
Abstract
Faba bean seeds are generally large which limits its adoption as cover crop and/or dual-purpose cover crop/cash crop due to the high seed cost. The purpose of this study was to evaluate six faba bean varieties for their yield and yield components, using phenology, morphology and physiological growth pattern. Faba bean varieties included Early Violletto, Aquadulce, Delle Cascine, Windsor, Early White, and D’Aquadulce, which studied in 2015 and 2016. Aquadulce produced the highest grain yield followed by Delle Casine. These two varieties also produced the highest biomass, which potentially can provide more N to the succeeding crop. Aquadulce and Delle Cascine had the highest HI (40.1 and 41%, respectively) indicating an efficient distribution of assimilates to their seeds.. Delle Cascine produced the highest number of seeds plant−1 and seed weight, respectively. Plant height variation also revealed that varieties Early White was the shortest (40.6 cm) and the tallest (76.0 cm), respectively. Significant differences were observed in regard to the growing degree days required to reach flowering and pod formation stages. Aquadulce and Delle Cascine ranked early among the tested varieties, providing the opportunity for dual purpose and double cropping in short-season areas.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Faba bean is a cool-season grain legume crop with high nutrition value. Recently more attention has been made on using faba bean as a multi-purpose legume crop (L’opez-Bellido et al. 2005; Etemadi et al. 2015; Landry et al. 2015) and/or in intercropping systems (Zhang et al. 2004; Song et al. 2007; Li et al. 2009). Faba beans fit nicely into various crop rotations, including double cropping with other vegetables and grains (Turk and Tawaha 2002; Agegnehu et al. 2006). Due to the diverse and significant ecological services (Köpke and Nemecek 2010) faba bean has increasingly received attention. Currently, faba bean is widely cultivated in many regions of the world as a source of food and as a break crop in cereal production to minimize the occurrence of cereal cyst nematode (Heterdera avenae) (Sattell et al. 1998) and some soil-borne pathogens (Landry et al. 2016).
In legumes, the total biomass production plays an additional role in sustainability of cropping systems (Zandvakili et al. 2012; Etemadi et al. 2018). Arguably, the most important contribution of faba bean to agricultural ecosystems is its high capability of fixing atmospheric nitrogen (Cline and Silvernail 2002; Hoffmann et al. 2007; Horst et al. 2007; Ruisi et al. 2017; Etemadi et al. 2018). It is well documented that the magnitude of N contribution to the soil nitrogen largely depends on the amount of final biomass of the legume crop (Finney et al. 2016; Jahanzad et al. 2016; Mirsky et al. 2017).
Integrating faba bean into the crop rotation systems requires some information about the growth pattern and morphological characteristics in addition to the potential yield of various genotypes (Agegnehu et al. 2006). Etemadi et al. (2015) recommended that faba bean should be sown in spring as early as late March to provide the opportunity for successful double cropping in areas with relatively short growing season. In cold areas, faba bean can also be grown in summer by mid-July as dual-purpose cash/cover crop (Etemadi et al. 2015; Landry et al. 2015). Domestication of faba bean began approximately 4000 years ago through selection among wild ecotypes for larger seeds and plants. The selection for larger plants resulted in higher potential ecological benefits including N fixation (Köpke and Nemecek 2010). The ability of faba bean to produce large quantity of biological N fixation is well documented (Unkovich and Pate 2000; Jensen et al. 2010; Dayoub et al. 2017; Denton et al. 2017; Barłóg et al. 2018; Gebremariam and Assefa 2018). Therefore, faba bean can significantly contribute to the soil N that lowers the need for the N by the subsequent cash crop (Etemadi et al. 2018).
