
Abstract
Three bearing mid-late Oxfordian and early Kimmeridgian ammonite stratigraphic sections, corresponding to the Santiago Formation, located in the surroundings of Tamán (San Luis Potosí), Tenango, and La Mesa-El Pescado (both in Hidalgo) towns were studied. The presence of the genera Passendorferia, Neumannia, Sequeirosia (Gemmellarites), Perisphinctes (Otosphinctes), and Gregoryceras support the existence of an extensive communication via with the Tethys during the Plicatilis-Transverarium Chrons. On the other hand, for the Bifurcatus Chron, fauna with Tethys affinity is still recognized, including the genera Neoprionoceras, Subdiscosphinctes, Ochetoceras, and Euaspidoceras. In addition, is possible to establish a paleobiogeographic affinity with Cuba, Chile, and Argentina based on the occurrence of Vinalesphinctes, Perisphinctes (Cubasphinctes), and Caracoliceras, which is also supported by the presence of Physodoceras and Subnebrodites in the Hypselum/Bimammatum Chrons. Finally, the record of Passendorferia, Ochetoceras, and Euaspidoceras throughout the entire stratigraphic succession confirms that the connection with the Tethys was present throughout the Oxfordian and early Kimmeridgian. Based on these paleogeographic interpretations, the Tethys influence on the Oxfordian-Kimmeridgian Mexican ammonite faunas is confirmed. For areas currently integrated into Cuba, the record in Mexico of forms with Cuban affinity reveals the existence of a paleobiogeographic entity in the northern margin of the western portion of the Hispanic Corridor; while the faunal affinity with Chile and Argentina is interpreted as evidence of a connection with these epicontinental areas, which favors dispersal by marine currents.
Resumen
Se estudiaron tres secciones estratigráficas de amonites del Oxfordiano medio-tardío y Kimmeridgiano temprano, correspondientes a la Formación Santiago, ubicadas en los alrededores de los poblados de Tamán (San Luis Potosí), Tenango y La Mesa-El Pescado (ambas en el estado de Hidalgo). La presencia de los géneros Passenderforia, Neumannia, Sequeirosia (Gemmellarites), Perisphinctes (Otosphinctes) y Gregoryceras apoyan la existencia de una extensa comunicación a través del Tethys durante los Crones Plicatilis-Transverarium. Por otro lado, para el Cron Bifurcatus la fauna con afinidad del Tethys es ya reconocida, incluidos los géneros Neoprionoceras, Subdiscosphinctes, Ochetoceras y Euaspidoceras. Además, es posible establecer una afinidad paleobiogeográfica con Cuba, Chile y Argentina basada en la ocurrencia de Vinalesphinctes, Perisphinctes (Cubasphinctes) y Caracoliceras, lo cual es soportado por la presencia de Physodoceras y Subnebrodites en los Crones Hypselum/Bimammatum. Finalmente, el registro de Passendorferia, Ochetoceras y Euaspidoceras a lo largo de toda la sucesión estratigráfica confirma que la conexión con el Tethys estuvo presente durante todo el Oxfordiano y el Kimmeridgiano temprano. Con base en estas interpretaciones paleogeográficas se confirma la influencia del Tethys en las faunas de amonites mexicanos del Oxfordiano-Kimmeridgiano. Para áreas actualmente integradas a Cuba, el registro en México de formas con afinidad cubana revela la existencia de una entidad paleobiogeográfica en el margen norte de la porción occidental del Corredor Hispánico; mientras que la afinidad faunística con Chile y Argentina se interpreta como evidencia de una conexión con estas áreas epicontinentales, lo que favorece la dispersión por corrientes marinas.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Reliable biostratigraphic information has been generated as a result of the development of detailed studies with strict stratigraphic control. The aforementioned opens possibilities for carrying out research where biogeographic and paleogeographic interpretations are involved (i.e. Buitrón, 1984; Cantú-Chapa, 2001; Villaseñor et al., 2005). It is worth mentioning that these studies have been based mainly on ammonite faunas, which are recognized as the main faunal elements to establish relative ages in stratigraphy, as well as in regional and global correlation; being very important for paleobiogeographic, paleobiological and evolutionary studies, both of the marine Paleozoic and Mesozoic.
There are some publications discussing the paleogeographic distribution of Oxfordian-Kimmerdigian ammonite associations in Mexico, and relating it to the Caribbean Province (Cariou, 1973; Myczyński & Meléndez, 1990). Besides, it has been considered that during the middle Oxfordian the northwestern portion of the Proto-Caribbean basin was probably connected with the eastern part of Mexico, existing a connection with the fauna of the Andean Province (Myczyński, 1994).
In particular, according to the occurrence of the genus Vinalesphinctes SPATH, 1931 in Chile and Cuba, Melendez and Myczyński (1994) point out the existence of biogeographic connections between these two areas through the Hispanic Corridor, and mention the possibility of a Pacific origin of this genus; although they express the need for more data to corroborate this. On the other hand, López-Palomino et al. (2006) and López-Palomino (2013) highlight the importance of Vinalesphinctes in biostratigraphic correlations in the Upper Jurassic (Oxfordian) of America and its paleobiogeographic dynamics.
These authors mention that the geographical distribution of this genus, restricted to Cuba, Chile, and Mexico, gives it an endemic character for the Americas, which would mean that during the Oxfordian these areas were close, representing a discontinuous epicontinental marine environment, connected with both the western Tethys and the eastern Pacific. Likewise, they mention that the presence of different species in Mexico and Cuba agrees with the interpretations made by Myczyński et al. (1998, p. 201) on the increase in endemism in ammonite fauna during the Bifurcatus Chron. On the other hand, Parent et al. (2006) document the occurrence of Subvinalesphinctes WIERZBOWSKI, 1976, Vinalesphinctes, and Cubasphinctes JUDOLEY Y FURRAZOLA-BERMÚDES, 1968 from the middle Oxfordian (lower part of the Plicatilis Zone) to the late Oxfordian (lower part of the Bifurcatus Zone) in Cuba, Mexico, Chile, and Argentina; as well as their probable origin from the species Subvinalesphinctes pseudokranaus, would mainly indicate Tethys and Tethys-Caribbean biogeographic affinities.
Furthermore, the occurrence in Mexico of the genus Gregoryceras SPATH, 1924, beyond its biostratigraphic significance, has allowed important paleobiogeographic and paleoenvironmental interpretations for the Oxfordian Mexican marine deposits (Transversarium Chron), which represent an epicontinental platform related to transgressive events. The aforementioned is based mainly on the morphological and ontogenetic traits of Gregoryceras.
Olóriz et al. (2008, 2010) agree with the importance of the record of Gregoryceras in Mexico, pointing out that it is the most complete continuous paleobiogeographic range of this genus in the Americas, in an intermediate area between the records of northern Chile and the northern edge of the Gulf of Mexico Basin. Those authors also mentioned that the occurrence of Gregoryceras, together with perisphinctids previously documented and others reported for the very first time in Mexico, corroborates Tethys origin for the ammonite association, at least during the Oxfordian. Additionally, they mentioned that the presence of Gregoryceras in Mexico and Chile, Dichotomosphinctes BUCKMAN, 1926 in Mexico, and ochetoceratids in Mexico and Cuba, may be the result of vicariant events produced by flooding processes and subsequent isolation of ammonite associations during the middle Oxfordian in areas of Mexico-Caribbean and Chile.
