
Abstract
Backgrounds
A fixed root:shoot ratio is widely used to estimate the underground biomass in the carbon (C) cycle in grassland ecosystems, although edaphic and climatic factors may influence the root:shoot ratio. Our current understanding of the effects of the soil texture and water potential on the root:shoot ratio is rather poor.
Methods
Thus, we conducted a pot experiment where we measured the dry weights of the roots and shoots to investigate the responses of ryegrass (Lolium perenne, L.) and alfalfa (Medicago sativa, L.) in terms of C allocation to different soil textures and water potentials.
Results
The root:shoot ratios in ryegrass and alfalfa were significantly affected by the soil texture, but not by the soil water potential (–400 kPa to –40 kPa). Integrated analysis of previous studies indicated a significant average negative correlation between the root:shoot ratio and clay content according to a linear mixed model (k = –0.0045). This correlation might be related to the hydraulic properties of the root-soil contact zone.
Conclusions
We assume that the soil water potential in our experiment was insufficient to affect root hydraulic conductivity, but further evidence is required. Our findings could be useful for future estimates of ecosystem C stocks.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The root biomass is the core link in the soil organic matter allocation and process (Tsegay and Meng 2021). Huang et al. (2021) suggested that the stabilization efficiency of root-derived carbon (C) in the soil is two times greater than that of aboveground residues. However, compared with the aboveground biomass, measuring the root biomass is more challenging due to the difficulty of accurately collecting the root biomass (Tian et al. 2018).
The root:shoot ratio is generally used to estimate the root biomass (Mokany et al. 2006). Fixed allocation coefficients are widely applied although the root:shoot ratio is known to be affected by biotic and abiotic factors (Qi et al. 2019), including the plant species (Warren et al. 2015), stand condition (Costa et al. 2014; Zhang et al. 2022), climate (Reich et al. 2014), and nutrition (Martin et al. 2022). Previous studies (Khan et al. 2012; Moore and Lawrence 2013) demonstrated that the root:shoot ratio is influenced by the soil physical properties, particularly by the differences in the bulk density and thus the penetration resistance, as well as differences in the water-holding capacity and availability of nutrients. Studies have indicated that the soil texture is the main factor that affects the root:shoot ratio (Zhang et al. 2015; Moore and Lawrence 2013). The soil bulk density and shear strength are both related to the soil texture, and they can considerably affect root growth due to mechanical constraints (Barbosa et al 2018; Cai et al. 2021; Nunes et al. 2021). Poeplau and Katterer (2017) found that the soil texture rather than nutrient availability was the most important factor that affected root growth in spring barley, and they also hypothesized that this correlation was due to texture-related differences in water availability. Our current understanding of the effect of soil water on the root:shoot ratio is unclear. In particular, Ma and Fang (2006) reported that the soil water content had no significant effect on the root:shoot ratio. By contrast, Mrak et al. (2019) reported that soil water deficit increased the root:shoot ratio because more roots were needed to absorb water. These different results might be explained by differences in the soil types, indexes analyzed, and plant species used in their studies. The distributions of the soil texture and rainfall usually exhibit some zonality at a regional scale. Therefore, it is important to understand the effects of the soil texture and soil water on the root:shoot ratio to obtain more precise estimates of root-derived C inputs. The soil water potential is determined by the soil texture and water content, and our current understanding of the effect of the soil water potential on plant C allocation is rather poor. It has been shown that the soil water potential is particularly useful for understanding the movement of water from the soil to plants (Kutlu et al. 2018). However, further research is required to determine whether the soil water potential can be used to obtain better prediction of the root:shoot ratio.
In general, the effects of the soil texture and water potential on the root:shoot ratio are not considered adequately when estimating belowground C inputs. Thus, in the present study, we conducted a pot experiment to determine whether the soil texture or water potential is a better predictor of the root:shoot ratio. The plants used in this study were ryegrass and alfalfa, which are major perennial herbaceous plants on the Loess Plateau in China.