Faba bean is a new crop in New England and Windsor is currently the dominant variety available to the growers however, Windsor has not performed well in our previous studies and out-yielded by several other varieties (Etemadi et al. 2017). Thus evaluating more varieties for their potential yield and suitable morphological characteristics can provide valuable information to the researchers and growers. Seed yield is a function of DM accumulation and partitioned assimilates into the seeds (Ricciardi et al. 2001). Phenology is the study of the timing of biological events, the cause of their timing with regards to biotic and abiotic forces, and the interrelation among phases of the same or different species (Hunt and Yamada 2003). Phenological development described as the rate of progress through growth stages. Temperature and photoperiod can control phonological growth stages, more specifically the reproductive growth (Peng et al. 2004; Wilczek et al. 2010). Therefore, variation in time to reach each phenotypic stage relates to the sensitivity of genotypes to changes in the photo-thermal regimes thus affecting days to maturity, adaptation to the environment, and yield in divers growing conditions (Peng et al. 2004; Etemadi and Hashemi 2012). Different types of faba beans including spring, winter and Mediterranean exhibit various phenological characteristics therefore induction and maintenance of flowering differs among these classes (Patrick and Stoddard 2010).
Plant breeders have recognized the importance of larger seed in the production of food crops and have been breeding crops for the trait. General conception is that the larger seeds may have more food reserves thus germinate faster, establishes more successfully and out yield smaller seeds within the same variety/cultivar (Gan et al. 2004). The desired population density traditionally determined by seeding rate, which primarily relate to the number of seeds per unit weight (kg). Therefore, seeding rate of cultivars with smaller seed size can be significantly lower than larger seeds. Since the cost of faba bean seeds is relatively high, smaller seeds with high yield potential are preferable and can significantly reduce the production costs.
Despite the significant economic and ecological importance of faba bean, the relationship between various phenology, morphology, and growth pattern characteristics of various faba bean varieties in short growing season regions is not extensively investigated.
The present study aimed to assess selected traits that are important for adaptation of faba bean in short-season areas. The phenology, morphology, and seed yield of six faba bean varieties studied in a 2-year field experiment.
Materials and Methods
Experimental Site and Weather Condition
A 2-year field experiment was conducted in 2015 and 2016 at the University of Massachusetts Amherst Agricultural Experiment Station, Crops and Animal Research and Education Farm in South Deerfield (42°28′37″N, 72°36′2″W). Weather condition including growing degree-days (GDD) and precipitation throughout the experiment period in both years and the norm (average of 20 years) for the region presented in Table 1. The soil type at the experimental site was a Hadley fine sandy loam (nonacid, mesic TypicUdifluvent). Composite soil sample (0–0.2 m depth) taken prior to planting indicated soil pH (1:1, soil/H2O) was 6.6, cation exchange capacity (CEC) was 8.1 meq/100 g and available P, K and Mg were all in the optimum range thus no fertilizer was applied to the experimental plots in both years. Throughout the experiment, weeds periodically removed by hand when necessary. No irrigation used in this experiment, as irrigation is not a common practice for agronomic field crops in Massachusetts.
Experiment Layout and Measurements
Six faba beans varieties including; Aquadulce, Early Violletto, Delle Cascine, Early White, D’Aquadulce, and Windsor, were hand seeded on April 16 in 2015 and April 14 in 2016. A complete randomized block design experiment with four replications used in this study. Each plot consisted of three rows, 76 cm apart and 15 cm space between the plants, which is equivalent to a population of 60,000 plants ha−1. Morphological characteristics including height of plant, number of lateral branches per plant, height of the first pod from the ground measured on five randomly tagged faba bean plants in each plot. Plant height was determined by measuring the distance between the ground and the terminal growing point. Number of lateral branches counted at the harvest. First pod height was determined by measuring the distance between the soil surface and the first pod presence in the plant before the harvest. The major phenological stages of the varieties were determined when 50% + 1 of plants in the center row reached the corresponding stage. The recorded phenological stages included the days required to reach three leaf stage, flowering, first pod formation, and pod maturity. The number of Growing Degree Days (GDD) accumulated for reaching each phenological stage was calculated using Eq. 1:
where Tmax and Tmin are the maximum and the minimum daily air temperature (°C), Tbase is the base temperature equal to 4 °C.
Crop Growth Rate (CGR) was calculated (Eq. 2) for entire growth period (Aghaalikhani et al. 2012). At each harvest, five plants from first and third rows of each plot cut at the soil surface and oven dried at 50 °C for 72 h in a forced-air oven to reach a constant weight.