This paper aims to analyze the Oxfordian-Kimmeridgian ammonite association from Mexico, Cuba, Chile, and Argentina, to establish their faunistic affinities and define the factors that determine the paleogeographic distribution and faunal exchanges among the areas mentioned above.
For this purpose, we consider the results of the latest detailed systematic-paleontological studies, such as those carried out by López-Palomino (2002, 2013), Villaseñor et al. (2004), López-Palomino et al. (2006), and Olóriz et al. (2008); whose studied material comes from 3 stratigraphic sections (Tamán, San Luis Potosí State, Tenango and La Mesa-El Pescado, Hidalgo State). In addition, we also include previously documented information (Burckhardt, 1930; Cantú-Chapa, 1969, 1971, 1984; Olóriz et al., 2010).
To analyze beta diversity, the Jaccard similarity coefficient was used (as a quantitative similarity index). This index expresses the degree to which two samples are similar based on the species present in them (to calculate the dissimilarity index 1- similarity). In the Jaccard similarity coefficient, values close to zero indicate that no species are shared between sites, while values close to one indicate that the sites are more similar (Moreno, 2001). This analysis was performed using the software PAST 4.10 (Hammer et al., 2001).
2 Study area and geological setting
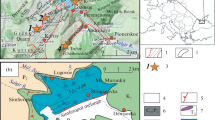
The material documented here comes from three stratigraphic sections measured and sampled in the Huasteca region, central-eastern Mexico (Fig. 1). The study area is located in the Sierra Madre Oriental Physiographic Province, which has an NNW-SSE orientation, reaching heights in the area from 2,000 to 3,000 m above sea level; and it is located in the Geologic Province “Cinturón de Pliegues y Cabalgaduras”, which in accordance with Suter (1990) and Ortega-Gutiérrez et al. (1992), was formed by compression due to the Laramide deformation of the Cordilleran orogeny.
The first stratigraphic section corresponds to the Tamán section (TAM-1) which is located in the neighborhoods of Tamán town, on the west flank of the Moctezuma River, approximately 7 km to the south of Tamazunchale, southeastern San Luis Potosí (21° 13’ 35’’ N–21° 13’ 48’’ N; 98° 52’ 34’’ W–98° 52’ 48’’ W). The second section is the Tenango section (TEN-1), which is located in the surroundings of Tenango town, about 7.6 km to the southeastern of Chapulhuacán, northern Hidalgo State, on the dirt road that connects Tenango and the Cahuasas Ranch, (21° 9’ 51” N − 21° 9’ 56” N; 98° 52’ 40” W–98° 52’ 43” W). The third measured section corresponds to the La Mesa-El Pescado section (LM-EP), located around 3 km to the northeast of Chapulhuacán, Hidalgo State, on the road that connects the towns La Mesa and El Pescado (21° 13’ 35” N–21° 13’ 48” N; 98° 52’ 34” W–98° 52’ 48” W).

Geographical location and geological map of the study area.
Previously, Suter (1990) carried out the geological survey of the study area corresponding to the Geological Chart of Tamazunchale, scale 1:100,000, which covers part of the Sierra Madre Oriental.
3 Litostratigraphy
The measured sections belong to the Santiago Formation, originally studied by Reyes (1964) and formally defined by Cantú-Chapa (1969, p. 5) as a sequence conformed by dark gray calcareous shales intercalated with clay-limestone in layers up to 40 cm thick, brown to reddish-brown on weathered surfaces, with diagonal crossing to the stratification plane and interspersed calcareous nodules.
3.1 Tamán section (TAM-1, Fig. 2a)
This section corresponds to the type locality of the Santiago Formation. At this site, the lower contact does not crop out while the upper contact conformably underlies the Tamán Formation. This section is about 186 m thick, although it is worth mentioning that structurally this outcrop exhibits changes in the orientation of the layers, which could affect the measurement of its real thickness. From base to top, the Santiago Formation comprises approximately 1.90 m of calcareous levels followed by a 31 m thick package of crumbly dark-gray siltstone in fresh surfaces and whitish-brown in weathering surfaces, in beds from 20 to 50 cm, with limestone nodules ranging from 40 to 50 cm containing well-preserved ammonite fauna. Interbedding with siltstone is possible to observe gray limestone layers 30 cm thick as maximum. Above this package, 69 m covered are observed, within which there is an interbedded level, approximately 37 m (level 57 in the stratigraphic section). In the middle part of the section, 26 m thick, a rhythmic intercalation of limestone and siltstone is observed, in beds from 0.20 to 0.30 cm, and 10 cm respectively.

Stratigraphic columns of the Santiago Formation in central-eastern Mexico with ammonite taxa distribution. (a) Tamán section, discontinuous lines at the top indicate repetition of the sequence, (b) Tenango section and (c) La Mesa-El Pescado section. Doted lines showing possible correlation between profiles
Finally, the upper part of this profile corresponds to 129 m, which is mainly composed of dark-brown slightly calcareous siltstone. The structural change is also present, so the faunal position was useful to identify the stratification planes. In some levels, 20 to 30 cm in diameter limestone nodules are observed. Likewise, thin layers of limestone like those above mentioned are sporadically observed. At the top, there are some levels of siltstone with nodular appearance.
3.2 Tenango section (TEN-1, Fig. 2b)
In this section, locally, the Santiago Formation overlies the Tepexic Formation in a concordantly abrupt contact, underlying conformably and transitionally the Tamán Formation. The contact between the Santiago and the Tepexic formation is marked at the base of the former by a thin conglomeratic horizon about 5 cm thick (level 18), constituted mainly by quartz grains and oyster shells; and underlies the Tamán Formation in a concordant and transitional manner. This section comprises a 42 m thick package conformed by gray limestone 2 m interbedded with gray siltstone, whose thickness ranges from 10 to 45 cm. Above these are about 6.5 m of argillaceous limestone interbedded with 5 to 45 cm siltstone strata, as well as some sandstone beds. Later there are 5 m of sheeted siltstone 10 to 18 cm thick with some strata of gray limestone. Finally, 29.5 m are represented by gray limestone with thicknesses that vary between 10 and 70 cm interbedded with thin layers of gray siltstone 5 to 10 cm thick.
3.3 La Mesa-El Pescado section (LM-EP, Fig. 2c)
At this site, the Santiago Formation is similar to TEN-1, overlying the Tepexic Formation with a conglomeratic contact between them (level 18). At its base there are approximately 3 m of gray limestone in beds ranging from 20 to 52 cm; while the rest of the stratigraphic section is constituted by an intercalation of gray limestone and siltstone in layers from 11 to 50 cm, and lesser extent thin siltstones beds in a range of 5 to 14 cm.
