
Abstract
The aim of this study was to determine the effects of past N fertilizations on soil N mineralization and crop yield formation in the succeeding season in a rice paddy. A field experiment was conducted to compare soil N mineralization and yield attributes of the succeeding crop in a rice paddy following four N application rates (0, 50, 100, and 200 kg N ha−1) during the previous six consecutive crop seasons. Results showed that soil N mineralization potential and maximum and mean N mineralization rates increased with the increase of rates of N applied in the previous seasons, with increases by 14–30% at the N rate of 200 kg N ha−1. Total N uptake by the succeeding crop was significantly and positively related to soil N mineralization potential and maximum and mean N mineralization rates, showing determination coefficients of 0.701–0.729. There was a significant negative relationship between internal N-use efficiency and total N uptake in the succeeding crop; namely, internal N-use efficiency for biomass production decreased by 4 g g−1 for each 1 g m−2 increase in total N uptake. The previous six seasons of N fertilizer applications did not significantly affect grain yield, yield components, total biomass production, and harvest index of the succeeding crop. These results indicate that past N applications can accelerate soil N mineralization and consequently lead to a partial increase in total N uptake by the succeeding crop, but the subsequent crop’s grain yield is not necessarily affected because the increased total N uptake can be offset by the decrease in internal N-use efficiency for biomass production. The finding of this study highlights the need to fully consider soil nutrient supply and crop nutrient-use efficiency when developing new nutrient management practices for rice production.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Rice is one of the most important staple foods in the world, where more than 3.5 billion people rely on it for more than 20% of their daily caloric intake (Seck et al. 2012). Between 1961 and 2019, global rice production has increased from 216 to 755 million tons (Food and Agriculture Organization 2021) and has greatly improved the world’s food supply. Swaminathan (2007) predicted that global rice production will need to increase to 800 million tons by 2025 to meet food demands of the world’s growing population. To reach this goal, great efforts are required to improve rice yields by overcoming yield-limiting factors such as soil nutrient deficiencies (Huang 2021).
Nitrogen (N) is the most limited nutrient for rice growth and development in almost all environments (Yoshida 1981; Chen and Wang 2014). With the development of the fertilizer industry, applying synthetic N fertilizers has been a common practice to supplement N-deficient indigenous soils for rice production and has made an important contribution to increase rice yields (Huang et al. 2020).
In addition to providing a direct source of plant-available N for the in-season crop, the N fertilizer applied to soils can alter the N supply already present in the indigenous soil by changing soil properties such as soil pH and N content. In this regard, it has been reported that the application of N fertilizers may result in decreases in soil pH and increases in soil N content (Guo et al. 2010; Yan et al., 2014), in which both can lead to significant accelerations in soil N mineralization (Li et al. 2019). Moreover, the accelerated soil N mineralization due to N fertilization occurs not only during the in season but also in succeeding seasons (Glendining et al. 1996; Yan et al. 2006; Huang et al. 2018). Therefore, past N fertilizations may influence crop yield formation and N application rates in the succeeding season by accelerating soil N mineralization. However, such information is limited for rice, especially under field conditions.
To bridge this information gap, this study determined the effects of different N application rates for six consecutive seasons on soil N mineralization and yield attributes of the succeeding crop in a rice paddy.
2 Materials and Methods
2.1 Experimental Details
A field experiment was conducted at the research farm of the Crop and Environment Research Center (28°09′ N, 113°37′ E, 43 m asl) at Hunan Agricultural University, China in 2019. The experimental site has a clay soil (Fluvisol, FAO taxonomy) and a humid mid-subtropical monsoon climate. A double-season rice cropping system, i.e., successively growing early- and late-season rice from March to November within a single calendar year, is the most common rice production system in the region surrounding the experimental site.
The field selected for this study had been used for a N-application-rate experiment over six consecutive rice growing seasons (consisting of the late-rice growing season in 2016, the early- and late-rice growing seasons in 2017 and 2018, and the early-rice growing season in 2019) before this study began. In this previously established N-application experiment, four treatments of increasing N fertilization rates (0, 50, 100, and 200 kg N ha−1) were arranged in randomized blocks of treatments with three replicates per treatment. Each plot was 40 m2 and fixed in place throughout the six consecutive seasons. In each season, N fertilizer was applied in three splits: 50% at 1 day before transplanting, 30% at 7 days after transplanting, and 20% at panicle initiation. P fertilizer (60 kg P2O5 ha−1) was applied at 1 day before transplanting. K fertilizer (105 kg K2O ha−1) was applied in two splits: 50% at 1 day before transplanting and 50% at panicle initiation. A floodwater depth of 5–10 cm was maintained from transplanting to 7 days before maturity, when plots were drained. Weeds, insects, and pathogens were intensively controlled by chemicals to avoid yield loss. Crop residues were removed from plots after harvest in each season.
