
Abstract
Altitudinal enclaves of rainforest are fragile and highly disturbed ecosystems located in high-altitude plateaus and mountains that are surrounded by Caatinga dry forest. Regardless of its biological importance, few studies regarding dung beetle assemblage were performed in an altitudinal enclave of rainforest. The aim of this study was to describe the structure of dung beetle assemblage from an altitudinal enclave of rainforest in the municipality of Triunfo, Pernambuco, Northeastern Brazil. Dung beetles were surveyed in the beginning of the rainy season using pitfall traps baited with excrement. Beetle diversity patterns were subsequently explored using Hill numbers. A total of 217 individuals belonging to 13 species and nine genera were recorded, and Dichotomius carbonarius (Mannerheim 1829), Uroxys bahianus Boucomont 1927, and Eurysternus nigrovirens Génier 2009 were the most abundant species. According to Hill numbers, there were five abundant species and three dominant species in the dung beetle assemblage. Also, there were two distinct beetle assemblages in the region of Triunfo, being most of the species commonly collected in Caatinga dry forest, as Deltochilum verruciferum Felsche 1911, Deltochilum sp., and U. bahianus. This study contributes to the scarce knowledge regarding diversity of dung beetles that inhabits altitudinal enclaves of rainforest of Northeastern Brazil, with a total of 43 species being currently recorded in such forests.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In the Neotropical region, Caatinga highlights as the largest of Seasonally Tropical Dry Forest (STDF), which is distributed mostly in Northeastern Brazil (Silva et al. 2018). Embedded at Caatinga, there are altitudinal enclaves of rainforest (AER), which are located in high-altitude plateaus and mountain ranges up to 1,000 m.a.s.l. (Queiroz et al. 2018). These small enclaves harbor distinct climatic conditions (e.g. high annual precipitation), which are very contrasting from those of the surrounding Caatinga (Andrade et al. 2018; Silva et al. 2019). Consequently, AERs harbor a distinct vegetation cover, structured as semi-deciduous and evergreen forests that can present typical elements of humid forests of South America, such as the Coastal Atlantic Forest and Amazonian forest (Queiroz et al. 2018).
The plant communities in AERs are assembled mainly by species from Amazonian and Atlantic forest ecosystems (Queiroz et al. 2018), which may suggest past connections between these two humid forests (Santos et al. 2007). Due to the contrasting climatic condition between Caatinga and AER, species composition from both ecosystems are strikingly distinct (Santos et al. 2007). High levels of species diversity and endemism are reported for the AERs scattered thought Caatinga domain (Pôrto et al. 2004). This pattern of biodiversity is frequently attributed to the mesic climatic conditions of AERs and the evolutionary history of ecological communities in this ecosystem (Smith et al. 2014; Dantas et al. 2015; Cabanne et al. 2016). Thus, AERs can be seen as refugia for rainforest taxa (Dantas et al. 2015; Cabanne et al. 2016). Unfortunately, the combination of mesic temperatures and elevated annual precipitation makes the AERs an attractive environment for agricultural purposes (Tabarelli and Santos 2004; Dantas et al. 2015), and only ca. 15% of their original forest are still conserved (Pôrto et al. 2004).
To investigate how habitat quality affects biodiversity, dung beetles feature as an effective indicator group, acting as a surrogate of local diversity (Nichols et al. 2007). Such beetles have ca. 6,200 species described worldwide (Tarasov and Dimitrov 2016), and ca. 720 species described in Brazil (Vaz-de-Mello 2020). To our knowledge, only three studies of dung beetle assemblage were performed in an AER (Silva et al. 2007; Alvarado et al. 2020; Barretto et al. 2021), which assessed the effect of seasonality, bait attractiveness, and habitat disturbance. Due to the fragility of this altitudinal ecosystem, it is important to understand the diversity patterns of its ecological communities, thus providing information regarding the biological importance of such reservoirs and giving basis for planning conservation strategies. In this context, the aim of this study was to describe the structure of dung beetle assemblage from an AER surrounded by Caatinga dry forest.
