
Abstract
The present study is an attempt to access the effect of two different silvicultural tree species on the process of nitrogen mineralization. Seasonal and depth wise variation in nitrogen mineralization of Populus deltoides (PAS) and Tectona grandis (TAS) based agrisilvicultural systems were studied using buried-bag technique. N-mineralization and nitrification rates differ significantly across the sites and seasons. Net N-mineralization rate varied from 5.88 ± 0.41 to 7.46 ± 1.44 µg g−1 month−1. Ammonium and nitrate concentration decreased with increasing soil depth at both the sites. The annual N-mineralization and nitrification rates were high in PAS than TAS system. The results suggested that variations in rate of N-mineralization and nitrification in the studied agrisilvicultural systems are related to differences in soil moisture content, nutrient status and vegetation cover in combination with other environmental factors.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen (N) is one of the key nutrients limiting plant growth in terrestrial ecosystems. Ammonium and nitrate are the main N forms assimilated by tree roots in temperate regions. Nitrification is one of the key ecological processes, since it converts less readily mobile ammonium into mobile nitrate that is easily leached out of the soil profile or absorbed in water by roots. Mineralization of organic N and immobilization of mineral N, two opposite processes take place simultaneously in soils, but at a given time the two processes are likely to be spatially partitioned into different microhabitats (Chen and Stark 2000; Myrold and Bottomley 2008). Within ecosystems, the mineralization of soil organic N plays an important role in the N cycling (Chapin et al. 2002). An understanding of N and other nutrient cycling processes are necessary to achieve the potential benefits and identify possible constraints associated with agroforestry systems (Nyadzi et al. 2003). The rate of soil N-mineralization and nitrification also indicate the capacity of soil to retain N, especially after disturbances (Haynes 1986).
Soil nutrients are closely related to land use types and their management (Wang et al. 2010). Land-use practices affect the distribution and supply of soil nutrients by directly altering soil properties and by influencing biological transformations in the rooting zone (Bouchoms et al. 2016; Bargali et al. 2018, 2019). Differences in human activities and vegetation have a strong influence on the changes in biological properties and stand structure (Manral et al. 2020). Plantations impart a favourable role in the biological reclamation due to modification of the soil characteristics. Different tree species are responsible for the variations in the quality of organic matter, which microbes receive from the leaf litter (Pastor and Post 1986). Soil microbes associated with various tree species often decompose the organic matter at different rate (Melillo et al. 1982), N-mineralization (Vitousek and Matson 1984; Zak and Pregiter 1990) and nitrification (Lovett and Rueth 1999).
Agroforestry is one of the best known traditional practices for livelihood, suitable land management and sustainable development (Kittur and Bargali 2013; Parihaar et al. 2014, 2015) and commonly practiced in the region to get the additional benefits of tree components. Trees based agroforestry systems (agrisilvicultural system), reduce the impact of climate change through carbon sequestration and boost the livelihood through the production of food, fodder and fuel wood (Arora et al. 2011; Singh et al. 2008; Bargali et al. 2009) and also increase the macro and micronutrients of soils (Sirohi and Bangarwa 2017).
Although the agroforestry system improves the soil quality but the quantitative data on N-mineralization in these systems of this region is limited. The objective of the present study was to analyze the seasonal and depth wise variations in N-mineralization and nitrification rates in soil under poplar (Populus deltoides) and teak (Tectona grandis) based agrisilviculture systems in Central Himalayan region of India. An attempt was made to quantify the influence of planted tree species on rates of N-mineralization and nitrification and to recommend the suitable plant species to grow in association with crops in agroforestry systems, so that an understanding of the mineralization rate could help to improve management practices.
