
Abstract
Little is known about the effect of water availability on P pools in the detritusphere. Detritusphere was generated with two plant residues: 100% barley straw, C/N 95, C/P 255; 75% barley straw with 25% young faba bean shoot, C/N 74, C/P 200. Residues were placed between two PVC caps filled with soil at − 0.078 MPa, separating them from the soil surface with fine nylon mesh. Unamended controls were without residue between the two meshes. After 2 weeks, soil at 0–2 mm distance from the surface was collected and the soil water availability was either maintained at − 0.078 MPa or reduced to − 0.320 and − 1.700 MPa by drying in a fan-forced oven. Bioavailable P pools, available N, and microbial N were measured 1, 14, and 28 days after adjusting to different water availabilities. Soil respiration was measured over 28 days. Soil water availability had a stronger effect on respiration, available N, and microbial biomass N (MBN) in the mix than the control or barley. With the mix compared with − 1.700 MPa, cumulative respiration from day 0 to 14, available N and MBN were five, two, and three-fold higher at − 0.078 MPa. In the control or with barley, differences between the two water contents were two or less fold. Low water availability limits microbial activity and nutrient fluxes at high substrate availability as in the mix but has little effect when substrate availability is low even at high water availability as in the control and with barley.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Dry periods are common in Mediterranean ecosystems and their intensity and frequency are expected to increase in the future (Houghton et al. 2001). Soil water content is a major factor affecting soil nutrient availability (Clancy et al. 1995; Díaz and Roldán 2000; Schimel et al. 2007) and decomposition of organic amendments through its effect on microbial activity.
Soil water availability is best described as soil water potential (McLaren and Skujins 1968). At low soil water content, water potential is more negative (lower) because water is bound more tightly to soil particles and held in smaller pores than at high water content (Brady and Weil 2002). Thus, water is less available for microbes as water potential becomes more negative. At low soil water content, substrate diffusion is restricted due to the thin water films around aggregates and water may be drawn out of microbial cells. Consequently, soil water content is an important regulator of microbial activity. But at or above saturation, microbial activity is low due to poor aeration and limited availability of O2 (Rovira 1953). The effect of water availability on soil nutrient availability and microbial activity has been studied extensively using bulk soil samples. For example, Poll et al. (2008) showed that litter mass loss declined with soil water content. Soil respiration declines with decreasing (more negative) water potential (Christian and Waltho 1966; Mozumder and Caroselli 1970).
In experiments where crop residues were mixed into soil and maintained at water content optimal for microbial activity, residues with C/P above 200 induce net P immobilisation and depletion of available P (Khuyen and Marschner 2017). In contrast, decomposition of residues with lower C/P ratio results in higher P availability and other P pools (Alamgir et al. 2012; Umrit and Friesen 1994). During decomposition of crop residues, P availability increases in the soil due to (i) release of P, (ii) exchange of organic acid anions produced during residue decomposition with sorbed P (Bolan et al. 1994; Hue et al. 1994), and organic acid anions forming metal complexes which reduces P sorption. For example, compared to unamended soil, mixing of soil with low (63; faba bean) or medium (232; chickpea) C/P crop residues increased available P, labile P pools, microbial P, and NaHCO3-extractable inorganic P (Alamgir and Marschner 2013). But these P pools were little affected by amendment with high C/P residue (640; white lupin). Decomposition rate of residues is not only influenced by the C/nutrient ratio, but also by their organic C composition as cellulose or lignin is more difficult to decompose than less complex or water-soluble compounds (Xu et al. 2017). Mature plant residues such as straw have a higher concentration of cellulose and lignin than residues of young plant tissues (Rencoret et al. 2011).
The detritusphere is defined as the layer of soil directly adjacent to the decomposing crop residue (< 5 mm into the soil), and usually characterised by a higher concentration of easily available compounds than the bulk soil, particularly in the early stages of residue decomposition (Poll et al. 2010; Liu et al. 2011). For example, C and N turnover can be higher within a distance of 3–4 mm from litter than in bulk soil (Gaillard et al. 1999; Kandeler et al. 1999). However, little is known about the influence of water availability on P pools and P dynamics in the detritusphere of crop residues with different C/P ratios. In the field, the detritusphere is an important interface between soil and crop residues, for example under a layer of residues on the soil surface or around residue clumps in the soil. Particularly in semi-arid regions, water availability in the top soil can vary depending on rainfall and temperature (Sun et al. 2017). When crop residues are used to improve plant nutrient uptake, it is important to understand nutrient availability in the detritusphere at different water availabilities.
