
Abstract
The fall armyworm has emerged as a major invasive pest of maize in multiple regions of Africa and Asia, affecting the crop at different stages. Pesticides have been widely used to manage fall armyworm in maize, however, there is always a need to explore other management options, including biological control. The current study aimed to explore the occurrence of the biological control agents in maize field, along with the predatory activity of selected groups under lab conditions. For live observations of the predatory arthropods in the field, weekly observations were made from five different maize fields located in Multan. For estimating predatory potential, selected groups of predators were collected using hand picking method and released in the Petri dishes under laboratory conditions. Diversity indices including the Shannon’s and Simpson’s diversity indices, showed significant differences in diversity of predators across various months and sites. The abundance of arthropod predators was significantly higher during September, October, March, and April. During fall 2022, spiders, red ants and lady bird beetles showed higher abundance followed by rove beetles and black ants. During spring 2023, spiders showed the highest mean abundance followed by red ants, rove beetles and lady bird beetles. The laboratory predation experiments showed lady bird beetles as an effective predator against the 1st larval instar of fall armyworm, followed by spiders and black ants, respectively. Results of the current study will help to identify the beneficial insects providing natural control of fall armyworm.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Maize, Zea mays L. (Poaceae), is an important food and feed crop across the globe (Luo et al. 2022). Among cereal crops in Pakistan, it is ranked forth after wheat, rice and cotton (Rehman et al. 2015). According to Pakistan Economic Survey 2022-23, maize showed 4.1% increase in cultivation area, i.e., 1720 thousand hectares and yield of 10.18 million tons. Overall, maize contributes 3% of total agricultural value and 0.7% of national GDP of Pakistan (Economic Survey of Pakistan 2022-23, 2023). Approximately 85% of maize has been utilized in livestock feed manufacturing and industrial processing. The maize germ has 45–50% oil and extracted oil has been used for cooking purposes (Shah et al. 2016). Maize also finds its utility in producing fermented drinks and leather bronzing chemicals (Watson 1988).
Lepidopteran pests including stem borers, cutworms, armyworms and ear borers inflict major losses to maize crop (Kotey et al. 2020). Among these pests, borers and fall armyworm (FAW) (Lepidoptera: Noctuidae) inflict major losses (Kumar 2002). FAW is a native to tropical and subtropical areas of the American continents and after 2016, invaded in different parts of Africa, Asia and Australia (Goergen et al. 2016), and in 2019, invaded in Pakistan (Naeem-Ullah et al. 2019). It is a highly migratory polyphagous pest with a host range of 353 plants from 76 families (Montezano et al. 2018). If unmanaged, maize yield may decrease up to 21–53% (Omwoyo 2021; Bhandari et al. 2020).
Natural enemies serve as an essential component of FAW pest management (Miller 1983; Abbas et al. 2022). In the USA (Hogg et al. 1982; Ashley 1986; Meagher et al. 2016), Mexico and Central America (Castro and Pitre 1989; Molina-Ochoa et al. 2003), and South America (Beserra et al. 2002; Murúa et al. 2009), surveys have been conducted to explore the natural enemies of fall armyworm. Multiple recent studies highlight the value of biological control to manage the fall armyworm in maize (Li et al. 2023; Durocher-Granger et al. 2023; Ratnakala et al. 2023; Gao et al. 2023; Mohamed et al. 2023, Fallet et al. 2022; Abbas et al. 2022; Perier et al. 2022; Kadzamira et al. 2022).
More than 21 arthropod predators have been reported to feed on at least one stage of fall armyworm (Khan 2021b). Most frequently reported predators include Solenopsis spp. (Hymenoptera: Formicidae) (Wyckhuys and O’Neil 2006), Orius insidiosus (Say) (Hemiptera: Anthocoridae) (Varella et al. 2015), and Doru spp. (Diptera: Forficulidae) (Toscano et al. 2012). A study reported formicids, forficulids, blattellids, pentatomids, and spiders as active predators and with formicids being most abundant in the early (55.85%) and late (95.67%) cropping seasons (Omoregie et al. 2023). Another study reported predators belonging to families: Coccinellidae (Coccinella septempunctata L.), Forficulidae [Forficula senegalensis (Serville)] and Formicidae [Pheidole megacephala (Fabricius) and Lasius niger (Linnaaeus)]. Out of these predators, the family Formicidae was most abundant (Adjaoke et al. 2023). Another study reported the diversity of predators belonging to different taxa: Carabidae (Calleida sp.), Coccinellidae [Cheilomenes sulphurea (Olivier)], Forficulidae [Diaperasticus erythrocephalus (Olivier), Forficula senegalensis (Serville)], Formicidae [Pheidole megacephala (Fabricius) and others], Mantidae, Pentatomidae and Reduviidae (Rhynocoris sp.). The most abundant predators were members of the Forficulidae, followed by Formicidae and Coccinellidae (Ahissou et al. 2021). Another work reported three predator species active against fall armyworm: Pheidole megacephala (F.) (Hymenoptera: Formicidae), Haematochares obscuripennis Stål, and Peprius nodulipes (Signoret) (both Heteroptera: Reduviidae) (Koffi et al. 2020).