After the rice harvest in the early-rice growing season in 2019, each plot was equally divided into two subplots for this study. Two inbred rice cultivars, Guangluai 4 and Zhongjiazao 17, were grown, one in each of the subplots to establish a split plot design. These two cultivars were selected because they have been widely grown by rice farmers and are among the top ten inbred cultivars grown in rice planting areas in China (China Rice Data Center 2021).
All crop establishment and management practices were the same for each cultivar. Pre-germinated seeds were sown in a seedbed on 7 July. Twenty-day-old seedlings were manually transplanted at a hill spacing of 20.0 cm × 16.7 cm with three seedlings per hill. N fertilizer (150 kg N ha−1) was applied in three parts: 75 kg N ha−1 was applied 1 day before transplanting, 45 kg N ha−1 was applied on the seventh day after transplanting, and 30 kg N ha−1 was applied when rice reached the panicle initiation stage. P fertilizer (60 kg P2O5 ha−1) was applied once the day before transplanting. K fertilizer (105 kg K2O ha−1) was applied equally in two parts, one-half on the day before transplanting and the other half when panicle initiation began. The experimental field was irrigated to maintain a water depth of 5–10 cm from the date of transplanting to 7 days before maturity, when the field was drained. Weeds, insects, and pathogens were controlled by chemicals as required.
2.2 Sampling and Measurements
A homogenized soil sample, consisting of 25 soil cores (3-cm diameter and 20-cm depth), was taken from each 40-m2 plot before initiation of the experiment. The soil cores were collected following a W pattern on the plot. Soil samples were air-dried at room temperature. About 150 g of air-dried soil was randomly taken from each sample and ground and passed through an 18-mesh sieve for determining soil pH, total and available N contents, and N mineralization.
The soil pH was measured with a digital pH meter (PHS-3C, Shanghai Precision & Scientific Instrument Inc., Shanghai, China) after water extraction (soil: water = 1:2.5, w: v). The soil total N content was determined using a Skalar SAN Plus segmented flow analyzer (Skalar Inc., Breda, The Netherlands) after HClO4-H2SO4 digestion. The soil available N content was measured with the NaOH hydrolysis method. The soil N mineralization was determined according to the following procedures: twelve 10-g subsamples of sieved soils were taken from each sample and each subsample was placed in a 100-ml plastic bottle. The bottles were capped after adding 25 ml of deionized water into each of them, and then they were placed in a dark incubator at 35 °C. At 0, 7, 14, 21, 28, and 35 days of incubation, two randomly selected bottles of each sample were taken out from the incubator, 25 ml of extraction solution (4 mol L−1 KCl) was added to each bottle, and then bottles were shaken at 180 rpm for 1 h. The suspension was filtrated using medium-speed qualitative filter papers, and the N concentration in the filtrate was measured by a Skalar SAN Plus segmented flow analyzer. Total mineralized N was calculated by the following equation: Total mineralized N = N concentration in the filtrate × total volume of water added to the soil and extraction solution/soil sample weight, where the total volume of water added to the soil and extraction solution was 50 ml and the soil sample weight was 10 g.
Ten hills of rice plants were sampled diagonally from each subplot at rice maturity, and the plants were separated into straw and panicles. Panicles were counted and hand-threshed, and filled and unfilled spikelets were separated from each other by submerging them in tap water. After air-drying, three subsamples of 30-g filled spikelets and all of the unfilled spikelets were taken to count the number of spikelets. The straw, rachis, and filled and unfilled spikelets were oven-dried at 70 °C to a constant weight to determine their biomass and N concentrations. Each type of plant organ was ground and digested in H2O2-H2SO4, and the N concentration in the digested solution was measured using the Skalar SAN Plus segmented flow analyzer. Panicles per m2, spikelets per panicle, spikelet-filling percentage, grain weight, total biomass production, harvest index, total N uptake, and internal N-use efficiency for biomass production (total biomass production/total N uptake) were calculated. Grain yield was determined from a 5-m2 area in each subplot and adjusted to the standard moisture content of 14%.