2 Materials and Methods
2.1 Soil Samples
Sandy soil and silty clay loam soil samples were collected from the towns of Shenmu (109°30′34.3′′E, 38°48′22.6′′N) and Yangling (108°02′6.4′′E, 34°16′46.7′′N), respectively, in Shaanxi province, China. The sandy soil and silty clay loam soil samples were air dried and passed through a 2-mm sieve. Selected properties of the two soil samples are listed in Table 1. The soil texture was analyzed by laser diffraction using a Mastersizer 2000 (MS-2000, Malvern, UK). The bulk densities of both soil types were measured by using the core sampling method at the beginning and end of the experiment, and they did not vary greatly during the experiment (sandy soil, 1.60 g cm–3; silty clay loam soil, 1.40 g cm–3; and equal mixture of sandy soil and silty clay loam soil, 1.50 g cm–3). Total nitrogen (N) was measured using the Kjeldahl method (Guo et al. 2019). The NH4+-N contents were measured using a Seal Auto Analyzer. Total phosphorus (P) was determined by melt-molybdenum, antimony, and scandium colorimetry. The plant-available P was extracted with bicarbonate, and the concentration was determined with a molybdenum blue colorimetric method using ultraviolet-photometry.
2.2 Soil Characteristics
The sandy soil and silty clay loam soil samples were characterized by clay fractions of 3.36% and 32.31%, respectively, and sand fractions of 90.47% and 6.51% (Table 1). The N and P contents were low in both samples (Table 1). Thus, external N and P were fully supplied.
The relationships between the soil water content and three soil water potentials (–40 kPa, –100 kPa, and –400 kPa) were determined at 20 °C for the sieved soils during drying down using the centrifuge method (Nimmo and Akstin 1988) with a high speed centrifuge (CR21G, Japan). As shown in Table 2, the soil water content increased as the water potential increased in the same soil type. The soil water content of the silty clay loam soil was higher than those of the mixture and sandy soil at the same water potential. At a soil water potential of –40 kPa, the soil water content of the silty clay loam soil was 3.6 times higher and 1.5 times higher than those of the sandy soil and mixture, respectively. At a soil water potential of –400 kPa, the soil water content of the silty clay loam soil was 4.3 times higher and 1.4 times higher than those of the sandy soil and mixture, respectively.
The relationships between the soil water content and hydraulic conductivity were determined by using a retention curve (RETC) computer model (Peters et al. 2021). As shown in Table 2, the hydraulic conductivity increased as the water potential increased in the same soil type. The hydraulic conductivity of the sandy soil was lowest at a soil water potential of –40 kPa.
2.3 Pot Experiment
The sieved soil was packed into prepared pots with a diameter of 15 cm and height of 20 cm. Each treatment was replicated four times, with a total of 36 pots (three soil textures × three soil water potentials × four pots). External N and P were supplied as carbamide and calcium superphosphate, respectively, at approximate rates of 150 and 120 μg g−1 dry soil. The fertilizer was weighed and mixed with the soil samples before packing. No other fertilizer was supplied throughout the experiment. All of the soil substrates were incubated for 60 days before sowing the ryegrass and alfalfa seeds. Thirty seeds were sown in each pot, and the seedlings were thinned to 20 plants per pot at 2 weeks after emergence. The target water content was maintained in each pot by irrigation according to Table 2, and the contents were verified by weighing every 3 days. The pot experiment based on the growth of ryegrass and alfalfa under different soil textures and water potentials is illustrated in Fig. 1, and the amounts of water applied are shown in Fig. 2. The shoots were cut off in the ryegrass and alfalfa flowering periods. The roots were then carefully separated from the soil by washing and sieving in a water tub. The shoots and roots were dried in an oven at 60 °C for 48 h and weighed separately. The 64-day experiment was conducted from June to August during 2019 in a glasshouse at the Institute of Soil and Water Conservation, Yangling, Shaanxi, China. The temperature in the greenhouse was kept between 20 and 30 °C, with no supplemental lighting or other additional conditions.