A few studies have reported the predatory activity on fall armyworm, as most of the studies reports activity of parasitoids. Identification of the active predatory groups and integrating it into the pest management programs is essential (Gross and Pair 1986; Riggin et al. 1993; Figueiredo et al. 2006; Wyckhuys and O’Neil 2006–2007). Keeping in view this research gap in the maize growing area of Pakistan, the current study was performed to explore the occurrence of predatory arthropods in the maize fields. The study also evaluates the predation of selected predators against first instar of fall armyworm.
Materials and methods
Experiment 1: occurrence of Predatory arthropods in Maize Fields
Study site
We selected five maize fields in District Multan, Pakistan: Location 1: 30.125732, 71.441996; Location 2: 30.135324, 71.467226; Location 3: 30.120331, 71.469452; Location 4: 30.096679, 71.493568; and Location 5: 30.115108, 71.474317. The locations were located within the same city. The minimum distance between the two fields was 2 km. The maize fields were surrounded by wheat, cotton, and vegetables during our experiments.
Maize: sowing and management
The study was conducted from August to September 2022 and February to May 2023. All the observations were taken from the same fields in both seasons. However, the planted cultivar was different in both seasons. In fall 2022, maize hybrids, including YH-5482, FH-988, and YH-5427, were planted, and in spring 2023, YH-1898, FH-1046, and YH-5427 (AARI, Faisalabad, Pakistan) were planted. The smallholders managed all five fields. Before sowing, a deep cultivator was used to plough the fields, followed by a rotavator to prepare the difficult terrain, and finally, a bed shaper was used to make the ridges for sowing.
Sowing was completed in the last week of July 2022 and the first week of February 2023. Farmers sowed two to three seeds per ‘hill’ arranged in rows. Planting hills and rows were spaced apart on average ~ 20 cm and ~ 100 cm, respectively. Weekly visits to maize fields were made during the Fall 2022 and Spring 2023 growing seasons, which corresponded to different phenological phases of the crop. The ‘leaf collar method’ was used to determine the developmental stage of 50 randomly chosen plants at each visit (Nielsen 2004; Wyckhuys and O’Neil 2006).
Arthropod Sampling
Sampling was performed on weekly basis. We chose five transects of 10 plants each at random for each field visit, and we counted the number of arthropod predators found in each plant. To prevent edge effects, transects were placed inside the field. Plants were gently pressed to coerce the residents to exit maize whorls in order to count the number of earwigs and other arthropods living there. Unidentified predators were collected and transported to the lab for identification (Wyckhuys and O’Neil 2006).
Data analyses
The data were analyzed using the statistical software STATISTIX 8.1. ANOVA was performed, followed by the Tukey HSD All-Pairwise Comparisons Test to compare the means. Shannon’s diversity index (H), Shannon’s equitability (EH), Simpson’s diversity index (D), and Simpson’s equitability (ED) were calculated for predatory arthropods observed in maize fields during fall 2022 and spring 2023.
Experiment 2: laboratory evaluation of predation against fall armyworm
Collection of predators
Black ants, ladybird beetles, and spiders were collected from nearby maize fields using handpicking. All predators were identified by order level using a taxonomic key and used for lab predation trials. A few representative specimens were first washed with xylene and then preserved in 70% ethanol vials for further identification.
