
Abstract
Polistes is considered as one of the most widely and abundantly distributed, speciose genera of primitively eusocial wasps. The biology and behaviour of Polistes have been crucial to understanding the cooperation and conflicts among the nest members and have contributed greatly to our understanding of the functionality and evolution of eusociality. However, despite the wide geographical distribution, our perception of the genus has been shaped largely by the studies on the temperate species of Polistes. The diversity of Polistes species in Asia has been reflected through taxonomic reports but their nesting biology and behaviour are mostly unknown. Empirical studies on their behaviour in response to alternate conditions, like experimental removal of the queens, availability of extra food, etc. are also rare. The geographical origin of Polistes is also disputed. The speculation that Polistes originated in oriental regions can neither be confirmed nor refuted due to the lack of knowledge on Asian Polistes. Such missing information creates a large lacuna in our understanding of the origin and routes of dispersal of the genus. Here, we review the scattered studies on Asian Polistes and attempt to summarise and analyse the available taxonomic, behavioural and phylogenetic information. To date, 76 Polistes species have been reported from 31 Asian countries, out of which Vietnam, India and Japan have the maximum numbers of species. We also review the available behavioural studies on Polistes, mostly qualitative and conducted in Japan to show how these can provide a different perspective on Polistes biology. We also try to point out the major questions that need to be answered to get a broader overview of Asian Polistes. We suggest that more behavioural and phylogenetic studies on various species of Asian Polistes could provide the answers to questions like the geographic origin of the species, the evolution of hibernation (reproductive diapause) in the species, the diversification of nest founding strategies, the queen control on worker reproduction, etc.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The primitively eusocial, paper waspPolistes (Hymenoptera, Vespidae, Polistinae) is one of the most widely distributed genera of social insects. More than 200 species belonging to four subgenera1, 2 of Polistes have been reported from all continents except Antarctica. Understandably, Polistes has gained its due attention from the taxonomists but the genus has also been an attractive model for behavioural studies. The primitively eusocial status of the genus, i.e. reproductive caste differentiation in the absence of apparent morphological differences among the reproductives and the workers, has intrigued social insect biologists, ethologists and evolutionary biologists. Attempts to decode how the queens achieve their reproductive status have given rise to a plethora of behavioural, biochemical, physiological and theoretical studies. However, the most interesting interpretations about Polistes biology came from behavioural studies.
Polistes, as a typical member of Subfamily Polistinae, construct gymnodomous (uncovered) nests with less than a hundred individuals. Often these nests are made in man-made structures and are present in close proximity to human settlements. Due to such nesting habits and small colony size, the behaviour of individual wasps and that of the whole colony can be easily observed and recorded. Many interesting behavioural observations and empirical studies on various species of the genus have shaped our understanding of the biology of Polistes. However, most of these studies have been carried out on temperate species of Europe, North America and the neotropical species of South America.3,4,5 Although there are faunal records for at least 76 species from 31 Asian countries (Table 1) only a few species from Japan have been subjected to behavioural studies. There are at least two well-documented invasive species of Polistes, P. dominula, which invaded North America from Europe and P. chinensis which invaded Australia from Asia. Understandably, most studies have also been conducted on these two morphologically similar and phylogenetically close species.6, 7 Nevertheless, there is another widely distributed species in Asia – P. wattii, which has been reported from 12 Asian countries and appears to have a wide distribution like P. dominula. P. olivaceous, a phylogenetically close species to P. wattii,8 has been reported from 8 countries. Here, we attempt to review the available studies (Supplementary Tables 1 and 2) on various Asian species of Polistes and explore the need for more studies on Asian Polistes.
2 Diversity and Distribution of Polistes in Asia
In Asia, the focus of scientific work on Polistes has been mostly confined to taxonomic reporting. Using the available literature on such taxonomic records, we have prepared a database of species distribution of Polistes in Asia (Table 1, supplementary table 1). Carpenter1 described four subgenera of Polistes, Gyrostoma, Polistella and Polistes s.str and Aphanilopterus. Species belonging to first three subgenera occur in Asia, while subgenus Aphanilopterus mainly includes the South American species. According to the literature, Vietnam has the maximum number of reported species (35). Other species-rich countries in Asia are India (30), Japan (17), South Korea (11), China (10) Iran (10), Pakistan (9), and Nepal (9) (Table 1, Fig. 1). Plotting the numbers of species reported from different countries on the map (Fig. 2) shows the wide distribution and species density of Polistes in Asia. Despite the small geographic area, the species richness (number of species) of Vietnam can be contributed to the favourable, coastal, warm and humid climate and strategic biogeographic position as many species in Vietnam overlap with India and Japan. The species richness in India is also not surprising, given the large geographic area and various climatic regions. However, the same does not seem to be true for China. The relatively low species richness of China is probably due to the reporting bias caused by a lack of interest in taxonomic work or due to the publication of such work in regional language, which were not accessible to us. Nepal has 9 reported species, but 8 of them are also available in India. These species seem to have a continuous distribution due to the absence of any biogeographic barrier between the two countries.

Polistes species richness of different Asian countries. Only the countries with at least two reported species are shown here.

Distribution and density of Polistes species in Asia. The gradient is representative of numbers of species reported from different countries.
Some species have a wider distribution and have gained more attention from social insect biologists. The most common species (according to available literature) are P. wattii, P. olivaceous, P. rothneyi, P. stigma, P. sagittarius, P. strigosus, P. tenebricosus, P. chinensis, P. nimpha, P. indicus, P. biglumis and P. gigas (Fig. 3). P. chinensis has been studied in both Asia (mostly Japan) and New Zealand because of its invasive status.