We hypothesised that (1) soil respiration, microbial biomass, and P pools are greater at high (− 0.078 MPa) than low (− 1.700 MPa) water availability, due to higher water and substrate availability, and (2) the difference in soil respiration, microbial biomass, and P pools between high (− 0.078 MPa) and low (− 1.700 MPa) water availability will be greater with low C/P residue than one with high C/P ratio. The second hypothesis is based on the assumption that with high C/P residue, substrate availability will limit soil respiration, microbial biomass, and P pools.
2 Materials and Methods
A loamy sand was collected from 0 to 10 cm on Waite Campus, The University of Adelaide, South Australia (Longitude 138° 38′ E, Latitude 35° 6′ S); the site had been under permanent pasture over 80 years but recently cropped with oats. This area has a Mediterranean climate, characterised by cool, wet winters and hot, dry summers with occasional short, heavy rainfall events. The soil is a Chromosol in Australian soil classification and a Rhodoxeralf in US Soil Taxonomy. Soil from six different sites was combined and mixed. The soil was dried at 40 °C and sieved to 2 mm. The properties of the soil are as follows: pH 6.8 (1:5 soil/water), EC (1:5) 0.1 dS m−1, clay 25%, sand 37%, silt 37%, total P 302 mg kg−1, total organic C 17 g kg−1, total organic N 1.5 g kg−1, bulk density 1.3 g cm−3, and maximum water holding capacity (WHC) 349 g kg−1.
2.1 Experimental Design
Detritusphere soil was generated over 14 days at − 0.078 MPa to maximise the effect of the crop residues on detritusphere soil properties. Two crop residues used are mature barley straw (Hordeum vulgare L.) alone or mixed with young faba bean residue (Vicia faba L.) at a 75:25 ratio. The mix was used because, in a previous study, faba bean alone had resulted in very high P pools and N availability compared to barley alone (Erinle et al. 2018). Results of the two residues could only be statistically compared after log-transformation of the data. The residues were dried in a fan-forced oven (40 °C) ground and then sieved to 0.25–2 mm (Table 1).
Detritusphere soil was generated in microcosms as described in Ha et al. (2007) and Erinle et al. (2018). Briefly, 90 g dry soil was filled into caps of PVC tubes (2 cm height, 7 cm diameter) and packed to a bulk density of 1.3 g cm−1. To activate the soil microbes, reverse osmosis (RO) water was added to achieve 50% WHC (equivalent to − 0.078 MPa), then the soil was incubated in the dark at 20–25 °C for 7 days during which it was maintained at 50% WHC. In other experiments with this soil, soil respiration was maximal at − 0.078.
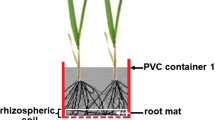
After the pre-incubation, the open ends of the PVC caps were covered with fine nylon mesh cut into circles (mesh size 0.1 mm × 0.8 mm; ⌀ = 85 mm). Barley residues alone or the 75:25 mix of barley and faba bean residue (3.6 g per microcosm, equivalent to 20 g kg−1) were spread on the mesh of one PVC cap and then covered by the open end of a second. The open ends of the PVC caps were pressed together and secured with rubber bands. With this residue amount, a thin residue layer covered the entire mesh area and the cap ends could be closely fitted to avoid loss of residue during the experiment Thus, the thin layer of residue was between the two open ends of the caps, separated from the soil by nylon mesh. The control had no crop residue between the meshes. The microcosms were placed in the dark at 20–25 °C for 2 weeks during which soil moisture was maintained at − 0.078 MPa by weight. The closed ends of the PVC caps had four holes to allow gas exchange and addition of water to maintain the soil water content. The two-week duration was used because, in a preliminary study, detritusphere P pools changed little after 2 weeks (data not shown). After 2 weeks, the two PVC caps were separated and the two layers of mesh with the residues in between were removed without disturbing the soil surface. Residue dry weight after 2 weeks was 2.7 g for barley straw and 2.1 g for the mix. Soil in 0–2 mm from the surface was collected as the detritusphere soil. Fifteen grams of soil were collected from each of the two PVC caps of a microcosm which were combined to 30 g per replicate for the following period.