4 Comments on Oxfordian ammonites of Mexico
Oxfordian Mexican ammonites have been studied since 1912 by Burckhardt, who recognized and described a diverse ammonite fauna from San Pedro del Gallo, Durango, proposing the differentiation of two layers: “Couches inférieures” and “Couches supérieures”, which were assigned to the upper Oxfordian. Later, the same author (Burckhardt, 1930) delimitates more precisely the age of these layers naming them “Couches a Persiphinctes gr. plicatilis” (lower layers), and Couches à Ochetoceras” (upper layers), proposing a middle Oxfordian age, Transversarium Zone for the former; and a late Oxfordian age, Bimammatum Zone (= Hypselum Zone, according to Wierzbowski et al., 2023) for the latter.
Between 1930 and 1990, the knowledge of the Oxfordian ammonite fauna, both in northern and central-eastern Mexico, increased due to several studies carried out during those years (Imlay, 1939, 1980, 1984; Cantú-Chapa, 1969, 1971, 1984; Contreras et al., 1988). Among these studies, only Imlay (1939) recognized middle Oxfordian fauna, while the others identified late Oxfordian fauna but without specifying its age within this substage.
After that, based on bibliographic data, Callomon (in Hillebrandt et al., 1992) compared the Mexican ammonite faunas documented by Burckhardt, Imlay, and Cantú-Chapa, with European ammonite faunas. This led the author to recognize the middle Oxfordian, Plicatilis Zone (not extreme base) and Transversarium Zone (not the higher part), as well as the upper Oxfordian (Bimammatum Zone = Hypselum Zone, according to Wierzbowski et al., 2023). After him, also based on bibliographic data and some field observations, Myczyński et al. (1998) reinterpreted the Oxfordian age for the ammonite fauna of Mexico, determining the presence of the middle Oxfordian (Plicatilis Zone)-late Oxfordian (Planula Zone possibly, Planula subzone = lower most Kimmeridgian, according to Wierzbowski et al., 2023).
Particularly in the study area, López-Palomino (2002) established an age from the middle Oxfordian (Plicatilis Zone, Antecedens Subzone) to the late Oxfordian (Bimammatum Zone, probable Hypselum Subzone) for the Tamán section (TAM-1). However, a subsequent study carried out by Villaseñor et al. (2012; Fig. 12, part 2) suggests that the specimens referred to Praeataxioceras correspond to a “Subdiscosphinctes” sp. 1 gr. carribeanus, which allows to determine that the upper part of the Santiago Formation in the Tamán Section corresponds to the Bifurcatus Zone (lower most Kimmeridgian, according to Wierzbowski et al., 2023).
For the Tenango Section (TEN-1), according to López-Palomino (2013), the interpreted age represents the middle Oxfordian (Transversarium Zone)-late Oxfordian (Bimammatum Zone); however, based on the most recent interpretation by Wierzbowski et al., (2023), the age of the Tenango section would correspond to the middle Oxfordian (Transversarium Zone)-late Oxfordian (Hypselum Zone)/early Kimmeridgian (Bimmamatum Zone?).
Finally in the La Mesa-El Pescado section (LM-EP), only the middle Oxfordian (Transversarium Zone) is represented.
According to the mentioned before, is certainly established the presence of the middle Oxfordian (Plicatilis Zone)- late Oxfordian (Hypselum Zone)/early Kimmeridgian (Bimammatum Zone?) ammonites in Mexico.
5 Ammonite association
5.1 Mexican associations
The Oxfordian ammonite fauna in the study area is represented in each of the sections (some well-preserved ammonites were selected and illustrated in Figs. 3 and 4) as follows:

(a) Ochetoceras (Cubaochetoceras) burckhardti, IGM-6096, left side view showing lateral groove (Olóriz et al., 2008, Fig. 5, a-g); (b) Ochetoceras (Cubaochetoceras) submexicanum, IGM-13,358, right side view showing lateral groove; (c) Perisphinctes (Dichotomosphinctes) durangensis, IGM-6093, left side view (Olóriz et al., 2008, Fig. 4, g, i-j); (d) Euaspidoceras (Euaspidoceras) aff. costatum, IGM-6110, right side view with small periumbilical bullae (Olóriz et al., 2008, Fig. 6, d-e); (e) Gregoryceras sp., IGM-6090, left side view (Villaseñor et al., 2004, Figs. 1, 2 and 3); (f) Mirosphinctes sp., IGM-13,359, right side view of incomplete specimen; (g) Vinalesphinctes tamanensis, IGM-6170, left side view with suture line preserved (López-Palomino et al., 2006, Fig. 5, a-b, d); (h) “Discosphinctes” acandai, IGM-13,360, right side view showing fine ribbing

(a) Euaspidoceras occonellae, IGM-6553, right side view (Villaseñor et al., 2012, pl. I, Fig. 6); (b) Perisphinctes (Cubasphinctes) sp. cf. cubanensis, IGM-13,361, right side view; (c) Caracoliceras aff. dunkeri, IGM-13,362, left side view; (d) Passendorferia (Passendorferia) sp. gr. tenuis-uptonioides, IGM-13,363, left side view of inner-middle whorls; (e) Sequeirosia (Gemmellarites) sp. cf. trichoplocoides, IGM-13,364, left side view showing incrusted brachiopods; (f) Vinalesphinctes tenangensis, IGM-6175, right side view (López-Palomino et al., 2006, Fig. 9, a-d)
5.1.1 Tamán section (Fig. 2a)
Ochetoceras (Ochetoceras) sp., O. (Cubaochetoceras) burckhardtii (O’CONNELL, 1920) (Fig. 3a), Ochetoceras (Cubaochetoceras) sp. cf. mexicanum (BURCKHARDT, 1912), Ochetoceras (Cubaochetoceras) sp. cf. pedroanum (BURCKHARDT, 1912), Perisphinctes (Perisphinctes) sp., Perisphinctes (Otosphinctes) sp. group. patturatensis (DE LORIOL, 1901), Perisphinctes (Dichotomosphinctes) durangensis (BURCKHARDT, 1912) (Fig. 3c), Perisphinctes cf. plicatilis (DE RIAZ in BURKHARDT, 1912), Passendorferiinae gen. et sp. indet., Euaspidoceras (Euaspidoceras) aff. costatum (DORN, 1930) (Fig. 3d), Euaspidoceras sp., Subdiscosphinctes (Subdiscosphinctes) sp. cf. aeneas (GEMMELLARO, 1875), “Discosphinctes” acandai (JUDOLEY & FURRAZOLA-BERMÚDEZ, 1968) (Fig. 3h) (Olóriz et al., 2008), Gregoryceras sp. (Fig. 3e) (Villaseñor et al., 2002, 2004), Vinalesphinctes tamanensis LÓPEZ-PALOMINO, 2002 (Fig. 3g) (López-Palomino et al., 2006), Ochetoceras (Cubaochetoceras) submexicanum (JUDOLEY & FURRAZOLA-BERMÚDEZ, 1968) (Fig. 3b), Neoprionoceras sp., Mirosphinctes sp. (Fig. 3f), Perisphinctes (Dichotomosphinctes) sp. cf. chloroolithicus (NIKITIN in BURCKHARDT, 1912), Perisphinctes (Cubasphinctes) jaworskii JUDOLEY & FURRAZOLA-BERMÚDEZ, 1968, cf. Larcheria sp., Subdiscosphinctes (Subdiscosphincte) sp. cf. carribeanus (JAWORSKI, 1940), Subdiscosphictes (Subdiscosphinctes) moctezumensis LÓPEZ-PALOMINO, 2002, Subdiscosphictes (Subdiscosphinctes) spp., Sequeirosia (Gemmellarites) trichoplocoides (BURCKHARDT, 1912), Euaspidoceras (Euaspidoceras) spp. (López-Palomino, 2002), Euaspidoceras oconnellae (SÁNCHEZ-ROIG, 1920) (Fig. 4a), “Subdiscosphinctes” sp. 1 gr. carribeanus (JAWORSKI, 1940) (Villaseñor et al., 2012).
