochemical processes to help plants to tolerate abiotic stresses like salinity (Mekdad et al. 2021a; El-Mageed et al. 2022). Selective K+ uptake to maintain a high K+/Na+ ratio in plant throughout the growing season is an effective method for enhancing plants capacity to withstand salt stress (Pi et al. 2016; Tittal et al. 2021). In plant tissue, mainly the highly appealed K transporters (HAKTs) and a channel arbitrated low affinity transporter (LAT) are accounted for K+ uptake (Van Zelm et al. 2020). Nonselective cation channels (NCCs), on the other hand, are most likely the primary transporter responsible for Na+ uptake at root-soil interface (Assaha et al. 2017). Due to similar physicochemical characteristics, excessive Na+ in saline soil inhibits K+ uptake by plants through HAKTs and NCCs, even though there is an adequate amount of K+ present in the soil (Wu et al. 2018; Raddatz et al. 2020). Thus, by disturbing the cytoplasmic K+/Na+ homeostasis, salt stress leads to the increasing Na+ to K+ ratio in the plant cytosol (Javed et al. 2020). Therefore, reducing plant salt stress includes more than just adapting to Na+ toxicity; it also demands consuming enough K+ (Zhang et al. 2010; Pi et al. 2016).
Rice (Oryza sativa L.) is the most extensively consumed starchy cereal grain in Bangladesh, with 144.5 kg of consumption per person annually (Yunus et al. 2019), and rice security is considered the ultimate food security in this country as well as in South Asia. However, because of salinity stress, rice production usually declines by 30 to 50% of total production (Eynard et al. 2005). The increasing saline water intrusion as a consequence of global climate change is thus frustrating the food security in Bangladesh. Available K+ in soil is considered a crucial factor for rice production (Yang et al. 2003). The modern high yielding varieties (HYVs) are reported to remove significant amount of K from soil, sometimes, even more than N to ensure the yield at optimum level (Sharma et al. 2013; Wakeel 2013). Based on a report of Bangladesh Rice Research Institute (BRRI), rice crops remove 19.13 to 22.31 kg K for every ton of rice production (Choudhury et al. 2013). Unexpectedly, like several other Asian nations, Bangladeshi farmers typically overlook K fertilization and focus on N and P application for improving crop production (Islam and Muttaleb 2016). Potassium acts as a counter cation for anion accumulation and helps to mitigate salt stress by decreasing the Na+/K+ ratio in a wide range of crops (Ali et al. 2019; Jan et al. 2020). According to Banerjee et al. (2018), an increase in K+ uptake in the initial developmental stage of rice alleviates the negative impact of Na+ uptake. Further, K plays a significant role in plant physiology, viz., osmoregulation, photosynthesis, enzyme activation, starch, and protein synthesis (Ahanger and Agarwal 2017; Mardanluo et al. 2018). In addition, K application can increase nutrients solubility in saline soil (Niu et al. 2020).
However, in Bangladesh, the research knowledge regarding the rice response under K fertilization in saline soil is still limited and diverse to recommend K for lessening the detrimental impact of salinity. Therefore, we hypothesized that the K application could be an effective method to reduce salinity effect while improving nutrients availability and rice yield. The current research work was thus undertaken (i) to investigate the effect of different K levels on the yield attributes and production of rice, (ii) to study the macro-nutrients uptake trends as well as use efficiencies, and (iii) to find out the K+/Na+ ratio of rice in a salt-affected soil.
2 Material and Methods
2.1 Experiment Site and Soil Description
A pot study was conducted from January to May 2020 at the net house of Soil, Water and Environment Discipline, Khulna University, Bangladesh, where all the prevailing biotic and abiotic environmental factors were present like a real field experiment. Throughout the study, recorded temperature and humidity data for the study site were 15.4–34.3 °C and 74–79%, respectively.
Soil sample was collected from a salinity-affected agriculture field of Batiaghata Upazila, Khulna, before the plowing of the field for the next crop. The soil developed from Gangetic Alluvium parent material and belongs to the Inceptisol order. The associated agro-ecological zone (AEZ) of the experimental soil was the Ganges tidal river floodplain. After collection, the soil was air dried, grinded, and passed through 2-mm sieve. Preliminary soil properties were determined by the standard methods prior to the experiment (see Table 1). Analyses showed that the soil was saline in nature and had a high K content (Table 1).
2.2 Experimental Setup and Cultural Operations
A salinity-tolerant rice variety, BRRI-47 (148–152-day duration) was selected for the experiment. This variety can withstand salinity level up to 12–14 dSm−1 at seedling stage and 8–10 dSm−1 during the vegetative growth stages. The study comprised of complete randomized design (CRD) with seven K treatments (0, 20, 40, 60, 80, 100, and 120 kg ha−1) and three replications. Muriate of potash (KCl) was used as the source of K (60% K2O). Apart from the K treatments, recommended doses of N, P, S, and Zn were also applied into each of the experimental pots at a rate of 125, 24, 6, and 1.5 kg ha−1, respectively, as suggested by Bangladesh Agricultural Research Council (BARC 2018). Fertilizers were applied on a g pot−1 basis from kg ha−1 after calculation by using Eq. 1. The sources of N, P, S, and Zn were urea, triple super phosphate (TSP), gypsum, and zinc sulfate (ZnS), respectively. Muriate of potash (treatment) was applied into 2 splits: 50% before transplantation, during the final soil preparation, and 50% at 25 days after transplantation. The others recommended fertilizers (TSP, gypsum, and ZnS) were applied as basal dose in all the pots at the time of soil preparation. Urea was applied into three splits: 7, 30, and 50 days after transplantation, respectively. Following that, each pot was filled with 5 kg of soil, and three 23-day-old seedlings were transplanted into each pot after puddling under irrigated condition. For cultural operations, proper crop management measures including irrigation and weeding were followed throughout the growing period and kept uniform across all experimental pots. Harvesting was done when the grain reached at 80% of its maturity stage, and afterwards, the cut crops were threshed, winnowed, cleaned, and packed separately for laboratory analyses.