Distribution of the most frequently reported species in Asia.
It can be assumed that taxonomic documentation of Polistes has not yet been conducted in many countries of Asia, or many regions of each country. The taxonomic studies are often published in print-only journals or in regional languages and have limited accessibility. We may have missed some species due to the non-accessibility of literature. Therefore, the species richness of each country and the distribution of each species must be an underestimation. Many species have been reported from multiple countries (Fig. 3) but we do not have an accurate distribution map for most of these species due to a lack of proper phylogeographic studies. Furthermore, we plotted the distribution and species richness according to political boundaries (Fig. 3) but political boundaries are not representative of physical landscapes.
There are many papers on the occurrences and morphological descriptions of many Polistes and other Vespid wasps from India. However, data from India (30 species) could also be underestimated, given that different climate specific zones and landscapes are present in India and a thorough sampling from all the states is awaited. The species diversity of India has been assessed by morphological identification and awaits molecular confirmation and phylogenetic interpretation. Despite the limitations and flaws, the database presented here is an attempt to understand and analyse the distribution of Polistes in Asia.
3 Origin of Polistes
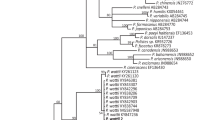
Polistes is undoubtedly the most abundant and widely distributed genus of Polistine wasps. The other well-studied and diverse genera of Polistine wasps, the neotropical Mischocyttarus, afrotropical Belanogaster and Ropalidia and palearctic and oriental Parapolybia, are restricted to narrower geographic distribution. Ecological success of Polistes over a wide range of geographical areas has given rise to many hypotheses including its geographic origin. The rich species diversity of Polistes in Asia has led to the hypothesis that Polistes may have originated in the oriental tropics or South East Asia and invaded the new world through Beringia and other parts of the world; the subsequent species diversity of the American continents is the result of such secondary radiation.6, 9 Molecular phylogenetic analysis failed to unambiguously confirm or reject this hypothesis but proposed that Polistes colonised the new world through South America and not Beringia.6 From such trees made with sequences that were available for Polistes from different parts of the world, it appears that the old world and new world taxa form sister clades and the former originated in Southeast Asia. Phylogenetic trees made with morphology or behavioural traits show that Asian species usually group together.6 Recent molecular phylogenetic analysis conducted with almost all species found in Vietnam shows the presence of subgroups even within the subgenus Polystella.10 Interestingly, P. wattii was never included in any phylogenetic tree until now despite its wide distribution in Asia. The recent inclusion of P. wattii in a phylogenetic tree shows that it forms a separate clade with P. olivaceous. P. wattii and P. olivaceous are two of the most reported species with overlapping geographical distribution in Asia (Fig. 3, Table 1) and their separate clade formation in the phylogenetic tree suggests the possibility of their unique ecological and behavioural traits.8 In this context, ecological comparison of old world and new world species and phylogenetic analysis with more Asian species may help resolve the questions regarding the origin of Polistes.
4 Polistes of Asia: Behavioural Data (Supplementary Table 2)
There are some behavioural studies available on following Asian Polistes – P. chinensis antennalis, P. rothneyi, P. japonicus, P. jokahamae, P. snelleni P. riparius. P. nimpha, P. olivaceous, P. biglumis, P. stigma and P. wattii. Most species reported from Japan have been subjected to observation-based natural historical studies or behavioural experiments. Such studies reveal interesting but scattered facts like P. chinensis and P. rothneyi have short nesting cycles (approximately four months) in Okushiri Islands, Japan11; the smaller females were more successful in founding nests in P. jadwigae12 etc.
There are at least 3 species (P. nipponensis, P. riparius and P. stigma) that are reported and studied in both Japan and Vietnam. It would not be surprising if these species are present in eastern China or Taiwan. Observation-based studies have also been conducted on other Polistes species in Vietnam. Although these species have not been extensively studied, yet the available information still alludes to the diversity in nesting biology, nest architecture, queen–worker dynamics, male production and mating behaviour (see below).
4.1 Nest Architecture
For some Asian species, the description of nests is available. Most species reported from Japan make small- to medium-sized, horizontally or vertically aligned single comb nests.13 Nest architecture of P. riparius and P. biglumis includes construction of long empty cells in the lower periphery of the nest which probably work as air chambers like the envelopes of vespine nests13,14,15 P. olivaceous16 and P. wattii8 nests may become very large with more than 1000 cells. Such large nests are not commonly seen in temperate Polistes with the only exception of P. annularis.5 P. wattii nests can be made of multiple combs (see later) which is extremely rare among Polistes species. P. riparius has also been reported to make large nests.14 This is in contrast to western temperate Polistes, which usually makes small- to medium-sized, circular nests with a single stalk.3 The extended period of warmer days in South Asia could be a reason for such flexible shapes and sizes.8 P. gigas, P. mandarinus and P. snelleni construct cells eccentrically while P. chinensis and P. jadwigae (= fadwigae) make concentric nests.17 P. mandarinus makes nests within vegetation, particularly among cedar trees. The pupal caps of P. mandarinus have a characteristic bright yellow colour, which is probably an adaptation for camouflage among leaves.18 Nest reconstruction after a hornet attack has been observed in P. jadwigae.19
4.2 Nesting Biology
Globally, both solitary and multiple founding are seen in Polistes. Some species strictly follow one of the strategies and some can employ both. When a species can use both solitary and multiple founding, multiple founding appears to be a more successful strategy (Reviewed in20). Some authors have suggested that Japanese Polistes follow only a solitary founding strategy. But multiple founding strategy has also been seen in some Japanese species (Table 2).21 Other Asian species like P. olivaceous and P. stigma occasionally use multiple founding strategy. In tropical areas, multiple founding should be more adaptive for various reasons22 and the same should be expected for P. stigma, P. olivaceous or P. wattii (in spring), which are more common in warmer countries. However, solitary founding is more prevalent even in these species. It should be noted that given the wide distribution of these species, and the low sample size in the available literature, variability within these phenotypes should be expected.