Images showing the pot experiments conducted with ryegrass (a) and alfalfa (b) before cutting

Amounts of irrigation water applied to ryegrass (a) and alfalfa (b) over time. S40, sandy soil at a soil water potential of –40 kPa; S100, sandy soil at a soil water potential of –100 kPa; S400, sandy soil at a soil water potential of –400 kPa; M40, mixture at a soil water potential of –40 kPa; M100, mixture at a soil water potential of –100 kPa; M400, mixture at a soil water potential of –400 kPa; C40, silty clay loam soil at a soil water potential of –40 kPa; C100, silty clay loam soil at a soil water potential of –100 kPa; C400, silty clay loam soil at a soil water potential of –400 kPa
2.4 Statistical Analysis
The effects of the soil texture, water potential, and their interactions on the shoot biomass, root biomass, and root:shoot ratio were determined by two-way analysis of variance, followed by the least significant difference test. We also compared our results with data obtained in previous studies to assess their generality and representativeness. In particular, eight previous studies that investigated the effects of the soil texture on the root:shoot ratio in different crops (Table 4) were selected. A linear mixed effect model was used to test the effects of different soil textures on the root:shoot ratio, and the continuous clay content variable was treated as a fixed effect. All statistical analyses were conducted using IBM SPSS 20 software. Statistically significant differences were accepted at the 5% significance level.
3 Results
3.1 Biomass Production
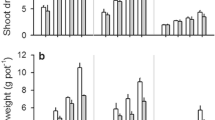
The ryegrass and alfalfa shoot biomasses under different soil textures and water potentials are shown in Fig. 3. The highest shoot biomasses in ryegrass (10.48 ± 0.87 g pot−1) and alfalfa (3.50 ± 0.03 g pot−1) occurred in silty clay loam soil at a soil water potential of –40 kPa. The lowest shoot biomasses in ryegrass (1.16 ± 0.32 g pot−1) and alfalfa (0.72 ± 0.05 g pot−1) occurred in sandy soil at a soil water potential of –400 kPa. The shoot biomass increased as the soil water potential increased (Fig. 3). At the same water potential, the shoot biomass was higher in silty clay loam soil than the mixture and sandy soil. Table 3 shows that the water potential and soil texture had significant (P < 0.001) effects on the shoot biomass in ryegrass and alfalfa.

Shoot biomasses in ryegrass (a) and alfalfa (b) under each treatment. Different uppercase letters (A, B, and C) indicate significant differences in means (P < 0.05) between different soil textures under a specific soil water potential. Different lowercase letters (a, b, and c) indicate significant differences in means (P < 0.05) between different soil water potentials under a specific soil texture
Figure 4 shows the ryegrass and alfalfa root biomasses under different soil textures and water potentials. The highest root biomasses in ryegrass (3.02 ± 0.28 g pot−1) and alfalfa (1.80 ± 0.19 g pot−1) occurred in silty clay loam soil at a soil water potential of –40 kPa. The lowest root biomass in ryegrass (0.63 ± 0.21 g pot−1) and alfalfa (0.46 ± 0.05 g pot−1) occurred in sandy soil at a soil water potential of –400 kPa. The changes in the shoot biomass were similar to those in the root biomass. Figure 4 and Table 3 show that the water potential and soil texture had significant (P < 0.001) effects on the root biomass in ryegrass and alfalfa.

Root biomasses in ryegrass (a) and alfalfa (b) under each treatment. Different uppercase letters (A, B, and C) indicate significant differences in means (P < 0.05) between different soil textures under a specific soil water potential. Different lowercase letters (a, b, and c) indicate significant differences in means (P < 0.05) between different soil water potentials under a specific soil texture
3.2 Root:shoot ratio
Figure 5 shows that the root:shoot ratio was higher in sandy soil than the mixture and silty clay loam soil. The highest root:shoot ratios in ryegrass (0.54 ± 0.03 g pot−1) and alfalfa (0.64 ± 0.08 g pot−1) occurred in sandy soil at a soil water potential of –400 kPa. The lowest root:shoot ratio in ryegrass (0.25 ± 0.02 g pot−1) occurred in silty clay loam soil at a soil water potential of –400 kPa. The lowest root:shoot ratio in alfalfa (0.50 ± 0.02 g pot−1) occurred in silty clay loam soil at a soil water potential of –400 kPa. The average root:shoot ratios for ryegrass were 0.51 in sandy soil, 0.33 in the mixture, and 0.27 in silty clay loam soil, and those for alfalfa were 0.61 in sandy soil, 0.52 in the mixture, and 0.51 in silty clay loam soil. Table 3 shows that the soil textures had significant effects on the root:shoot ratios in ryegrass (P < 0.001) and alfalfa (P = 0.036). A linear function (Y = aX + b) was applied to describe the relationship between the shoot/root biomass (Y) and clay content (X). Average positive slopes of 25.11 (shoot biomass and soil texture) and 5.25 (root biomass and soil texture) were obtained for ryegrass and average positive slopes of 6.67 (shoot biomass and soil texture) and 3.07 (root biomass and soil texture) for alfalfa. Therefore, the significant effect of soil texture on the root:shoot ratio was due to a sharp increase in the shoot biomass and a slow increase in the root biomass as the soil texture changed from coarse to fine-textured soil (Fig. 3, Fig. 4, Table 3). However, the water potential had no significant effects on the root:shoot ratios in ryegrass (P = 0.105) and alfalfa (P = 0.854).