Here, TD or RD represents treatment dose or recommended dose.
2.3 Growth Parameters and Yield Measurement
The plant height (PH), number of tillers hill−1 (TN), and the number of panicle hill−1 (PN) of rice was measured in this experiment. The PH was measured from the base of the plant to the tip of the tallest panicle, and the TN and PN were enumerated manually. After threshing, filled grains panicle−1 (FG) and unfilled grains panicle−1 (UFG) were differentiated by dipping the seeds into water. The FG was settled down, and the light floating UFG was removed from the water. Fresh grain yield (GY) and straw yield (SY) were recorded in g pot−1 basis, and the yield was then converted into t ha−1 unit (Eq. 3). Dry matter yield (DMY) was estimated after the green biomass was oven-dried at 65 °C temperature. Agronomic efficiency of K (AEK) is the grain yield response of rice due to per unit of applied K, and partial factor productivity (PFPK) is the estimation of K fertilizer productivity. Following that, grain harvest index (GHI), agronomic K use efficiency (AEK), and partial factor productivity (PFPK) were calculated as Eq. 4, Eq. 5, and Eq. 6, respectively.
2.4 Internal Nutrient Use Efficiency
Internal nutrient use efficiency (INUE) is a concept of taking up and utilizing nutrients for maximum grain yield. The nutrient uptake and internal nutrient use efficiencies (INUE) were calculated followed by Eq. 7 and Eq. 8 (Yang et al. 2004).
Here, DMY represents Dry Matter Yield in kg ha−1, which was estimated from Eq. 2
2.5 Statistical Analysis
One-way analysis of variance (ANOVA) was performed by using Minitab (version 19.0, Minitab LCC, Triola Statistics Company, 1972) at 95% significant level. The linear regression graph was illustrated for demonstrating the correlations between nutrient uptake parameters and treatments by using Origin 2021 (Origin Lab, Northampton, MA, USA). Besides, principal component analysis (PCA) was done to depict the correlations of a group of agronomic attributes and nutrient uptake parameters using R 3.6.1 (R Core Team 2013).
3 Results
3.1 Growth Parameters of BRRI-47
In this experiment, different K levels did not show any significant influence (p ≤ 0.05) on the plant height (PH). However, the result revealed a significant effect on tiller number (TN) and panicle number (PN) under the application of K (Table 2). Observation showed that treatment K100 had the maximum TN (16), whereas the minimum TN (9) was recorded from treatment K0. Treatment K100 resulted in maximum PN (16) and was statistically identical to treatment K40, K60, K80, and K120, while the minimum PN (7) was attained from treatment K0. A significant correlation was observed for TN (r = 0.87, p ≤ 0.001) and PN (r = 0.81, p ≤ 0.001) with different K rates (Fig. 1).

Pearson correlation matrix among rice growth and yield parameter and nutrient uptake parameters. The blue, red, and white color correspond positive, negative, and neutral correlation, respectively. *, **, and *** represent significance levels at p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively. Here: Treatment = 0, 20, 40, 60, 80, 100, and 120 kg K ha−1; PH, plant height; TN, number of tillers hill−1; PN, number of panicle hill−1; FG, filled grain panicle−1; UFG, unfilled grain panicle−1; 1000-G.wt, 1000-grain weight; GY, grain yield; SY, straw yield; DMY, dry matter yield; GHI, grain harvest index; AE, agronomic use efficiency of K; PFP, partial factor productivity of K
The filled grain panicle−1 (FG) of BRRI-47 was positively (p ≤ 0.05) affected by different K treatments (Table 2). The FG was maximum at treatment K100, accounting for 29% more FG than control. The magnitude of different treatments for FG was recorded as K100 > K120 > K80 > K40 > K60 > K20 > K0. A significant correlation (r = 0.84, p ≤ 0.001) was obtained between the FG and increasing K doses (Fig. 1). However, no significant effect of K was obtained on unfilled grain panicle−1 (UFG) and 1000-grain weight (100-G.wt) production of rice (Table 2).
3.2 Yield Parameter of BRRI-47
The grain yield (GY) and straw yield (SY) were significantly influenced by different K levels (p ≤ 0.05) (Table 3). Statistically no significant variation was found in GY and the SY from treatment K80 to K120. Treatment K0 accounted for lower GY (3.51 t ha−1) compared to treatments getting higher K dose. GY in K100 was increased by 49% over K0 (control) and 19.13% over treatment K40 (current recommended rate). Under different K doses, K120 produced the maximum SY (8.08 t ha−1) followed by K100. On the contrary, minimum SY (5.67 t ha−1) was obtained from control treatment. Increasing K application had a significant correlation (r = 0.89, p ≤ 0.001) with the GY (Fig. 1). Variable K treatments appeared to have a significant influence on dry matter yield (DMY) production (Table 3). Treatment K100 and K120 showed the highest DMY response, and the lowest response was obtained from treatment K0. Treatment K120 produced 46% more DMY than K0. Statistically, an identical DMY was obtained from treatment K60, K80, K100, and K120. However, application of K did not show a significance effect on the grain harvest index (GHI %) of rice (Table 3).