2.3 Statistical Analysis
Data were analyzed using DPS 18.10 (Analytical Software, Hangzhou, China). The change in total mineralized N (y) with incubation time (x) was fitted by the logistic regression model: y = k/(1-exp(a-bx)), where k indicates the N mineralization potential, and a and b are parameters of the equation (Fig. 1). Maximum and mean N mineralization rates were calculated using the following two equations: (1) maximum N mineralization rate = kb/4 and (2) mean N mineralization rate = 0.99 k/active mineralization duration, where the active mineralization duration was estimated with y at 99% of k (0.99 k) and solved for x. The soil and crop data were analyzed using analysis of variance and multiple comparisons were analyzed by the LSD test when necessary. Linear regression was employed to assess potential relationships of total N uptake by rice plants with soil available N content and N mineralization parameters (N mineralization potential and maximum and mean N mineralization rates) and between internal N use efficiency and total N uptake in rice.

A schematic diagram for fitting the change in total mineralized N (y) with incubation time (x) by the logistic regression model, y = k/(1-exp(a-bx)), where k indicates the N mineralization potential, and a and b are parameters related to the active mineralization duration, which was estimated with y at 99% of k (0.99 k) and solving for x. Each circle represents the observed total mineralized N at a given incubation time
3 Results
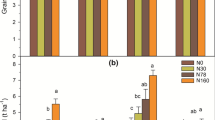
Soil pH decreased with the increase of rates of N applied in the previous seasons (Fig. 2A), whereas opposite trends were observed for soil total and available N contents (Fig. 2B and C). Compared with zero-N application during the previous six consecutive seasons, the past application of 200 kg ha−1 N season−1 for six consecutive seasons decreased soil pH by 7% and increased soil total and available N contents by 20% and 25%, respectively (Fig. 2A–C).

Soil pH (A), total N content (B), and available N content (C) for rice paddy soils following four rates of N application during the previous six consecutive crop seasons. N0, N50, N100, and N200 represent 0, 50, 100, and 200 kg N ha−1 season−1, respectively. Data are means of three replicates, and error bars represent standard errors. For each parameter, data not sharing any lowercase letter are significantly different at the 0.05 probability level
Soil N mineralization potential and maximum and mean N mineralization rates increased with the increase of rates of N applied in the previous seasons (Fig. 3A and B). Compared with zero-N application during the previous six consecutive seasons, the past application of 50, 100, and 200 kg ha−1 N season−1 for six consecutive seasons increased soil N mineralization potential by 9%, 13%, and 14%, respectively, the past application of 200 kg ha−1 N season−1 for six consecutive seasons increased maximum N mineralization rate by 30%, and the application of 100 and 200 kg ha−1 N season−1 for six consecutive seasons increased mean N mineralization rate by 12% and 25%, respectively.

N mineralization potential (A) and maximum and mean N mineralization rates (B) for rice paddy soils following four rates of N application during the previous six consecutive crop seasons. N0, N50, N100, and N200 represent 0, 50, 100, and 200 kg N ha−1 season−1, respectively. Data are means of three replicates, and error bars represent standard errors. For each parameter, data not sharing any lowercase letter are significantly different at the 0.05 probability level
The main effects of past N application rate and rice cultivar and their interactive effects on grain yield, yield components (panicles per m2, spikelets per panicle, spikelet filling percentage, grain weight), total biomass production, and harvest index were not significant (Table 1). Total N uptake and internal N-use efficiency for biomass production in rice were significantly affected by past N application rates (Table 2). Compared with zero-N application during the previous six consecutive seasons, the past application of 200 kg ha−1 N season−1 for six consecutive seasons increased total N uptake by 19% and decreased internal N-use efficiency for biomass production by 11% in the succeeding crop. The main effects of rice cultivar and the interactive effects of past N application rate and rice cultivar on total N uptake and internal N-use efficiency were not significant.
Total N uptake by rice plants was significantly and positively related to soil available N content, with a determination coefficient of 0.767 (Fig. 4A). Total N uptake by rice plants was also significantly and positively related to all three measured soil N mineralization parameters, i.e., soil N mineralization potential and maximum and mean N mineralization rates, showing determination coefficients of 0.701–0.729 (Fig. 4B–D). There was a significant negative relationship between internal N-use efficiency for biomass production and total N uptake in rice (Fig. 5). Internal N-use efficiency for biomass production decreased by 4 g g−1 for each 1 g m−2 increase in total N uptake.