Root:shoot ratios in ryegrass (a) and alfalfa (b) under each treatment. Different uppercase letters (A, B, and C) indicate significant differences in means (P < 0.05) between different soil textures under a specific soil water potential. Different lowercase letters (a, b, and c) indicate significant differences in means (P < 0.05) between different soil water potentials under a specific soil texture
3.3 Integration of Previous Studies
Effects of different soil textures on the root:shoot ratio have been demonstrated in some previous studies. Thus, we collected and analyzed eight previous studies that investigated the relationship between the clay content and root:shoot ratio (Al-Khafaf et al. 1989; Evers and Parsons 2003; Wakeel et al. 2005; Li et al. 2005; Khan et al. 2012; Moore and Lawrence 2013; Poeplau and Katterer 2017; Jia et al. 2019). The linear regression equations and R2 values calculated between the root:shoot ratio are shown in Table 4. A fixed effects linear mixed model was applied to investigate the effect of the clay content on the root:shoot ratio and a significant (P < 0.001) average negative slope of –0.0045 ± 0.0012 (standard error; Table 5) was obtained for the different plant species.
4 Discussion
The root and shoot biomasses in ryegrass and alfalfa increased significantly as the clay content increased. In addition, the root:shoot ratios in ryegrass and alfalfa decreased significantly as the clay content increased. These changes were due to a sharp increase in the shoot biomass and a slow increase in the root biomass as the soil texture changed from coarse to fine-textured soil. Eight previous studies (Table 4) showed that the root:shoot ratio was affected by the soil texture, thereby agreeing with the results obtained in the present study, despite the different plant species investigated. Similarly, Poeplau and Katterer (2017) reported that the soil texture is a major factor that controls the root:shoot ratio in cereals. In addition, Herkelrath et al. (1977) suggested that roots in coarse soil are partly exposed to large air-filled pores, which create a physical discontinuity to hinder the movement of water from the soil to the roots, thereby forcing the plant to increase the root surface area. The physical discontinuity between the root surface and soil particles was experimentally confirmed using the thin-section technique (Kooistra et al. 1992). North and Nobel (1997) investigated and confirmed the extent and size of the root-soil contacts that developed during soil drying. Li et al. (2005) provided evidence that root-soil contact is related directly to the hydraulic conductance at the soil:root interface. The root-soil contact increased as the aggregate or soil particle size decreased according to estimates based on three-dimensional X-ray microtomographs (Schmidt et al. 2012). Cai et al. (2021) suggested that the soil texture is the main factor that determines the uptake of water and soil–plant hydraulics under drought. Therefore, the observed effect of the soil texture on the root:shoot ratio was probably due to differences in the direct root:soil interface and texture-related differences in water availability (Saha et al. 2020), thereby resulting in greater plant C allocation to the underground organs to promote hydraulic conductivity in the coarse-textured soil.