3.3 Potassium Use Efficiency
In saline soil, added K found to have a significant influence on the agronomic efficiency of K (AEK) and partial factor productivity of K (PFPK). Data perusal showed a declining trend of AEK and PFPK indices with the increment of K doses (Table 3). Application of K120 minimized the AEK (12.01) and PFPK (41.26). On the contrary, K20 resulted in maximum AEK (29.21) and PFPK (204), respectively.
3.4 Macro-nutrients Uptake of Rice
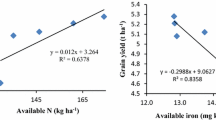
The nitrogen (N), potassium (K), phosphorus (P), and sulfur (S) uptake of rice were significantly influenced by increasing rates of K (Table 4). Nitrogen uptake did not show any significant difference (p ≤ 0.05) from treatment K40 to K100. The study showed that in saline soil, K fertilization increased the N uptake by 164% in treatment K100 and 188% in treatment K120 over control treatment. Overall, the highest response in terms of N uptake (112.9 kg ha−1) was obtained from treatment K120, while the lowest response of K on N uptake (39.18 kg ha−1) was recorded in treatment K0. Lowest P uptake (12.16 kg ha−1) by rice was noticed in treatment K0 which gradually increased the uptake to 27.75 kg ha−1 in treatment K100. Effects of added K on N and P uptake can be expressed by Eq. 9 and Eq. 10, respectively. The N uptake and P uptake by rice with K treatments were linear and accounted for 74% (Fig. 2a) and 60% (Fig. 2b) of the variability in this experiment.
where y = quantity of nutrient uptake (kg ha−1), x = amount of applied K (kg ha−1), and *** = p ≤ 0.001.

General linear regression between K application and nitrogen (a), phosphorus (b), potassium (c), and sulfur (d) uptake by BRRI -47
Result revealed that K uptake by rice was significantly (p ≤ 0.05) affected by the application of different K doses in saline soil (Table 4). In this experiment, a progressive increment of K uptake was noticed with the increasing levels of K fertilization. The mean K uptake for all treatments ranged from 54.48 to 136.8 kg ha−1. Treatment K80, K100, and K120 resulted in better performance in terms of K uptake compared to treatments receiving lower K dose. Treatment K120 resulted 150% more K uptake than K0. Total S uptake by BRRI-47 registered a significant (p ≤ 0.05) effect with the improved K doses. Minimum S uptake was obtained by treatment K0 (6.54 kg ha−1), whereas treatment K120 showed the maximum uptake (13.83 kg ha−1) and was significantly higher than treatment K60, K40, K20, and K0. Influence of applied potassium on K and S uptake can be explained by Eq. 11 and Eq. 12, which accounted for 92% (Fig. 2c) and 89% (Fig. 2d) of the variability, respectively.
where y = quantity of nutrient uptake (kg ha−1), x = amount of applied K (kg ha−1), and *** = p ≤ 0.001.
3.5 K + /Na + Ratio of Plant
The K+/Na+ ratio of studied rice responded significantly (p ≤ 0.05) with the application of higher K doses (Fig. 3). The K+/Na+ ratio progressively increased with the increasing K levels. Among the treatments, K120 demonstrated the highest ratio (0.95) followed by K100 (0.94) which were 206% and 203% more than the K control treatment, respectively. A strong significant correlation (r = 0.95, p ≤ 0.001) was noticed between K+/Na+ ratio and the application of K (Fig. 1).

K+/Na+ ratio of rice with K treatments. Data included as mean ± SE (standard error). Small common letters denote no significant difference in mean values at 0.05% level of significance. Here, Treatment K0 = control, K20 = 20 kg K ha−1, K40 = 40 kg K ha−1, K60 = 60 kg K ha−1, K80 = 80 kg K ha−1, K100 = 100 kg K ha−1, K120 = 120 kg K ha.−1
3.6 Internal Nutrient Use Efficiency (INUE)
In this study, different K doses implied a significant effect on the internal N use efficiency (IUEN) and internal K use efficiency (IUEK). The influence, on the other hand, was indifferent for internal P and S use efficiencies (Table 5). Highest IUEN (89.86) was obtained by 0 kg K ha−1 that significantly decreased to 44.65 with the application of 120 kg K ha−1. Similarly, mean IUEK was at maximum (64.45) in treatment K0 and decreased to 38.56 in treatment K120.
3.7 Principal Component Analysis and Correlation
Principal component analysis (variables (Fig. 4a) and bi-plot PCA (Fig. 4b)) of growth, yield, and nutrient uptake were done as influenced by K. Based on the findings, first two principal components (PC) accounted for 74.77% of dataset variance, where PC1 accounted for 61.89% of the total variance (Fig. 4a, b). PCA bi-plot evidently represented observation distribution corresponding to every treatment and the interaction between two main features. Throughout the dataset, N uptake and GY showed the maximum contribution, while PH and GHI showed the least contribution.