Rearing of fall armyworm
The fall armyworm larvae were identified by recognizing the pattern of black spots on the dorsal side of the larvae, as well as the inverted Y-shape on the top of their heads. After onsite identification, the collection was done from the maize field using the handpicking method. To prevent cannibalism, the larvae were kept in separate Petri dishes. The chopped maize leaves were also provided as food source. The Petri dishes, having fall armyworm larvae, were cleaned daily. A photoperiod of 16 L:8D hours. The temperature of 28 °C and relative humidity of 60 ± 10% were maintained in the laboratory. After pupation, the emerging adults were kept in the nylon mesh cage (0.028 m3). Adults were fed a 10% sugar solution. Potted maize plants were introduced into the cages, and eggs were laid on the leaves and cage walls. The eggs were harvested daily and wrapped in the white basal soft portion of the leaf. After hatching, the young caterpillars created small window-like openings on the leaves, which indicated the feeding progress. The first instar larvae were separated using a camel hair brush and deposited individually into Petri dishes. Fresh succulent leaves of maize were given after two days. After pupation, brown, reddish pupae were collected and transferred to other Petri dishes with a moist paper towel. After that, these Petri dishes were moved to the rearing cage with the potted maize plant and a 10% sugar solution. After ~ 9 days, adults emerged, and the whole process was repeated (Phambala et al. 2020).
Predation trial
Predators, including spiders, ladybird beetles, and black ants, were tested in a no-choice feeding trial against the 1st instar larvae of FAW. Fifteen 1st instar larvae, feeding on the maize leaves, were transferred to a single experimental unit, i.e., a Petri dish. After separating the fall armyworm larvae, a camel hair brush was used to release one 24-hour-old starved spider, lady bird beetle, and black ant in each of the experimental units. The experiment was replicated five times. The remaining larvae were counted, and a new batch of fifteen first-instar larvae were shifted to the experimental unit (Mohsin et al. 2015).
Results
Occurrence of predatory arthropods in maize fields
In fall 2022, significantly higher number of biological control agents were recorded in September and October 2022 (F = 71.4, df = 3, P < 0.001) (Fig. 1A). In spring 2023, significantly higher abundance recorded in March (F = 76.8, df = 3, P < 0.001) (Fig. 1B). The site wise abundance values fluctuated in both years. In fall 2022, the abundance of predatory arthropods was significantly higher on sites 1 and 4 compared to site 5 (Fig. 1C). In spring 2023, the abundance of predatory arthropods on sites 1–4 was significantly higher than on site 5 (Fig. 1D).

Mean (± SE) abundance of biocontrol agents across different months during (A) Fall 2022 (B) Spring 2023 and across different sites during (C) Fall 2022 (D) Spring 2023. Different letters above the bars are the significant differences (Tukey Kramer test, P < 0.05)
For the individual abundance of arthropods per 50 plants, significant differences were recorded during fall 2022 (F = 133.76, df = 16, P < 0.001) and spring 2023 (F = 159.16, df = 16, P < 0.001) (Table 1). In fall 2022, spiders, red ants and lady bird beetle were observed more as compared to other groups, while in spring 2023, spiders were abundant (Table 1).
The month wise distribution of arthropods during fall 2022 showed higher numbers of ants, lady bird beetles, rove beetles, and spiders in August. In September, red ants were detected significantly higher as compared to other groups. In October, red ants, spiders, and rove beetles were higher, while in November, spiders and lady bird beetles were abundant (Table 2).
The month wise distribution of arthropods during spring 2023 showed that lady bird beetles and spiders were significantly more abundant in February. In March, rove beetles and spiders were significantly higher in live observations. In April, red ants, spiders and rove beetles were significantly higher, while in May 2023, lady bird beetles, red ants and spiders were significantly higher in live observations (Table 3).
In fall 2022, site wise data of individual biological control agents also showed significant variations in the arthropods at different sites (Table 4). The overall abundance of the arthropods across different sites and different months also showed significant differences, with the highest abundance in September, across all sites (Table 5).
In spring 2023, site wise data of individual biological control agents also showed variations in the arthropods at different sites (Table 6). The overall abundance of the arthropods was significantly higher in March as compared to other spring months during the year 2023 (Table 7).
The Shannon’s (H) and Simpson’s diversity indices (D), and Shannon equitability index (EH) for live observations of biological control agents showed significantly higher values during September and October in fall 2022 and during March and April in spring 2023 (Table 8). However, Simpson’s equitability index (ED) was not significantly different during months of fall 2022 while showed higher values during the month of February during spring 2023 (Table 8). However, there were negligible differences among five different sites (Table 9).