Relationships of total nitrogen (N) uptake by rice plants with soil available N content (A), N mineralization potential (B), maximum N mineralization rate (C), and mean N mineralization rate (D). Data were obtained from a field experiment of two rice cultivars (Guangluai 4 and Zhongjiazao 17) grown in paddy soils that had been applied with one of four rates of N fertilizer (0, 50, 100, and 200 kg N ha−1 season−1) during the previous six consecutive crop seasons. Each data point is the mean of three replicates for each cultivar in each fertilizer treatment, and error bars represent standard errors. * and ** denote significant relationships at the 0.05 and 0.01 probability levels, respectively

Relationship between internal nitrogen (N)-use efficiency for biomass production and total N uptake in rice. Data were obtained from a field experiment of two rice cultivars (Guangluai 4 and Zhongjiazao 17) grown in paddy soils that had been applied with one of four rates of N fertilizer (0, 50, 100, and 200 kg N ha−1 season−1) during the previous six consecutive crop seasons. Each data point is the mean of three replicates for each cultivar in each fertilizer treatment, and error bars represent standard errors. ** denotes a significant relationship at the 0.01 probability level
4 Discussion
Consistent with previous studies (Glendining et al. 1996; Yan et al. 2006; Huang et al. 2018), this study also shows that soil N mineralization was accelerated by past N applications. The accelerated soil N mineralization due to past N applications was associated with decreased soil pH and increased total soil N content (Li et al. 2019). These outcomes highlight the need for a greater fundamental understanding of the effect of past N applications on soil microbes, which play critical roles in the soil N transformation (Robertson and Groffman 2015). In addition, our results indicate that the accelerated effect of applying N fertilizers on soil N mineralization depended on the application rate. Based on the results of this study and the N application rates (150–165 kg ha−1 season−1) used by farmers in the study region (Qin et al. 2013), we estimated that N mineralization potential has risen by more than 10% (or 30 mg kg−1) in this region due to the application N fertilizers (Fig. 6).

Model estimation of the effect of past N application rates on soil N mineralization potential. Data presented in Fig. 1B was used for this model. The red part of the curve shows the soil N mineralization potential at local farmers’ N application rates (150–165 kg ha−1 season−1) in the study region (Qin et al. 2013)
N mineralization is a driver of crop N uptake (Yin et al. 2020). In this study, past N applications led to increased total N uptake by the succeeding crop, which could be partially explained by the accelerated soil N mineralization due to past N applications. Another factor responsible for the increased total N uptake by the succeeding crop was that past N applications resulted in increased soil available N content. Because the magnitude of increased soil available N content was higher than that of increased soil total N content, the increased soil available N content was attributable to both increased soil total N content and accelerated soil N mineralization. These findings indicate that further investigations are required to quantify the contribution of changes in soil N mineralization and availability induced by past N applications to the change in total N uptake by the succeeding crop.
Notably, the N application in previous crop seasons led to increased total N uptake but not any significant effects on grain yield in the subsequent crop. In this regard, it has been well-documented that the effect of N on crop yield depends not only on N uptake by the crop but also on the internal N-use efficiency of the crop (Dobermann 2007; Lobos Ortega et al. 2016; Xu et al. 2021). In this study, a significant negative relationship was observed between total N uptake and internal N-use efficiency for biomass production. This was possibly why the accelerated soil N mineralization due to the past N application led to increased total N uptake but did not have significant effects on grain yield in the succeeding crop. The negative relationship observed between total N uptake and internal N-use efficiency for biomass production could be attributed to the fact that increases in N uptake decreases photosynthetic N-use efficiency (i.e., the ratio of net photosynthetic rate to nitrogen content per unit leaf area) by altering canopy N and light distributions (Jiang et al. 2005; Gu et al. 2017). Moreover, the insignificant effect of accelerated soil N mineralization on grain yield also indicates that the in-season N application rate (150 kg ha−1) used in this study was over the optimum value for crop yield. Therefore, it is feasible to decrease N application rates to less than 150 kg ha−1 in the study region under long-term N fertilization conditions.
In addition to serving as a source for crop N uptake, N mineralized from soil organic matter is also a source of N losses from crop production systems (Martínez-Lagos et al. 2015; Huang et al. 2018; Yin et al. 2020). Over the past several decades, increases in N fertilizer inputs have led to large amounts of N lost by leaching, volatilization, and denitrification (Gao et al. 2020; Huang and Zou 2020) and a range of environmental impacts, including surface water eutrophication (Le et al. 2010), soil acidification (Guo et al. 2010), increased greenhouse gas (N2O) emissions (Davidson 2009; Hu et al. 2020), enhanced N deposition (Liu et al. 2013), and biodiversity loss (Christopher and Tilman 2008). These negative impacts suggest that further investigations are needed to understand the contributions to N losses in crop production from the accelerated soil N mineralization caused by the N application.