The root and shoot biomasses in ryegrass and alfalfa increased significantly as the soil water potential increased. The soil water potential reflects the availability of soil water to plants, which might influence plant growth (Vogel et al. 2016). A higher soil water potential is more beneficial for the absorption and utilization of soil water by plants, thereby promoting plant growth (Cai et al. 2018). However, the soil water potential had no clear effect on the root:shoot ratio in the present study because of the similar responses by the shoot biomass and root biomass. Liu et al. (2015) reported the occurrence of progressively greater root shrinkage, loss of root-soil contact, and increased resistance to water flow at the soil:root interface as the soil water potential decreased. In addition, Cai et al. (2021) suggested that the soil hydraulic conductivity dropped by several orders of magnitude and became even lower than the root hydraulic conductivity at more negative soil water potentials. However, further studies are needed to determine how the soil hydraulic conductivity changes the root hydraulic conductivity under different soil textures, which might help explain the relationship between the soil water potential and root:shoot ratio.
The results obtained in this study highlight the importance of considering the soil texture for accurately estimating the underground biomass and root-derived C inputs. A 5% increase in the clay content would decrease the annual root-derived carbon input by 0.07 Mg ha−1 year−1 when an average coefficient of − 0.0045 is applied. This demonstrates that considerable errors might occur when constant root:shoot ratio values are used to estimate root-derived C inputs.
5 Conclusions
In the present study, we determined whether the soil texture or water potential is a better predictor of the root:shoot ratio. Experimental observations demonstrated that the root:shoot ratios in ryegrass and alfalfa decreased significantly as the clay content increased, whereas the soil water potential had no clear effect on the root:shoot ratio. Integrated analysis of previous studies also showed that effects of different soil textures on the root:shoot ratio were observed in several other crop types. These effects might be attributable to the weaker root-soil contacts in coarse-textured soil leading to the production of a greater root area to increase the hydraulic conductance. Thus, estimating root-derived carbon inputs by assuming a constant root:shoot ratio for a given soil may lead to considerable errors if the soil texture is not considered, and it is independent of the water potential over a medium range. Understanding root biomass dynamics is essential for improving our knowledge of carbon allocation and storage in terrestrial ecosystems. According to our novel findings, the incorporation of plant-specific root:shoot ratios under different soil textures in models could substantially improve the accuracy of root biomass estimates for applications such as carbon accounting and studies of ecosystem dynamics. Thus, the novel results obtained in the present study might help to improve the accuracy of fine-scale ecosystem carbon stock estimates on local or global scales, which could clearly deviate greatly from the estimates utilized widely for policy and strategy development. Furthermore, more accurate root biomass estimates may facilitate better predictions of local and global carbon cycles under future conditions, particularly under ongoing climate change.
Data Availability
The data used to support the findings of this study are available from the corresponding author upon request.
References
Al-Khafaf S, Aziz FG, Salih HM, Jack FJ (1989) Shoot and root growth and nutrients uptake of wheat as affected by soil layers. Plant Soil 117:59–66. https://doi.org/10.1007/BF02206257
Barbosa LC, de Souza ZM, Franco HC, Otto R, Neto JR, Garside AL, Carvalho JLN (2018) Soil texture affects root penetration in Oxisols under sugarcane in Brazil. Geoderma Reg 13:15–25. https://doi.org/10.1016/j.geodrs.2018.03.002