Materials and methods
Study site
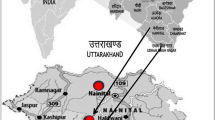
The two study sites were selected under the bhabhar region [an upland zone and foot of Himalayas with low water table having stones and rocky soil, made up of debris washed down from the hill ranges (Karki et al. 2021)] in Nainital district of Uttarakhand state in Indian Central Himalaya, situated between 29° 13′ and 29° 15′ N latitude and 79° 27′ and 79° 30′ E longitudes. The altitude ranges between 391 and 418 m above mean sea level (Fig. 1). Climatically the study area falls within sub-tropical zone with the year divisible into summer (April–mid June), rainy (late June–September) and winter (November–February) season. The mean monthly minimum temperature ranges between 9 and 26 °C and the mean monthly maximum between 22 and 37 °C. The mean monthly rainfall during study period was 0–475 mm (Fig. 2).

Location map of studied sites

Meteorological data of studied sites during study period (2016–2017)
Populus deltoides (PAS) and Tectona grandis (TAS) based agroforestry systems having similar age of stand (i.e., 8 years old tree plantation) were selected. The similar seasonal agricultural crops with the similar agricultural practices (i.e., Oryza sativa and Triticum aestivum) were grown in both the studied systems of the region. The standard tree density (Joshi et al. 1997) of poplar plantation was 833 trees ha−1 with 3 × 4 m spacing while density of teak plantation was 250 trees ha−1 with 5 × 8 m spacing.
Methodology
Soil samples were collected seasonally during the annual cycle from November, 2016 to October, 2017 in three depths (i.e., 0–20 cm, 20–40 cm and 40–60 cm) with the help of a soil corer in three replicates. Soil texture was examined by using sieves of different mesh size (Indian Standard 1965). Soil pH was analysed by digital pH meter (Jackson 1958). Water holding capacity (WHC) was estimated by the formula given by (Piper 1950). Bulk density was determined through the formula of Black (1965). Organic carbon was assessed by rapid titration method (Walkley and Black 1934). Total soil nitrogen was extracted by following (Subbiah and Asija 1956). The C/N ratios were derived from total soil organic carbon and total soil nitrogen contents (Jackson 1958).
N-mineralization
In the present study, net N-mineralization, nitrification and ammonification were determined seasonally following a short-term buried-bag technique (in field) incubation method (Eno 1960). In this method soil sample of the cores from each pair was sealed in sterile polyethylene bag after removing coarse roots and larger organic debris, to avoid nutrient immobilization during the incubation (Schimel and Parton 1986) and reinserted to its respective depth. The other fresh soil cores were brought to the laboratory to determine the ammonium and nitrate concentrations. After 1 month, the buried bags were retrieved and the soil samples were pooled according to depth and analyzed for final ammonium and nitrate concentrations. Changes in ammonium and nitrate concentrations were obtained by subtracting initial concentration from the corresponding final concentration, and the resultant values were referred to as ammonification and nitrification rates, respectively. Net N mineralization was calculated as the sum of changes in extractable ammonium (NH4+) N and nitrate (NO3−) N over 1 month. All the analyses were done in triplicate and the results are expressed on the basis of mean values of an oven-dry soil samples.
Statistical analysis
The data collected from different sites during the study were compiled and processed for statistical treatment using the Microsoft Excel. The replicates were analysed for the mean and standard error, while MANOVA by using SPSS software were done to prove the statistical significance of the results.
Results
Soil properties
The physico-chemical properties of soils of two investigated sites are presented in Table 1. The soil textures were clay loam in PAS site and loam in nature in TAS site (according to USDA classification). WHC, soil organic carbon and total nitrogen showed decreasing trend with increasing soil depth, while pH and C/N ratio showed a fluctuating trend. In contrast, bulk density increased with soil depth and ranged between 1.18 and 1.36 g cm−3. WHC of the soil was higher in PAS (32.61–42.79%) than TAS (31.24–35.06%) site. pH was alkaline in PAS (7.10–7.24) and acidic in TAS (6.62–6.95) site. At each depth SOC (0.93–1.88%) and total nitrogen (0.11–0.16%) were higher in PAS site as compared to TAS site (Table 1). C/N ratio showed fluctuating trend with soil depth and was higher in PAS site than TAS site (Table 1).
