
Abstract
The El Castellar Formation (upper Hauterivian-lowermost Barremian, Lower Cretaceous) in the southwest of the Maestrat Basin (Spain) has yielded diverse dinosaur remains, mostly fragmentary bones. This study describes, morphometrically analyses, and compares several isolated postcranial fossils of large ornithopods from three new different sites in the municipality of Cabra de Mora (Peñagolosa sub-basin, Teruel Province). The fossils are attributed to styracosternan ornithopods, and some of them are related to the species Iguanodon galvensis, representing the oldest known record related to this taxon in the Iberian Peninsula. Moreover, this study verifies the hypothesis of the coexistence of at least two different styracosternans—a medium-sized form and a large-sized one—during the late Hauterivian-early Barremian in the Peñagolosa sub-basin. In addition, we formally describe the first dinosaur tracksite of the El Castellar Formation in this sub-basin. All ichnites are preserved as natural casts, and their trackmakers are likely to have been related to large styracosternans. Hence, the evidence provided here by both bones and tracks supports the idea that large ornithopods were the predominant dinosaurs in the wetlands of an extensive coastal plain.
Resumen
La Formación El Castellar (Hauteriviense superior-Barremiense basal) en el suroeste de la Cuenca del Maestrazgo (España) ha proporcionado diversos fósiles de dinosaurios bastante fragmentarios. Aquí describimos, analizamos morfométricamente y comparamos varios restos postcraneales aislados de grandes ornitópodos procedentes de tres yacimientos en el municipio de Cabra de Mora (subcuenca de Peñagolosa, provincia de Teruel). Los fósiles son atribuidos sistemáticamente a grandes ornitópodos estiracosternos, y algunos de ellos son relacionados con Iguanodon galvensis, lo que implica que estos restos pueden representar el registro más antiguo relacionado con dicho taxón en la Península Ibérica. Además, este estudio comprueba la hipótesis de la presencia de al menos dos formas de estiracosternos durante el Hauteriviense tardío-Barremiense temprano en la subcuenca de Peñagolosa, uno de talla grande y otro de talla mediana. Adicionalmente, se describe formalmente el primer yacimiento de icnitas de dinosaurios en la Formación El Castellar de esta subcuenca. Todas las icnitas están preservadas como hiporrelieves convexos y sus productores podrían estar relacionados también con grandes estiracosternos. Por lo tanto, la evidencia aquí proporcionada tanto por los restos directos como por las icnitas respaldan la idea de que los grandes ornitópodos eran los dinosaurios predominantes en los diferentes humedales desarrollados en una extensa llanura costera.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The Early Cretaceous sediments on the Iberian Peninsula have yielded numerous dinosaur fossil finds (e.g. Alcalá et al., 2018; Pereda-Suberbiola et al., 2012). In relation to the Hauterivian-Barremian section, the saurischians are represented by brachiosaurid and rebbachisaurid sauropods (e.g. Fernández-Baldor et al., 2011; Royo-Torres et al., 2017), tetanuran theropods such as spinosaurids (e.g. Isasmendi et al., 2020; Malafaia et al., 2020a, 2020b), carcharodontosaurids (Gasca et al., 2014a; Ortega et al., 2010), and coelurosaurs (e.g. Torrente et al., 2018) such as ornithomimosaurs (Cuesta et al., 2021; Pérez-Moreno et al., 1994). The ornithischian dinosaurs are mainly ankylosaurs (Gasulla et al., 2011; Pereda-Suberbiola et al., 2007) and ornithopods. This latter group of dinosaurs is the most abundant and highly diverse. They are represented by small basal ornithopods related to Hypsilophodon (e.g. Ruiz-Omeñaca et al., 2012), rhabdodontomorphans (Dieudonné et al., 2016), dryosaurids (e.g. Galton, 2009), and medium to large-sized non-hadrosaurid styracosternans (e.g. Fuentes-Vidarte et al., 2016; Gasca et al., 2014b; Gasulla et al., 2014, 2015; Verdú et al., 2015, 2019). The diversity of Iberian styracosternans of the Hauterivian-Barremian range includes Magnamanus soriaensis, in the upper Hauterivian-lower Barremian of Soria Province (Fuentes-Vidarte et al., 2016); Iguanodon galvensis, in the lower Barremian of Teruel Province (Verdú et al., 2015, 2018, 2020); Portellsaurus sosbaynati, in the lower Barremian of Castellón Province (Santos-Cubedo et al., 2021); Iguanodon bernissartensis and Mantellisaurus atherfieldensis, in the upper Barremian of Cuenca and Castellón Provinces (Gasulla et al., 2014; Llandres et al., 2013; Sanguino & Buscalioni, 2018; Sanz et al., 1982); and Morelladon beltrani, in the upper Barremian of the latter province (Gasulla et al., 2015).