Material and methods
Study area
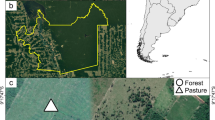
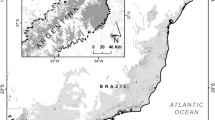
Surveys were conducted in January 2020 in Triunfo municipality, state of Pernambuco, Northeastern Brazil (38°05′W, 07°50′S; Fig. 1). This area can be characterized as a mosaic of native vegetation and crops (e.g. banana, mango and manioc). More specifically, the canopy height of the native vegetation reaches up to 17 m (Potapov et al. 2020), and the area of the sampling remnant was estimated in ca. 27 km2 (Foerster et al. 2019). Such native vegetation remnants are scattered throughout the geographical extent of the municipality of Triunfo, being often disposed in patches interspersed by a matrix of modified landscapes composed by both natural (e.g. large rock outcrops) and human-induced elements (e.g. croplands). Relief is mainly undulating and marked by expressive slopes, with soil types compounded by cambisols, latosols and litosols (Jacomine et al. 1973), being mostly shallow and blended with rocky conglomerates. The average annual temperature is 25 ± 13.7° C (mean ± standard deviation) and mean annual rainfall ranges from 634 to 1,190 mm (Karger et al. 2017). Vegetation structure of natural areas are classified as ‘brejo de altitude’ (see Andrade-Lima 1966) – a relictual AER immersed in Caatinga dry forest (Pôrto et al. 2004).

Location of study area located in Triunfo municipality, Pernambuco, Brazil
Dung beetle sampling
Surveys were performed at the beginning of the rainy season. To capture dung beetles, pitfall traps baited with ca. 50 g of human excrement were used. The pitfall trap consisted of a cylindric plastic jar (15 cm diameter × 13 cm height) with a bait-holding recipient installed ca. 3 cm above it. Besides, a solution of ca. 250 ml of a liquid mixture of water, detergent, and salt (ca. 1000 mL: 10 mL: 10 g) was placed inside the plastic container to conserve the captured dung beetles. We used exclusively human excrement since it attracts most of the dung beetle species in the Neotropics (e.g. Larsen et al. 2006), being therefore considered an efficient method to sample dung beetles. Furthermore, a study performed in AERs showed no clear difference on the dung beetle diversity between traps using excrement and other bait types (i.e. carrion) (Silva et al. 2007). Dung beetles were recovered 48 h after trap installation. Each sample consisted in a pair of pitfall traps, which were installed 5 m apart (each pair were displaced 50 m apart). A total of 14 traps were used in this study, comprising an area of ca. 5,500 m2 considering a radius of 25 m2 as the attractive area comprised by each trap set (see Larsen and Forsyth 2005, but see Silva and Hernández 2015). Specimens were stored at the Entomological Section of the Zoological Collection of the Federal University of Mato Grosso (CEMT, Brazil). Information regarding dung beetle diet type and dial activity period were obtained from literature (Aidar et al. 2000; Hernández 2007; Edmonds and Zidek 2010; Gillet et al. 2010; Lopes et al. 2011; Vieira and Silva 2012; Dinghi et al. 2013; Matavelli et al. 2013; Iannuzzi et al. 2016).
Data analysis
We assessed sampling coverage of the entire sampling extent, which was estimated according to Chao and Jost (2012). Sampling coverage was obtained using the iNEXT online software (Chao et al. 2016). Patterns of dung beetle diversity were explored by using Hill numbers (Jost 2006), which allowed us to quantify the alpha, beta, and gamma diversities. We used the diversity components (qD) of orders q = 0 (species richness), q = 1 (exponential of Shannon entropy), and q = 2 (inverse of Simpson diversity) (Hill 1973; Jost 2006). 0D is insensitive to species abundance, thus assigning a higher weight to rare species when compared to 1D, which considers the relative abundance of species (i.e. abundant species of the assemblages). 2D assigns a higher weight to abundant species than 0D and 1D, therefore indicating the number of dominant species of the assemblages (Jost 2006). We calculated diversity numbers using the entropart library in R software version 3.2.0 (Marcon and Hérault 2015; R Core Development Team 2015).