5.1.2 Tenango section (Fig. 2b)
Phylloceras sp., Ochetoceras (Cubaochetoceras) mexicanum, Ochetoceras (Cubaohetoceras) spp., Perisphinctes (Cubasphinctes) sp. cf. cubanensis O’CONNELL, 1920 (Fig. 4b), Caracoliceras aff. dunkeri STEINMANN, 1881 (Fig. 4c), Perisphinctes (Dichotomosphinctes) sp., Perisphinctes (Cubasphinctes) sp., Neumannia sp. cf. cyrilli NEUMANN, 1907, Passendorferia (Passendorferia) sp. gr. tenuis ENAY, 1966- uptonioides ENAY, 1966 (Fig. 4d), Subnebrodites sp., Euaspidoceras (Euaspidoceras) sp. cf. oconnellae, Physodoceras sp. cf. wolfi (NEUMAYR, 1873) (López-Palomino, 2013), Vinalesphinctes tenangensis LÓPEZ-PALOMINO et al., 2006 (Fig. 4f) (López-Palomino et al., 2006).
5.1.3 La Mesa-El Pescado section (Fig. 2c)
Perisphinctes (Otosphinctes) sp., Sequeirosia (Gemmellarites) sp. cf. trichoplocoides (BURCKHARDT, 1912) (Fig. 4e), Sequeirosia sp. (López-Palomino, 2013).
All shared records will be analyzed particularly with the fauna associations for Cuba, Chile and Argentina, presented below.
5.2 Cuban associations
Oxfordian ammonites from Cuba have been studied by Sánchez-Roig (1920), O’Connell (1920), Jaworski (1940), Imlay (1942), and Judoley and Furrazola-Bermúdez (1968). However, despite the important data these studies have provided, biostratigraphic and paleobiogeographic interpretations were only carried out until the investigations conducted by Myczyński (1976), Wierzbowski (1976), and Myczyński et al. (1998). Among the taxa recognized in Cuba related to Mexico, it is possible to find representatives of the genera Phylloceras SUESS, 1865 (Imlay, 1942), Ochetoceras HAUG, 1885 and Cubaochetoceras SÁNCHEZ-ROIG, 1951 (O’Connell, 1920; Sánchez-Roig, 1920; Jaworski, 1940; Judoley & Furrazola-Bermúdez, 1968), Neoprionoceras SPATH, 1928 (Jaworski, 1940), Gemmellarites MELÉNDEZ, 1989 (Sánchez-Roig, 1920), Mirosphinctes SCHINDEWOLF, 1926 (Myczyński, 1976), Dichotomosphinctes (Sánchez-Roig, 1920; Judoley & Furrazola-Bermúdez, 1968), Cubasphinctes (O’Connell, 1920; Judoley & Furrazola-Bermúdez, 1968), Euaspidoceras SPATH, 1931 (Judoley & Furrazola-Bermúdez, 1968; Myczyński, 1976; Wierzbowski, 1976), Vinalesphinctes (Jaworski, 1940; Judoley & Furrazola-Bermúdez, 1968; Wierzbowski, 1976), and Physodoceras HYATT, 1900 (Imlay, 1942).
5.3 Chilean associations
Research on Oxfordian ammonites from Chile dates since the contributions made by Steinmann (1881) and Burckhardt (1903). Subsequent and relevant studies were carried out by Leanza (1947), Harrington (1961 in García, 1967), Tobar et al. (1968), Hillebrandt (1970), Gröschke and Hillebrandt (1985), and Meléndez and Myczyński (1987). In particular, the work of Gygi and Hillerandt (1991) provided valuable data about the Oxfordian Chilean ammonites and their correlation with other American faunas. In addition, Hillebrandt and Gröschke (1995) and Hillebrandt et al. (2000) provide information that greatly contributes to biostratigraphic and paleobiogeographic correlations. The ammonites recognized in Chile with affinities to Mexico are the genera Ochetoceras, Cubaochetoceras, Perisphinctes WAAGEN, 1869, Larcheria TINTANT, 1961, Gregoryceras, Euaspidoceras, Vinalesphinctes, Subdiscosphinctes MALINOWSKA, 1972 (Gygi & Hillebrandt, 1991), Mirosphinctes (Gygi & Hillebrandt, 1991; Hillebrandt & Gröschke, 1995), Otosphinctes BUCKMAN, 1926 (Hillebrandt & Gröschke, 1995), and Caracoliceras HILLEBRANDT et al., 2000 (Hillebrandt et al., 2000).
5.4 Argentinian associations
The first contribution to the knowledge of ammonites from Argentina was made by Gottsche (1878). After him, the study of this taxon and its Oxfordian biostratigraphy is mainly restricted to the works performed by Stipanicic (1951), Stipanicic et al. (1975), and Riccardi (1984). Some later authors reanalyzed the data of the aforementioned authors providing new information, among which are Parent (1998, 2006), Parent et al. (2006), Palma et al. (2012), and Parent and Garrido (2015). The recognized ammonites in Argentina with affinities to Mexico are Passendorferia BROCHWICZ-LEWINSKI, 1973 (Parent, 1998, 2006), Mirosphinctes, Otosphinctes (Parent, 1998; Palma et al., 2012), Dichotomosphinctes (= Cubasphinctes durangensis, Parent, 2006), Perisphinctes (Parent, 1998; Parent & Garrido, 2015), Cubasphinctes (Parent, 2006; Parent & Garrido, 2015), Subdiscosphinctes (Parent, 2006; Palma et al., 2012), Euaspidoceras (Parent, 1998, 2006; Parent & Garrido, 2015) and Vinalesphinctes (Parent, 2006; Parent et al., 2006; Parent & Garrido, 2015).
6 Discussion
The paleobiogeographic relationships between the ammonite faunas of Mexico and other regions of America and Europe have been difficult to establish, mainly due to insufficient knowledge of the ammonite fauna of the highest part of the middle and upper Oxfordian (lower most Kimmeridgian, according to Wierzbowski et al., 2023) of Mexico. However, with the information discussed here, it is possible to recognize a certain affinity between the Oxfordian Mexican ammonite faunas with the Cuban, Chilean, Argentinian, and Tethys associations.