In this study, we report the discovery of new skeletal remains and tracks of large ornithopod dinosaurs in the El Castellar Formation (Salas, 1987) of the Peñagolosa sub-basin (Maestrat Basin, Teruel, Spain; Salas & Guimerà, 1996). Despite their fragmentary and isolated nature, vertebrate remains are highly abundant and diverse in this unit. The dinosaur evidence recorded in the El Castellar Formation of this sub-basin verifies a high diversity of medium to large styracosternan remains (Cobos et al., 2012; Guerrero & Cobos, 2017; Verdú et al., 2019). Specifically, some fossils have been attributed to a large-sized morphotype related to the genera Iguanodon and Magnamanus, and others to a medium-sized one related to Morelladon (Verdú et al., 2019).
Although dinosaur tracks are also moderately abundant in the Barremian of the Peñagolosa sub-basin (Cobos et al., 2013), no tracks from the El Castellar Formation have been formally described yet. Despite this, several tracks of ornithopod dinosaurs preserved as convex hyporeliefs, also called infillings or natural casts, have been mentioned and described in the consecutive Barremian Camarillas and Artoles formations in this sub-basin (Benito et al., 2015; Cobos & Gascó, 2012; Cobos et al., 2013, 2016). Some of these large ornithopod ichnites (as Iguanodontipus in Cobos & Gascó, 2012) have been assigned to the Caririchnium ichnogenus (Díaz-Martínez et al., 2015).
The aims of this study were: (a) to perform a systematic study of the large ornithopod skeletal remains found in the CM-3, CM-4, and CM-8 sites in Cabra de Mora (Teruel Province), describing and comparing them with other related taxa, (b) to test them in a morphological multivariate analysis with other iguanodontians, and (c) to describe and interpret some dinosaur tracks recorded in this municipality.
Institutional abbreviations FCPTD, Fundación Conjunto Paleontológico de Teruel-Dinópolis, Teruel, Spain; GPIT, Institut für Geowissenschaften, Tübingen, Germany; IWCMS, Dinosaur Isle, Sandown, UK; MAP, Museo Aragonés de Paleontología, Fundación Conjunto Paleontológico de Teruel-Dinópolis, Teruel, Spain; MDS, Museo de Dinosaurios de Salas de los Infantes, Salas de los Infantes, Spain; MNS, Museo Numantino de Soria, Soria, Spain; MPT, Museo Provincial de Teruel, Teruel, Spain; NHMUK, The Natural History Museum, London, UK; RBINS, Royal Belgian Institute of Natural Sciences, Brussels, Belgium; USNM, National Museum of Natural History, Smithsonian Institute, Washington, USA; YPM, Yale Peabody Museum of Natural History, New Haven, USA.
Other abbreviations AR, Mina de Santa María de Ariño site, Teruel, Spain (material deposited at MAP); CM, Cabra de Mora site, Cabra de Mora, Teruel, Spain (all material with this acronym is deposited at MAP); CMP-MS, Mas de la Parreta Quarry-Mas de Sabater site, Morella, Spain; CT, El Castellar site, El Castellar, Spain.