4.3 Hibernation and Nesting Cycle
One of the most prominent features of most Polistes species is reproductive diapause or hibernation of adult wasps. The females produced in the late colony phase prior to winter mate with males and pause the social life for a few months. During this time, the females find shelters (hibernacula) or gather at the back of the nest and spend the winter in total inactivity. These wasps emerge from diapause in spring or the beginning of summer and initiate nests alone or with other foundresses.3 All Asian (including Indian) Polistes for whom behavioural data is available, also follow the annual cycle. This widespread trait of overwintering or hibernation or diapause can be interpreted as an adaptation to survive harsh winter and also suggests a possible temperate origin of the genus.6 The absence of hibernation and ability to initiate nests anytime of the year in neotropical Polistes species of South America4 can be an adaptive variation.
4.4 Queen–Worker Dynamics
In primitively eusocial insects, the queen usually leads the dominance hierarchy and controls the workers through physical aggression. In western temperate Polistes species, the queen is the most dominant individual and she controls worker reproduction and centralised functioning of the colony through aggression.23 There are at least three reported instances, where the Polistes queens maintain reproductive dominance without being physically aggressive. Incidentally, all three species with the non-aggressive queen, P. jokahamae,24 P. japonicus25, 26 and P. snelleni27 are from Asia. The absence of physical aggression by the queen is typically found in highly eusocial species and only a few primitively eusocial species are known to control worker reproduction without physical aggression. Often, the lack of a dominant queen leads to worker reproduction (reviewed in28). Indeed, studies on worker reproduction using microsatellites reveal that worker reproduction, particularly male production by workers takes place in many Asian Polistes.29 Workers of P. snelleni and P. chinensis antennalis are capable of mating and producing female offspring in the presence of the queen or in orphaned colonies27, 30, 31 although production of female offspring only by a singly mated foundress has also been reported in P. snelleni.32 In Ropalidia marginata, the behaviour of the docile queen and the succession of the next queen through aggression has given rise to many interesting empirical studies.28, 33 Such empirical and observational studies of more Asian Polistes may change our view about how we perceive physical aggression in primitively eusocial organisms.
4.5 Worker Reproduction and Behaviour
Ovarian development in workers is known in Polistes.8, 34 In queenless (orphan) colonies, egg-laying by multiple workers has been reported in P. chinensis antennalis35 and P. jadwigae.36 However, physical aggression among the workers was seen in the former but not in the latter species. P. jadwigae workers produced a few female offspring while P. chinensis antennalis workers produced only male offspring.36 In orphaned colonies of P. nipponensis, workers could continue the nest and produce reproductives implying the mated status of workers. P. stigma workers also lay eggs in the presence of the queen.37 In case of P. wattii in late colony phase, the nests attain large colony sizes and development of worker ovaries is a common phenomenon. Worker behaviour with respect to division of labour, dominance or nest construction has been reported in some studies26, 37,38,39 by the Japanese social insect biologists but many such behavioural studies lack quantification of behaviour and statistical comparison of data.
4.6 Male Production
Males are usually produced late in the colony cycle in western temperate Polistes. The gynes mate with the males prior to hibernation. However, male production during early colony cycle (spring) has been reported in P. exclamans, P. gallicus and P. f. variatus.40, 41 In P. exclamans, P. metricus and P. annularis the early male production has been associated with frequent queen death and subsequent takeover by mated workers.40
We do not have much information on the male production of Asian species but early male production is also reported here. In case of P. chinensis, P. rothneyi and P. jadwigae,42 the males can be produced early in the colony cycle and the gynes can mate with them post hibernation in Spring (reviewed in3). In P. chinensis diploid males have been reported in early colony cycle.29, 43 In P. wattii, large number of males are produced in late colony cycle but occasionally males are also seen in spring.8
In social insects, males usually do not participate in any nest maintenance behaviour. Nevertheless, in some Polistine wasps, males were occasionally seen to feed larvae with food received from female workers.44 In Asian Polistes, this rare behaviour has been observed in P. jadwigae and P. snelleni.13, 45
4.7 Mating Behaviour
In comparison to social behaviour, mating behaviour has received very little attention in Polistes. Polistes males usually aggregate in leks to mate with foraging workers and gynes. Such territorial behaviour of males and their mating behaviour or mating associated behaviours have been observed in P. jadwigae,46 P. snelleni, P. chinensis and P. mandarinus.47,48,49 The timing of mating in Polistes may vary with respect to colony phase3 and the same is true for the Asian species. In P. chinensis, although the gynes destined for hibernation, emerge from August to September, mating occurs in November.49 A similar late mating has also been observed in P. jadwigae (reviewed in49). However, the unavailability of information on mating behaviour for most Asian species restricts us from making any comprehensive comparison of Asian Polistes with their western temperate or neotropical counterparts regarding mating or mating associated behaviours.