The existence of marine communication between eastern and western Tethys with the Pacific during the Late Triassic to the Late Jurassic is widely documented, through the so-called Hispanic Corridor (Damborenea, 2000; Arias, 2007, 2009; Stevens, 2007, among others).
Here, we propose that during the Plicatilis-Transversarium Chrons a wide communication with the Tethys was established, as suggests the presence of the genus Passenderforia, whose origin has been proposed in the southern margin of the Tethys, and later widely distributed throughout the entire Tethys (Meléndez et al., 2009). Additionally, the genera Neumannia GLOWNIAK, 2002, Sequeirosia (Gemmellarites), Perisphinctes (Otosphinctes), among others, clearly show Tethys affinities, mainly with the Mediterranean Province, during the Oxfordian.
A remarkable case corresponds to the genus Gregoryceras, which is widely used in global correlations due to its clearly Tethys affinity (Gygi & Hillebrandt, 1991; Villaseñor et al., 2004; Olóriz et al., 2010). In particular, Gygi and Hillebrandt (1991) for Chile, Young & Olóriz (1993) for the southeastern United States, and Villaseñor et al. (2002, 2004) for Mexico, reported the existence of forms with similar morphologies to those from the Tethys during the Plicatilis Chron (middle Oxfordian).
On the other hand, for the Bifurcatus Chron in Mexico, the presence of the genus Vinalesphinctes suggests a paleogeographic affinity with Cuba, Chile, and Argentina; the occurrence of Perisphinctes (Cubasphinctes)-Vinalesphinctes could imply affinities with Cuba and Argentina; while Caracoliceras only with Chile. Moreover, fauna with Tethyas affinity was recognized (Neoprionoceras, Subdiscosphinctes, Ochetoceras, and Euaspidoceras) (Fig. 5).

Oxfordian (Bifurcatus Chron) paleogeographic map modified from Smith et al. (1995) showing the distribution of the genera Vinalesphictes, Cubasphinctes and Caracoliceras
The presence of this fauna in these regions is explained by the existence of a connection between the western Tethys and the eastern Pacific (López-Palomino et al., 2006). In particular, the occurrence of Vinalesphinctes, with a biochronostratigraphic distribution included in the Bifurcatus Chron and restricted paleogeographically, gives it an undoubted endemic character. Furthermore, the presence of the genus Cubasphinctes, as well as to the record in Mexico of the genus Caracoliceras, which had been previously documented in Chile, agrees with this interpretation. All the above is congruent with the proposal of Myczyński (1994) that during the middle Oxfordian, the northwest part of the proto-Caribbean Basin was probably connected with the eastern portion of Mexico, as well as there was a possible connection with the fauna from the Andean Province.
Finally, during the Hypselum Chron (late Oxfordian)/Bimammatum Chron (early Kimmeridgian) the presence of Physodoceras and Subnebrodites, taxa with a clearly Tethys affinity, support the existence of a marine connection at least since the middle Oxfordian to early Kimmeridgian between both areas. Also this connection is confirmed by the presence througthout the studied succesions of Passendorferia, Ochetoceras, and Euaspidoceras genera typically considered as Tethys origin.
The data presented for mexican and carribean faunas, allows us to confirm the existance of communication with the Tethys via the Hispanic Corridor since particularly in the Plicatilis Chron and continued, perhaps intermittently, until the Cretaceous.
In general terms, the results of the application of the Jaccard index show that for the mid-late Oxfordian, at generic level Mexico presents greater affinity with the Tethys and in second place with Cuba (Fig. 6a, e); the dendrogram show that the Tethys and Mexico areas are united into one group, being Cuba the closest area to them; whiles Chile is the area more similar to the aforementioned areas, and Argentina the more different (Fig. 6c). On the other hand, at specific level, the greatest similarity is with Cuba and in second place with the Tethys (Fig. 6b, f). The dendrogram derived from the Jaccard analysis shows a group with Argentina and Tethys as sister areas, with Chile close to them, and Cuba and Mexico in a separate clade.
The ammonites records presented as well as the similarity analysis carried out allows us to confirm the existence of communication between Mexican, Caribbean and Tethys faunas via the Hispanic Corridor since Plicatilis Chron and continued, perhaps intermittently, until the Cretaceous.

Similarities between Mexico, Cuba, Chile, Argentina, and Tethys regions. (a, b) Values of the Jaccard similarity index for ammonites (a, genera, b, species); (c, d) Cluster dendrograms (UPGMA) derived from the Jaccard index constructed from the presence/absence data (c, genera Cophenetic correlation: 0.8302; d, species Cophenetic correlation: 0.9189); (e, f) pie charts showing similarity values, genera and species, respectively
7 Conclussions
-
1)
The record of genera and/or species that have been reported in Mexico and in other regions such as Cuba, Chile, and Argentina clearly evidence the connection of these areas during the mid-late Oxfordian and would support the existence of the so-called Hispanic Corridor at this time.
-
2)
The faunal affinity with Chile and Argentina is interpreted as evidence of a connection with these epicontinental areas favoring dispersion by marine currents.
-
3)
The interpretation of Olóriz (1987) is confirmed, regarding that the Mexican platforms should have acted as an area of immigration and later evolution in situ, rather than as a center of immigration and subsequent migration of the ammonite faunas to other more distant areas, such as Europe and the western Pacific. This is evident by the endemism at species level in Ochetoceras, and at genus level in Vinalespinctes.
-
4)
Due to the oldest record in Chile and Mexico of the genera Vinalesphintes and Cubasphinctes, their Pacific origin is corroborated, as it was interpreted by Meléndez & Myczyński (1994).
-
5)
The interpretation of Myczyński (1994) is reinforced that during the mid-late Oxfordian the northwest part of the Proto-Caribbean Basin was probably connected to the eastern part of Mexico and a possible connection with the fauna of the Andean Province.
-
6)
The presence of the typically Tethyan genera Ochetoceras, Euaspidoceras, and Subdiscosphinctes, from the middle to late Oxfordian, corroborates the existence of the connection between the Tethys and the Pacific throughout the Oxfordian.
Data availability
The datasets generated and/or analyzed during the current study are not publicly available because it involves participant’s personal information, but are available from the corresponding author on reasonable request.
References
Arias, C. (2007). Pliensbachian-Toarcian ostracod biogeography in NW Europe: Evidence for water mass structure evolution. Palaeogeography Palaeoclimatology Palaeoecology, 251, 398–421. https://doi.org/10.1016/j.palaeo.2007.04.014
Arias, C. (2009). Extinction pattern of marine Ostracoda across the Pliensbachian-Toarcian boundary in the Cordillera Ibérica, NE Spain: Causes and consequences. Geobios, 42, 1–15. https://doi.org/10.1016/j.geobios.2008.09.004
Brochwicz-Lewinski, W. (1973). Some remarks on the origin of the subfamily Idoceratinae SPATH, 1934 (Perisphinctidae, Ammonoidea). Acta Paleontologica Polonica, 18(3), 299–320.
Buckman, S. (1909–1930). Yorkshire type ammonites. Wheldon & Wesley (Eds.), London. 1–7, 790.