Detritusphere soil (30 g) was placed into PVC cores (1.85 cm radius, 5 cm height) which had one end covered by nylon mesh base (7.5 μm, Australian Filter Specialist); soil bulk density was adjusted to 1.3 g cm−1. Then soil water availability was either maintained at − 0.078 MPa or adjusted to − 0.320 MPa and − 1.700 MPa by placing the cores in a fan-forced oven at 30 °C for 4 and 10 h, respectively. This water availability was maintained in the following 28 day period by weight and adding water if necessary. These water availabilities were selected because Xue et al. (2017) showed that respiration and microbial growth in this soil decreased with decreasing soil water availability from − 0.078 to − 1.700 MPa in planted and unplanted soil. Cores were destructively sampled for analyses 1, 14, and 28 days after adjusting to different water availabilities. There were four replicates per treatment and sampling time. After reaching the desired water content, cores sampled on day 1 were placed individually into 1-L jars with gas-tight lids which had septa to allow quantification of the headspace CO2 concentration (see below). The jars were incubated in the dark at 20–25 °C. After removal of the cores on day 1, the cores for the day 14 sampling were placed in the jars. This was repeated on day 14, placing the cores for the day 28 sampling in the jars. Cores not in jars were incubated in the same conditions in trays.
2.2 Measurements
Analyses were carried out as described in Erinle et al. (2018). Soil maximum water holding capacity was measured matric potential = − 10 kPa (Wilke 2005). Soil texture was determined according to Ge and Or (2002). Soil pH was determined in a 1:5 (w/v) soil to reverse osmosis (RO) water ratio (Rayment and Higginson 1992). Total organic C of soil and residues was determined by wet oxidation (Walkley and Black 1934). Total N in soil and plant residues was determined using the Kjeldahl method (McKenzie and Wallace 1954). Total P in soil and plant residues was determined by digesting samples with a mixture of HNO3 and HClO4 and measured in the digest by the phosphovanadomolybdate method (Hanson 1950). To determine water-soluble P in the residues, 30 mL of hot (85 °C) RO water was added to 1 g residue, then shaken for 2 h and filtered through a Whatman #42 filter paper (modified from Konieczynski and Wesolowski 2007). The filtrate P concentration was measured colorimetrically (630 nm) using the malachite green method (Ohno and Zibilske 1991). Water extractable carbon was determined following a similar procedure, but with 0.25 g residue. Organic C in the filtrate was determined by the Walkey and Black method.
Soil respiration was measured as described in Setia et al. (2011) using a Servomex 1450 infra-red analyser (Servomex Group, Crowborough, UK). Linear regression based on the injection of known amounts of CO2 in similar jars was used to define the relationship between CO2 concentration and analyser reading. Residue decomposition was measured separately using 3.6 g residue mixed with 1.8 g soil. Respiration was measured over 2 weeks as described above. Cumulative respiration was about twofold higher with the mix than with barley (Table 1).
Soil P pools were measured using a modification of DeLuca et al. (2015). Two pools of the DeLuca method were not measured. CaCl2 P (representing the soluble and weakly adsorbed inorganic P) was replaced by anion extractable P, both are available P pools. Instead of phosphatase labile organic P, microbial biomass P was measured because we found in earlier studies that the phosphatase P concentration in this soil was very low and variable. Biomass P is a potentially available P pool from which P is released upon biomass turnover. To determine Citric acid- and HCl-extractable P, 0.5 g of soil was extracted in parallel by shaking with 10 ml of each extractant (10-mM citric acid or 1 M HCl, respectively) on an end-over-end shaker in separate 50-mL falcon tubes for 3 h. P pools extracted are potentially bioavailable inorganic P pools sorbed to clay particles or weakly bound in inorganic precipitates, respectively. Two other pools, resin (available) P and microbial biomass P (MBP), were determined using the anion exchange resin method (Kouno et al. 1995) with hexanol fumigation. The P concentration in the extracts was by the malachite-green method as described in Ohno and Zibilske (1991). MBP is the difference in P concentration between fumigated and un-fumigated soil (Kouno et al. 1995). No correction factor was used because the recovery of a P spike in this soil was 98% (Butterly et al. 2010). Available N (ammonium and nitrate) concentration was measured after 1 h shaking with 2 M KCl in a 1:5 soil to extractant ratio. Ammonium-N and nitrate-N were determined after Willis et al. (1996) and Miranda et al. (2001), respectively. Microbial biomass N was determined by chloroform fumigation-extraction with 0.5 M K2SO4 at 1:4 soil to extractant ratio (Moore et al. 2000) followed by ammonium in the extract according to Willis et al. (1996). Microbial biomass N is the difference in NH4+ concentration between fumigated and non-fumigated samples divided by 0.57 (Moore et al. 2000).