Seed yield (kg ha−1) and seed size (100-seed weight) were determined based on harvesting pods from 2 m2 from the middle row. Seed yield is corresponding to the total seed weight of multiple harvests and seed size is the average weight of multiple groups of 100 seeds. At maturity, all plants from the 5 m2 of the center row harvested manually by cutting at the soil surface for determination of biological yield. Harvest index (HI) derived as the proportion of grain yield to total biological yield.
Statistical Analysis
Data subjected to analysis of variance using Proc GLM procedure of SAS software (SAS Institute 2003). Data shown in figures are the arithmetic means of four replicates of each treatment. Effects considered significant at P ≤ 0.05 by the F test, and when the F test was significant, Fisher’s Least Significant Difference Test (LSD) was used for mean separations. Regression equations (averaged of 2 years) for faba bean CGR based on days after planting was determined by fitting the data to a quadratic trend using SAS linear regression procedure (PROC REG). The relationships between the traits analyzed using PROC CORR of SAS software.
Results and Discussion
Weather
Cumulative growing degree days (GDD 4 °C) at Orange Airport, MA (roughly 27 km away from the research site), during the growth period of faba bean (April–July), were 2484 and 2461 in 2015 and 2016, respectively which were similar to the norm for this location (Table 1). Since irrigation is not a common practice for field crops production in Northeast U.S., plants are depend on the amount of precipitation they receive during the growing season. In 2016, faba bean plants received almost 50% less precipitation than in 2015 during April through July, the months that spanned the crop growth period. More specifically, lower moisture availability in the month of June 2016 when pods and seeds are normally formed, resulted in 28 and 29% reduction in biomass and seed yield, respectively when averaged across all faba bean varieties (Table 3).
Phenology
The life cycle of grain legumes identifies by several major phenological stages. These stages include seed emergence, vegetative growth, flowering, pod set, pod filling, and physiological maturity. In faba bean, the induction of flowering, retention of flowers and development of seeds are all therefore critical processes in final yield determination.
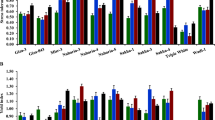
Figure 1 illustrates the mean thermal units (above 4 °C) of the varieties that needed to reach different phonological stages. While the varieties did not show significant differences in required GDD to reach their 3-leaf stage, large variations recorded for other stages of growth among the varieties. Aquadulce and Delle Cascine required 534 GDD to produce their first flower, which was almost 25% lower than the Early White, which flowered last. Interestingly, the flowering period of Aquadulce and Delle Cascine was longer than other varieties except in Windsor. Aquadulce matured earlier than all other varieties and required 33% less GDD than the second earliest variety, Delle Cascine. Considering that faba bean is sensitive to high temperature and low moisture availability, days to flowering and maturity is very important (L’opez-Bellido et al. 2005). Earlier maturity potentially considered as a positive characteristic for the varieties grown in short growing season areas such as New England. This is because firstly, the plant will not coincide with high temperature in summer, which makes faba bean plants sensitive to chocolate spot bacteria. Secondly, earlier harvest provides the opportunity for successful double cropping which is crucial to the growers’ income. However, early maturity varieties must demonstrate an efficient distribution of assimilates to the economic sinks to produce high pods/seeds yield. Other reports indicated that faba bean varieties exhibited large variations in terms of maturity (Tafere et al. 2012; Talal and Munqez 2013; Mitiku and Wolde 2015). Our results however, does not confirm the earlier report by Girma and Haila (2014) who concluded that physiological maturity of faba bean varieties was not different in irrigated and rain fed conditions.

Growing degree days required to reach various growth stages of six faba bean varieties
Morphology and Growth
Growing season had a significant influence on all measured morphological and growth indices (Table 2). As mentioned earlier, the amount and distribution of precipitation was more suitable in 2015 than 2016 (Table 1). As a result, average all varieties, plant height, height of first pod from the soil surface, number of lateral branches, and maximum CGR were higher in 2015 compared with 2016. However, the interaction of year and variety was not statistically significant therefore; an average of the 2 years of experiment presented in Table 2.