Buitrón, B. E. (1984). Late Jurassic bivalves and gastropods from northern Zacatecas, Mexico, and their biogeographic significance. In G. E. G. Westermann (Ed.), Jurassic-Cretaceous biochronology and paleogeography of North America. Geological Association of Canada, Special Paper, 27, 89–98.
Burckhardt, C. (1912). Faunes Jurassiques et Cretaciques de San Pedro del Gallo. Boletín del Instituto Geológico de México, 29, 264.
Burckhardt, C. (1930). Ètude syntétique sur le Mésozoïque mexicain. Mémoires de la Société Paléontologique Suisse, 49–50, 280.
Burckhardt, C. (1903). Beitrage zur Kenntnis der Jura-und Kreideformation der Cordillere. Palaeontographica L. 1–444.
Callomon, J. H. (1992). Upper Jurassic, especially of Mexico. Part IV: Biochronology. Ammonite zones of the Circum-Pacific region. In A. Hillebrandt, P. Smith, G. E. G. Westermann, J. H. Callomon). In G. E. G. Westermann (Eds.), The Jurassic of the Circum-Pacific (pp. 261–273). Cambridge University Press.
Cantú-Chapa, A. (1969). Estratigrafía del Jurásico Medio-Superior del subsuelo de Poza Rica, Ver. (Área de Soledad-Miqueta). Revista del Instituto Mexicano del Petróleo, 1(1), 3–9.
Cantú-Chapa, A. (1971). La Serie Huasteca (Jurásico Medio-Superior) del Centro-Este de México. Revista del Instituto Mexicano del Petróleo, 3(2), 17–40.
Cantú-Chapa, A. (2001). Mexico as the western margin of Pangea based on biogeographic evidence from the Permian to the Lower Jurassic. In C. Bartolini, R. T. Bubbler, & A. Cantú-Chapa (Eds.), The western Gulf of Mexico Basin: Tectonics, sedimentary basins, and petroleum systems. America Association of Petroleum Geologist Memoir, 75, 1–27. https://doi.org/10.1306/m75768c1
Cantú-Chapa, A. (1984). El Jurásico Superior de Tamán, San Luis Potosí, Este de México. In M. C. Perrilliat (Ed.), Memoria Tercer Congreso Latinoamericano de Paleontología (pp. 207–215). Instituto de Geología, Universidad Nacional Autónoma de México. México, D.F.
Cariou, E. (1973). Ammonites of the Callovian and Oxfordian. In A. Hallam (Ed.), Atlas of Paleobiogeography (pp. 287–295). Elsevier.
Contreras, B., Martínez-Cortés, A., & Gómez-Luna, M. E. (1988). Bioestratigrafía y sedimentología del Jurásico Superior en San Pedro del Gallo, Durango, México. Boletín del Instituto Mexicano del Petróleo, 20(3), 5–27.
Damborenea, S. E. (2000). Hispanic Corridor: its evolution and the biogeography of bivalve molluscs. In R. L. Hall & P. L. Smith (Eds.), Paleontology and Biogeography. GeoResearch Forum, 6, 369–380.
De Loriol, P. (1901). Etude sur les Mollusques et brachiopodes de l’Oxfordien superieur et moyen du Jura bernois. Mémoires de la Société Paléntologique Suisse, 1er suppl 1, 23.
Dorn, P. (1930). Die Ammoniten-Fauna des untersten Malm der Frankenalb. Paläeontographica Stuttgart Band, 73–74, 158.
Enay, R. (1966). L’Oxfordien dans la moitié Sud du Jura Francais. 1. Étude Stratigraphique. 2. Étude Paléontologique. Nouvelles Archives du Muséum d’Histoire Naturelle de Lyon, 8, 624
García, F. (1967). Geología del Norte Grande de Chile. Santiago de Chile, Sociedad Geológica de Chile, Simposium sobre el geosinclinal andino 1962, 1–138.
Gemmellaro, G. G. (1875). Sui Fossili della zona con Peltoceras Transversarium QUENSTEDT delle provinci di Palermo e di Trapani. Atti dell’Academi di Scienze e Lettere di Palermo, 4, 113–124.
Glowniak, E. (2002). The ammonites of the family Perisphinctidae from the plicatilis Zone (lower Middle Oxfordian) of the Polish Jura Chain (Central Poland); their taxonomy, phylogeny and biostratigraphy. Acta Geologica Polonica, 52(3), 307–364...
Gottsche, C. (1878). Uber jurassische versteinerungen aus der argentinischen cordillere. PhD dissertation, Universität Münschen, 50.
Gröschke, M., & Hillebrandt, A. (1985). Trias und Jura in der Mittleren Cordillera Domeyko von Chile (23 30’-24c 30’). Neues Jahrbuch für Geologie Und Paläontologie Abhandlungen, 170(2), 129–166. https://doi.org/10.1127/njgpa/170/1985/129
Gygi, R. A., & Hillerandt, A. (1991). Ammonites (mainly Gregoryceras) of the Oxfordian (Late Jurassic) in northern Chile and time-correlation with Europe. Schweizerische Paläontologische Abhandlungen, 113, 137–185.
Schindewolf, O. (1926). Zur Systematik der Perisphincten. Neues Jahrbuch für Mineralogie Geologie und Paläontologie, Band 55(Abt. A), 497–517.
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4(1), 1–9.
Haug, E. (1885). Beiträge zu einer Monographie der Ammonitengattung Harpoceras. Neues Jahrbuch für Mineralogie Geologie und Paläeontologie, Beilage-Band, 3, 585–722.
Hillebrandt, A. (1970). Zur Biostratigraphie und Ammoniten-Fauna des südamerikanischen Jura (insbes Chile). Neues Jahrbuch für Geologie Und Paläontologie Abhandlungen, 139(2), 166–211.
Hillebrandt, A., & Gröschke, M. (1995). Ammoniten aus dem Callovium/Oxfordium-Grenzbereich von Nordchile. Berliner Geowissenschaftliche Abhandlungen A, 169, 1–40.
Hillebrandt, A., Kossler, A., & Gröschke, M. (2000). Caracoliceras, a new Oxfordian (Upper jurassic) ammonites genus from northern Chile. Revue Paléobiologie, 8, 65–81. Special Volume.
Hyatt, A. (1900). Cephalopoda. In Zittel, K.A., Eastman, W. (Eds.), Text-Book of Palaeontology, 1, 502–604.
Imlay, R. W. (1939). Upper Jurassic ammonites from Mexico. Geological Society of America Bulletin, 50(1), 1–78. https://doi.org/10.1130/GSAB-50-1
Imlay, R. W. (1942). Late Jurassic fossils from Cuba and their economic significance. Bulletin of the Geological Society of America, 53(10), 1417–1478. https://doi.org/10.1130/GSAB-53-1417
Imlay, R. W. (1980). Jurassic paleobiogeography of the Conterminous United States in its continental setting. United States Geological Survey Professional Paper, 1062, 1–134. https://doi.org/10.3133/pp1062
Imlay, R. W. (1984). Jurassic ammonite successions in North America and biogeographic implications. In G.E.G. Westermann (Ed.), Jurassic-Cretaceous biochronology and paleontology of North America. Geological Association of Canada, Special Paper 27, 1–12.