2.3 Statistical Analysis
There were four replicates per treatment and sampling time. Data was analysed by two-way repeated measures ANOVA with fixed factors water availability (− 0.078 MPa, − 0.320 MPa, and − 1.700 MPa) and residue treatment (control without residue, barley, and barley+faba bean residues). Separately for each sampling time, Tukey’s multiple comparison test was used to significant effects among treatments (P ≥ 0.05). The statistical analyses were carried out in Genstat v18.2 (VSN International Ltd., UK).
3 Results
Compared with the control, respiration rates were about twofold higher with barley and threefold higher with the mix (faba bean:barley residue at 25:75) (Fig. 1a–c). Respiration rates were stable in the control but declined over time with residues, particularly with the mix. At − 0.078 MPa compared with − 1.700 MPa, respiration rates were about fivefold higher with the mix, threefold with barley and twofold higher with the control. Water availability had no effect on cumulative respiration in the control or with barley (Fig. 2). With the mix, where cumulative respiration was two to threefold higher than in the control, cumulative respiration was about fivefold higher at − 0.078 MPa than at − 1.700 MPa.

Respiration rate over 28 days in a unamended control, b detritusphere soil of mature barley straw, and c detritusphere of barley straw + faba bean residue (ratio 0.75:0.25) at different water availabilities (− 0.078 MPa, − 0.320 MPa, and − 1.700 MPa) (n = 4)

Cumulative respiration from day 0 to 14 and day 15 to 28 in control and detritusphere soil of barley and barley-faba bean mix at different water availabilities (− 0.078 MPa, − 0.320 MPa, and − 1.700 MPa). Columns in with different letters are significantly different (P ≥ 0.05, n = 4)
One day after water availability adjustment, P pools, MBP, and available N were higher with the mix than in the control at all water availabilities (Fig. 3). There was little or no difference in P pools, available N and MBN between control and with barley. In all residue treatments (residues and control), citrate-P was about 10% higher at − 1.700 MPa than with higher water availability (Fig. 3a). Citrate-P was about threefold higher with the mix than the control or with barley. HCl-P decreased with increasing water availability and was about 20% higher with − 1.700 MPa than at − 0.078 MPa. HCl-P was about 40% higher with the mix than in the control and with barley (Fig. 3b). In all residue treatments, resin-P was about twofold higher at − 1.700 MPa than at − 0.078 MPa. Resin-P was two to threefold higher with the mix than with barley or the control (Fig. 3c). In the control and with the mix, MBP was higher at − 0.078 MPa than − 1.700 MPa with greater differences in the control than the mix (Fig. 3d). MBP with barley was not influenced by water availability. Compared to the control, MBP with barley was threefold higher at − 1.700 MPa and 25% higher at − 0.078 MPa. With the mix, MBP was two to threefold higher than the control at − 1.700 MPa and − 0.078 MPa.

Citrate-P (a), HCl-P (b), resin-P (c), microbial biomass P (d), available N (e), and microbial biomass N (f) (mg kg−1) 1 day after drying to different water availabilities (− 0.078, − 0.320, and − 1.700 MPa) in control  , detritusphere soil of barley straw
, detritusphere soil of barley straw  and barley straw + faba bean residue (ratio 0.75:0.25)
and barley straw + faba bean residue (ratio 0.75:0.25)  . Columns with different letters are significantly different (P ≥ 0.05, n = 4)
. Columns with different letters are significantly different (P ≥ 0.05, n = 4)
Only in the mix, available N was higher at − 1.700 MPa than at − 0.078 MPa, where it was about twofold higher at − 1.700 MPa (Fig. 3e). Available N 1 day after water availability, adjustment was about 10-fold higher with the mix than in the control or with barley. In all residue treatments, MBN was lowest at − 1.700 MPa. MBN did not differ between control and detritusphere at − 1.700 and − 0.320 MPa, but was about twofold higher with the mix than in the control and with barley at − 0.078 MPa (Fig. 3f).