Results
A total of 217 individuals belonging to 13 species and nine genera were recorded (Table 1). Dichotomius carbonarius (Mannerheim 1829), Uroxys bahianus Boucomont 1927, and Eurysternus nigrovirens Génier 2009 were the most abundant species, accounting for 80.18% of the total abundance of dung beetles recorded in this study (Fig. 2). Six species were singleton: Canthon sp. 1, Canthon sp. 2, Deltochilum verruciferum Felsche 1911, Dichotomius nisus (Olivier 1789), Ontherus appendiculatus (Mannerheim 1829), and Onthophagus sp; Coprophanaeus cyanescens (d’Olsoufieff 1924) was doubleton. According to the sampling coverage estimator, 97.2% of the total expected number of species were obtained.

Species abundance rank of dung beetles collected at the rural region of Triunfo municipality, Pernambuco, Brazil
Diversity patterns at landscape level (gamma diversity) indicated approximately five abundant species (1Dγ = 4.82) and three dominant species (2Dγ = 3.28) in Triunfo region. According to diversity patterns per sample (alpha diversity), there were approximately five species per sample (0Dα = 5.55), two abundant species (1Dα = 2.58), and two dominant species (2Dα = 1.79). According to species turnover (beta diversity), there were two distinct communities based on species richness (0Dβ = 2.34), and nearly two distinct communities based on the exponential of Shannon (1Dβ = 1.86) and the inverse of Simpson (2Dβ = 1.83).
Discussion
Acting like a prominent biodiversity reservoir (Pôrto et al. 2014), the AERs scattered throughout Caatinga play a pivotal role on the maintenance of local biodiversity, since these remnants harbor several taxa adapted to both rainforests and dry environments. However, there are almost no studies encompassing dung beetle diversity in such altitudinal enclaves of tropical rainforests (except Silva et al. 2007; Alvarado et al. 2020; Barretto et al. 2021). Here we provide new data highlighting dung beetles species distribution and diversity patterns in the AER of Triunfo.
A total of 13 species were recorded in the municipality of Triunfo. From those, D. carbonarius was the most abundant species, encompassing more than half of the collected dung beetles. The AER of Triunfo was originally a closed-canopy forest, and currently it has been turned into a mosaic composed mostly by anthropogenic land-use, as pastures, agricultural crops, and urban settlements. Apparently, D. carbonarius thrives under open habitat conditions, as grasslands and native open canopy areas (see Louzada and Silva 2009; Tissiani et al. 2017). Therefore, we believe that the open-canopy structure that exists in Triunfo promote the widespread distribution of D. carbonarius in this region. In addition, it is important to note that the genus Dichotomius Hope, 1838 often comprises the dominant species in rainforests of Northeastern Brazil (i.e. Atlantic rainforest, see Filgueiras et al. 2015; Salomão et al. 2019). Since morphological, behavioral and physiological traits are related to the spatial distribution of dung beetles in tropical ecosystems (Nichols et al. 2007; França et al. 2016; Salomão et al. 2019), we suggest that further studies assess the relation between these traits and the dominance of Dichotomius species in tropical rainforests of Northeastern Brazil. By shedding light in this question, it is possible to reinforce the paradigm of traits related to dominant species in ecological communities.
Caatinga and AERs are contrasting ecosystems in which species from each region apparently does not move across them (Monteiro et al. 2016; Foerster et al. 2019). According to the rationale, in the AER studied herein we expected to record species that did not occur in Caatinga. However, we found two distinct assemblages with taxa from both Caatinga and Atlantic rainforest, and most of the species are commonly collected in Caatinga region. The species D. verruciferum, Deltochilum sp. (related to D. irroratum (Castelnau, 1840)), Onthophagus ranunculus Arrow, 1913, and U. bahianus have been recorded in some areas of Caatinga (Hernández 2007; Vieira and Silva 2012; Salomão and Iannuzzi 2017). It is important to note that dung beetle assemblages from open-canopy forests are highly distinct from those of closed-canopy forests (Andresen 2008; Filgueiras et al. 2015). Therefore, the conversion of closed forests (e.g. native rainforest) into open forests at Triunfo may have led to a replacement of dung beetle species that benefits from drier conditions promoted by the open vegetation. Accordingly, D. verruciferum, a dominant species in different regions of Caatinga (Hernández 2007; Salomão and Iannuzzi 2017), was rare in this study. Ecological communities from Caatinga are highly adapted to the dry conditions of this STDF ecosystem (Silva et al. 2018). Thus, the higher moisture and lower temperatures in AERs compared to Caatinga (Pôrto et al. 2004) may limit dung beetle species occupation to move across both ecosystems. Furthermore, the alarming deforestation levels observed in Triunfo region (Santos and Tabarelli 2004) may be an important factor leading to the high number of dung beetle species from Caatinga ecosystem.