2 Geographic and geological settings
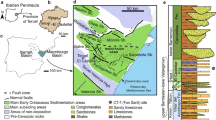
Dinosaur bonesites CM-3, CM-4, and CM-8 are geographically located in the western surroundings of Cabra de Mora Village, in the northwest of the Gúdar-Javalambre region (Teruel Province, Spain; Fig. 1a).

Geographical and geological situation of the study area. a Geographical location of Cabra de Mora Village (Teruel, Spain). b Geological map of Cabra de Mora (Teruel, Spain) with the location of CM-3, CM-4 and CM-8 bonesites, redrawn from Fig. 2a in Campos-Soto et al. (2017). c Simplified stratigraphic section in the surrounding area of Cabra de Mora (modified from the unpublished Doctoral Thesis of Cobos, 2011 and based on data obtained from Campos-Soto et al., 2017, 2019). Kimm. Kimmeridgian, Tithon. Tithonian, Haut. Hauterivian
Geologically, they are included in one of seven sub-basins, the so-called Peñagolosa, controlled by synsedimentary faults which make up the Maestrat Basin (Salas & Guimerà, 1996), in outcrops of the El Castellar Formation (Salas, 1987). In this sub-basin, the El Castellar Formation usually overlies the Villar del Arzobispo Formation (Kimmeridgian-Tithonian sensu Campos-Soto et al., 2017), the uppermost of which is considered to be the Aguilar del Alfambra Formation (Tithonian-Berriasian) by Aurell et al. (2016). However, in the area of Cabra de Mora, this lithostratigraphic unit rests over the Mora de Rubielos Formation (upper Berriasian-lower Valanginian sensu Salas et al., 2001 and Campos-Soto et al., 2017) and is overlain by the Camarillas Formation (lower Barremian, Martín-Closas, 1989; Schudack & Schudack, 2009; Villanueva-Amadoz et al., 2015; Fig. 1b, c).
The El Castellar Formation of the Peñagolosa sub-basin is dated to the upper Hauterivian-lowermost Barremian through an association of charophytes (Martín-Closas, 1989). In general, this lithostratigraphic unit is lithologically divided into two stages (Cobos et al., 2012). The lower one consists of an alternation of violet and grey shales with passes of evaporites and channelled bodies of sandstones. The upper part is composed of grey marls intercalated with limestones (Cobos et al., 2012). According to Fernández-Labrador (2016, unpublished data), the sedimentary environment of the El Castellar Formation in the Peñagolosa sub-basin may have been a coastal wetland system.
The CM-3, CM-4, and CM-8 sites are located in the lower part of the upper section of the El Castellar Formation, where there is a sequence of carbonate layers with some bodies of sandstones (Fig. 1c). Two of these layers, located above the levels with fossil bones, present different tracks preserved as natural casts on their underside, and they extend longitudinally along 350 m. Hereafter, we refer to these layers as Level 1 and Level 2.
3 Material and methods
The described fossils comprise 10 vertebral centra and a fibular fragment of three large ornithopods (Table 1). The remains were found ex-situ at the sites CM-3, CM-4, and CM-8 sites, as a result of the palaeontological tasks carried out by the FCPTD in the municipality of Cabra de Mora, and later housed in the MAP. The materials from each site were described individually and separately.
The tracks studied here include a convex hyporelief with tridactyl morphology found ex situ, and others found in situ at Levels 1 and 2.
In this study, new ornithopod remains from the CM-3, CM-4, and CM-8 sites were systematically studied following the evolutionary hypothesis of Iguanodontia proposed by Verdú et al. (2018). Furthermore, these fossil bones were compared with those of other ornithopod dinosaurs from the Lower Cretaceous of the Iberian Peninsula and other regions, mainly Europe, through first-hand study of their remains housed in different institutions or by consulting the available descriptions. For these comparisons, the genus name was indicated when this is the subject to be discussed and not its species, and the non-abbreviated specific name is used throughout the entire study. Assignations in open nomenclature were made following Sigovini et al. (2016).