Predation against fall armyworm
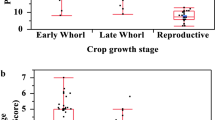
For the predation trial, there were significantly higher predation potential shown by lady bird beetles as compared to spiders and black ants, respectively (F = 59.0, df = 2, P < 0.001) (Fig. 2A). Overall, a higher predation potential was observed after 24 h and lowest was observed after 96 h of predatory activity (F = 21.34, df = 3, P < 0.001). A higher predation was observed after 24 h, while no significant difference in consumption rate were observed for 48 and 72 h followed by 96 h (Fig. 2B).

Mean (± SE) of consumption rate of predators (A) and mean consumption rates during different time periods (B). Different letters above the bars are the significant differences (Tukey Kramer test, P < 0.05)
For individual predatory potential by time, ladybird beetles showed higher predation potential after 24 h. Spiders performed significantly higher after 24 and 48 h as compared to later times. Black ants showed higher predation after 72 h (Table 10).
Discussion
Our research findings indicate a higher abundance of biological control agents in maize crops during the months of September and October, as well as in March and April. Spiders, ants, and predatory coleopterans showed the highest abundance in the field. An earlier study identified ants, spiders, and earwigs as abundant predators of the fall armyworm at the whorl stage of maize. Among these predators, earwigs were the most abundant, followed closely by ants (Wyckhuys and O’Neil 2006). Another study found 32 different predators and among those ants were the most abundant followed by the beetles and spiders (Litsinger et al. 2007). One study from Pakistan reported the abundance of big-eyed (Hemiptera) and ladybird beetles (Coleoptera) in maize fields well before the invasion of fall armyworm (Sahito et al. 2012).
Multiple studies have reported the biodiversity of beneficial arthropods associated with the fall armyworm. One study documented Formicidae, Forficulidae, Blattellidae, Pentatomidae, and Araneae, with formicids highly abundant (Omoregie et al. 2023). Similarly, formicids, coccinellids, and forficulids were found to be the most abundant predators (Ahissou et al. 2021; Adjaoke et al. 2023). El-Heneidy and Abbas (2005) found that in August, rove beetles were the most common predators in maize fields, while in September and October, the minute pirate bug and eleven-spotted lady beetles were the most abundant predators. Our findings also show a higher abundance of rove beetles in August, while bugs and lady beetles were higher in September and October months.
Ants, ladybird beetles and spiders showed predatory potential on fall armyworm larvae. In China, functional response experiments measuring the predatory potential of the insidious flower bug (Orius sauteri) and the Asian ladybeetle (Harmonia axyridis) showed high predatory potential of ladybird beetle against the eggs and larvae of fall armyworm (Di et al. 2021). Similarly, in turfgrass system, ants have been documented as the most active group preying upon the fall armyworm (Khan and Joseph 2022). Lady beetles have also been reported to consume immature stages of lepidopterans and coleopterans (Evans 2009). This indicates that the ladybird beetles have a potential to decrease the population of lepidopteran pests, like fall armyworm. Similarly, spiders have also been reported to manage the lepidopterans. Jeyaparvathi et al. (2013) did laboratory assays for estimating predatory potential of four spider species against lepidopteran and hemipteran pests, with green lynx spider Peucetia viridans (Hentz) (Araneae: Oxyopidae) showing the highest predation rate, based on daily feeding rates. These studies show the potential of these tested arthropod predators to be incorporated in the integrated pest management programs.
Spiders were the most abundant predator during both years. Previous studies have also highlighted the role of spiders in managing pests. Miliczky and Calkins (2002) found eleven spider species preying on caterpillars, with sac spiders consuming 65% during night time. Mohsin et al. (2015) study evaluated five spider species against major cotton pests. Results showed each predatory spider consumed at least one pest species, with higher predation rates due to unavailability of other feeding sources. A higher spider numbers, as found in the current study, may contribute to low pesticide usage and suggest their potential as effective predators. The current study also highlights the need for conservation efforts to increase the number of active predators, such as earwigs, which were reported to be low. Previously, the management strategy has been reported to influence the abundance of earwigs (Malagnoux et al. 2015). The lower number of earwigs may be attributed to their higher sensitivity to pesticide use. So, developing an integrated pest management program is crucial to maintain the diversity of beneficial arthropods.