Mineral-N (NH4 +–N + NO3 −–N)
Present study showed that, extractable NH4+ was more than the extractable NO3− (Table 2). The value of ammonium and nitrate showed decreasing trend with increasing soil depth at both the investigated sites. Higher inorganic N pool (14.13 µg g−1) was recorded in PAS as compared to TAS (13.80 µg g−1).
N-mineralization rates
In PAS, ammonification rate varied from 4.74 µg g−1 month−1 (40–60 cm soil depth) to 5.58 µg g−1 month−1 (0–20 cm soil depth), and nitrification rate between 1.34 µg g−1 month−1 (40–60 cm soil depth) and 1.88 µg g−1 month−1 (0–20 cm soil depth). On the other hand in TAS, maximum ammonification rate (4.53 µg g−1 month−1) and nitrification rate (1.75 µg g−1 month−1) were observed at 0–20 cm soil depth and minimum ammonification rate (4.32 µg g−1 month−1) was observed in 20–40 cm soil depth. Net N-mineralization rate varied from 6.02 µg g−1 month−1 (40–60 cm soil depth) to 7.46 µg g−1 month−1 (0–20 cm soil depth) across the sites (Table 2). Ammonification rate varied significantly (p < 0.05) between the studied sites while season and depth showed insignificant effects.
Seasonal variation in ammonification and nitrification
In PAS, maximum ammonification rate was recorded during summer season while, in TAS it was highest during winter season. The minimum ammonification rate was observed during winter season in PAS site while, during summer season in TAS site. Nitrification rate was significantly (p < 0.05) correlated with the seasons. Highest rate of nitrification was encountered during the summer season whereas; it was lowest during the winter at both the studied sites. In PAS site, maximum N-mineralization was recorded during summer season followed by rainy and winter season. In TAS site, highest value of N-mineralization was observed during winter season followed by rainy and summer season (Fig. 3).

Seasonal variation in ammonification, nitrification and Net N-mineralization at two investigated sites
Discussion
Variation in soil properties
Soil is a complex system where chemical, physical and biochemical factors are held in dynamic equilibrium. Similar to many other soil properties, WHC too is affected by the organic matter content (Yuksek et al. 2009). Land use changes affect the WHC of soils because of changes produced in infiltration, surface runoff, and evaporation (Demir et al. 2007). Soil hydraulic properties were strongly influenced by land use management, vegetation type (Yuksek et al. 2009). In the present study, higher bD was observed in PAS site than the TAS site possibly due to high tree density at PAS site. The soil bulk density increased with the increase in the soil depth due to poor organic matter content at deeper depth.
The soil pH varied with soil depths at both the systems due to the differences in management practices such as application of manure and fertilizers in the field, crop rotation, ploughing, water management strategies and effect of tree species present in different agrisilviculture systems. Negative correlation was observed between SOC and bD in the present study which was in support of Sharma and Qahar (1989), Gupta and Sharma (2008) and Bhuyan et al. (2014) as they also reported a negative correlation between bD and SOC in Himalayan ecosystems. Variation in SOC under different agro-ecosystems may be due to the crop plant composition and the soil fertility management. Ploughing causes the breakdown of aggregates, may further increase the degradation processes by exposing organic material to biodegradation and oxidative agents (Padalia et al. 2018) and increase the available pore space (Bargali et al. 1993a) for microbial activities and root growth (Padalia et al. 2018).
Variation in ammonification and nitrification
The rate of ammonification was higher than the nitrification at both the sites. Similar results were also reported by Das et al. (1997) from the subtropical humid forest of northeast India, Tanjang et al. (2009) from the Areca catechu based agroforestry system of northeast India and Bhuyan et al. (2014) from the Paddy (AES) and homegarden system. The differences of plant species have long been recognized (Waksman and Tenney 1927) and the study of Aerts and Chapin (2000) stated that species composition is directly related to nitrogen pools and fluxes. Lower rate of nitrification than ammonification is helpful in conserving N loss through leaching (Das et al. 1997). Greater ammonium N than nitrate N concentrations in the present study also agree with Chao et al. (1993) who attributed it to the slightly acidic nature of the soil, which might have inhibited the growth and activity of autotrophic nitrifiers in the soil. Ammonification rate was higher in PAS (4.74–5.58 µg g−1 month−1) while, nitrification rate was higher in TAS (1.62 µg g−1 month−1) towards lower soil depth. This could be attributed to plant material (litter quality) because nitrogen mineralization is biologically mediated release of organically bound N from a substrate and its conversion to the inorganic form, ammonium and nitrate (Bargali et al 1993b, 2015; Bargali 1996).