Varieties used in this experiment showed significant differences in terms of plant height, height of first pod from the soil surface, and number of branches (Table 2). Windsor was the tallest variety followed by Aquadulce, while the Early White was the shortest variety. Except in Early Violletto, the height of the first pod from the ground ranged from 15 to 19 cm. The height of the first pod is important for the ease of fresh pods hand harvesting and for mechanical harvesting of dry seeds. Moreover, the lowest pods may contaminate with soil particles due to the splash of raindrops. Number of lateral branches per plant varied between 2–4 and the difference between the varieties was significant. Number of branches is mainly influenced by agronomic management practices including plant density (Loss et al. 1998a, b; Gezahegn et al. 2016), time of planting (Badran and Ahmed 2010) and environment conditions such as moisture availability (Toker 2004; Hegab et al. 2014). Number of lateral branches per plant can also differ based on the differences in the genotype (Mwanamwenge et al. 1998; Sozen and Karadavut 2016). In the current experiment, since a uniform population density and environmental condition existed, the difference in the number of branches primarily related to the genotypic differences rather than agronomic practices and/or environmental conditions. In general, producing several branches per plant should not be considered as a good characteristic since lateral branches are not as productive as the main stem. Therefore optimum canopy closure should be reached through increasing population density rather than the capacity of varieties for branching capability.
The maximum CGRs of the varieties tested in this study were considerably different and varied between 2.7 g m−2 day−1 in low yielding variety, D’Aquadulce, and 6.6 g m−2 day−1 in high producing varieties, Delle Cascine and Aquadulce (Table 2). The highest value obtained in this study was significantly lower than the maximum CGR recently reported by Barłóg et al. (2018). The lower maximum CGR in our study could be attributed in part to the growing conditions in the Northeast U.S., which usually has more cloudy days, and due to the lack of irrigation practice. The time to reach the peak CGR was also significantly different among the varieties and ranged from 62 days in Aquadulce to 75 days in Windsor. Intensive growth period of faba bean occurs during flowering stage, when the crop reaches its highest leaf area index (LAI) and CO2 fixation rate (Coelho and Pinto 1989; Pilbeam et al. 1991). Our results indicated that the days to reach the maximum CGR in all varieties, regardless of their maturity, coincided during the flowering period. Interestingly it turned out that the earliest (Aquadulce) and latest (Windsor) maturity varieties required the fewest and the highest days to reach their maximum CGR, respectively (Fig. 1). As the time passed, dry matter accumulation rate decreased with the aging of the leaves, thus CGR reduced at final stages of growth.
Biomass, Seed Yield, and Harvest Index
Biomass yield was significantly different in the two growing seasons and averaged all varieties was 28% higher in 2015. Biomass also varied greatly among the faba bean varieties and ranged from 2162 kg ha−1 in D’Aquadulce in 2016 up to 7970 kg ha−1 in Aquadulce in 2015 (Table 3). Biological yield is one of the most sensitive traits to environmental conditions and varies greatly among the faba bean genotypes (Toker 2004; Abdalla et al. 2015; Flores et al. 1996; Gasim et al. 2015; Mitiku and Wolde 2015). Final biological yield represents the photosynthetic efficiency of the genotypes whereas the seed yield primarily related to the effective translocation of assimilates to the economic sinks. However, often a high correlation exists between the total biomass and final seed yield (Losset al. 1998b).
In current study, we found similar ranking of biomass and seed yield among the varieties (Table 3). For example, Aquadulce with highest biological yield out-yielded other varieties and produced the maximum seed yield. However, Lo’pez-Bellido et al. (2003) reported that although there is a certain correlation in faba bean crops between biological yield and seed yield, the high biological yield does not necessarily corresponds with the high seed yield.
Statistical analysis indicated that varieties of faba bean tested in this experiment produced significantly (P < 0.05) different seed yield (Table 3). Aquadulce and Delle Cascine, which produced the highest biomass, also ranked top as the most productive varieties and yielded 2756 and 2352 kg seed ha−1, respectively. Depite of being late maturity varieties, D’Aquadulce and Windsor performed poorly and produced only 900 and 993 kg seed ha−1, respectively. Interestingly Windsor is the dominant variety currently available to the growers in New England. The results of this study further confirmed the necessity of varieties/cultivars testing trials for each region. Final seed yield also depends on distribution efficiency of genotypes to translocate assimilates to the target sinks. The harvest index is a physiological criterion that represents the assimilation distribution efficiency and influenced by both the growing season and the genotype (Table 3). The harvest index range obtained in the current study (35–41%) is in the range of several other reports (Agung and McDonald 1998; Gebremeskel et al. 2011; Abdalla et al. 2015; Gezahegn et al. 2016). Agung and McDonald (1998) who stated that the HI of faba beans differed little among different genotypes have reported contrasting result.