Currently, pesticides dominate as the sole pest management option in Pakistan (Khan et al. 2023). The current evidence indicates the presence and activity of biological control agents in maize grown in southern Punjab, Pakistan, which could be utilized towards developing integrated pest management programs to manage the fall armyworm and other lepidopteran pests of maize (Khan et al., 2021; Khan et al. 2021c). Future studies should not only focus on testing the predation potential and functional response of different predator species against fall armyworm but also on the estimation of the predation under field conditions. Moreover, studies observing the effects of pesticides on the predators under field conditions are also needed. Future studies can also include the effects of climate change and farming practices on predatory species (Khan and Joseph 2024). It is crucial to develop conservation strategies for these predators, particularly in Pakistan where farmers mostly depend on the usage of pesticides. The central goal of the future studies should be to successfully incorporate predators into integrated pest management strategies.
Conclusion
The current study reports on the predatory fauna in relation to the fall armyworm in the maize system in Pakistan. Moreover, the study demonstrates the potential of ladybird beetles, spiders, and ants as effective predators.
Data availability
All the data associated with this research is in the manuscript.
References
Abbas A, Ullah F, Hafeez M, Han X, Dara MZ, Gul H, Zhao CR (2022) Biological control of fall armyworm, Spodoptera frugiperda. Agronomy 12:2704. https://doi.org/10.3390/agronomy12112704
Adjaoke AM, Yotto GT, Adandonon A (2023) Assessment of damage and biocontrol agents of fall armyworm Spodoptera frugiperda (JE Smith, 1797) (Lepidoptera, Noctuidae) on maize in southern of Benin republic. JAPS 33. https://www.thejaps.org.pk/
Ahissou BR, Sawadogo WM, Bonzi S, Baimey H, Somda I, Bokonon-Ganta A, Verheggen F (2021) Natural enemies of the fall armyworm Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) in Burkina Faso. https://doi.org/10.25518/2295-8010.1881. Tropicultura 39
Ashley TR (1986) Geographical distributions and parasitization levels for parasitoids of the fall armyworm, Spodoptera frugiperda. Fla Entomol 516:24. https://doi.org/10.2307/3495384
Beserra EB, Dias CT, Parra JR (2002) Distribution and natural parasitism of Spodoptera frugiperda (Lepidoptera: Noctuidae) eggs at different phenological stages of corn. Fla Entomol. https://doi.org/10.1653/0015-4040(2002)085[0588:DANPOS]2.0.CO;2 85:588– 93
Bhandari S, Bhattaraia R, Pandey KR, Adhikari S (2020) Assessment Of Infestation Of Spodoptera Frugiperda (JE Smith) On Maize And Its Implemented Management Practices With Their Efficacy In Kailali, Nepal. Malaysian J. Sustainable Agric. 5:10– 5. https://doi.org/10.26480/mjsa.01.2021.10.15
Castro MT, Pitre HN, Meckenstock DH (1989) Populations of fall armyworm, Spodoptera frugiperda (JE Smith), larvae and associated natural enemies in sorghum and maize cropping systems in southern Honduras. Trop Agric. https://journals.sta.uwi.edu/ojs/index.php/ta/article/view/1875
Di N, Zhang K, Xu Q, Zhang F, Harwood JD, Wang S, Desneux N (2021) Predatory ability of Harmonia axyridis (Coleoptera: Coccinellidae) and Orius sauteri (Hemiptera: Anthocoridae) for suppression of fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 12:1063. https://doi.org/10.3390/insects12121063
Durocher-Granger L, Fiorito S, Mudenda SK, Chiboola MM, Kansiime MK, Ludwig D, Leeuwis C (2023) Investigating the feasibility of developing a collective action for biological control of fall armyworm among smallholder farmers in rural communities of Zambia. CABI A&B 4:1–5. https://doi.org/10.1186/s43170-023-00154-6
Economic survey of Pakistan 2022–2023 (2023) https://www.finance.gov.pk/survey/chapters_23/02_Agriculture.pdf