Effect of soil depth on N-mineralization
In general, N-mineralization rates decreased with increasing soil depth Chen et al. (2005). In this study, at PAS site ammonification, nitrification and net N-mineralization rates decreased with increasing soil depth (Table 2). At TAS site, nitrification rate decreased with increasing soil depth, ammonification and N-minralization rate showed a fluctuating trend (Fig. 5). The higher percentage of N-mineralization at upper soil depth (0–20 cm) revealed that surface soil should be considered as the key part during study on N-mineralization though the differences between soil depths were not significant (Table 3). High N-mineralization rate tended to accumulate less N in the soil and it could therefore, be expected that there would be a more rapid depletion of labile soil N-pools. Between the sites, difference between ammonification and nitrification rate were more pronounced in upper layer and difference decreases with increasing soil depth (Fig. 5). The importance of root exudates in maintaining a larger microbial biomass closer to the trees has also been reported by Browaldh (1997). The rapid decline in microbial biomass with depth in the soil and distance from the trees in the present study could be attributed to the quantity and quality of organic inputs.
Effect of season on N-mineralization
Present study sites correspond to strong seasonality in N-mineralization and nitrification rates (Fig. 4). Nitrification and net N-mineralization were significantly affected by seasons (Table 3). As compared to TAS, seasonality was more pronounced in PAS (Fig. 4). In PAS maximum N-mineralization was recorded during summer season while, minimum N-mineralization was recorded during winter season. High concentration of mineral N during the dry summer season reflected low nutrient demand by vegetation and increase in supply due to microbial cell death (Singh and Kashyap 2007). The increased mineralization with increase in temperature is due to the increased microbial activities and decomposition of organic matter. In contrast, in TAS maximum N-mineralization was recorded during winter season and minimum N-mineralization was recorded during summer season (Fig. 4). Prieme and Christensen (2001) suggested that due to the death of microorganisms in winter they release nutrients and can facilitate the decomposition of organic detritus and mineralization of nitrogen.

Seasonal variations in Net N-mineralization in different soil depth at PAS (a) and TAS (b) sites
Difference in seasonal patterns of N-mineralization in PAS and TAS sites might be associated with the leaf fall pattern of planted species. Since site and season interaction was significant in the present investigation (Table 3), it may be suggested that the action of seasons across the study sites was different and may be related to topographic characteristic of each site, including vegetation cover, quality and quantity of soil organic matter, organic-N and or N-mineralization and nitrification rates.
Effect of Tree species on N-mineralization
This study indicated that the N-mineralization and nitrification rates differ across the two study sites (Fig. 5) though the differences were significant for ammonification only (Table 3). As recorded, PAS site performed better than TAS in terms of nitrogen mineralization. Annual average ammonification rate was 1.20 times and net mineralization rate was 1.13 times higher in PAS site while, nitrification rate was almost same in both the sites. Poplar based agro-forestry system prevent land degradation more efficiently by which biological production could be restored sustainably (Chauhan et al. 2010). It has been suggested that vegetation cover and associated factors greatly influence the rate of N-mineralization and nitrification (Singh and Kashyap 2007). Thus, high organic C in the soil of PAS site (1.33%) may be attributed to higher nitrogen mineralization and nitrification rates than TAS site (1.11%). Differences in ammonification, nitrification and net N-mineralization rates between two sites were more pronounced at uppermost (0–20 cm) soil layer and decreased with increasing depth (Fig. 4). This may be due to difference in quality and quantity of litter at two sites. In agrisilviculture land use system Rani et al. (2016) reported that leaf litter production was markedly higher for P. deltoides (7.8 t ha−1) as compared to T. grandis (1.83 t ha−1). They also reported that nitrogen concentration of P. deltoides (2.27%) leaf litter was significantly higher than T. grandis (1.45%) leaf litter.