In Triunfo we also recorded dung beetle species that are more commonly recorded in mesic than dry ecosystems, which is the case of E. nigrovirens (Genier 2009). Caatinga environments apparently act as an ecological barrier for Eurysternus Dalman, 1824 distribution. Dung beetles of this genus has an uncommon resource relocation strategy (i.e. dwellers) that consists in feeding and breeding in the resource patch, different from the other taxa (i.e. rollers and tunnellers), which relocate food material (Halffter and Edmonds 1982). We believe that the resource removal strategy presented by dwellers result on a higher exposure to climatic conditions when compared to roller and tunneller dung beetles, thus restraining Eurysternus distribution in harsh habitats. In Los Tuxtlas, Mexico, Eurysternus individuals were almost absent in the dung beetle assemblages analyzed right after the decrease of intense deforestation activities of this conservation unit (Salomão et al. 2020). Such result supports the idea of dweller sensitivity to harsh conditions. According to the assemblage structure observed herein, we may suggest that the anthropogenic disturbance regimes established in Triunfo led to an assemblage dominated mostly by Caatinga species. Considering the biological and biogeographic importance of altitudinal forests, future studies should focus on understanding the consequences concerning the conversion of forest into open areas for the ecological communities from AERs.
The AER of Triunfo showed the one of the lowest species richness when compared to the other two AERs that had their dung beetle diversity surveyed. Silva et al. (2007) recorded 28 species and 1,540 beetles at the AER of Brejo dos Cavalos, while Alvarado et al. (2020) recorded 22 species and 696 beetles at the AER of Areia. In a recent study, Barretto et al. (2021) studied the dung beetle diversity from six AERs, and diversity ranged widely among each AER (from 7 species and 27 beetles to 14 species and 492 beetles). There is a clear difference of sampling effort among the four studies: Silva et al. (2007) set a total of 240 traps at pasturelands near forest edges, distributed throughout 10 months; Alvarado et al. (2020) set a total of 60 traps (in a single month) at pasturelands, forest edge and forest core; Barretto et al. (2021) set 10 traps per AER (single sampling event), which were installed in forested sites. In this study, we set a total of 14 traps in a heterogeneous mosaic of habitats, installed during a one-month period. It is clear that with a higher sampling effort (p.e. a higher number of traps, bait types, sampling months, and area covered) we would attain a higher number of dung beetle species. However, due to logistic limitations, we were unable to expand the sampling effort used herein. Regarding the differences in species richness, it is important to consider the distances from each AER towards the nearest continuum of tropical rainforest (i.e. Atlantic rainforest). While our study area is in the middle of Caatinga dry forest, the AERs studied by Silva et al. (2007), Alvarado et al. (2020), and most of AERs studied by Barretto et al. (2021) are in the ecotone between Caatinga and Atlantic rainforest. According to theory of island biogeography (MacArthur and Wilson 1967) we could expect that AERs that are nearer from the ‘mainland’ (Atlantic rainforest) could sustain a higher species diversity than AERs that are more isolated. Future studies could focus on disentangling whether species richness in AERs follow the rules proposed by biogeography island theory.
In summary, we found that dung beetle diversity in Triunfo is comprised mostly by Caatinga species, with a few elements commonly found at rainforests, as E. nigrovirens. Although the dung beetle records from AER of Triunfo are far from complete, they are of significantly importance, as they reduce the large gap in AER knowledge regarding dung beetle fauna. This study contributes to the poorly known diversity of dung beetles that inhabits AERs of Northeastern Brazil, and a total of 43 species are currently recorded in such forests (Table 2). Altitudinal enclaves located in Caatinga are some of the relictual forests that previously represented a continuum from the tropical Atlantic and Amazonian rainforests (Pôrto et al. 2004; Santos et al. 2007). Due to the colonization history of AERs, which could be considered as an ‘oasis’ in the middle of a dry region, currently there are few well-preserved remnants. We encourage future studies to aim exploring the biodiversity of AERs, thus promoting a rational anthropogenic use of these ecosystems.