5 Asian Polistes: The Reasons to Study Them
Despite the wide diversity of Polistes, our perception of their behaviour is largely formed by the studies of temperate species of Europe, North America and Japan and the neotropical species of South America.3, 4 The research on Asian Polistes biology has been dominated by the work on P. chinensis in their native and invaded locations.21, 50,51,52,53 Behaviourally, P. chinensis is similar to its look-alike (particularly to novice eyes) and phylogenetically close species P. dominula (based on mitochondrial genes6). Studies on various Japanese and Vietnamese species show different features but it is difficult to infer a pattern on the behaviour of Asian Polistes from these unrelated studies.
All kinds of geographic and climatic conditions are available in Asia, which might be a reason for the rich species diversity of Polistes in this continent. It can be assumed that the environmental diversity and species diversity are also reflected in the behavioural diversity of different species. Exploration of their behaviour in a systematic manner, therefore, may result in the discovery of unique phenotypic traits and thus may open newer directions of social insect biology. A comparative behavioural study of the three subgenera of Polistes, that occur in Asia and a phylogenetic study including various species belonging to these three subgenera from different locations will also be helpful in getting a comprehensive view of the genus in Asia. However, currently, we lack systematic behavioural work and molecular data on different Asian species to conduct such a study.
Most behavioural work on Asian Polistes has been conducted on Japanese temperate species. We did not find any extreme difference between the western temperate Polistes and Japanese (temperate-zone) Polistes in literature. The Vietnamese and Indian species are representative of the tropical weather but in in both countries, Polistes research has been dominated by taxonomy and faunal records (but see8, 54). Although we can expect behavioural or geographic and climatic adaptations in Polistes from warmer Asian countries, till now, there are very few studies to support that. From India, we have found a unique nesting cycle in P. wattii, which is very different from either the temperate or the neotropical nesting cycles.
6 Polistes wattii: A Lesser-Known Species with Interesting Features
Indian scientists have contributed an enormous amount of knowledge in the field of wasp biology. Ropalidia marginata, the Indian Polistine (Tribe: Ropalidini) wasp is one of the most extensively studied social insect species in the world.28 However, we lag in our studies on Indian Polistes. In the last seven years, we have studied the nesting biology of P. wattii (Figs. 4, 5), which is abundantly found in North India and other Asian countries.8 Observational data indicate that this species is unique in many ways and it differs from all other known tropical and temperate Polistes. All temperate and most tropical Polistes species make almost circular small nests with a single stalk at the back and the nest usually hangs from a substrate.3 P. wattii makes nests of variable sizes and shapes and they can make nests in open spaces or in cavities (Figs. 4b, e, 5a). The nest architecture is extremely flexible and depends on the available space. Multiple combs are rarely found in Polistes with the exception of a few tropical species, as reviewed in.5 But in late summer, large nests of P. wattii with multiple combs stacked one below the other is a very common sight in North India (Fig. 5). Small combs are either connected to the substrate or to a previously built comb through one or more pedicels of varied widths.

A collage of Polistes wattii: a A forager collecting water b a solitary foundress nest (nest initiated by a single female) in an open space. c Two wasps sitting as guards at the nest entrance. d Workers sitting outside the entrance of a nest built inside a cavity. e A multiple foundress nest (nest initiated by multiple females) in an artificial cavity.

a A nest in October with multiple combs (circular/oval nest parts that contain many hexagonal cells, where eggs are laid). b Deepak Nain collecting wasps from the nest. c Ruchira Sen collecting wasps from the nest entrance.
Polistes wattii is also different than other known Polistes species in their nest founding strategies. Temperate Polistes wasps initiate nests in spring, produce workers in the first batch and then males and gynes in the second or the last batch. These nests can be started by either a single female (solitary foundress) or by multiple females. In winter, the mated gynes survive the unfavourable conditions by hibernation/diapause. Neotropical wasps in South America do not hibernate and can initiate nests any time of the year.4, 5 Although P. wattii is an annual species that hibernates in winter, the nest founding strategy of P. wattii is different from any other known temperate and tropical Polistes. This species follows a strictly solitary founding strategy in spring but these nests do not survive beyond May–June and in summer, they use multiple founding strategy and initiate nests again. Such a biannual, alternative nesting strategy has not been reported in any other Polistes species. In the middle of summer nest-free clusters of healthy and stylopised wasps are seen, which may be linked with the initiation of summer nests by multiple foundresses.
7 Reasons Behind Lack of Behavioural Studies on Polistes in Asia
It can be assumed that the wide range of environmental and geographical conditions may have induced many behavioural specialisations in the Asian Polistes species but there are not many studies to support this assumption. Apart from the danger of painful sting imparted by female Polistes, there are some other challenges that restrict behavioural studies. First, quantitative behavioural experiments require standardisation of ethogram and long, meticulous observations. Those who are ready to do that often focus on the economically important organisms. Second, the convenience of collecting wasps and studying their morphological features in the lab has led to an abundance of taxonomic studies on wasps in Asian countries. Behavioural studies on Polistes have been mostly monopolised by western scientists on western species. It is, however, possible that there are some behavioural studies published in regional languages or in print-only journals in different countries of Asia but they remain hidden to current researchers. We may not have a proper explanation for the lack of studies on Asian Polistes till now but for a better understanding of the evolution and diversification of genus we need more behavioural and phylogenetic studies. One major exception is the biology of another polistine wasp, Ropalidia marginata, which has been extensively studied by Professor Raghavendra Gadagkar and his group.28 However, it is not enough to make a comprehensive estimation of the social wasps of the continent and more such studies are required to follow particularly on Polistes.