(a) Variables and (b) bi-plot PCA diagrams demonstrated the performance of K on various growth, yield, and nutrient uptake features of BRRI-47. Here, smaller angles within the vectors depict the greater correlation. In variables PCA red colors are responsible for larger contribution, whereas blue one accounted for less contribution. Parameters included: PH, plant height; TN, number of tiller panicle−1; FG, filled grain panicle−1; 1000-G.wt, 1000-grain weight; GY, grain yield; SY, straw yield; GHI, grain harvest index; AE, agronomic use efficiency of K; PFP, partial factor productivity of K; N uptake, nitrogen uptake
4 Discussion
In this study, an insignificant effect of K on the PH was noticed that is in consistent with the findings of Ardakani et al. (2016). Under saline condition, interim stress of K fertilization might be accounted for the reduced PH (Adhikari et al. 2020). However, a significant effect on rice PH upon K application was obtained by Hartati and Purnomo (2018) that contradicted with our results. The difference in crop salt tolerance mechanisms, soil physiography, or the cropping patterns perhaps was the probable reasons for this contrast finding. The experiment showed a positive effect of K on the TN and PN of rice, which is supported by some previous perusals indicating higher levels of K minimized plant salt stress and increased the effective TN and PN production of rice (Zain and Ismail 2016; Bhagat et al. 2019). Increasing TN and PN per unit plant was one of the factors for rising rice yield (Banerjee et al. 2018). The study implies that rice productivity is collectively regulated by its vegetative growth as the GY noticed to be significantly correlated (r = 0.74, p ≤ 0.001; r = 0.69, p ≤ 0.001) with the TN and PN production, respectively. Adequate K supplies improved N uptake during the growth stages of rice, and this cumulative N uptake increased TN and PN production, which may be due to more N availability that facilitated stomatal movement and cell division of rice plant (Souri and Hatamian 2019; Kundu et al. 2020).
In saline environment, assimilation of CO2, stomatal opening, and seed production are largely affected by the presence of excess salt ions in nutritional medium (Lima et al. 2020). The FG production of this present study was significantly affected by different K treatment that is identical to the findings reported by Islam et al. (2016). Due to K-mediated carbohydrate metabolism, higher rates of K fertilizer yield huge quantities of starch that facilitated the increasing number of FG production and heavier rice grain (Jiang et al. 2019; Khokhar et al. 2022). In our experiment, the effect of K was indifferent on the UFG and 1000-G. wt. of BRRI-47. Islam and Muttaleb (2016) also found an insignificant effect of K on rice UFG and 1000-G. wt. that is in coherence with our observation. However, a significant response of K on 1000-G. wt. was reported by a number earlier investigations (Hartati and Purnomo 2018; Li et al. 2018). We assumed that these contradict results probably caused by the variations in rice varieties, soil, climate, and cultivation practices, as majority of these studies are conducted in agricultural fields that reflected the farmer’s different management conditions. Despite the fact that early rice growth phases are extremely sensitive to salinity impact, as it significantly affected both green and dry rice yield during this stage (Acosta-Motos et al. 2017), we obtained an increase in rice GY and SY with higher K doses. This could be because rice vegetative growth and yield have a clear relationship with photosynthesis rate that improved under the supply of elevated K levels (Zain and Ismail 2016; Kundu et al. 2020). Additionally, potassium is known to promote photosynthetic pigments and enzyme activity that has a positive effect on K-mediated carbohydrate metabolism and biological activity of plants (Islam et al. 2016). We recorded that application of 100 kg K ha−1 increased the grain yield by 19.13% over the current recommended dose (40 kg K ha−1). This demonstrates the better response of rice in saline soil under increased K supply compared to the traditional practice. Similar to our findings, a significant variation in GY and SY of rice under different K treatments was also documented in some earlier studies (Pal et al. 2019; Mehmood et al. 2020).
DMY of BRRI-47 was significantly higher upon rising K doses. Islam et al. (2015) stated that use of inorganic K fertilizer improved rice plant growth via increasing photosynthetic capability and translocation of photo-assimilate availability of N, P, and K which consequently increased the DMY yield of rice. The result is identical to that of Banerjee et al. (2018) who concluded that K application resulted in greater DMY by increasing net photosynthesis and CO2 assimilation at heading and ripening phases. AEK and PFPK are two indices that represent the efficiencies of added K. The variation in AEK and PFPK indices were closely connected with the amount of used fertilizer and the values trend to decrease with the increase in K rates. According to Sarangi et al. (2016), the decrease in AEK and PFPK values might be associated the arced return to the transformation of K to grain as the yield approaches the crop canopy at elevated K levels. The present finding is in agreement with some other studies reporting K use efficiency was positively affected by K fertilization (Li et al. 2018; He et al. 2022).
Rice response to external K application largely depends on the amount of time, the sources and mode of fertilizer application, and the relationship of K with other nutrients (Vijayakumar et al. 2019). Potassium supply at higher levels could increase N, K, and P uptake by plant (Dhillon et al. 2019; Kumar et al. 2022). In this study, an increase in macro-nutrient uptake was observed with the rising K doses. The finding is supported by several past investigations mentioning that applied K increased the nutrient acquisition in saline soil by interacting with Na (Kibria et al. 2015; Merwad 2016). In saline soil, K supplementation favors higher stomatal conductance in plants that influenced C absorption and photosynthetic product distribution to the root system (Hou et al. 2019). Therefore, it stimulates the root system growth and development and enhanced the essential nutrient uptake in saline soil. An increase in N, K, P, and S acquisition was observed in this study demonstrating that salinity issue improved with the application of K. In this experiment, 120 kg ha−1 K had showed the maximum K uptake. However, the grain yield at this K level was statistically identical to treatment K100 and K80. In this case, the higher K uptake without any significant improvement of yield could lead to luxury consumption of K (Ravichandran and Sriramachandrasekharan 2011).