El-Heneidy AH, Abbas MS (1984) Population dynamics of certain insect predators associated with aphids in maize fields in the Giza region. Beitr Trop Landwirtsch Veterinaermed 22:407–413. https://www.medscape.com/viewpublication/2942
Evans EW (2009) Lady beetles as predators of insects other than Hemiptera. Biol Control 51:255–267. https://doi.org/10.1016/j.biocontrol.2009.05.011
Fallet P, Bazagwira D, Guenat JM, Bustos-Segura C, Karangwa P, Mukundwa IP, Kajuga J, Degen T, Toepfer S, Turlings TC (2022) Laboratory and field trials reveal the potential of a gel formulation of entomopathogenic nematodes for the biological control of fall armyworm caterpillars (Spodoptera frugiperda). Biol Control 176:105086. https://doi.org/10.1016/j.biocontrol.2022.105086
Figueiredo MD, Martins-Dias AM, Cruz I (2006) Relationship between fall armyworm and their natural biological control agents in the maize crop. Pesquisa Agropecuária Brasileira 41:1693–1698. https://doi.org/10.1590/S0100-204X2006001200002
Goergen G, Kumar PL, Sankung SB, Togola A, Tamò M (2016) First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 11:e0165632. https://doi.org/10.1371/journal.pone.0165632
Gross HR Jr, Pair SD (1986) The fall armyworm: status and expectations of biological control with parasitoids and predators. Fla. Entomol. 502– 15. https://doi.org/10.2307/3495383
Hogg DB, Anderson RE, Pitre HN (1982) Early-season parasitization of fall armyworm (Lepidoptera: Noctuidae) larvae in Mississippi. Florida Entomol 65(4):584–585
Jeyaparvathi S, Baskaran S, Bakavathiappan GA (2013) Biological control potential of spiders on the selected cotton pests. Int J Pharm Life Sci. 4. https://www.ijplsjournal.com/home.html
Kadzamira MA, Chaudhary M, Williams F, Dutta NK (2022) A non-linear approach to the establishment of local biological control agent production units: a case study of fall armyworm in Bangladesh. CABI Agric biol sci 3:48. https://doi.org/10.1186/s43170-022-00115-5
Khan FZA (2021b) Understanding the Role of Predators for the Management of Fall Armyworm (Lepidoptera: Noctuidae) in Turfgrass (Doctoral dissertation, University of Georgia). https://esploro.libs.uga.edu/esploro/outputs/9949420529302959
Khan FZA, Joseph SV (2022) Assessment of predatory activity in residential lawns and sod farms. Biol Control 169:104885. https://doi.org/10.1016/j.biocontrol.2022.104885
Khan FZA, Joseph SV (2024) Influence of short-term, water-deprived bermudagrass on Orius insidiosus predation and Spodoptera frugiperda larval survival and development. Biocontrol Sci Technol, 1–14
Khan FZA, Manzoor SA, Gul HT, Ali M, Bashir MA, Akmal M, Haseeb M, Imran MU, Taqi M, Manzoor SA, Lukac M (2021a) Drivers of farmers’ intention to adopt integrated pest management: a case study of vegetable farmers in Pakistan. Ecosphere 10:e03812. https://doi.org/10.1002/ecs2.3812
Khan FZA, Manzoor SA, Akmal M, Imran MU, Taqi M, Manzoor SA, Lukac M, Gul HT, Joseph SV (2021c) Modeling pesticide use intention in Pakistani farmers using expanded versions of the theory of planned behavior. Hum Ecol Risk Ass: Int J 27:687–707
Khan FZA, Paudel S, Saeed S, Ali M, Hussain SB, Ranamukhaarachchi SL, Siddique M, Gireesh M, Sidhu GS, Nanayakkara D, Pandey S, Sharma T, Kaur P, Sharma M, Singh A, Epitakumbura LS, Thasneen S, Jayawardhana S, Kuruppuarachchi D, Ghimire S, Adhikari B, Karki B, Soti A, Mujtaba F, Imran MU, Haseeb M, Siddique F, Mehmood H, Manzoor SA (2023) Mitigating the impact of the invasive fall armyworm: evidence from south Asian farmers and policy recommendations. Int J Pest Manag 1–9. https://doi.org/10.1080/09670874.2023.2205834
Koffi D, Kyerematen R, Eziah VY, Agboka K, Adom M, Goergen G, Meagher RL (2020) Natural enemies of the fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) in Ghana. Fla Entomol 103:85–90. https://doi.org/10.1653/024.103.0414
Kotey DA, Siamey J, Egbadzor KF, Bissah MN (2020) Towards the identification of new sources of resistance to stem borer pests of maize in Ghana: preliminary findings and matters arising. Ghana J Forestry 36:22–33. https://www.ajol.info/index.php/gjf