The 100-seed weight was significantly different among the varieties (Table 3). The average seed size of Aquadulce, as the highest yielding variety, was 265 g 100-seed−1 which was the smallest among the tested varieties. Seed size of faba bean varieties varies greatly (Duc 1997; Landry et al. 2016; Etemadi et al. 2017) and such significant variation makes faba bean a unique legume crop when it comes to the cultivar evaluation. Faba bean varieties are classified as large-seeded (> 1.0 g seed−1), medium-seeded (0.5–1.0 g seed−1) and small seeded (< 0.5 g seed−1) (Etemadi et al. 2015). Regardless of the seed size, faba bean yield generally increases with increased population density and reaches its peak at different plant populations (L’opez-Bellido et al. 2005). Growers commonly use seed weight per unit land area to achieve the recommended population density. Therefore, in large seed crops such as faba bean, the seed size plays a significant role in determining the seed cost. Aquadulce not only out-yielded other varieties, also because of its smaller seed size, requires lower seeding rates. Lower seed cost promotes faba bean cultivation as cash/cover crop dual purpose.
Conclusion
The present study provides evidence that the faba bean genotypes exhibited dramatic variations in thermal units required to reach various phenological stages. This variation allows growers to select the proper genotype that favors their environmental conditions. The current study revealed that late-maturity varieties are not necessarily superior over early-maturity genotypes. Thus in short-season regions, high yielding early maturity varieties such as Aquadulce provide the opportunity for double cropping. Moreover, our findings confirm the existence of high correlation between biomass and seed yield. This correlation is very important in cultivation of legumes since production of higher biomass additionally provides higher N yield, which benefits the succeeding crop.
Abbreviations
- N:
-
Nitrogen
- HI:
-
Harvest index
- CGR:
-
Crop growth rate
References
Abdalla, A. A., Naim, E. L., Ahmed, A. M., & Taha, M. F. M. B. (2015). Biological yield and harvest index of faba bean (Vicia faba L.) as affected by different agro-ecological environments. World Journal of Agricultural Research, 3, 78–82.
Agegnehu, G., Ghizaw, A., & Sinebo, W. (2006). Yield performance and land-use efficiency of barley and faba bean mixed cropping in Ethiopian highlands. European Journal of Agronomy, 25(3), 202–207.
AghaAlikhani, M., Etemadi, F., & Ajirlo, A. F. (2012). Physiological basis of yield difference for grain sorghum (Sorghum bicolor (L.) Moench) in a semi-arid environment. ARPN Journal of Agricultural and Biological Science, 7, 488–496.
Agung, S., & McDonald, G. K. (1998). Effects of seed size and maturity on the growth and yield of faba bean (Vicia faba L.). Australian Journal of Agricultural, 49, 79–88.
Badran, M. S. S., & Ahmed, M. Z. D. (2010). Effect of sowing dates and planting methods on growth characters, seed yield and its components of faba bean in newly reclaimed lands. Journal of Agriculture and Environmental Science, Alexandria University, Egypt, 9(1), 53–66.
Barłóg, P., Grzebisz, W., & Łukowiak, R. (2018). Faba bean yield and growth dynamics in response to soil potassium availability and sulfur application. Field Crops Research, 219, 87–97.
Cline, G. R., & Silvernail, A. F. (2002). Effect of cover crops, nitrogen, and tillage on sweet corn. Hort Technology, 12(1), 118–125.
Coelho, J. C., & Pinto, P. A. (1989). Plant density effects on growth and development of winter faba bean (Vicia faba var. minor). Fabis Newsletter, 25, 26–30.