At all water availabilities, citrate-P, HCl-P, and resin P 14 and 28 days after water availability adjustment were between two and threefold higher with the mix than in the control which differed little from barley detritusphere (Fig. 4). The only exception was citrate-P at − 0.078 MPa on day 14 which was 50% lower with barley and 30% higher with the mix compared to the control (Fig. 4a). On day 14, citrate-P with barley and the mix was about 20% higher at − 1.700 MPa than at − 0.078 MPa. But in the control, it was 40% higher at − 0.078 MPa than at − 1.700 MPa. On day 28, citrate-P was not influenced by water availability. HCl-P on day 14 was 10–20% higher at − 1.700 MPa than at − 0.078 MPa with the mix and barley (Fig. 4b). HCl-P in the control on day 14 and in all residue treatments on day 28 was not influenced by water availability. Resin-P with barley and the mix was about 10–20% higher at − 1.700 MPa than at − 0.078 MPa on days 14 and 28 (Fig. 4c). In the control, resin-P was 20% higher at − 1.700 MPa than at − 0.078 MPa on day 14 but not influenced by water availability on day 28. MBP on day 14 was about 25% lower at − 1.700 MPa than at − 0.078 MPa in all residue treatments (Fig. 4d). But on day 28, MBP was influenced by water availability only with the mix where it was 30% lower at − 1.700 MPa than at − 0.078 MPa. On both days, MBP was between 50% and twofold higher with barley and two to three-old higher with the mix compared with the control. Available N on days 14 and 28 was about twofold higher at − 1.700 MPa than at − 0.078 MPa (Fig. 4e). Compared with the control, available N on day 14 was similar with barley and about twofold higher with the mix at 1.700 MPa and − 0.078 MPa. But residue treatments differed a little in available N at − 0.320 MPa. Available N on day 28 was twofold higher with the mix than the control at − 1.700 MPa, but the two treatments did not differ in available N at higher water availability. Available N on day 28 was about 20% higher in the control than with barley. In all residue treatments and both sampling days, MBN increased with water availability (Fig. 4f). It was about threefold higher at − 0.078 MPa than at − 1.700 MPa. Compared with the control, MBN was about twofold higher with barley and threefold higher with the mix.

Citrate-P (a), HCl-P (b), resin-P (c), microbial biomass P (d), available N (e), and microbial biomass N (f) (mg kg−1) after 14- and 28-day incubation at different water availabilities (− 0.078, − 0.320, and − 1.700 MPa) in control  , detritusphere soil of barley straw
, detritusphere soil of barley straw  and barley straw + faba bean residue (ratio 0.75:0.25)
and barley straw + faba bean residue (ratio 0.75:0.25)  . For each sampling time separately, columns with different letters are significantly different (P ≥ 0.05, n = 4)
. For each sampling time separately, columns with different letters are significantly different (P ≥ 0.05, n = 4)
4 Discussion
Based on the results of this experiment, Hypothesis 1 (soil respiration, microbial biomass, and P pools are greater at high (− 0.078 MPa) than low (− 1.700 MPa) water availability, due to higher water and substrate availability) can be confirmed. Hypothesis 2 (the difference in soil respiration, microbial biomass, and P pools between high (− 0.078 MPa) and low (− 1.700 MPa) water availability will be greater with low C/P residue than one with high C/P ratio) can only be confirmed for some sampling times and measured parameters.