5 Conclusions
Applying N fertilizers for six consecutive seasons accelerated soil N mineralization in a rice paddy, which was further stimulated by the increase in N application rates. Although the increased mineralization rates due to the past N application led to a partial increase in total N uptake by the succeeding crop, the subsequent crop’s grain yield was not significantly affected because the increased total N uptake was offset by the decrease in internal N-use efficiency for biomass production. The results of this study highlight the need to take full consideration of soil nutrient supplies and crop nutrient-use efficiency when developing new nutrient management practices for rice production.
References
Chen LS, Wang K (2014) Diagnosing of rice nitrogen stress based on static scanning technology and image information extraction. J Soi Sci Plant Nutr 14:382–393. https://doi.org/10.4067/S0718-95162014005000030
China Rice Data Center (2021) Database of rice cultivars and genealogies in China. Available via DIALOG. https://www.ricedata.cn/variety/index.htm. Accessed 22 June 2021
Christopher MC, Tilman D (2008) Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 451:712–715. https://doi.org/10.1038/nature06503
Davidson EA (2009) The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860. Nat Geosci 2:659–662. https://doi.org/10.1038/ngeo608
Dobermann A (2007) Nutrient use efficiency—measurement and management. In: Krauss A, Isherwood K, Heffer P (eds) Fertilizer Best Management Practices: General Principles, Strategy for Their Adoption and Voluntary Initiatives Versus Regulations. International Fertilizer Industry Association, Paris, France, pp 1–28
Food and Agriculture Organization (2021) FAOSTAT database: crops. Available via DIALOG. http://www.fao.org/faostat/en/#data/QC. Accessed 22 June 2021
Gao J, Luo J, Lindsey S, Shi Y, Sun Z, Wei Z, Wang L (2020) Benefits and risks for the environment and crop production with application of nutrification inhibitors in China. J Soi Sci Plant Nutr 21:497–512. https://doi.org/10.1007/s42729-020-00378-9
Glendining MJ, Powlson DS, Poulton PR, Bradbury NJ, Palazzo D, Li X (1996) The effect of long-term applications of inorganic nitrogen fertilizer on soil nitrogen in the Broadbalk Wheat Experiment. J Agric Sci 127:347–363. https://doi.org/10.1017/S0021859600078527
Gu J, Chen Y, Zhang H, Li Z, Zhou Q, Yu C, Kong X, Liu L, Wang Z, Yang J (2017) Canopy light and nitrogen distributions are related to grain yield and nitrogen use efficiency in rice. Field Crops Res 206:74–85. https://doi.org/10.1016/j.fcr.2017.02.021
Guo JH, Liu XJ, Zhang Y, Shen JL, Han WX, Zhang WF, Christie P, Goulding KWT, Vitousek PM, Zhang FS (2010) Significant acidification in major Chinese croplands. Science 327:1008–1010. https://doi.org/10.1126/science.1182570
Hu Q, Liu T, Jiang S, Cao C, Li C, Chen B, Liu J (2020) Combined effects of straw returning and chemical N fertilization on greenhouse gas emissions and yield from paddy fields in northwest Hubei Province, China. J Soi Sci Plant Nutr 20:392–406. https://doi.org/10.1007/s42729-019-00120-0
Huang M, Lei T, Cao F, Chen J, Shan S, Zou Y (2020) Grain yield responses to nitrogen rate in two elite double-cropped inbred rice cultivars released 41 years apart. Field Crops Res 259:107970. https://doi.org/10.1016/j.fcr.2020.107970
Huang M, Shan S, Cao F, Chen J, Zou Y (2018) The potential of naturally occurring fallow weeds to scavenge nitrogen in rice cropping systems. Ecol Indic 93:183–187. https://doi.org/10.1016/j.ecolind.2018.05.002
Huang M, Zou Y (2020) Reducing environmental risk of nitrogen by popularizing mechanically dense transplanting for rice production in China. J Integr Agr 19:2362–2366. https://doi.org/10.1016/S2095-3119(20)63155-0