8 Conclusion
Evans (1958) suggested, Polistes is the key genus to understand the evolution of eusociality.55 Currently, our knowledge about this genus is biased with the available information on the new world species. We need to explore (but not limit ourselves to) the following questions to get a comprehensive view of the biology and behaviour of Polistes. (1) Systematic behavioural ecology on representative species from different phylogenetic clades. (2) Population ecology of the more abundant species to understand the nature of their dispersal. (3) Behavioural experiments to document the mating behaviour and studying behavioural isolation among the sympatric species with similar colouration and markings. (4) The adaptive values of the intersexual and intrasexual body markings. (5) Identification of possible strepsipteran parasites from different species of Asian Polistes (reported from only P. gigas, P. rothneyi87 and P. wattii8 and the effect of Polistes–Strepsiptera association on the behaviour and biology of Polistes. (6) The reason for the abundance of solitary founding in Asian Polistes.
Although there are some behavioural studies on Japanese Polistes and taxonomic studies from different countries of Asia, we know very little about the species that occur in the warmer countries of West and South Asia (Supplementary Table 1). For a better understanding of evolution of eusociality it is crucial that we ask questions regarding the geographic origin of the genus, the evolution of hibernation in the genus, the diversification of nest founding strategies, the queen–worker and foundresses dynamics, occurrence of worker reproduction etc. Such questions have been explored in temperate and neotropical species but our understanding will remain incomplete without studying the Asian species. We need to explore the behavioural, chemical and molecular ecological aspects of Polistes from different Asian countries to get a comprehensive perspective of the genus.
Data availability
This is a review article and therefore does not contain any raw data.
References
Carpenter JM (1996) Distributional checklist of species of the genus Polistes (Hymenoptera: Vespidae: Polistinae: Polistini). Am Museum Novit 3188:1–40
Gawas SM, Girish Kumar P, Pannure A, Gupta A, Carpenter JM (2020) An annotated distributional checklist of Vespidae (Hymenoptera: Vespoidea) of India. Zootaxa 4784(1):001–087
Reeve HK (1991) Polistes. In: Ross MRW, Keneth G (eds) The Social biology of wasps. Cornell University Press
Prezoto SA, Nascimento F, Barbosa BC, Somavilla A (2021) Neotropical social wasps basic and applied aspects. Springer Nature Switzerland AG
Hunt JH (2007) The evolution of social wasps. Oxford University Press
Santos BF, Payne A, Pickett KM, Carpenter JM (2015) Phylogeny and historical biogeography of the paper wasp genus Polistes (Hymenoptera: Vespidae): implications for the overwintering hypothesis of social evolution. Cladistics 31(5):535–549. https://doi.org/10.1111/cla.12103
Pickett KM, Carpenter JM, Wheeler WC (2006) Systematics of Polistes (Hymenoptera: Vespidae), with a phylogenetic consideration of Hamilton’s haplodiploidy hypothesis. Ann Zool Fennici 43(5–6):390–406
Sen R, Malhotra K, Gupta M, Kaur R, Bawa D, Duhan M, Sandhi S, Songara P, Nain D, Raychoudhury R (2022) Coping with the ‘Indian summer’: unique nesting cycle and nest architecture of the paper wasp, Polistes wattii. Sci Nat 109(31):1–12. https://doi.org/10.1007/s00114-022-01801-0
Van der Vecht J (1965) The geographic distribution of social wasps (Hymenoptera: Vespidae). In: Proc. 12th International Congr. Entomol. London, pp 440–441
Nguyen LTP, Nguyen AD, Nguyen TTP, Perrard A, Carpenter JM (2018) Molecular phylogeny of the paper wasp subgenus Polistes (Polistella) Ashmead, 1904 (Hymenoptera : Vespidae : Polistinae) from Vietnam. J Asia Pac Entomol 21(2):638–644. https://doi.org/10.1016/j.aspen.2018.04.006
Yamane S (1972) Life cycle and nest architecture of Polistes wasps in the Okushiri Island, Northern Japan (Hymenoptera, Vespidae ). J Fac Sci Hokkaido Univ. Ser. VI, Zool. 18(3):440–459
Tsuchida K (1991) Effects of female body size and timing of nest foundation on the reproduciton of the Japanese Paper Wasp, Polistes jadwigae Dalla Torre (Hymenoptera: Vespidae). Res Popul Ecol 33:361–366
Soichi Y (1969) Preliminary observations on the life history of two polistine wasps, Polistes Snelleni and P. biglumis in Sapporo, Northern Japan. J Fac Sci Hokkaido Univ Ser. VI, Zool 17:78–105
Yamane S, Kudo S, Tajima T, Nihon’yanagi K, Shinoda M, Saito K, Yamamoto H (1999) Comparison of investment in nest construction by the foundresses of Consubgeneric Polistes wasps, P. (Polistes) riparius and P. (P.) chinensis (Hymenoptera, Vespidae). J Ethol 16(1986):97–104
Hozumi S, Yamane S (2001) Incubation ability of the functional envelope in paper wasp nests (Hymenopteta, Vespidae, Polistes): I. Field measurements of nest temperature using paper models. J Ethol 19:39–46