Soil water availability had a stronger effect on cumulative respiration, available N, and MBN in the mix than the control or barley. The effect of water availability on P pools was smaller than that on respiration, available N, and MBN probably because of flux of P among P pools. Depletion of a given P pool, particularly available P, and MBP triggers the release of P from other pools including citrate and HCl-P and possibly pools not assessed by the Deluca method. Compared with − 0.078 MPa, the low water availability at − 1.700 MPa will result in thinner water films around aggregates and disruption of pore continuity (Tecon and Or 2017). This will restrict substrate diffusion and also diffusion of enzymes away from microbial cells (Geisseler et al. 2012). The restricted diffusion will have a greater impact on substrate availability when substrate supply is high at − 0.078 MPa as in the mix. During the 2 weeks of detritusphere generation, only 0.9 g of barley straw had been decomposed compared with 1.5 g of the mix, and cumulative respiration during this time was twofold higher with the mix than with barley. Mass loss during detritusphere generation was not only due to CO2 release but also diffusion of water-soluble compounds into the detritusphere, particularly with the mix. Small residue particles that fell through the mesh may also contribute to the mass loss. Due to the high C/N ratio and complex nature of organic C of mature barley straw, any barley residue particles that had fallen through the mesh would have been more difficult to decompose than faba bean residues from the mix. Therefore, substrate availability in barley detritusphere soil was lower than in the detritusphere of the mix. Available N, MBN, and P pools were up to two or more fold greater with the mix than barley although the C/N and C/P ratio differed little (C/N 95 and 74, C/P 250 and 200, respectively). This suggests that presence of a small proportion of easily decomposable faba bean residue induced a priming effect of both barley residue and native organic matter (Stewart et al. 2015), particularly at high water availability. It is also possible that the presence of sustained C source from barley enhanced respiration and thus decomposition of faba bean residues. With barley and the control, it is likely that substrate availability limited microbial activity even at high water availability and a reduction in water availability had little effect on the measured parameters. With the mix, the higher substrate availability − 0.078 MPa compared with − 1.700 MPa led to higher MBN and MBP which, in turn, resulted in lower available N and resin-P. Differences between − 0.320 MPa and the other two water availabilities only occurred with the mix. This suggests that only with high substrate supply the measured parameters are influenced by small changes in water availability. The higher resin-P, citrate-P, HCl-P, and available N at − 1.700 MPa could also be due to chemical processes induced by drying or by extraction for analysis. Drying can increase P extractability, possibly due to disruption of aluminium-phosphate complexes (Erich and Hoskins 2011) or by inducing death and lysis of microbial cells due to desiccation (Miller et al. 2005; Schimel et al. 2007). Addition of an extractant to the dry soil can cause cell lysis (Turner et al. 2003) and disruption of soil aggregates which exposes additional organic matter and clay surfaces similar to the effect of rapid rewetting of dry soil (Erich and Hoskins 2011; Wang et al. 2014). The latter could increase extractability of P bound to soil particles. The low MBN and MBP at − 1.700 MPa may therefore be due not only to low in situ microbial biomass but also to cell lysis induced during extraction for analysis.
The higher P concentration in the mix compared to barley induced higher citrate-P, HCl-P, and resin P at all sampling times. MBP on the other hand differed a little between mix and barley 1 day after adjustment of water availability. This suggests that microbes had not yet been able to take up P released from the mix within 1 day even at high water availability. Later however, P released during the decomposition of the mix resulted in higher MBP than with barley.
Available N, MBN, and P pools were lower after 14 and 28 days than after 1 day. This may be due to the conversion of available N into microbial metabolites, microbial biomass turnover; and for P, conversion into P pools not measured by the Deluca method, i.e. more stable, not directly biologically available. Relative differences between residue and water availability treatments were similar at all sampling times for available N and MBN, resin P, and MBP. But for citrate-P and HCl-P, treatment differences that occurred on day one had disappeared on day 28, probably due to the flux of P between these and more stable P pools.
5 Conclusion
This study showed that low water availability limited microbial activity and nutrient supply with high substrate supply, however, had little effect when substrate supply was low even at high water availability. This suggests that low soil water content will have a stronger negative impact on nutrient release in the detritusphere of nutrient-rich residues than of nutrient-poor residues.