Salinity tolerance is an intricate mechanism, involving the ability of plant to resist ionic and osmotic stresses produced by highly salinized soil (Chiconato et al. 2019). Consequently, the mitigation measures for salt stress should not emphasize only on reducing the Na+ buildup in vigorous photosynthetic tissue (Shabala and Cuin 2008) but instead on K+ homeostasis and the maintenance of a high K+/Na+ ratio (Barus et al. 2018). K+/Na+ ratio is considered a key factor for assessing crop performance in saline soil (Mohapatra et al. 2021). Under saline condition, the controlled acquisition and translocation of K+ act as an antagonistic parameter to prevent the buildup of harmful ions like Na+ and chloride (Cl−) ions (Kumari et al. 2021). This occurs mostly because of decreased K+ efflux, which causes significant intracellular retention of K+ following the exclusion of Na+ (Gul et al. 2019). Thus, the application of increasing K levels improved the K+/Na+ ratio in rice plant, which aids rice’s ability to withstand salt stress. Munir et al. (2019) reported that under saline-sodic environment, application of K at higher levels could ameliorate plant physiological and biochemical traits by increasing the K+/Na+ ratio in plant tissues that is in line with our present study. An identical effect of K on the K+/Na+ ratio of plant was also recorded by Li et al. (2022).
INUE is an important factor for all nutrients to outline a point of recommendation for future development. In our study, “dilution effect” apparently accounted to maximize the IUEN and IUEK in K control pot rather that in K-treated pots. Similar to our findings, Islam and Muttaleb (2016) reported that plants trend to exert the nutrients more efficiently at biochemical level in control pot compared to the pots where the element quantity was relatively high. In this experiment, highly significant correlations between most of the agronomic components and nutrient uptake parameters of rice reflected the greater influence of K fertilization. As a whole, the increasing doses of K fertilizer contributed in lessening salinity stress by ensuring a higher K+ to Na+ ratio and significantly enhanced most of the growth, yield, and nutrient uptake traits of rice.
5 Conclusions
The coastal tract of Bangladesh has a very high agricultural potential. But due to salinity issue, the crop productivity in this region cannot able to meet the national average production. The findings of our present investigation imply that the current recommended K rate for rice cultivation (40 kg K ha−1) is insufficient to assist obtainable rice yield potential, and the level might need an improvement to 100 kg K ha−1 to maximize the rice productivity and effective nutrient management. Therefore, this project will help to alleviate salinity effect and enhance rice productivity under saline condition. However, optimum K supply for rice generally varies with the variety, yield target, and fertility status of soil. Conclusively, the study is limited to a single rice variety that does not accurately represent the overall situation. Further research is still needed to establish 100 kg ha−1 K as a recommended dose for improving rice production and for managing salinity impact in the coastal belts of Bangladesh.
Data Availability
The field and lab (raw) data used to support the findings of this study are available from the corresponding author upon request.
References
Abbasi H, Jamil M, Haq A, Ali S, Ahmad R, Malik Z, Parveen (2016) Salt stress manifestation on plants, Mechanism of salt tolerance and potassium role in alleviating it: a review. Zemdirbyste-Agric 103:229–238. https://doi.org/10.13080/z-a.2016.103.030
Acosta-Motos JR, Ortuño MF, Bernal-Vicente A, Diaz-Vivancos P, Sanchez-Blanco MJ, Hernandez JA (2017) Plant responses to salt stress: adaptive mechanisms. Agronomy 7:18. https://doi.org/10.3390/agronomy7010018
Adhikari B, Dhungana SK, Kim ID, Shin DH (2020) Effect of foliar application of potassium fertilizers on soybean plants under salinity stress. J Saudi Soc Agric Sci 19:261–269. https://doi.org/10.1016/j.jssas.2019.02.001
Ahanger MA, Agarwal RM (2017) Potassium improves antioxidant metabolism and alleviates growth inhibition under water and osmotic stress in wheat (Triticum aestivum L.). Protoplasma 254:1471–1486. https://doi.org/10.1007/s00709-016-1037-0
Ali A, Maggio A, Bressan RA, Yun D-J (2019) Role and functional differences of HKT1-type transporters in plants under salt stress. Int J Mol Sci 20:1059. https://doi.org/10.3390/2Fijms20051059
Allison LE, Richards LA (1954) Diagnosis and improvement of saline and alkali soils. U.S. Department of Agriculture
Ardakani A, Armin M, Filehkesh E (2016) The effect of rate and application method of potassium on yield and yield components of cotton in saline condition. Iran J Field Crops Res 14(3):514–525
Arif Y, Singh P, Siddiqui H, Bajguz A, Hayat S (2020) Salinity induced physiological and biochemical changes in plants: an omic approach towards salt stress tolerance. Plant Physiol Biochem 156:64–77. https://doi.org/10.1016/j.plaphy.2020.08.042