Alam SM (1959) Some interesting revelations about the nest of Polistes hebroeus Fabr". (Vespidae, Hymenoptera)-the common yellow wasp of India. Proc Zool Soc 11(2):113–122
Iwata K (1965) Description of the nest of the giant paperwasp, Polistes gigas Kirby from Formosa, with some ethological considerations (Hym., Vespidae). Konytû 33(3):284–290
Hagiwara Y, Kojima J (1994) Options in Construction behavior for Polistes mandarinus Saussure nesting on Japanese Cedar Twigs (Hymenoptera: Vespidae). J Kan Entomol Soc 67(1):126–128
Hibino Y, Kasuya E, Ito Y (1980) Reconstruction of Nests by Polistes jadwigae and Polistes chinensis antennalis workers. Appl Entomol Zool 15(2):190–193
Tibbetts EA, Reeve HK (2003) Benefits of foundress associations in the paper wasp Polistes dominulus: increased productivity and survival, but no assurance of fitness returns. Behav Ecol 14(4):510–514
Hoshikawa T (1979) Observations on the Polygynous Nests of Polistes chinensis antennalis Perez (Hymenoptera: Vespidae) in Japan. Kontyu 47(3):239–243
Suzuki T, Ramesh M (1992) Colony founding in the social wasp, Polistes stigma (Hymenoptera Vespidae), in India. Ethol Ecol Evol 4(4):333–341
Jandt JM, Tibbetts EA, Toth AL (2014) Polistes paper wasps: a model genus for the study of social dominance hierarchies. Insect Soc 61:11–27. https://doi.org/10.1007/s00040-013-0328-0
Yoshimura H, Yamada J, Yamada YY (2019) The queen of the paper wasp Polistes jokahamae (Hymenoptera: Polistinae) is not aggressive but maintains her reproductive priority. Sociobiology 66(1):166–178. https://doi.org/10.13102/sociobiology.v66i1.3577
Ishikawa Y, Yamada YY, Matsuura M, Tsukada M, Tsuchida K (2011) Polistes japonicus (Hymenoptera, Vespidae) queens monopolize ovipositing but are not the most active aggressor in dominant-subordinate interactions. Insectes Soc 58:519–529. https://doi.org/10.1007/s00040-011-0173-y
Ishikawa Y, Yamada YY, Matsuura M, Tsukada M, Tsuchida K (2010) Dominance hierarchy among workers changes with colony development in Polistes japonicus (Hymenoptera, Vespidae) paper wasp colonies with a small number of workers. Insectes Soc 57(4):465–475. https://doi.org/10.1007/s00040-010-0106-1
Tsuchida K, Yamasaki K (2014) Orphaning does not affect the colony productivity of the primitive eusocial wasp Polistes snelleni. Insect Soc 61:133–140. https://doi.org/10.1007/s00040-013-0336-0
Gadagkar R (2001) The social biology of Ropalidia marginata toward understanding the evolution of eusociality, Harvard University Press, Cambridge, Masachusetts London England
Tsuchida K, Saigo T, Tsujita S, Takeuchi K (2004) Early male production is not linked to a reproductive strategy in the Japanese paper wasp, Polistes chinensis antennalis (Hymenoptera: Vespidae). J Ethol 22(1):119–121. https://doi.org/10.1007/s10164-003-0109-8
Tsuchida K, Saigo T, Tsujita S, Takeuchi K, Ito N, Sugiyama M (2003) Polymorphic microsatellite loci for the Japanese paper wasp, Polistes chinensis antennalis (Hymenoptera: Vespidae). Mol Ecol Notes 3(3):384–386. https://doi.org/10.1046/j.1471-8286.2003.00457.x
Suzuki T (2005) Insemination of workers prior to assuming the position of queen in a temperate paper wasp Polistes snelleni Saussure ( Hymenoptera Vespidae ). Ethol Ecol Evol 17:335–339. https://doi.org/10.1080/08927014.2005.9522587
Sayama K, Takahashi JI (2005) Mating structure and genetic relatedness among gynes in the primitively eusocial wasp Polistes snelleni (Hymenoptera: Vespidae). Entomol Sci 8(1):27–31. https://doi.org/10.1111/j.1479-8298.2005.00097.x
Gadagkar R (2009) Interrogating an insect society. Proc Natl Acad Sci USA 106(26):10407–10414. https://doi.org/10.1073/pnas.0904317106
Gobbi N, Noll FB, Penna MAH (2006) ‘Winter’ aggregations, colony cycle, and seasonal phenotypic change in the paper wasp Polistes versicolor in subtropical Brazil. Naturwissenschaften 93(10):487–494. https://doi.org/10.1007/s00114-006-0140-z
Miyano S (1986) Colony development, worker behaviour and male production in orphan colonies of a Japanese paper wasp, Polistes chinensis antennalis Perez (Hymenoptera: Vespidae). Res Popul Ecol 28:347–361
Miyano S (1991) Worker reproduction and related behavior in orphan colonies of a Japanese paper wasp, Polistes jadwigae (Hymenoptera, Vespidae ). J Ethol 9:135–146
Suzuki T (1996) Natural history and social behaviour of the cofoundresses in a primitively eusocial wasp, Polistes stigma (Fabricius) (Hymenoptera: Vespidae), in India: a case study. Jpn J Ent 64(1):35–55
Kasuya E (1980) Behavioral ecology of Japanese paper wasps Polistes spp (Hymenoptera: Vespidae) 1. Extranidal activities of Polistes chinensis antennalis. Res Popul Ecol 22(2):242–254
Tsuchida K (1991) Temporal behavioural variation and division of labor among workers in the primitively eusocial wasp, Polistes jadwigae Dalla Torre. J Ethol 9:129–134