References
Alamgir Md, Marschner P (2013) Changes in phosphorus pools in three soils upon addition of legume residues differing in carbon/phosphorus ratio. Soil Res 51(6):484
Alamgir M, McNeill A, Tang C, Marschner P (2012) Changes in soil P pools during legume residue decomposition. Soil Biol Biochem 49:70–77
Bolan NS, Naidu R, Mahimairaja S, Baskaran S (1994) Influence of low-molecular-weight organic acids on the solubilization of phosphates. Biol Fertil Soils 18:311–319
Brady N, Weil R (2002) Soil phosphorus and potassium the nature and properties of soils, 13th edn. Upper Saddle River, Prentice-Hall, Inc
Butterly CR, Marschner P, McNeill AM, Baldock JA (2010) Rewetting CO2 pulses in Australian agricultural soils and the influence of soil properties. Biol Fertil Soils 46:739–753
Christian JH, Waltho JA (1966) Water relations of Salmonella oranienburg; stimulation of respiration by amino acids. Microbiology 43:345–355
Clancy KM, Wagner MR, Reich PB (1995) Ecophysiology and insect herbivory. In: Ecophysiology of coniferous forests. Elsevier, pp 125–180
DeLuca TH et al (2015) A novel biologically-based approach to evaluating soil phosphorus availability across complex landscapes. Soil Biol Biochem 88:110–119
Díaz E, Roldán A (2000) Effects of reafforestation techniques on the nutrient content, photosynthetic rate and stomatal conductance of Pinus halepensis seedlings under semiarid conditions. Land Degrad Dev 11:475–486
Erich MS, Hoskins BR (2011) Effects of soil drying on soil pH and nutrient extractability. Commun Soil Sci Plant Anal 42:1167–1176
Erinle KO, Li J, Doolette A, Marschner P (2018) Soil phosphorus pools in the detritusphere of plant residues with different C/P ratio – influence of drying and rewetting. Biol Fertil Soils 54:841–852
Gaillard V, Chenu C, Recous S, Richard G (1999) Carbon, nitrogen and microbial gradients induced by plant residues decomposing in soil. Eur J Soil Sci 50:567–578
Ge G, Or D (2002) Particle size analysis. In: Dane J, Topp G (eds) Methods of soil analysis. Part 4. Physical methods. Soil Sci Soc Am J, Madison, pp 255–294
Geisseler D, Joergensen RG, Ludwig B (2012) Potential soil enzyme activities are decoupled from microbial activity in dry residue-amended soil. Pedobiologia 55:253–261
Ha K, Marschner P, Bünemann E, Smernik R (2007) Chemical changes and phosphorus release during decomposition of pea residues in soil. Soil Biol Biochem 39:2696–2699
Hanson WC (1950) The photometric determination of phosphorus in fertilizers using the phosphovanado-molybdate complex. J Sci Food Agric 1:172–173
Houghton JT, Ding Y, Griggs DJ, Noguer M (2001) Climate change 2001: the scientific basis. Contribution of working group I to the third assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, United Kingdom, New York
Hue NV, Ikawa H, Silva JA (1994) Increasing plant-available phosphorus in an Ultisol with a yard-waste compost. Commun Soil Sci Plant Anal 25:3291–3303
Kandeler E, Luxhøi J, Tscherko D, Magid J (1999) Xylanase, invertase and protease at the soil–litter interface of a loamy sand. Soil Biol Biochem 31:1171–1179
Khuyen TKH, Marschner P (2017) Plant and microbial-induced changes in P pools in soil amended with straw and inorganic P. J Soil Sci Plant Nutr 17:1088–1101
Konieczynski P, Wesolowski M (2007) Water extractable forms of nitrogen, phosphorus and iron in fruits and seeds of medicinal plants. Acta Pol Pharm 64:385–391
Kouno K, Tuchiya Y, Ando T (1995) Measurement of soil microbial biomass phosphorus by an anion exchange membrane method. Soil Biol Biochem 27:1353–1357
Liu M, Chen X, Chen S, Li H, Hu F (2011) Resource, biological community and soil functional stability dynamics at the soil–litter interface. Acta Ecol Sin 31:347–352
McKenzie H, Wallace HS (1954) The Kjeldahl determination of nitrogen: a critical study of digestion conditions-temperature, catalyst, and oxidizing agent. Aust J Chem 7:55–70
McLaren A, Skujins J (1968) The physical environment of microorganisms in soil, vol 3. Liverpool University Press Liverpool, Liverpool