a Ammonification, b nitrification, c net N-mineralization at two investigated sites
In PAS site, N-mineralization showed significant positive correlation with total soil nitrogen (Nmin = 3.015 + 28.32 TN; r2 = 0.972) as well as total inorganic nitrogen (Nmin = 1.07 + 0.35 IN; r2 = 0.915). In TAS though N-mineralization was positively correlated with total soil nitrogen as well as total inorganic nitrogen but the correlations were not significant.
Conclusions
We concluded that both the study sites had distinct rate of N-mineralization, possibly in response to distinct vegetation cover, soil moisture content and nutrient status. P. deltoides based agrisilvicultural system showed the better rate of N-mineralization as compared to the T. grandis based system, due to preferable soil conditions i.e., water holding capacity, organic C and N contents are higher in PAS system. These characteristics make the poplar tree as one of the ideal agroforestry species for improving soil fertility. However, teak based land-use system can also help in improving soil nutrient status. Therefore, tree based land-use systems must be promoted for better soil conditions in the bhabhar belt of Central Himalayan region.
References
Aerts R, Chapin FS (2000) The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Adv Ecol Res 30:1–67. https://doi.org/10.1016/S0065-2504(08)60016-1
Arora VPS, Bargali SS, Rawat JS (2011) Climate change: challenges, impacts, and role of biotechnology in mitigation and adaptation. Progress Agric 11:8–15
Bargali SS (1996) Weight loss and nitrogen release in decomposing wood litter in an age series of eucalypt plantation. Soil Biol Biochem 28:699–702
Bargali SS, Singh RP, Joshi M (1993a) Changes in soil characteristics in eucalypt plantations replacing natural broad leaved forests. J Veg Sci 4:25–28
Bargali SS, Singh SP, Singh RP (1993b) Pattern of weight loss and nutrient release in decomposing leaf litter in an age series of eucalypt plantations. Soil Biol Biochem 25:1731–1738
Bargali SS, Bargali K, Singh L, Ghosh L, Lakhera ML (2009) Acacia nilotica based traditional agroforestry system: effect on paddy crop and management. Curr Sci 96(4):581–587
Bargali SS, Shukla K, Singh L, Ghosh L, Lakhera ML (2015) Leaf litter decomposition and nutrient dynamics in four tree species of Dry Deciduous Forest. Trop Ecol 56(2):57–66
Bargali K, Manral V, Padalia K, Bargali SS, Upadhyay VP (2018) Effect of vegetation type and season on microbial biomass carbon in Central Himalayan forest soils, India. CATENA 171(12):125–135
Bargali SS, Padalia K, Bargali K (2019) Effects of tree fostering on soil health and microbial biomass under different land use systems in central Himalayas. Land Degrad Dev 30(16):1984–1998. https://doi.org/10.1002/ldr.3394
Bhuyan SI, Tripathi OP, Khan ML (2014) Effect of season, soil and land use pattern on soil N-mineralization, ammonification and nitrification: a study in Arunachal Pradesh, Eastern Himalaya. Int J Environ Sci 5(1):88–97
Black CA (1965) Methods of soil analysis. Academic Press Inc., New York, p 369
Bouchoms S, Wang ZG, Vanacker V, Doetterl S, Van Oost K (2016) Modelling longterm soil organic carbon dynamics under the impact of land cover change and soil redistribution. CATENA 151:63–73
Browaldh M (1997) Nitrogen release from leaves and prunings of different tree species used as green manure. Biol Agri Hortic 14(4):309–322
Chao WL, Gan KD, Chao CC (1993) Nitrification and nitrifying potential of tropical and subtropical soils. Biol Soil Fertil 15:87–90