Assaha DV, Ueda A, Saneoka H, Al-Yahyai R, Yaish MW (2017) The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front Physiol 8:509. https://doi.org/10.3389/fphys.2017.00509
Banerjee H, Ray K, Dutta SK, Majumdar K, Satyanarayana T, Timsina J (2018) Optimizing potassium application for hybrid rice (Oryza sativa L.) in coastal saline soils of West Bengal. Indian J Agron 8:292. https://doi.org/10.3390/agronomy8120292
BARC (2018) Fertilizer recommendation guide. Bangladesh Agricultural Research Council, Farmgate, Dhaka
Barus WA, Abdul R, Rosmayati AR, Chairani H (2018) Study of nutrient uptake in some varieties of rice by foliar application of potassium phosphate fertilizer on saline soil. Int J Sci Technol Res 7(1)
Bhagat MA, Mahar AR, Rajpar I, Channa, et al (2019) Potassium applications on rice (Oryza sativa L.) genotypes under saline environment. Sindh Univ Res J 51:333–338. https://doi.org/10.26692/sujo/2019.6.54
Billah M, Aktar S, Brestic M, Zivcak M, Khaldun ABM, Uddin et al (2021) Progressive genomic approaches to explore drought-and salt-induced oxidative stress responses in plants under changing climate. Plants 10:1910. https://doi.org/10.3390/plants10091910
Chapman HD (1965) Cation-exchange capacity. In: Methods of Soil Analysis. John Wiley & Sons, Ltd, pp 891–901
Chiconato DA, Cruz FJR, da Silva SJG, dos Santos DMM, Munns R (2019) Adaptation of sugarcane (Saccharum officinarum L.) plants to saline soil. Environ Exp Bot 162:201–211. https://doi.org/10.1016/j.envexpbot.2019.02.021
Choudhury ATMA, Saleque MA, Zaman SK, Bhuiyan NI, Shah AL, Rahman MS (2013) Nitrogen fertilizer management strategies for rice production in Bangladesh: nitrogen fertilizer for rice production. Biol Sci-PJSIR 56:167–174
Dewis J, Freitas F (1970) Physical and chemical methods of soil and water analysis. FAO Soils Bull 10:275
Dhillon JS, Eickhoff EM, Mullen RW, Raun WR (2019) World potassium use efficiency in cereal crops. Agron J 111:889–896. https://doi.org/10.2134/agronj2018.07.0462
El-Mageed TA, Mekdad AA, Rady MO, Abdelbaky AS, Saudy HS, Shaaban A (2022) Physio-biochemical and agronomic changes of two sugar beet cultivars grown in saline soil as influenced by potassium fertilizer. J Soil Sci Plant Nutr 22:3636–3654. https://doi.org/10.1007/s42729-022-00916-7
Eynard A, Lal R, Wiebe K (2005) Crop response in salt-affected soils. J Sustain Agric 27:5–50
Gul M, Wakeel A, Steffens D, Lindberg S (2019) Potassium-induced decrease in cytosolic Na+ alleviates deleterious effects of salt stress on wheat (Triticum aestivum L.). Plant Biol 21:825–831. https://doi.org/10.1111/plb.12999
Hartati SS, Purnomo D (2018) Effectiveness and efficiency of potassium fertilizer application to increase the production and quality of rice in entisols. IOP Conf Ser Earth Environ Sci 142:012031. https://doi.org/10.1088/1755-1315/142/1/012031
Hasan MH, Rahman MR, Haque A, Hossain T (2019) Soil salinity hazard assessment in Bangladesh coastal zone. In: International Conference on Disaster Risk Management.https://www.researchgate.net/publication/330383801_SOIL_SALINITY_HAZARD_ASSESSMENT_IN_BANGLADESH_COASTAL_ZONE. Accessed 7 November 2022
He B, Xue C, Sun Z, Ji Q, Wei J, Ma W (2022) Effect of different long-term potassium dosages on crop yield and potassium use efficiency in the maize–wheat rotation system. Agronomy 12:2565. https://doi.org/10.3390/agronomy12102565
Hou W, Xue X, Li X, Khan MR, Yan J, Ren T, Lu J (2019) Interactive effects of nitrogen and potassium on: grain yield, nitrogen uptake and nitrogen use efficiency of rice in low potassium fertility soil in China. Field Crops Res 236:14–23
Hunt J (1980) Determination of total sulphur in small amounts of plant material. Analyst 105:83–85
Isayenkov SV, Maathuis FJ (2019) Plant salinity stress: many unanswered questions remain. Front Plant Sci 10:80. https://doi.org/10.3389/fpls.2019.00080
Islam A, Muttaleb A (2016) Effect of potassium fertilization on yield and potassium nutrition of Boro rice in a wetland ecosystem of Bangladesh. Arch Agron Soil Sci 62:1530–1540. https://doi.org/10.1080/03650340.2016.1157259
Islam A, Chandrabiswas J, Sirajul Karim AJM, Salmapervin MST, Saleque MD (2015) Effects of potassium fertilization on growth and yield of wetland rice in grey terrace soils of Bangladesh. Res Crop Ecophysiol 10:64–82
Islam A, Saha PK, Biswas JC, Saleque MA (2016) Potassium fertilization in intensive wetland rice system: yield, potassium use efficiency and soil potassium status. Int J Agric Pap 1:7–21
Jackson ML (1973) Soil chemical analysis. Pentice hall of India Pvt. Ltd. New Delhi India 498:151–154
Jan M, Anwar ulHaq M, ulHaq T, Ali A, Hussain S, Ibrahim M (2020) Protective effect of potassium application on NaCl induced stress in tomato (Lycopersicon esculentum L.) genotypes. J Plant Nutr 43:1988–98. https://doi.org/10.1080/01904167.2020.1766071