Miller AE, Schimel JP, Meixner T, Sickman JO, Melack JM (2005) Episodic rewetting enhances carbon and nitrogen release from chaparral soils. Soil Biol Biochem 37:2195–2204
Miranda KM, Espey MG, Wink DA (2001) A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide 5:62–71
Moore JM, Klose S, Tabatabai MA (2000) Soil microbial biomass carbon and nitrogen as affected by cropping systems. Biol Fertil Soils 31:200–210
Mozumder B, Caroselli N (1970) Water relations of respiration of Verticillium alboatrum conidia. Phytopathology 60:915–916
Ohno T, Zibilske LM (1991) Determination of low concentrations of phosphorus in soil extracts using malachite green. Soil Sci Soc Am J 55:892–895
Poll C, Marhan S, Ingwersen J, Kandeler E (2008) Dynamics of litter carbon turnover and microbial abundance in a rye detritusphere. Soil Biol Biochem 40:1306–1321
Poll C, Brune T, Begerow D, Kandeler E (2010) Small-scale diversity and succession of fungi in the detritusphere of rye residues. Microb Ecol 59:130–140
Rayment GE, Higginson FR (1992) Australian laboratory handbook of soil and water chemical methods. Inkata Press Pty Ltd, Melbourne
Rencoret J, Gutiérrez A, Nieto L, Jiménez-Barbero J, Faulds CB, Kim H, Ralph J, Martínez ÁT, del Río JC (2011) Lignin composition and structure in young versus adult Eucalyptus globulus plants. Plant Physiol 155:667–682
Rovira A (1953) Use of the Warburg apparatus in soil metabolism studies. Nature 172:29–30
Schimel J, Balser TC, Wallenstein M (2007) Microbial stress-response physiology and its implications for ecosystem function. Ecology 88:1386–1394
Setia R, Marschner P, Baldock J, Chittleborough D, Smith P, Smith J (2011) Salinity effects on carbon mineralization in soils of varying texture. Soil Biol Biochem 43:1908–1916
Stewart CE, Moturi P, Follett RF, Halvorson AD (2015) Lignin biochemistry and soil N determine crop residue decomposition and soil priming. Biogeochemistry 124:335–351
Sun Q, Meyer WS, Koerber RK, Marschner P (2017) Prior rainfall pattern determines response of net ecosystem carbon exchange to a large rainfall event in a semi-arid woodland. Agric For Meteorol 247:112–119
Tecon R, Or D (2017) Biophysical processes supporting the diversity of microbial life in soil. FEMS Microbiol Rev 41:599–623
Turner BL, Driessen JP, Haygarth PM, Mckelvie ID (2003) Potential contribution of lysed bacterial cells to phosphorus solubilisation in two rewetted Australian pasture soils. Soil Biol Biochem 35:187–189
Umrit G, Friesen DK (1994) The effect of C:P ratio of plant residues added to soils of contrasting phosphate sorption capacities on P uptake by Panicum maximum (Jacq.). Plant Soil 158:275–285
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38
Wang X, Mohamed I, Xia Y, Chen F (2014) Effects of water and potassium stresses on potassium utilization efficiency of two cotton genotypes. J Soil Sci Plant Nutr 14:833–844
Wilke B-M (2005) Determination of chemical and physical soil properties. In: Monitoring and Assessing Soil Bioremediation. Springer, pp 47–95
Willis RB, Montgomery ME, Allen PR (1996) Improved method for manual, colorimetric determination of total Kjeldahl nitrogen using salicylate. J Agric Food Chem 44:1804–1807
Xu Y, Chen Z, Fontaine S, Wang W, Luo J, Fan J, Ding W (2017) Dominant effects of organic carbon chemistry on decomposition dynamics of crop residues in a Mollisol. Soil Biol Biochem 115:221–232
Xue R, Shen Y, Marschner P (2017) Soil water content during and after plant growth influence nutrient availability and microbial biomass. J Soil Sci Plant Nutr 17:702–715
Acknowledgements
Kehinde O. Erinle receives a postgraduate scholarship from the University of Adelaide.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Erinle, K.O., Marschner, P. Soil Water Availability Influences P Pools in the Detritusphere of Crop Residues with Different C/P Ratios. J Soil Sci Plant Nutr 19, 771–779 (2019). https://doi.org/10.1007/s42729-019-00076-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42729-019-00076-1