Huang M (2021) The decreasing area of hybrid rice production in China: causes and potential effects on Chinese rice self-sufficiency. Food Secur. https://doi.org/10.1007/s12571-021-01199-z
Jiang L, Dong D, Gan X, Wei S (2005) Photosynthetic efficiency and nitrogen distribution under different management and relationship with physiological N-use efficiency in three rice genotypes. Plant Soil 271:321–328. https://doi.org/10.1007/s11104-004-3116-9
Le C, Zha Y, Li Y, Sun D, Lu H, Yin B (2010) Eutrophication of lake waters in China: Cost, causes, and control. Environ Manage 45:662–668. https://doi.org/10.1007/s00267-010-9440-3
Li Z, Tian D, Wang B, Wang J, Wang S, Chen HYH, Xu X, Wang C, He N, Niu S (2019) Microbes drive global soil nitrogen mineralization and availability. Glob Chang Biol 25:1078–1088. https://doi.org/10.1111/gcb.14557
Liu X, Zhang Y, Han W, Tang A, Shen J, Cui Z, Vitousek P, Erisman JW, Goulding K, Christie P, Fangmeier A, Zhang F (2013) Enhanced nitrogen deposition over China. Nature 494:459–462. https://doi.org/10.1038/nature11917
Lobos Ortega I, Alfaro M, Martinez-Lagos J (2016) Soil nitrogen contribution to grasslands yield in southern Chile its implication for nitrogen use efficiency. J Soil Sci Plant Nutr 16:310–322. https://doi.org/10.4067/S0718-95162016005000027
Martínez-Lagos J, Salazar F, Alfaro M, Rosas M, Macías F (2015) Nitrogen mineralization in a silandic andosol fertilized with dairy slurry and urea. J Soil Sci Plant Nutr 15:60–75. https://doi.org/10.4067/S0718-95162015005000006
Qin J, Impa SM, Tang Q, Yang S, Yang J, Tao Y, Jagadish KSV (2013) Integrated nutrient, water and other agronomic options to enhance rice grain yield and N use efficiency in double-season rice crop. Field Crops Res 148:15–23. https://doi.org/10.1016/j.fcr.2013.04.004
Robertson GP, Groffman PM (2015) Nitrogen transformations. In: Paul EA (ed) Soil Microbiology, Ecology and Biochemistry. Academic Press, Burlington, pp 421–446
Seck PA, Diagne A, Mohanty S, Wopereis MCS (2012) Crops that feed the world 7: Rice. Food Secur 4:7–24. https://doi.org/10.1007/s12571-012-0168-1
Swaminathan MS (2007) Science and shaping the future of rice. In: Aggarwal PK, Ladha JK, Singh RK, Devakumar C, Hardy B (eds) Science, Technology, and Trade for Peace and Prosperity. International Rice Research Institute, Los Baños, Philippines, pp 3–14
Xu L, Yuan S, Wang X, Yu X, Peng S (2021) High yields of hybrid rice do not require more nitrogen fertilizer than inbred rice: a meta-analysis. Food Energy Secur 10:341–350. https://doi.org/10.1002/fes3.276
Yan DZ, Wang DJ, Sun RJ, Lin JH (2006) N mineralization as affected by long-term N fertilization and its relationship with crop N uptake. Pedosphere 16:125–130. https://doi.org/10.1016/S1002-0160(06)60034-9
Yan X, Ti C, Vitousek P, Chen D, Leip A, Cai Z, Zhu Z (2014) Fertilizer nitrogen recovery efficiencies in crop production systems of China with and without consideration of the residual effect of nitrogen. Environ Res Lett 9:095002. https://doi.org/10.1088/1748-9326/9/9/095002
Yin X, Beaudoin N, Ferchaud F, Mary B, Strullu L, Chlébowski F, Clivot H, Herre C, Duval J, Louarn G (2020) Long-term modelling of soil N mineralization and N fate using STICS in a 34-year crop rotation experiment. Geoderma 357:113956. https://doi.org/10.1016/j.geoderma.2019.113956
Yoshida S (1981) Fundamentals of Rice Crop Science. International Rice Research Institute, Los Baños
Funding
This work was supported by the National Key R&D Program of China (2017YFD0301503).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Huang, M., Liu, Y., Cao, F. et al. Residual Effects of Nitrogen Application for Six Consecutive Crop Seasons on Soil Nitrogen Mineralization and the Succeeding Crop Yield in a Rice Paddy. J Soil Sci Plant Nutr 22, 1052–1059 (2022). https://doi.org/10.1007/s42729-021-00714-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-021-00714-7