Jaworski, E. (1940). Oxford-Ammoniten von Cuba. Neues Jahrbuch für Mineralogie Geologie und Paläeontologie, 83(1), 87–137.
Judoley, C. M., & Furrazola-Bermúdez, G. (1968). Estratigrafía y fauna de Jurásico de Cuba. Instituto Cubano de Recursos Minerales, 126.
Leanza, A. F. (1947). Ammonites coralinos en el Jurásico de Chile. Revista de la Sociedad Geológica de Argentina, 2(4), 285–295.
López-Palomino, I. (2013). Bioestratigrafía y análisis ecoestratigráfico de la Formación Santiago (Jurásico Superior-Oxfordiano) en el centro de México: Implicaciones paleoecológicas y paleobiogeográficas. México, D.F. PhD dissertation, Universidad Nacional Autónoma de México, 186.
López-Palomino, I. (2002). Análisis bioestratigráfico basado en fauna de ammonites del Oxfordiano (Jurásico Superior) en la región de Tamán, San Luis Potosí México. Master’s thesis, Universidad Nacional Autónoma de México, 146.
López-Palomino, I., Villaseñor, A. B., & Olóriz, F. (2006). Primer registro del género Vinalesphinctes (Ammonitina) en el Oxfordiano de México: Significación bioestratigráfica y consideraciones paleobiogeográficas en el Jurásico Superior de América. Revista Mexicana de Ciencias Geológicas, 23(2), 162–183.
Malinowska, L. (1972). The Middle Oxfordian Perisphinctidae of Zawodzie near Czestochowa (Poland). Acta Paleontologica Polonica, 17(2), 167–242.
Meléndez, G. (1989). El Oxfordiense en el sector central de la Cordillera Iberica (Provincias de Zaragoza y Teruel). Zaragoza-Teruel, España, PhD dissertation, Instituto Fernando El Católico, Instituto de Estudios Teruelenses, 418.
Meléndez, G., & Myczyński, R. (1987). Sobre la posición sistemática de los ammonites del Oxfordiense de los Andes Chilenos (Cordillera Domeyko, Chile, Provincia Andina). Geogaceta, 2, 12–14.
Meléndez, G., & Myczyński, R. (1994). On the origin and biogeographic, spread of Vinalesphinctes SPATH, 1931, and related Oxfordian, Perisphinctids from Chile and Cuba. Madrid, España, Sociedad, Española de Paleontología, Comunicaciones de las X Jornadas de Paleontología, 135–137.
Meléndez, G., Atrops, F., Bello, J., Brochwicz-Lewinski, W., D’Arpa, C., Fözy, I., Pérez-Urresti, I., Ramajo, J., & Sequeiros, L. (2009). The Oxfordian ammonite genus Passendorferia Brochwicz-Lewinski and the Tethyan subfamily Passendorferiinae Meléndez: Origin and palaeobiogeography. Volumina Jurassica, 7, 113–134.
Moreno, C. E. (2001). Métodos para medir la biodiversidad. M&T Manuales y Tesis, 1, 1–84.
Myczyński, R. (1976). A new ammonite fauna from the Oxfordian of the Pinar del Río Province, western Cuba. Acta Geologica Polonica, 26(2), 261–297.
Myczyński, R. (1994). Caribbean ammonite assemblages from Upper Jurassic– lower Cretaceous sequences of Cuba. Studia Geologica Polonica, 105, 91–108.
Myczyński, R., & Meléndez, G. (1990). On the current state of progress of the studies on Oxfordian ammonites from Western Cuba. Publicaciones del Seminario de Paleontología de Zaragoza, 2, 185–189.
Myczyński, R., Olóriz, F., & Villaseñor, A. B. (1998). Revised biostratigraphy and correlations of the middle-upper Oxfordian in the Americas (Southern USA, Mexico, Cuba and Northern Chile). Neues Jahrbuch für Mineralogie Geologie Und Paläontologie Abhandlungen, 207(2), 185–206. https://doi.org/10.1127/njgpa/207/1998/185
Neumann, R. (1907). Breitage zur Kenntniss der Kreideformation in Mittel-Peru. Neues Jarbuch für Mineralogie Geologie und Paläontologie Abhandlungen, 24, 69–132.
Neumayr, M. (1873). Die Fauna der Schichten mit Aspidoceras acanthicum. Abhandlungen Der Kaiserlich-Königlichen Geologischen Reichsanstal, 6, 141–257.
O’Connell, M. (1920). The jurassic ammonite fauna of Cuba. Bulletin of the American Museum of Natural History, 42(16), 643–692.
Olóriz, F. (1987). El Significado biogeográfico de las plataformas mexicanas en el Jurásico Superior. Consideraciones sobre un modelo eco-evolutivo. Revista de la Sociedad Mexicana de Paleontología, 1(1), 219–247.
Olóriz, F., Villaseñor, A. B., & López-Palomino, I. (2008). Middle Oxfordian ammonite biostratigraphy of the lower Santiago Formation at Tamán, San Luis Potosí, Mexico-with notes on Mexico-Caribbean ochetoceratins. Revista Mexicana de Ciencias Geológicas, 25(2), 261–283.
Olóriz, F., Villaseñor, A. B., & Grajales-Nishimura, M. (2010). New finding of genus Gregoryceras SPATH 1924 (Ammonitina) from SE Mexico, Veracruz. In L. I. Ruiz-Omeñaca, L. Piñuela, & J. C. García-Ramos (Eds.), Comunicaciones del V Congreso del Jurásico de España (pp. 112–114). Museo del Jurásico de Asturias (MUJA).
Ortega-Gutiérrez, F., Mitre-Salazar, L. M., Roldán-Quintana, J., Aranda-Gómez, J. J., Morán-Zenteno, D., Alaniz-Álvarez, S. A., & Nieto-Samaniego, A. F. (1992). Texto explicativo de la carta geológica de la República Mexicana, scale 1:2000,000. Universidad Nacional Autónoma de México, 5th edition, 1 map with text.
Palma, R. M., Kietzmann, D. A., Martín-Chivelet, J., López-Gómez, J., & Bressan, G. S. (2012). New biostratigraphic data from the Callovian-Oxfordian La Manga f Formation, Neuquén Basin, Argentina: Evidence from an ammonite condensed level. Revue de Paléobiologie, Special 11, 345–356.
Parent, H. (1998). Upper Callovian to upper Oxfordian Ammonite biostratigraphy of the transect Chacay Melehue-Sierra de Reyes, Argentina. Cuadernos de Geología Ibérica, 24, 261–275.
Parent, H. (2006). Oxfordian and Late Callovian ammonite faunas and biostratigraphy of the Neuquén-Mendoza and Tarapacá basins (Jurassic, Ammonoidea, western South-America). Boletín del Instituto de Fisiografía y Geología, 76(1–2), 1–70.