Cai G, Vanderborght J, Langensiepen M, Schnepf A, Hüging H, Vereecken H (2018) Root growth, water uptake, and sap flow of winter wheat in response to different soil water conditions. Hydrol Earth Syst Sc 22:2449–2470 (0.5194/hess–22–2449–2018)
Cai G, Carminati A, Abdalla M, Ahmed MA (2021) Soil textures rather than root hairs dominate water uptake and soil-plant hydraulics under drought. Plant Physiol 187:858–872. https://doi.org/10.1093/plphys/kiab271
Costa TL, Sampaio EVSB, Sales MF, Accioly LJO, Althoff TD, Pareyn FGC, Albuquerque ERGM, Menezes RSC (2014) Root and shoot biomasses in the tropical dry forest of semi–arid Northeast Brazil. Plant Soil 378:113–123. https://doi.org/10.1007/s11104-013-2009-1
Evers GW, Parsons MJ (2003) Soil type and moisture level influence on Alamo switchgrass emergence and seedling growth. Crop Sci 43:288–294. https://doi.org/10.2135/cropsci2003.0288
Guo XL, Chen L, Zheng RB, Zhang K, Qiu YP, Yue HT (2019) Differences in soil nitrogen availability and transformation in relation to land use in the Napahai wetland, Southwest China. J Soil Sci Plant Nutr 19:92–97. https://doi.org/10.1007/s42729-019-0013-0
Herkelrath W, Miller E, Gardner W (1977) Water uptake by plants: II. The root contact model. Soil Sci Soc Am J 41:1039–1043. https://doi.org/10.2136/sssaj1977.03615995004100060004x
Huang JS, Liu WX, Yang S, Yang L, Peng ZY, Deng M, Xu S, Zhang BB, Ahirwal J, Liu LL (2021) Plant carbon inputs through shoot, root, and mycorrhizal pathways affect soil organic carbon turnover differently. Soil Biol Biochem 160. https://doi.org/10.1016/j.soilbio.2021.108322
Jia JC, Yang XF, Zhang PP, Zhang XC (2019) Use of montmorillonite-enriched sandstone to remediate a sandy mined soil. Ecol Eng 130:101–109. https://doi.org/10.1016/j.ecoleng.2019.01.014
Khan SR, Abbasi MK, Hussan AU (2012) Effect of induced soil compaction on changes in soil properties and wheat productivity under sandy loam and sandy clay loam soils: a greenhouse experiment. Commun Soil Sci Plan 43:2550–2563. https://doi.org/10.1080/00103624.2012.711877
Kooistra MJ, Schoonderbeek D, Boone FR, Veen BW, Van Noordwijk M (1992) Root–soil contact of maize, as measured by a thin–section technique. Plant Soil 139:119–129. https://doi.org/10.1007/BF00012850
Kutlu T, Guber AK, Rivers ML, Kravchenko AN (2018) Moisture absorption by plant residue in soil. Geoderma 316:47–55. https://doi.org/10.1016/j.geoderma.2017.11.043
Li Y, Xu H, Cohen S (2005) Long-term hydraulic acclimation to soil texture and radiation load in cotton. Plant Cell Environ 28:492–499. https://doi.org/10.1111/j.1365-3040.2005.01291.x
Liu XP, Zhang WJ, Wang XY, Cai YJ, Chang JG (2015) Root-soil air gap and resistance to water flow at the soil-root interface of robinia pseudoacacia. Tree physiol 35:1343–1355. https://doi.org/10.1093/treephys/tpv075
Ma WH, Fang JY (2006) R: S ratios of temperate steppe and the environmental controls in Inner Mongolia. Acta Sci Natur Univer Pekin 42:774–778. https://doi.org/10.13209/j.0479-8023.2006.139
Martin T, Culman S, Sprunger CD (2022) Quality or quantity? Determining the impact of fine root traits on soil health in row crop agriculture. J Soil Sci Plant Nutr 22:1–12. https://doi.org/10.1007/s42729-022-00811-1
Mokany K, Raison RJ, Prokushkin AS (2006) Critical analysis of root:shoot ratios in terrestrial biomes. Global Change Biol 12:84–96. https://doi.org/10.1111/j.1365-2486.2005.001043.x
Moore SR, Lawrence KS (2013) The effect of soil texture and irrigation on Rotylenchulus reniformis and cotton. J Nematol 45:99–105
Mrak T, Štraus I, Grebenc T, Gričar J, Hoshika Y, Carriero G, Kraigher H (2019) Different belowground responses to elevated ozone and soil water deficit in three European oak species (Quercus ilex, Q. pubescens and Q. robur). Sci Total Environ 651:1310–1320. https://doi.org/10.1016/j.scitotenv.2018.09.246
Nimmo JR, Akstin KC (1988) Hydraulic conductivity of a sandy soil at low water-content after compaction by various methods. Soil Sci Soc Amer 52:303–310. https://doi.org/10.2136/sssaj1988.03615995005200020001x