Kumar H (2002) Plant damage and grain yield reduction by fall armyworm and stem borers on certain maize hybrids containing resistance genes from varying sources under experimental and farmers field conditions. Crop Prot 21:563–573. https://doi.org/10.1016/S0261-2194(01)00146-6
Li TH, de Freitas Bueno A, Desneux N, Zhang L, Wang Z, Dong H, Wang S, Zang LS (2023) Current status of the biological control of the fall armyworm Spodoptera frugiperda by egg parasitoids. J Pest Sci 18:1–9. https://doi.org/10.1007/s10340-023-01639-z
Litsinger JA, Dela Cruz CG, Canapi BL, Barrion AT (2007) Maize planting time and arthropod abundance in southern Mindanao, Philippines. II. Population dynamics of natural enemies. Int J Pest Manag 53:161–173. https://doi.org/10.1080/09670870701220176
Luo Y, Huang D, Miao H, Wu L, Zhu J (2022) Impact of advanced storage facilities on households’ maize storage losses and food security in China. Environ Dev Sustain 24:221–237. https://doi.org/10.1007/s10668-021-01406-z
Malagnoux L, Marliac G, Simon S, Rault M, Capowiez Y (2015) Management strategies in apple orchards influence earwig community. Chemosphere 124:156–162. https://doi.org/10.1016/j.chemosphere.2014.12.024
Meagher RL Jr, Nuessly GS, Nagoshi RN, Hay-Roe MM (2016) Parasitoids attacking fall armyworm (Lepidoptera: Noctuidae) in sweet corn habitats. Biol Control 95:66–72. https://doi.org/10.1016/j.biocontrol.2016.01.006
Miliczky ER, Calkins CO (2002) Spiders (Araneae) as potential predators of leafroller larvae and egg masses (Lepidoptera: Tortricidae) in central Washington apple and pear orchards. Pan-Pac Entomol 78:140–150. www.ars.usda.gov/research/publications/publication/?seqNo115=121837
Miller JC (1983) Ecological relationships among parasites and the practice of biological control. Environ Entomol 12:620–624. https://doi.org/10.1093/ee/12.3.620
Mohamed HO, El-Heneidy AH, Dahi HF (2023) Impact of egg mass layers and scale thicknesses of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), on the parasitic performance of Trichogrammatoidea bactrae (Hymenoptera: Trichogrammatidae). EJBPC 33:89. https://doi.org/10.1186/s41938-023-00733-9
Mohsin SB, Yi-jing L, Li-jie T, Maqsood I, Ma-sun T, Le-meng H, Khalil UR, Andleeb S, Muhammad SK, Saleem MA (2015) Predatory efficacy of cotton inhabiting spiders on Bemisia tabaci, Amrasca devastans, Thrips tabaci and Helicoverpa armigera in laboratory conditions. J Northeast Agric Univ (English Edition) 22:48–53. https://doi.org/10.1016/S1006-8104(16)30006-X
Molina-Ochoa J, Carpenter JE, Heinrichs EA, Foster JE (2003) Parasitoids and parasites of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas and Caribbean Basin: an inventory. Fla Entomol 86:254–289. https://doi.org/10.1653/0015-4040(2003)086[0254:PAPOSF]2.0.CO;2
Montezano DG, Sosa-Gómez DR, Specht A, Roque-Specht VF, Sousa-Silva JC, Paula-Moraes SD, Peterson JA, Hunt TE (2018) Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr Entomol 26:286–300. https://hdl.handle.net/10520/EJC-112bc26060
Murúa MG, Molina-Ochoa J, Fidalgo P (2009) Natural distribution of parasitoids of larvae of the fall armyworm, Spodoptera frugiperda, in Argentina. J Insect Sci 9(20). https://doi.org/10.1673/031.009.2001
Naeem-Ullah U, Ansari MA, Iqbal N, Saeed S (2019) First authentic report of Spodoptera frugiperda (JE Smith) (Noctuidae: Lepidoptera) an alien invasive species from Pakistan. Appl Sci Bus Econ 6:1–3. https://portal.issn.org/resource/issn/2312-9832
Nielsen RL (2014) Determining corn leaf stages. Corny News Netw Articles. 237.http://www.kingcorn.org/news/articles.03/LeafStaging-0520.html
Omoregie ME, Enobakhare DA, Emoefe JO, Omoregie AO (2023) Occurrence and Dynamics of Arthropod Predators Associated with fall armyworm, Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) on Maize in Benin City, Edo State. Nigeria J Appl Sci Environ Manag 27:1063–1067. https://doi.org/10.4314/jasem.v27i6.3