Chapin FS, Matson PA, Mooney HA (2002) Terrestrial nutrient cycling. In: Chapin FS, Matson PA, Mooney HA (eds) Principles of terrestrial ecosystem ecology. Springer, New York, pp 197–223
Chauhan SK, Sharma SC, Beri V, Ritu Yadav S, Gupta N (2010) Yield and carbon sequestration potential of wheat (Triticum aestivum)-poplar (Populus deltoides) based agri-silvicultural system. Indian J Agric Sci 80(2):129–135
Chen J, Stark JM (2000) Plant species effects and carbon and nitrogen cycling in a sagebrush-crested wheatgrass soil. Soil Biol Biochem 32:47–57
Chen E, Zeng D, Singh AN, Chen G (2005) Effects of soil moisture and soil depth on nitrogen mineralization process under Mongolian pine plantations in Zhanggutai sandy land, PR China. J For Res 16(2):101–104
Das AK, Boral L, Tripathi RS, Pandey HN (1997) Nitrogen mineralization and microbial biomass N in a subtropical humid forest of Meghalaya, India. Soil Biol Biochem 29:1609–1612
Demir M, Makineci E, Yilmaz E (2007) Harvesting impact on herbaceous understory, forest floor and top soil properties on skid road in a beech (Fagus orientalis Lipsky) stand. J Environ Biol 28:427–432
Eno CF (1960) Nitrate production in the field by incubating soil in polythene bags. Soil Sci Soc Am Proc 24:277–279
Gupta MK, Sharma SD (2008) Effect of tree plantation on soil properties, profile morphology and productivity index I. Poplar in Uttarakhand. Ann For Sci 16(2):209–224
Haynes RJ (1986) Nitrification. In: Haynes RJ (ed) Mineral nitrogen in the plant soil system. Academic Press Inc, London, pp 127–165
Indian Standard (1965) Part IV: grain size analysis. Indian Standard Institute, New Delhi, p 2720
Jackson ML (1958) Soil chemical analysis. Prentice Hall, Inc., Englewood Clift
Joshi M, Bargali K, Bargali SS (1997) Changes in physico-chemical properties and metabolic activity of soil in popular plantations replacing natural broad leaved forests. J Arid Environ 35:161–169
Karki H, Bargali K, Bargali SS (2021) Spatial and seasonal pattern of fine root biomass and turnover rate in different land use systems in Central Himalaya, India. Russ J Ecol 52(1):36–48
Kittur B, Bargali SS (2013) Perspectives of agroforestry: ppresent and future facets. J Progress Agric 4(2):91–94
Lovett G, Rueth H (1999) Nitrogen Soil nitrogen transformations in beech and maple stands along a nitrogen deposition gradient. Ecol Appl 9:1330–1344
Manral V, Bargali K, Bargali SS, Shahi C (2020) Changes in soil biochemical properties following replacement of Banj oak forest with Chir pine in Central Himalaya, India. Ecol Process. https://doi.org/10.1186/s13717-020-00235-8
Melillo JM, Aber J, Muratore JF (1982) Nitrogen and lignin control of hardwood leaf litter decomposition dynamics, microbial biomass and functional diversity in a hilly area of the loess plateau, China. Ecology 63:621–626
Myrold DD, Bottomley PJ (2008) Nitrogen mineralization and immobilization. In: Raun W, Schepers JS (eds) Nitrogen in Agricultural Soils. American Society of Agronomy, Madison, pp 157–172
Nyadzi GI, Janssen BH, Otsyina RM, Booltink HWG, Ong CK, Oenema O (2003) Water and nitrogen dynamics in rotational woodlots of five tree species in western Tanzania. Agrofor Sys 59:215–229
Padalia K, Bargali SS, Bargali K, Khulbe K (2018) Microbial biomass carbon and nitrogen in relation to cropping systems in Central Himalaya. India Curr Sci 115(9):1741–1750
Parihaar RS, Bargali K, Bargali SS (2014) Diversity and uses of ethno-medicinal plants associated with traditional agroforestry systems in Kumaun Himalaya. Indian J Agric Sci 84(12):1470–1476
Parihaar RS, Kiran B, Bargali SS (2015) Status of an indigenous agroforestry system: a case study in Kumaun Himalaya. India Indian J Agric Sci 85(3):442–447