North GB, Nobel PS (1997) Root-soil contact for the desert succulent Agave deserti in wet and drying soil. New Phytol 135:21–29. https://doi.org/10.1046/j.1469-8137.1997.00620.x
Nunes MR, de Lima RP, Tormena CA, Karlen DL (2021) Corn seedling root growth response to soil physical quality. Agron J 113:3135–3146. https://doi.org/10.1002/agj2.20705
Peters A, Hohenbrink TL, Iden SC, Durner W (2021) A simple model to predict-hydraulic conductivity in medium to dry soil from the water retention curve. Water Resour Res 57, https://doi.org/10.1029/2020WR029211
Poeplau C, Katterer T (2017) Is soil texture a major controlling factor of root:shoot ratio in cereals? Eur J Soil Sci 68:964–970. https://doi.org/10.1111/ejss.12466
Qi YL, Wei W, Chen CG, Chen LD (2019) Plant root-shoot biomass allocation over diverse biomes: a global synthesis. Glob Ecol Conserv 18:e00606. https://doi.org/10.1016/j.gecco.2019.e00606
Reich PB, Luo Y, Bradford JB, Poorter H, Perry CH, Oleksyn J (2014) Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots. PNAS 111:13721–13726. https://doi.org/10.1073/pnas.1216053111
Saha A, Rattan B, Sekharan S, Manna U (2020) Quantifying the interactive effect of water absorbing polymer (WAP)–soil texture on plant available water content and irrigation frequency. Geoderma 368:114310. https://doi.org/10.1016/j.geoderma.2020.114310
Schmidt S, Bengough AG, Gregory PJ, Grinev DV, Otten W (2012) Estimating root–soil contact from 3D X-ray microtomographs. Eur J Soil Sci 63:776–786. https://doi.org/10.1111/j.1365-2389.2012.01487.x
Tian T, Joann KW, Pierre D (2018) Agricultural practices for crop residue transformation into soil organic matter in cold humid temperate agroecosystems. CAB Reviews 13:1–15. https://doi.org/10.1079/PAVSNNR201813060
Tsegay G, Meng XZ (2021) Impact of Ex-Closure in above and below Ground Carbon Stock Biomass. Forests 12:130. https://doi.org/10.3390/f12020130
Vogel T, Votrubova J, Dusek J, Dohnal M (2016) Mesoscopic aspects of root water uptake modeling–hydraulic resistances and root geometry interpretations in plant transpiration analysis. Adv Water Resour 88:86–96. https://doi.org/10.1016/j.advwatres.2015.12.006
Wakeel A, Aziz T, Aziz T, ul-Hassan A, (2005) Growth and potassium uptake by maize (Zea mays L.) in three soils differing in clay contents. Emir J Food Agr 17:57–62. https://doi.org/10.9755/ejfa.v12i1.5048
Warren JM, Hanson PJ, Iversen CM, Kumar J, Walker AP, Wullschleger SD (2015) Root structural and functional dynamics in terrestrial biosphere models-evaluation and recommendations. New Phytol 205:59–78. https://doi.org/10.1111/nph.13034
Zhang H, Song TQ, Wang KL, Wang GX, Liao JX, Xu GH, Zeng FP (2015) Biogeographical patterns of forest biomass allocation vary by climate, soil and forest characteristics in China. Environ Res Lett 10:044014. https://doi.org/10.1088/1748-9326/10/4/044014
Zhang PP, Wang YQ, Xu L, Sun H, Li RJ, Zhou JX (2022) Factors controlling the spatial variability of soil aggregates and associated organic carbon across a semi-humid watershed. Sci Total Environ 809:151–155. https://doi.org/10.1016/j.scitotenv.2021.151155
Acknowledgements
We acknowledge the anonymous reviewers for their helpful suggestions that greatly improved this manuscript.
Funding
This work was funded by the Program of Shaanxi Academy of Sciences (2020 K–32; 2020 K–09), the Natural Science Foundation of Shaanxi Province (2022JQ–418), and the Open Research Fund of the State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau (A314021402–2019).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception, design, data collection, analyses, and writing and revising the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Junchao, J., Chaodong, L. & Zhangyue, C. Soil Texture Rather Than Water Potential Determines the Root:Shoot Ratio in Ryegrass and Alfalfa. J Soil Sci Plant Nutr 23, 1297–1305 (2023). https://doi.org/10.1007/s42729-022-01121-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-022-01121-2