Omwoyo CO (2021) Assessment of Yield Losses and Farmers’ Perceptions on Control Practices of Fall Armyworm: the Case of Maize Infestation in Trans-nzoia County, Kenya (Doctoral dissertation, University of Nairobi). http://erepository.uonbi.ac.ke/handle/11295/155738
Perier JD, Haseeb M, Kanga LH, Meagher RL, Legaspi JC (2022) Intraguild interactions of three Biological Control agents of the fall Armyworm Spodoptera frugiperda (JE Smith) in Florida. Insects 13:815. https://doi.org/10.3390/insects13090815
Phambala K, Tembo Y, Kasambala T, Kabambe VH, Stevenson PC, Belmain SR (2020) Bioactivity of common pesticidal plants on fall armyworm larvae (Spodoptera frugiperda). Plants 9:112. https://doi.org/10.3390/plants9010112
Ratnakala B, Kalleshwaraswamy CM, Rajkumar M, Mallikarjuna HB (2023) Biocontrol potential of entomopathogenic nematodes against invasive fall armyworm, Spodoptera frugiperda in India. Biol Control 185:105304. https://doi.org/10.1016/j.biocontrol.2023.105304
Rehman A, Jingdong L, Shahzad B, Chandio AA, Hussain I, Nabi G, Iqbal MS (2015) Economic perspectives of major field crops of Pakistan: an empirical study. Pac Sci Rev b: Int j Humanit soc sci. https://doi.org/10.1016/j.psrb.2016.09.002. 1:145– 58
Riggin TM, Espelie KE, Wiseman BR, Isenhour DJ (1993) Distribution of fall armyworm (Lepidoptera: Noctuidae) parasitoids on five corn genotypes in South Georgia. Fla Entomol 292–302. https://doi.org/10.2307/3495729
Rouf Shah T, Prasad K, Kumar P (2016) Maize—A potential source of human nutrition and health: a review. Cogent Food Agric 2:1166995. https://doi.org/10.1080/23311932.2016.1166995
Sahito HA, Abro GH, Talpur MA, Mal B, Dhiloo KH (2012) Population fluctuation of insect pests and predators in maize, Zea mays L. Wudpecker J Agric Res 1:466–473. http://www.wudpeckerresearchjournals.org
Toscano LC, Calado Filho GC, Cardoso AM, Maruyama WI, Tomquelski GV (2012) Impacto De inseticidas sobre Spodoptera frugiperda (Lepidoptera, Noctuidae) e seus inimigos naturais em milho safrinha cultivado em Cassilândia E Chapadão do Sul, MS. Arq Inst Biol 79:223–231
Varella AC, Menezes-Netto AC, Alonso JD, Caixeta DF, Peterson RK, Fernandes OA (2015) Mortality dynamics of Spodoptera frugiperda (Lepidoptera: Noctuidae) immatures in maize. PLoS ONE 10:e0130437. https://doi.org/10.1371/journal.pone.0130437
Watson SA (1988) Corn marketing, processing, and utilization. Corn corn Improv 18:881–940. https://doi.org/10.2134/agronmonogr18.3ed.c15
Wyckhuys KA, O’Neil RJ (2006) Population dynamics of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) and associated arthropod natural enemies in Honduran subsistence maize. Crop Prot 25:1180–1190. https://doi.org/10.1016/j.cropro.2006.03.003
Wyckhuys KA, O’Neil RJ (2007) Influence of extra-field characteristics to abundance of key natural enemies of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in subsistence maize production. Int J Pest Manag 53:89–99. https://doi.org/10.1080/09670870701245207
Acknowledgements
The authors are thankful to the maize farmers who cooperated during the field trials in Multan, Pakistan.
Funding
The current research is self-funded.
Author information
Authors and Affiliations
Contributions
The authors SR and MI conceived and designed the study. SR and MSQZ performed the experiments. MI, FZAK and MAM did project supervision. FZAK and GA performed statistical analysis of the data. SR and FZAK wrote the draft of the manuscript. FZAK edited the final draft of the manuscript. All authors approved the final draft of the manuscript.
Corresponding author
Ethics declarations
Ethical approval
No need of ethical approval for this research work.
Conflict of interest
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Riaz, S., Ishtiaq, M., Khan, F.Z.A. et al. Occurrence of natural enemies in maize and the predatory potential of selected arthropods against fall armyworm in Multan, Pakistan. Int J Trop Insect Sci 44, 1297–1307 (2024). https://doi.org/10.1007/s42690-024-01227-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42690-024-01227-3