Javed H, Aneela Rİ, Qureshi A, Javed K, Mujeeb F, Fraza IJ, Akhtar MS, Ali MA, Rehman GU, Aftab M (2020) Isolation, characterization and screening of PGPR capable of providing relief in salinity stress. Eurasian J Soil Sci 9:85–91. https://doi.org/10.18393/ejss.650546
Jiang W, Wang X, Xing Y, Liu X, Cui Z, Yang L (2019) Enhancing rice production by potassium management: recommended reasonable fertilization strategies in different inherent soil productivity levels for a sustainable rice production system. Sustainability 11:6522. https://doi.org/10.3390/su11226522
Khokhar A, Sharma V, Singh MJ, Yousuf A, Sandhu PS, Kumar V, Bhat MA (2022) Effect of potassium and magnesium application on growth, yield and nutrient uptake of rainfed maize in the sub-montaneous region of Punjab, India. J Plant Nutr 7:1–1. https://doi.org/10.1080/01904167.2022.2063732
Kibria MG, Farhad HM, Hoque MA (2015) Alleviation of soil salinity in rice by potassium and zinc fertilization. J Exp Agric Int 5:15–21
Kumar D, Ramesh K, Jinger D, Rajpoot SK (2022) Effect of potassium fertilization on water productivity, irrigation water use efficiency, and grain quality under direct seeded rice-wheat cropping system. J Plant Nut 45:2023–2038. https://doi.org/10.1080/01904167.2022.2046071
Kumari S, Chhillar H, Chopra P, Khanna RR, Khan MI (2021) Potassium: a track to develop salinity tolerant plants. Plant Physiol Biochem 167:1011–1023. https://doi.org/10.1016/j.plaphy.2021.09.031
Kundsen D, Peterson GA, Part PF (1982) Lithium, sodium and potassium. In: Methods of Soil Analysis, 2nd edn. LincMadision, Wisdon, USA
Kundu A, Raha P, Dubey AN, Rani M, Paul A, Patel R (2020) Differential responses of rice (Oryza sativa L.) to foliar fertilization of organic potassium salts. J Plant Nutr 44:1330–1348. https://doi.org/10.1080/01904167.2020.1862193
Li ST, Yu DUAN, Guo TW, Zhang PL, Ping HE, Majumdar K (2018) Sunflower response to potassium fertilization and nutrient requirement estimation. J Integr Agric 17:2802–2812. https://doi.org/10.1016/S2095-3119(18)62074-X
Li SJ, Wu GQ, Lin LY (2022) AKT1, HAK5, SKOR, HKT1;5, SOS1 and NHX1 synergistically control Na+ and K+ homeostasis in sugar beet (Beta vulgaris L.) seedlings under saline conditions. J Plant Biochem Biotech 31:71–84. https://doi.org/10.1007/s13562-021-00656-2
Lima GSD, Fernandes CGJ, Soares LADA, Gheyi HR, Fernandes PD (2020) Gas exchange, chloroplast pigments and growth of passion fruit cultivated with saline water and potassium fertilization. Rev Caatinga 33:184–194. https://doi.org/10.1590/1983-21252020v33n120rc
Makhlouf BSI, Khalil Soha RA, Saudy HS (2022) Efficacy of humic acids and chitosan for enhancing yield and sugar quality of sugar beet under moderate and severe drought. J Soil Sci Plant Nutr 24:1–6. https://doi.org/10.1007/s42729-022-00762-7
Mardanluo S, Souri MK, Ahmadi M (2018) Plant growth and fruit quality of two pepper cultivars under different potassium levels of nutrient solutions. J Plant Nutr Soil Sci 41:1604–1614. https://doi.org/10.1080/01904167.2018.1463383
Mehmood H, Abbasi GH, Jamil M, Malik Z, Ali M, Iqbal R, Ahmed S, Akhtar MW, Irfan M, Afzal S (2020) Potential of applied potassium in improving salt tolerance of Maize (Zea mays L.) hybrids. J Biol Environ Sci 16:35–43
Mekdad AA, Rady MM, Ali EF, Hassan FA (2021) Early sowing combined with adequate potassium and sulfur fertilization: promoting Beta vulgaris (L.) yield, yield quality, and K-and S-use efficiency in a dry saline environment. Agronomy 11:806. https://doi.org/10.3390/agronomy11040806
Mekdad AA, Shaaban A, Rady MM, Ali EF, Hassan FA (2021b) Integrated application of K and Zn as an avenue to promote sugar beet yield, industrial sugar quality, and K-use efficiency in a salty semi-arid agro-ecosystem. Agronomy 11:780. https://doi.org/10.3390/agronomy11040780
Merwad ARM (2016) Efficiency of potassium fertilization and salicylic acid on yield and nutrient accumulation of sugar beet grown on saline soil. Commun Soil Sci Plant Anal 47:1184–1192. https://doi.org/10.1080/00103624.2016.1166242
Mohapatra A, Jena D, Pradhan PP, Pattnaik D, Pal A, Pattanik M (2021) Interactive effects of potassium and saline water on yield, nutrient accumulation and uptake by rice. Int J Chem Stud 9:2936–43. https://doi.org/10.22271/chemi.2021.v9.i1ao.11676
Mohiti M, Ardalan MM, Torkashvand AM, Vahed HS (2011) The efficiency of potassium fertilization methods on the growth of rice (Oryza sativa L.) under salinity stress. Afr J Biotechnol 10:15946–15952. https://doi.org/10.5897/AJB11.1351
Munir A, Shehzad MT, Qadir AA, Murtaza G, Khalid HI (2019) Use of potassium fertilization to ameliorate the adverse effects of saline-sodic stress condition (ECw: SARw levels) in rice (Oryza sativa L.). Commun Soil Sci Plant Anal 50:1975–1985. https://doi.org/10.1080/00103624.2019.1648657
Niu J, Liu C, Huang M, Liu K, Yan D (2020) Effects of foliar fertilization: a review of current status and future perspectives. J Soil Sci Plant Nutr 21:104–118. https://doi.org/10.1007/s42729-020-00346-3
Olsen SR (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. U. S. Department of Agriculture, Washington D. C.