Dayoub, E., Naudin, C., Piva, G., Shirtliffe, S. J., Fustec, J., & Corre-Hellou, G. (2017). Traits affecting early season nitrogen uptake in nine legume species. Heliyon, 3(2), e00244. https://doi.org/10.1016/j.heliyon.2017.e00244
Denton, M. D., Phillips, L. A., Peoples, M. B., Pearce, D. J., Swan, A. D., Mele, P. M., et al. (2017). Legume inoculant application methods: Effects on nodulation patterns, nitrogen fixation, crop growth and yield in narrow-leaf lupin and faba bean. Plant and Soil, 419(1–2), 25–39.
Duc, D. (1997). Faba bean (Vicia faba L.). Field Crops Research, 53, 99–109.
Etemadi, F., & Hashemi, M., (2012). Morphological and phenological traits affecting grain sorghum yield. In: The second annual Life Sciences Graduate Research Symposium, University of Massachusetts (UMass), Amherst, USA.
Etemadi, F., Hashemi, M., Abbasi Shureshjani, R., & Autio, W. (2017). Application of data envelopment analysis to assess performance efficiency of eight faba bean varieties. Agronomy Journal, 109(4), 1225–1231.
Etemadi, F., Hashemi, M., Mangan, F., & Weis, S. (2015). Fava Beans; Growers guide in New England. Posted on Dairy, and livestock website: http://ag.umass.edu/sites/ag.umass.edu/files/research-reports/fava_bean_guide_2.pdf
Etemadi, F., Hashemi, M., Zandvakili, O., Dolatabadian, A., & Sadeghpour, A. (2018). Nitrogen contribution from winterkilled faba bean cover crop to spring-sown sweet corn in conventional and no-till systems. Agronomy Journal, 110(2), 455–462.
Finney, D. M., White, C. M., & Kaye, J. P. (2016). Biomass production and carbon/nitrogen ratio influence ecosystem services from cover crop mixtures. Agronomy Journal, 108, 39–52.
Flores, F., Moreno, M. T., Martinez, A., & Cubero, J. I. (1996). Genotype-environment interaction in faba bean: Comparison of AMMI and principal coordinate models. Field Crops Research, 47(2–3), 117–127.
Gan, Y. T., Miller, P. R., & McDonald, C. L. (2004). Response of kabuli chickpea to seed size and planting depth. Canadian Journal of Plant Science, 83(1), 39–46.
Gasim, S., Hamad, S. A. A., Abdelmula, A., & Mohamed Ahmed, I. A. (2015). Yield and quality attributes of faba bean inbred lines grown under marginal environmental conditions of Sudan. Food Science and Nutrition, 3(6), 539–547.
Gebremariam, A., & Assefa, F. (2018). The effect of inter cross-inoculation host group rhizobia on the growth and nitrogen fixation of Faba Bean (Vicia faba L.) varieties in North Showa, Amhara Regional State, Ethiopia. Journal of Agricultural Biotechnology and Sustainable Development, 10(2), 25–33.
Gebremeskel, Y., Estifanos, A., & Melaku, S. (2011). Effect of selected faba bean (Vicia faba L.) varietal difference on straw DM yield, chemical composition and nutritional quality. Journal of the Dry Lands, 4, 333–340.
Gezahegn, A. M., Tesfaye, K., Sharma, J. J., & Belel, M. D. (2016). Determination of optimum plant density for faba bean (Vicia faba L.) on vertisols at Haramaya, Eastern Ethiopia. Cogent Food & Agriculture, 2, 1224485. https://doi.org/10.1080/23311932.2016.1224485.
Girma, F., & Haila, D. (2014). Effect of supplemental irrigation on physiological parameters and yield of faba bean (Vicia faba. L) Varieties on the high land of Bale, Ethiopia. Journal of Agronomy, 13(1), 29–34.
Hegab, A. S. A., Fayed, M. T. B., Hamada, M. M. A., & Abdrabbo, M. A. A. (2014). Productivity and irrigation requirements of faba-bean in North Delta of Egypt in relation to planting dates. Annals of Agricultural Science, 59(2), 185–193.
Hoffmann, D., Jiang, Q., Men, A., Kinkema, M., & Gresshoff, P. M. (2007). Nodulation deficiency caused by fast neutron mutagenesis of the model legume Lotus Japonicus. Journal of Plant Physiology, 164, 460–469.