References
Aidar T, Koller WW, Rodrigues SR, Corrêa AM, da Silva JC, Balta ODS, Oliveira JM, Oliveira VLD (2000) Besouros coprófagos (Coleoptera: Scarabaeidae) coletados em Aquidauana, MS, Brasil. An Soc Entomol Bras 29:817–820
Alvarado F, Liberal CN, Dantas TAV, Bérgamo DB, Menezes RF (2020) Diversity and resource selection of dung beetles in a relictual mountain forest in Brazil. Int J Trop Insect Sci 2020: https://doi.org/10.1007/s42690-020-00327-0
Andrade EM, Aquino DN, Chaves LCG, Lopes FB (2018) Water as capital and its uses in the Caatinga. In: Leal IR, Tabarelli M (eds) Silva JMC. Caatinga, The Largest Tropical Dry Forest Region in South America. Springer Nature, pp 281–302
Andrade-Lima D (1966) Vegetação. In: Atlas Nacional do Brasil. Rio de Janeiro: IBGE, Conselho Nacional de Geografia.
Andresen E (2008) Dung beetle assemblages in primary forest and disturbed habitats in a tropical dry forest landscape in western Mexico. J Insect Conserv 12:639–650
Barretto J, Cunha JCS, da Silva F, Moura RC (2021) Dung beetle communities of altitudinal Atlantic forest remnants: diversity and composition Int J Trop Insect Sci 2021. https://doi.org/10.1007/s42690-021-00471-1
Cabanne GS, Calderón L, Trujillo Arias N, Flores P, Pessoa R, d’Horta FM, Miyaki CY (2016) Effects of Pleistocene climate changes on species ranges and evolutionary processes in the Neotropical Atlantic Forest. Biol J Linn Soc 119:856–872
Chao A, Jost L (2012) Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology 93:2533–2547
Chao A, Ma KH, Hsieh TC (2016) User’s Guide for iNEXT Online: Software for Interpolation and Extrapolation of Species Diversity. http://140.114.36.3/wordpress/wp-content/uploads/software/iNEXTOnline_UserGuide.pdf Accessed 1 Apr 20
Dantas LG, Esposito T, de Sousa ACB, Félix L, Amorim LL, Benko-Iseppon AM, Batalha-Amorim H, Pedrosa-Harand A (2015) Low genetic diversity and high differentiation among relict populations of the Neotropical gymnosperm Podocarpus sellowii (Klotz) in the Atlantic Forest. Genetica 143:21–30
Dinghi PA, Sánchez MV, Cantil LF, Sarzetti LC, Genise JF (2013) Leaf-litter brood chambers in Dichotomius (Luederwaldtinia) carbonarius (Mannerheim, 1829) (Coleoptera: Scarabaeidae): a novel behavior for dung beetles. Coleop Bull 67:388–396
Filgueiras BCK, Tabarelli M, Leal IR, Vaz-de-Mello FZ, Iannuzzi L (2015) Dung beetle persistence in human-modified landscapes: Combining indicator species with anthropogenic land use and fragmentation-related effects. Ecol Indic 55:65–73
Foerster SIA, DeSouza AM, Lira AFA (2019) Macroecological approach for scorpions (Arachnida, Scorpiones): β-diversity in Brazilian montane forests. Can J Zool 97:914–921
França F, Barlow J, Araújo B, Louzada J (2016) Does selective logging stress tropical forest invertebrates? Using fat stores to examine sublethal responses in dung beetles. Ecol Evol 6:8526–8533
Genier F (2009) Le genre Eurysternus Dalman, 1824 (Scarabaeidae: Scarabaeinae: Oniticellini), révision taxonomique et clés de détermination illustrées. Pensoft, Sofia
Hernández MIM (2007) Besouros escarabeíneos (Coleoptera: Scarabaeidae) da Caatinga paraibana, Brasil. Oecol Bras 11:356–364
Hill MO (1973) Diversity and evenness: a unifying notation and its consequences. Ecology 54:427–431
Karger DN, Conrad O, Böhner J, Kawohl T, Kreft H, Soria-Auza RW, Zimmermann NE, Linder HP, Kessler M (2017) Climatologies at high resolution for the earth’s land surface areas. Scientific Data 4:170122