Parent, H., & Garrido, A. C. (2015). The ammonite fauna of the La Manga Formation (Late Callovian-Early Oxfordian) of Vega de la Veranada, Neuquén Basin, Argentina. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 275(2), 163–217. https://doi.org/10.1127/njgpa/2015/0461
Parent, H., Scweigert, G., & Meléndez, G. (2006). Oxfordian perisphinctid ammonites from Chacay Melehué, Argentina. Paläontologische Zeitschrift, 80(4), 307–324. https://doi.org/10.1007/BF02990207
Reyes, E. (1964). El Jurásico Superior del área de Tamán, S. L. P. Petróleos Mexicanos, Technical Report, 508, 36 p. (Unpublished).
Riccardi, A. C. (1984). Las asociaciones de amonitas del Jurásico y Cretácico de la Argentina. San Carlos de Bariloche, Argentina, Servicio Geológico Nacional, Noveno Congreso Geológico 9, Actas 4, 559–595.
Sánchez-Roig, M. (1920). La fauna Jurásica de Viñales. Secretaría Agrícola Comercial del Trabajo, La Habana, Cuba. Boletín Especial, 1–61.
Sánchez-Roig, M. (1951). La fauna Jurásica de Viñales. Anales de la Academia de Ciencias Médicas Físicas y Naturales de La Habana, 89, 46–94.
Smith, L. G., Smith, D. G., & Funnell, B. M. (1995). Atlas of Mesozoic and Cenozoic coastlines. Cambridge University Press.
Spath, L. F. (1924). On the Blake collection of ammonites from Kachh, India. Paläeontographica Indica New Series, 9, 1–29.
Spath, L. F. (1927–1933). Revision of the Jurassic cephalopod faunas of Kachh (Cutch). Memoirs of the Geological Survey of India. Paleontologia Indica, New Series 9, Memoirs 2(1–6), 945.
Steinmann, G. (1881). Zur Kenntnis der Jura- und Kreideformation von Caracoles (Bolivia). Neues Jahrbuch für Mineralogie, Geologie und Palaeontologie [Beilage-Band], 239–301.
Stevens, G. R. (2007). The ammonite genus Harpoceras (Early Jurassic) in New Zealand, New Zealand. Journal of Geology and Geophysics, 50(4), 377–386.
Stipanicic, P. N. (1951). Sobre la presencia del Oxfordiense Superior en el Arroyo de La Manga (Provincia de Mendoza). Revista de la Asociación Geológica Argentina, 6(4), 213–239.
Stipanicic, P. N., Westermann, G. E. G., & Riccardi, A. C. (1975). The Indo-Pacific Ammonite Mayaites in the Oxfordiano of the Southern Andes. Ameghiniana Revista de la Asociación Paleontológica Argentina, 12(4), 281–305.
Suess, E. (1865). Über Ammoniten. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften in Wien. Mathematisch-Naturwissenschaftliche Klasse, 52(1), 71–89.
Suter, M. (1990). Geología de la hoja de Tamazunchale, Estado de Hidalgo, Querétaro y San Luis Potosí, Scale 1:100,000. Universidad Nacional Autónoma de México, Instituto de Geología, México, D.F., map with text, 55.
Tintan, H. (1961). Etudes sur les ammonites de I’Oxfordien Supérieur de Bourgogne. I. Les genres Platisphinctes nov. et Larcheria nov. Bulletin des Science de Bourgogne, 19, 109–1
Tobar, B. A., Salas, Y. I., & Kast, R. F. (1968). Cuadrángulos Camaraca y Azapa - Provincia de Tarapaca. Carta Geológica de Chile, Scale 1:50,000. Instituto de Investigaciones Geológicas Chile, Geological Sheet 19 y 20, 1–20.
Villaseñor, A. B., Olóriz, F., & López-Palomino, I. (2002). The finding of the ammonite genus Gregoryceras (Ammonitina) from Mexico. In L. Martire (Ed.), 6th International Symposium on the Jurassic System, Palermo. Abstrac and Program, p. 191.
Villaseñor, A. B., Olóriz, F., & López-Palomino, I. (2004). Inner whorls of Gregoryceras (Ammonitina, Peltoceratinae) as the first occurrence of the genus in Mexico. Rivista Italiana di paleontología e Stratigrafia, 110(1), 249–254. https://doi.org/10.13130/2039-4942/6295
Villaseñor, A. B., González-León, C., Lawton, T. F., & Aberhan, M. (2005). Upper Jurassic ammonites and bivalves from the Cucurpe Formation, Sonora (Mexico). Revista Mexicana de Ciencias Geológicas, 22(1), 65–87
Villaseñor, A. B., Olóriz, F., López-Palomino, I., & López-Caballero, I. (2012). Updated ammonite biostratigraphy from Upper Jurassic deposits in Mexico. Revue de Paléobiologie, Special 11, 249–267.
Waagen, W. (1869). Die Formenreihe des ammonites subradiatus. Geographisch Paläontologie Beitrage, Band 2(Helf 2), 181–256.
Wierzbowski, A. (1976). Oxfordian ammonites of the Pinar del Rio Province (Western Cuba); their revision and stratigraphical significance. Acta Geologica Polonica, 26(2), 137–260.
Wierzbowski, A., Barski, M., Coe, A. L., Hounslow, M. W., Matyja, B. A., Price, G. D., Wierzbowski, H., Wright, J. K., Atrops, F., Grabowski, J., Mattioli, E., Morton, N., Ogg, J. G., Olóriz, F., Page, K., Parent, H., Przybylski, P., Schweigert, G., & Villaseñor, A. B. (2023). The global stratotype section and point (GSSP) for the base of the Kimmeridgian Stage (Jurassic System), at Flodigarry, Staffin Bay, Isle of Skye, Scotland, UK. Episodes, 46(2), 281–307.
Young, K., Olóriz, F. Ammonites from the Smackover Limestone, Cotton Valley Field, Webster Parish, Lousiana, U.S.A. In S. Elmi, Ch. Mangold,Y. Alméras. 3ème Symposium International Céphalopodes Actuels et Fossiles (Symposium, F., & Roman (1993). Geobios Mémoire Spécial, 15, 401–40
Acknowledgements
The authors thank the Mexican Geological Survey and the Institute of Geology of the National Autonomous University of Mexico, for all the support received. Also, to Eng. Ana Luz Montoya-Vazquez (Geomatics, Mexican Geological Survey) who drafted the geological map. Thanks to M. Reolid, F.J. Rodríguez-Tovar, and J.A. Pérez-Claros for the invitation to participate in this special volume. Also, many thanks to two anonymous reviewers for suggestions that improved the original version. Finally, we express our gratitude to Federico Olóriz for his unconditional disposition in the field and office long days of work and for sharing his knowledge of the Jurassic.
Funding
This research was supported by the Mexican Geological Survey and the Institute of Geology of the National Autonomous University of Mexico.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
López-Palomino, I., Villaseñor, A.B. & Palma-Ramírez, A. Ammonites from the Late Jurassic (Oxfordian- early Kimmeridgian) of central-eastern Mexico and their paleobiogeographic relationship with Cuba, Chile, and Argentina. J Iber Geol (2024). https://doi.org/10.1007/s41513-024-00257-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s41513-024-00257-7