Pastor J, Post WM (1986) Influence of climate, soil moisture and succession on forest carbon and nitrogen cycles. Biogeochemistry 2:3–27
Piper CS (1950) Soil and plant analysis. The University of Adelaide, Academic Press, New York
Prieme A, Christensen S (2001) Natural perturbations, drying-welting and freezing thauring 322 cycles and the emission of nitrous oxide, carbon di oxide and methane from farmed organic soils. Soil Biol Biochem 33:2083–2091
Rani S, Benbi DK, Rajasekaran A, Chauhan SK (2016) Litterfall, decomposition and nutrient release patterns of different tree species in Taran district of Punjab. India J App Nat Sci 8(3):1260–1266
Schimel DS, Parton WJ (1986) Microclimatic controls of nitrogen mineralization and nitrification in short grass steppe soils. Plant Soil 93:347–357
Sharma PD, Qahar AQ (1989) Characterization of some outer Himalayan protected and eroded forest soils. J Int Soc Soil Sci 37:113–120
Singh JS, Kashyap AK (2007) Variations in soil N-mineralization and nitrification in seasonally dry tropical forest and savanna ecosystems in Vindhyan region, India. Trop Ecol 48(1):27–35
Singh L, Ghosh L, Bargali SS, Saxena RR (2008) Response of paddy planted under Butea monosperma trees in Chhattisgarh. Int J Ecol Environ Sci 34(1):63–66
Sirohi C, Bangarwa KS (2017) Effect of different spacings of poplar-based agroforestry system on soil chemical properties and nutrient status in Haryana, India. Curr Sci 113(07):1403
Subbiah BV, Asija GL (1956) A rapid procedure for the determination of available nitrogen in soil. Curr Sci 25:259–260
Tanjang S, Shukla AK, Arunachalam K, Arunachalam A (2009) Mineralization dynamics of nitrogen and phosphorus in Areca catechu L.-based traditional agroforestry system. Commun Soil Sci Plant Anal 40:3225–3237
Vitousek PM, Matson PA (1984) Mechanism of nitrogen retention in forest ecosystems: a field experiment. Science 225:51–52
Waksman SA, Tenney FG (1927) The composition of natural organic materials and their decomposition in the soil: III. The influence of nature of plant upon the rapidity of its decomposition. Soil Sci 24(5):317–334
Walkley A, Black IA (1934) An examination of Degtjareff method for determining soil organic matter and the proposed modification of the chromic acid titration method. Soil Sci 37(1):29–38
Wang L, Tang LL, Wang X (2010) Effects of alley crop planting on soil and nutrient losses in the citrusorchards of the Three Gorges Region. Soil Till Res 110(2):243–250
Yuksek T, Gol C, Yuksek F, Yuksel EE (2009) The effects of land-use changes on soil properties: the conversion of alder coppice to tea plantations in the Humid Northern Black sea Region. Afr J Agric Res 4(7):665–674
Zak DR, Pregiter KS (1990) Spatial and temporal variability of nitrogen cycling in North lower Michigan. For Sci 36:367–380
Acknowledgements
Authors are thankful to the Head, Department of Botany for providing necessary lab facilities and Uttarakhand Tea Development Board Bhowali, Nainital for analysis of soil samples. No financial support was given by any agency. We are thankful to both the reviewers and the editor for their closer and critical look in to the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Authors have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Karki, H., Bargali, K. & Bargali, S.S. Nitrogen mineralization patterns in Populus deltoides and Tectona grandis based agrisilvicultural practices in Central Himalaya, India. Vegetos 34, 86–93 (2021). https://doi.org/10.1007/s42535-021-00195-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42535-021-00195-0