Jacomine PKT, Cavalcanti AC, Burgos N, Pessoa SCP, Silveira CO (1973) Levantamento exploratório - Reconhecimento de solos do estado de Pernambuco. Boletim Técnico, SUDENE, Recife
Jost L (2006) Entropy and diversity. Oikos 113:363–375
Larsen TH, Forsyth A (2005) Trap spacing and transect design for dung beetle biodiversity studies. Biotropica 37:322–325
Larsen TH, Lopera A, Forsyth A (2006) Extreme trophic and habitat specialization by peruvian dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae). Coleopt Bull 60:315–324
Louzada JNC, Silva PRC (2009) Utilisation of introduced Brazilian pastures ecosystems by native dung beetles: diversity patterns and resource use. Insect Conserv Divers 2:45–52
MacArthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
Marcon E, Hérault B (2015) entropart: An R Package to Measure and Partition Diversity. J Stat Softw 67:1–26
Matavelli R, Campos AM, Silva FAB (2013) New records of Scarabaeinae (Coleoptera: Scarabaeidae) in a biogeographical transition zone in the state of Maranhão, Brazil. Check List 9:909–911
Monteiro LS, Garcia ACL, Oliveira GF, Rohde C (2016) High diversity of Drosophilidae in high-altitude wet forests in Northeastern Brazil. Neotrop Entomol 45:265–273
Nichols E, Larsen T, Spector S, Davis AL, Escobar F, Favila M, Vulinec K (2007) Global dung beetle response to tropical forest modification and fragmentation: A quantitative literature review and meta-analysis. Biol Conserv 137:1–19
Potapov P, Li X, Hernandez-Serna A, Tyukavina A, Hansen MC, Kommareddy A, Pickens A, Turubanova S, Tan H, Silva CE, Armston J, Dubayah R, Blair JB, Hofton M (2020) Mapping global forest canopy height through integration of GEDI and Landsat data. Environmen in press: 112165
Pôrto KC, Cabral JJP, Tabarelli M (2004) Brejos de Altitude em Pernambuco e Paraíba: História Natural e Conservação. Ministério do Meio Ambiente, Brasília
Queiroz LP, Cardoso D, Fernandes MF, Moro MF (2018) Diversity and Evolution of Flowering Plants of the Caatinga Domain. In: Leal IR, Tabarelli M (eds) da Silva JMC. Caatinga, The Largest Tropical Dry Forest Region in South America. Springer Nature, pp 23–64
R Core Development Team (2015) R: A language and environment for statistical computing. R foundation for statistical computing, Vienna
Salomão RP, Iannuzzi L (2017) How do regeneration stages of Caatinga forests influence the structure of dung beetle (Coleoptera: Scarabaeidae) assemblage? Coleop Bull 71:578–588
Salomão RP, Alvarado F, Baena-Díaz F, Favila ME, Iannuzzi L, Liberal CN, Santos BA, Vaz-de-Mello FZ, González-Tokman D (2019) Urbanization effects on dung beetle assemblages in a tropical city. Ecol Indic 103:665–675
Salomão RPS, Favila ME, González-Tokman D (2020) Spatial and temporal changes in the dung beetle diversity of a protected, but fragmented, landscape of the northernmost Neotropical rainforest. Ecol Indic 111:105968
Santos AMM, Tabarelli M (2004) Integridade, Esforço e Diretrizes para Conservação dos Brejos de Altitude da Paraíba e Pernambuco. In: Pôrto KC, Cabral JJP, Tabarelli M (eds) Brejos de Altitude em Pernambuco e Paraíba: História Natural e Conservação. Ministério do Meio Ambiente, Brasília, pp 309–318
Santos AMM, Cavalcanti DR, Silva JMCD, Tabarelli M (2007) Biogeographical relationships among tropical forests in north-eastern Brazil. J Biogeogr 34:437–446
Silva FAB, Hernández MIM, Ide S, Moura RC (2007) Copro-necrophagous scarab community (Coleoptera, Scarabaeidae) of the Brejo Novo region, Caruaru, Pernambuco, Brazil. Rev Bras Entomol 51:228–233