Strassmann JE (1981) Evolutionary implications of early male and satellite nest production in Polistes exclamans colony cycles. Behav Ecol Sociobiol 8(1):55–64. https://doi.org/10.1007/BF00302844
Page RE, Post DC, Metcalf RA (1989) Satellite nests, early males, and plasticity of reproductive behavior in a paper wasp. The Am Nat 134(5):731–748. https://doi.org/10.1086/285008
Kasuya E (1983) Social behaviour of early emerging males of a Japanese Paper Wasp, Polistes chinensis antennalis (Hymenoptera: Vespidae). Res Popul Ecol 25:143–149
Tsuchida K, Nagata N, Kojima J (2002) Diploid males and sex determination in a paper wasp, Polistes chinensis antennalis (Hymenoptera, Vespidae). Insectes Soc 49(2):120–124. https://doi.org/10.1007/s00040-002-8289-8
Sen R, Gadagkar R (2006) Males of the social wasp Ropalidia marginata can feed larvae, given an opportunity. Anim Behav 71(2):345–350. https://doi.org/10.1016/j.anbehav.2005.04.022
Makino S (1993) Sexual differences in larval feeding behavior in a paper wasp, Polistes jadwigae (Hymenoptera, Vespidae). J Ethol 11(1):73–75. https://doi.org/10.1007/BF02350009
Kasuya E (1981) Male mating territory in a Japanese Paper Wasp, Polistes jadwigae Dalla Torre (Hymenoptera, Vespidae). Kontyû 49(4):607–614
Suzuki T (1995) Seasonal timing of mate-searching activity in males of two congeneric paper wasps, Polistes snelleni and Polistes chinensis, (Hymenoptera, Vespidae). Jpn J Ent 63(2):425–432
Suzuki T (1998) Paradox of worker reproduction and worker mating in temperate paper wasps, Polistes chinensis and P. snelleni (Hymenoptera Vespidae). Ethol Ecol Evol 10(4):347–359. https://doi.org/10.1080/08927014.1998.9522848
Kojima T, Suzuki JI (1986) Timing of mating in five Japanese polistine wasps (Hymenoptera: Vespidae): anatomy of fall females. J Kansas Entomol Soc 59(2):401–404
Furuichi S, Kasuya E (2013) Costs, benefits, and plasticity of construction of nest defensive structures in paper wasps. Behav Ecol Sociobiol 68(2):215–221. https://doi.org/10.1007/s00265-013-1636-0
Tadashi S (1981) Male production by foundresses of Polistes chinensis antennalis (Hymenoptera, Vespidae) in the founding stage, Kontyû. Tokyo 49(3):519–520
Suzuki T (1981) Effect of photoperiod on male egg production by foundresses of Polistes Chinensis antennalis Perez (Hymenoptera, Vespidae). Jpn J Ecol 31(4):347–351. https://doi.org/10.18960/seitai.31.4_347
Kudô K (2003) Growth rate and body weight of foundress-reared offspring in a paper wasp, Polistes chinensis (Hymenoptera, Vespidae): no influence of food quantity on the first offspring. Insectes Soc 50:77–81. https://doi.org/10.1007/s000400300012
Pham PH (2014) Hibernation on the Nest of the Paper wasp, Polistes (Gyrostoma) olivaceus (De Geer) (Hymenoptera : Vespidae). Biol Forum - An Int J 6(1):116–119
Evans H (1958) The evolution of social life in wasps. In: Proceedings of the 10th international congress of entomology, Montreal, vol 2, pp 449–457
Jonathan JK, Roy SB, Kundu BG (2000) Insecta: Hymenoptera: Vespidae. In: Zool. Surv. India State Fauna Ser. 4 Fauna of Meghalaya, vol 7, pp 117–160
Yamane S, Yamane S (1979) Polistine wasps from Nepal (Hymenoptera, Vespidae). Insecta Matsumurana 15:1–37
Dorji P, Klein W, Nidup T, (2017) Taxonomic study of social vespid wasps (Hymenoptera : Vespidae : Vespinae & Polistinae) in Bhutan. J Insect Biodivers Syst 03(2):91–104
Dvořák L, Carpenter JM (2008) The first record of the paper wasp Polistes smithii neavei von schulthess, 1921, from the middle east (Hymenoptera: Vespidae), with notes on the social wasp fauna of Yemen. Zool Middle East 44(1):119–120. https://doi.org/10.1080/09397140.2008.10638298
Schmid-Egger C, Van Achterberg K, Neumeyer R (2017) Revision of the West Palaearctic Polistes Latreille, with the descriptions of two species - an integrative approach using morphology and DNA barcodes (Hymenoptera, Vespidae). Zookeys 713:53–112. https://doi.org/10.3897/zookeys.713.11335
Rahmani Z, Rakhshani E, Carpenter JM (2020) Updated checklist of Vespidae (Hymenoptera : Vespoidea) in Iran. J Insect Biodivers Syst 06(1):27–86
Yamane S, Kawamichi T (1975) Bionomic comparison of Polistes biglumis (Hymenoptera, Vespidae) at two different localities in Hokkaido, Northern Japan with reference to its probable adaptation to cold climate. Kontyû, Tokyo 43(2):214–232
Saito F, Kojima JL, Ubaidillah R, Hartini S (2005) Paper wasps of the genus in Eastern Lesser Sunda Islands (Hymenoptera: Vespidae). J Hym Res 14(1):102–114
Kim JK (2023) Taxonomic review of Polistes (Polistella) (Hymenoptera : Vespidae : Polistinae) in Korea, with a description of a new species and a new status species and a new status. Oriental Insects 57(3):908–934 https://doi.org/10.1080/00305316.2022.2163516