Horst, I., Welham, T., Kelly, S., Kaneko, T., Sato, S., Tabata, S., et al. (2007). Tilling mutants of Lotus Japonicus reveal that nitrogen assimilation and fixation can occur in the absence of nodule-enhanced sucrose synthase. Journal of Plant Physiology, 144, 806–820.
Hunt, C., & Yamada, S. (2003). Biotic resistance experienced by an invasive crustacean in a temperate estuary. Biological Invasions, 5, 33–43.
Jahanzad, E., Barker, A. V., Hashemi, M., Eaton, T., Sadeghpour, A., & Weis, S. (2016). Nitrogen release dynamics and decomposition of buried and surface cover crop residues. Agronomy Journal, 108, 1735–1741.
Jensen, E. S., Peoples, M. B., & Hauggaard-Nielsen, H. (2010). Review, Faba bean in cropping systems. 2010. Field Crops Research, 115(3), 203–216.
Köpke, U., & Nemecek, T. (2010). Ecological services of faba bean. Field Crops Research, 115(3), 217–233.
L’opez-Bellido, F. J., L’opez-Bellido, L., & L’opez-Bellido, R. J. (2005). Competition, growth and yield of faba bean (Vicia faba L.). European Journal of Agronomy, 23, 359–378.
L’opez-Bellido, R. J., L’opez-Bellido, L., L’opez-Bellido, F. J., & Castillo, J. E. (2003). Faba Bean (Vicia faba L.) Response to tillage and soil residual nitrogen in a continuous rotation with wheat (Triticum aestivum L.) under rainfed Mediterranean conditions. Agronomy Journal, 95, 1253–1261.
Landry, E., Coyne, C., & Hu, J. (2015). Agronomic performance of spring-sown faba bean (Vicia faba L.) in southeastern Washington. Agronomy Journal, 107, 574–578.
Landry, E. J., Coyne, C. J., McGee, R. J., & Hu, J. (2016). Adaptation of Autumn-Sown Faba Bean Germplasm to Southeastern Washington. Agronomy Journal, 108, 301–308.
Li, C., He, X., Zhu, S., Zhou, H., Wang, Y., Li, Y., et al. (2009). Crop diversity for yield increase. PLoS One. https://doi.org/10.1371/journal.pone.0008049.
Loss, S. P., Siddique, K. H. M., Jettner, R., & Martin, L. D. (1998a). Responses of faba bean (Vicia faba L.) to sowing rate in Southwestern Australia. Part I: Seed yield and economic optimum plant density. Australian Journal of Agricultural Research, 49, 989–997.
Loss, S. P., Siddique, K. H. M., Martin, L. D., & Crombie, A. (1998b). Responses of faba bean (Vicia faba L.) to sowing rate in Southwestern Australia. Part II: Canopy development, radiation absorption and dry matter partitioning. Australian Journal of Agricultural Research, 49, 999–1008.
Mirsky, S., Ackroyd, V., Cordeau, S., Curran, W., Hashemi, M., Reberg-Horton, S., et al. (2017). Hairy vetch biomass across the eastern U.S.: Effects of latitude, seeding rate and date, and termination timing. Agronomy Journal, 109(4), 1510–1519.
Mitiku, A., & Wolde, M. (2015). Effect of Faba Bean (Vicia faba L.) varieties on yield attributes at Sinana and Agarfa Districts of Bale Zone, Southeastern Ethiopia. Jordan Journal of Biological Sciences (JJBS), 4(4), 281–286.
Mwanamwenge, J., Loss, S. P., Siddique, K. H. M., & Cock, P. S. (1998). Growth, seed yield and water use of faba bean (Vicia faba L.) in a short-season Mediterranean-type Environment. Australian Journal of Experimental Agriculture, 38, 171–180.
Patrick, J. W., & Stoddard, F. L. (2010). Physiology of flowering and grain filling in faba bean. Field Crops Research, 115, 234–242.
Peng, S., Huang, J., Sheehy, J. E., Laza, R. C., Visperas, R. M., Zhong, X., et al. (2004). Rice yields decline with higher night temperature from global warming. Proceedings of the National academy of Sciences of the United States of America, 101(27), 9971–9975.