Pal SC, Chakrobortty J, Hashem MA, Hoque M (2019) Improvement of salinity tolerance in rice by efficient management of potassium and zinc fertilizers. Asian Australas J Biosci Biotechnol 4:154–161
Pi Z, Stevanato P, Sun F, Yang Y, Sun X, Zhao H, Geng G, Yu L (2016) Proteomic changes induced by potassium deficiency and potassium substitution by sodium in sugar beet. J Plant Res 129:527–538. https://doi.org/10.1007/s10265-016-0800-9
Raddatz N, Morales de los Ríos L, Lindahl M, Quintero FJ, Pardo JM (2020) Coordinated transport of nitrate, potassium, and sodium. Front Plant Sci 11:247. https://doi.org/10.3389/fpls.2020.00247
Ravichandran M, Sriramachandrasekharan MV (2011) Optimizing timing of potassium application in productivity enhancement of crops. Karnataka J Agri Sci 24:75–80
Sarangi SK, Maji B, Singh S, Sharma DK, Burman D, Mandal S, Haefele SM (2016) Using improved variety and management enhances rice productivity in stagnant flood-affected tropical coastal zones. Field Crop Res 190:70–81
Shabala S, Cuin TA (2008) Potassium transport and plant salt tolerance. Physiol Plant 133:651–669. https://doi.org/10.1111/j.1399-3054.2007.01008.x
Sharma S, Chander G, Verma TS, Verma S (2013) Soil potassium fractions in rice-wheat cropping system after twelve years of Lantana residue incorporation in a Northwest Himalayan acid alfisol. J Plant Nutr 36:1809–1820
Souri MK, Hatamian M (2019) Aminochelates in plant nutrition: a review. J Plant Nutr 42:67–78. https://doi.org/10.1080/01904167.2018.1549671
SRDI (2010) Saline soils of Bangladesh. Soil Resource Development Institute, Dhaka Bangladesh
R Core Team (2018). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Tittal M, Mir RA, Jatav KS, Agarwal RM (2021) Supplementation of potassium alleviates water stress-induced changes in Sorghum bicolor L. Physiol Plant 172:1149–1161. https://doi.org/10.1111/ppl.13306
Van Zelm E, Zhang Y, Testerink C (2020) Salt tolerance mechanisms of plants. Annu Rev Plant Biol 71:403–433. https://doi.org/10.1146/annurev-arplant-050718-100005
Vijayakumar S, Kumar D, Srivay YS, Sharma VK, Sharma DK, Saravanane P, Poornima S, Singh N (2019) Energy budgeting of aerobic rice (Oryza sativa L)-wheat (Triticum aestivum) cropping system as influenced by potassium fertilization. Indian J Agric Sci 89:159–163
Wakeel A (2013) Potassium–sodium interactions in soil and plant under saline-sodic conditions. J Plant Nutr Soil Sci 176:344–354. https://doi.org/10.1002/jpln.201200417
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38
Wu H, Zhang X, Giraldo JP, Shabala S (2018) It is not all about sodium: revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 431:1–17
Yang XE, Liu JX, Wang WM, Li H, Luo AC, Ye ZQ, Yang Y (2003) Genotypic differences and some associated plant traits in potassium internal use efficiency of lowland rice (Oryza sativa L.). Nutr Cycl Agroecosystems 67:273–282
Yang XE, Liu JX, Wang WM, Ye ZQ, Luo AC (2004) Potassium internal use efficiency relative to growth vigor, potassium distribution, and carbohydrate allocation in rice genotypes. J Plant Nutr 27:837–852
Yunus M, Rashid S, Chowdhury S (2019) Per capita rice consumption in Bangladesh: available estimates and IFPRI’s validation survey results. Intl Food Policy Res Inst. https://doi.org/10.2499/p15738coll2.133124
Zain NA, Ismail MR (2016) Effects of potassium rates and types on growth, leaf gas exchange and biochemical changes in rice (Oryza sativa L) planted under cyclic water stress. Agric Water Manag 164:83–90. https://doi.org/10.1016/j.agwat.2015.09.022
Zhang H, Xu M, Shi X, Li Z, Huang Q, Wang X (2010) Rice yield, potassium uptake and apparent balance under long-term fertilization in rice-based cropping systems in southern China. Nutr Cycl 88:341–349
Zörb C, Geilfus CM, Dietz KJ (2019) Salinity and crop yield. Plant Biol 21:31–38. https://doi.org/10.1111/plb.12884
Acknowledgements
The authors appreciate the support and laboratory facilities provided by Soil, Water and Environment Discipline, Khulna University, Bangladesh.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Akter, S., Kamruzzaman, M., Khan, M. et al. Enhanced Potassium Fertilization Improved Rice (Oryza sativa) Yield and Nutrient Uptake in Coastal Saline Soil of Bangladesh. J Soil Sci Plant Nutr 23, 1884–1895 (2023). https://doi.org/10.1007/s42729-023-01144-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-023-01144-3