Lien NTP (2020) Contribution to the taxonomy of the paper wasp genus Polistes (Polistella) (Hymenoptera: Vespidae: Polistinae) from Vietnam, with description of a new species. Zootaxa 4759(2):252–260
Petersen BR (1990) Polistine wasps of the subgenera Stenopolistes and Megapolistes from the Philippine, Moluccan and Bismarck Islands (Hymenoptera: Vespidae). Insect Syst Evol 21(1):53–66
Thi L, Nguyen P, Kojima J, Saito F (2011) Polistes (Polistella) wasps (Hymenoptera: Vespidae: Polistinae) from mountainous areas of northern Vietnam, with description of five new speceis. Zootaxa 3060:1–30
Kojima J (1997) Taxonomic notes on the social wasps in the Kanto Plain, central Japan (Insecta : Hymenoptera : Vespidae). Nat His Bull Ibaraki Univ 1:17–44
Dvořák L (2007) The social wasp fauna of Pakistan (Hymenoptera, Vespidae). Linzer Biol Beitr 39(1):5–55
Kojima J, Saito F, Thi L, Nguyen P (2011) On the species-group taxa of Taiwanese social wasps (Hymenoptera: Vespidae) described and/or treated by J. Sonan. Zootaxa 64(May):42–64
Barthélémy C (2012) Nest trapping, a simple method for gathering information on life histories of solitary bees and wasps. Bionomics of 21 species of solitary aculeate in Hong Kong. Hong Kong Entomol Bull 4(1):3–37
Kifune T, Maeta Y (1985) Taxonomical studies on the Genus Xenos (Strepsiptera, Stylopidae) parasitic on Vespa and Polistes (Hymenoptera, Vespidae) of Taiwan with descriptions of three new species, Konytû. Tokyo 53:426–435
Oh S, An S, Lee J (2012) Review of Korean Latibulus (Hymenoptera: Ichneumonidae: Cryptinae) and a key to the world species. Can Entomol 144:509–525. https://doi.org/10.4039/tce.2012.54
Song SN, Chen PY, Wei SJ, Chen XX (2016) The mitochondrial genome of Polistes jokahamae and a phylogenetic analysis of the Vespoidea (Insecta: Hymenoptera). Mitochondrial DNA 27(4):2783–2784. https://doi.org/10.3109/19401736.2015.1053065
Yoshimura H, Yamada YY (2021) Preimaginal caste - related bias in the paper wasp Polistes jokahamae is limited to the first brood. Insectes Soc 68(1):133–143. https://doi.org/10.1007/s00040-020-00805-1
Rafi MA et al (2017) The vespid fauna of Pakistan. Zootaxa 4362(1):1–28
Feng Y, Long S (2010) Common edible wasps in Yunnan Province, China and their nutritional value. In: Durst PB, Johinson DV, Leslie RN, Shono K (eds) Forest insects as food : humans bite back, food and agriculture organization of the United Nations regional office for Asia and the Pacific Bangkok, Thailand, p 93
Khan K et al (2018) Chemical composition, structure and architecture of the nest of various species of Vespidae (Insecta: Hymenoptera). Jokull J 68(3):22–34
Handru A, Nugroho H, Saito-Morooka F, Ubaidillah R, Kojima J (2020) Eusocial wasp fauna of Sulawesi Island, the central island of Wallacea (Hymenoptera: Vespidae; Polistinae, Vespinae). Zootaxa 4885(4):541–559
Yamane S, Kusigemati K (1985) Vespoidea and Scoliidae from the Fiji and Solomon Islands (Insecta, Hymenoptera). Kagoshima Univ Res Cent S Pac Occas Paper 5:75–79
Starr CK (1992) The Social Wasps (Hymenoptera: Vespidae) of Taiwan. Bull Natl Museum Nat Sci 3:93–138
Dorji P, Klein W, Nidup T (2016) Study of paper wasps (Hymenoptera: Vespidae: Polistinae) of Bhutan. Bio Bull 2(1):128–136
Choi MB, Kim JK, Lee JW (2013) Checklist and distribution of Korean Vespidae revisited. Korean Jou of Appl Entomol 52(2):85–91
Kudô K, Yamane S, Yamamoto H (1998) Physiological ecology of nest construction and protein flow in pre-emergence colonies of Polistes chinensis (Hymenoptera Vespidae): Effects of rainfall and microclimates. Ethol Ecol Evol 10(2):171–183. https://doi.org/10.1080/08927014.1998.9522865
Makino S, Yamane S, Sunose T, Aoki S (1987) Dispersion distance of queens from natal sites in the two haplometrotic paper wasps Polistes riparius and P. snelleni (Hymenoptera: Vespidae). Res Popul Ecol 29:111–117
Lorenzi MC, Cervo R (1995) Usurpations and late associations in the solitary founding social wasp, Polistes biglumis bimaculatus (Hymenoptera: Vespidae). J Insect Behav 8(4):443–451. https://doi.org/10.1007/BF01995318
Kathirithamby J (1993) Descriptions of Strespsiptera (Insecta) from Southeast Asia, with a checklist of the genera and species occurring in the region. Raffeles Bull Zool 41(2)
Acknowledgements
The authors thank Prof Raghavendra Gadagkar for the invitation to write this review, Dr Rhitoban Ray Choudhury for his comments on the manuscript and two anonymous reviewers for their comments on the earlier version of the manuscript.
Funding
This work was supported by the SERB-CRG grant CRG/2021/007010 awarded to RS. DN was supported by DST-INSPIRE fellowship (DST/INSPIRE/03/2021/000175, IF200146).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Nain, D., Sen, R. A Review of Our Meagre Knowledge of Asian Polistes, and a Call for More Studies. J Indian Inst Sci 103, 1049–1064 (2023). https://doi.org/10.1007/s41745-023-00402-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41745-023-00402-8