Pilbeam, C. J., Hebblethwait, P. D., Neyongesa, T. E., & Riketts, H. E. (1991). Effect of plant population density on determinate and indeterminate forms of winter field beans (Vicia faba) 2. Growth and development. The Journal of Agricultural Science, 116, 388–395.
Ricciardi, L., Polignano, G. B., & De Giovanni, C. (2001). Genotypic response of faba bean to water stress. Euphytica, 118, 39–46.
Ruisi, P., Amato, G., Badagliacca, G., Frenda, A. S., Giambalvo, D., & Miceli, G. D. (2017). Agro-ecological benefits of faba bean for rainfed Mediterranean cropping Systems. Italian Journal of Agronomy, 12, 233–245.
SAS Institute. (2003). SAS/STAT user’s guide. Version 9.1. Cary, NC: SAS Inst.
Sattell, R., Dick, R., & McGrath, D. (1998). Fava bean. Oregon State Univ. Ext. Serv. http://ir.library.oregonstate.edu/xmlui/bitstream/handle/1957/15226/em8697.pdf. Accessed 13 Mar 2016.
Song, Y. N., Zhang, F. S., Marschner, P., Fan, F. L., Bao, X. G., Sun, J. H., et al. (2007). Effect of intercropping on crop yield and chemical and microbiological properties in rhizosphere of wheat (Triticum aestivum L.), maize (Zea mays L.) and faba bean (Vicia faba L.). Biology and Fertility of Soils, 43, 565–574.
Sozen, O., & Karadavut, U. (2016). The yield traits of some cowpea (Vigna sinensis L.) genotypes cultivated in Kirșehİr province of Turkey. University of East Sarajevo, Faculty of Agriculture. CABI. Bosnia-Herzegovina. https://www.cabdirect.org/cabdirect/abstract/20173053773
Tafere, M., Tadesse, D., & Yigzaw, D. (2012). Participatory varietal selection of faba bean (ViciafabaL.) for yield and yield components in Dabat district, Ethiopia. Wudpecker Journal of Agricultural Research, 1, 270–274.
Talal, A. B., & Munqez, J. Y. (2013). Phenotypic Characterization of Faba Bean (Vicia faba L.) landraces grown in palestine. Journal of Agricultural Science, 5, 110–117.
Toker, C. (2004). Estimates of broad-sense heritability for seed yield and yield criteria in faba bean (Vicia faba L.). Hereditas, 140, 222–225.
Turk, M. A., & Tawaha, A. R. M. (2002). Impact of seeding rate, seeding date, rate and method of phosphorus application in faba bean (Vicia faba L. minor) in the absence of moisture stress. Biotechnology, Agronomy, Society and Environment, 6(3), 171–178.
Unkovich, M. J., & Pate, J. S. (2000). An appraisal of recent field measurements of symbiotic N2 fixation by annual legumes. Filed Crops Research, 65(2–3), 211–228.
Wilczek, A. M., Burghardt, L. T., Cobb, A. R., Cooper, M. D., Welch, S. M., & Schmitt, J. (2010). Genetic and physiological bases for phenological responses to current and predicted climates. Philosophical Transactions of the Royal Society B, 365, 3129–3147.
Zandvakili, O. R., Allahdadi, I., Mazaheri, D., Akbari, G. A., Jahanzad, E., & Mirshekari, M. (2012). Evaluation of quantitative and qualitative traits of forage sorghum and lima bean under different nitrogen fertilizer regimes in additive-replacement series. Journal of Agricultural Science, 4(6), 223–235.
Zhang, F., Shen, J., Li, L., & Liu, X. (2004). An overview of rhizosphere processes related with plant nutrition in major cropping systems in China. Plant and Soil, 260, 89–99.
Acknowledgements
This research partially supported by Northeast Sustainable Agriculture Research and Education, and University of Massachusetts Amherst Agricultural Experiment Station. The authors thank Dr. Sarah A. Weis (University of Massachusetts) for her contribution to the fieldwork.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Etemadi, F., Hashemi, M., Zandvakili, O. et al. Phenology, Yield and Growth Pattern of Faba Bean Varieties. Int. J. Plant Prod. 12, 243–250 (2018). https://doi.org/10.1007/s42106-018-0023-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42106-018-0023-1