Silva FAB (2011) First record of Coprophanaeus bellicosus (Olivier) (Coleoptera, Scarabaeidae) in a “Brejo de Altitude” forest in northeastern Brazil: a historical biogeographical approach. Rev Bras Entomol 55:615–2617
Silva PG, Hernández MIM (2015) Spatial patterns of movement of dung beetle species in a tropical forest suggest a new trap spacing for dung beetle biodiversity studies. PlosOne 10:e0126112
Silva JMC, Leal IR, Tabarelli M (2018) Caatinga: the largest Tropical Dry Forest region in South America. Springer
Silva ISD, Vasconcellos A, Moura FMD (2019) Termite assemblages (Blattaria, Isoptera) in two montane forest (Brejo de Altitude) areas in northeastern Brazil Biota Neotrop 19. https://doi.org/10.1590/1676-0611-bn-2018-0519
Smith BT, McCormack JE, Cuervo AM, Hickerson MJ, Aleixo A, Cadena CD, Emán-Pérez J, Burney CW, Xie X, Harvey MG, Faircloth BC, Glenn TC, Derryberry EP, Prejean J, Fields S, Brumfield RT (2014) The drivers of tropical speciation. Nature 515:406–409
Tabarelli M, Santos AMM (2004) Uma breve descrição sobre a História Natural dos Brejos Nordestinos. In: Pôrto KC, Cabral JJP, Tabarelli M (eds) Brejos de Altitude em Pernambuco e Paraíba: História Natural e Conservação. Ministério do Meio Ambiente, Brasília, pp 17–24
Tarasov S, Dimitrov D (2016) Multigene phylogenetic analysis redefines dung beetles relationships and classification (Coleoptera: Scarabaeidae: Scarabaeinae). BMC Evol Biol 16:257
Tissiani ASO, Vaz-de-Mello FZ, Campelo-Júnior JH (2017) Dung beetles of Brazilian pastures and key to genera identification (Coleoptera: Scarabaeidae). Pesq Agropec Bras 52:401–418
Valois MC, Vaz-de-Mello FZ, Silva FAB (2017) Taxonomic revision of the Dichotomius sericeus (Harold, 1867) species group (Coleoptera: Scarabaeidae: Scarabaeinae). Zootaxa 4277:503–530
Vaz-de-Mello FZ (2020) Scarabaeinae in Catálogo Taxonômico da Fauna do Brasil. http://fauna.jbrj.gov.br/fauna/faunadobrasil/127498. Accessed 14 Apr 2020
Vieira L, Silva FAB (2012) Dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) of the Floresta Nacional Contendas do Sincorá, Bahia, Brazil. Check List 8:733–739
Acknowledgements
We thank ‘Coordenação de Aperfeiçoamento de Pessoal de Nível Superior’ (CAPES) for granting a postdoctoral (PNPD PPG-Eco-INPA) to RPS, and ‘Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco’ (FACEPE) for postdoctoral scholarship (BFP-0010-2.05/19) to AFAL. The authors thank Dr. Fernando Augusto Barbosa Silva for the taxonomic revision of some dung beetle species.
Author information
Authors and Affiliations
Contributions
RPS contributed to the study conception and design. Material preparation, data collection and analysis were performed by RPS, AFAL and SIAF. FNZ identified all collected specimen. All authors contributed equally to manuscript preparation and approved their final version.
Corresponding author
Ethics declarations
Competing interest
The field collection was complied with Brazilian law. Voucher specimens were deposited in the scientific collections of the Universidade Federal do Mato Grosso following standard procedures, and there are no conflicts of interest (financial and non-financial).
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Salomão, R.P., Lira, A.F.A., Foerster, S.I.A. et al. Dung beetle assemblage (Coleoptera: Scarabaeinae) from an altitudinal enclave of rainforest surrounded by a Seasonally Tropical Dry Forest in the Neotropics. Int J Trop Insect Sci 42, 55–62 (2022). https://doi.org/10.1007/s42690-021-00517-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42690-021-00517-4