Furthermore, multivariate principal component analysis (PCA) was performed on a data set (modified from Verdú et al., 2019) including linear measurements (length, height, and width) obtained from cervical, dorsal, and caudal centra from CM sites and other iguanodontian specimens: the dryosaurids Dryosaurus altus and Valdosaurus canaliculatus, the basal ankylopollexian Camptosaurus dispar, and the styracosternans Magnamanus soriaensis, Iguanodon bernissartensis, Iguanodon cf. galvensis, Iguanodon sp., CT-16 and CT-17 styracosternans, Mantellisaurus atherfieldensis, Morelladon beltrani, and indeterminate tall-spined iguanodontian (see Table 1 in Supplementary Information). For this, we applied a methodology similar to that used by Verdú et al. (2019), which does not take into account deformation and distortion (among other taphonomic processes). In summary, this consisted of an analysis of each data set corresponding to cervical, dorsal, and caudal measurements by PCA using PAST v3.0 software (Hammer et al., 2001) in two ways: first, using the raw data, and second, removing the size component from the data set. Unlike in Verdú et al. (2019), the bootstrapping option was set to 9,999. For further details on the methods, please refer to Verdú et al. (2019).
4 Systematic palaeontology
DINOSAURIA Owen, 1842
ORNITHISCHIA Seeley, 1887 (sensu Madzia et al., 2021).
ORNITHOPODA Marsh, 1881 (sensu Madzia et al., 2021).
IGUANODONTIA Dollo, 1888 (sensu Madzia et al., 2021).
DRYOMORPHA Sereno, 1986 (sensu Madzia et al., 2021).
ANKYLOPOLLEXIA Sereno, 1986 (sensu Madzia et al., 2021).
STYRACOSTERNA Sereno, 1986 (sensu Madzia et al., 2021).
4.1 cf. Styracosterna indet
4.1.1 Material from CM-4 site
Two centra of the middle caudal vertebrae (MAP-4563 and MAP-4566; Fig. 2a, b).

Fossils from CM-4 and CM-8 sites. a, b Caudal vertebrae of cf. Styracosterna indet. from CM-4 site: a Middle caudal centrum MAP-4563 in anterior (a1), left lateral (a2), and ventral (a3) views; and b fragment of middle caudal centrum MAP-4566 in anterior or posterior (b1), and ventral (b2) views. c, d Vertebrae of Iguanodon cf. galvensis from CM-8 site: c Middle-to-anterior dorsal centrum MAP-8044 in posterior (c1), right lateral (c2), ventral (c3), and dorsal (c4) views; and d anterior caudal centrum MAP-8043 in anterior (d1), posterior (d2), right lateral (d3), left lateral (d4), and ventral (d5) views. Abbreviations: cf chevron facet; g groove; nc neural canal; np neural pedicels; tp transverse process. The anatomical profiles have been redrawn and modified from the caudal region of Iguanodon bernissartensis drawing in Norman (1980) and from the dorsal region of Mantellisaurus atherfieldensis drawing in Norman (1986)
Geographical and geological location These fossils were found in the CM-4 site in the Cabra de Mora municipality (Teruel, Spain; Fig. 1a). El Castellar Formation (Peñagolosa sub-basin, Maestrat Basin), upper Hauterivian-lowermost Barremian (Lower Cretaceous; Fig. 1b, c).
Description Caudal vertebrae. MAP-4563 and MAP-4566 are two centra of the middle caudal vertebrae, although the latter represents only a fragment of an articular facet. MAP-4563 (Fig. 2a) is an amphiplatyan (slightly platycoelous) centrum. The cranial articular facet is hexagonal and wider than high. In the lateral view, this centrum shows a rectangular profile that is longer than high. The ventral surface is concave, with a longitudinal groove and a chevron facet. On the other hand, the fragment MAP-4563 shows a cranial or a caudal articular facet less high and wide than both of MAP-4563, and apparently lacks the ventral groove (Fig. 2b). Both caudal vertebrae may have been positioned in the middle caudal region for having centra longer than wide and high, and with hexagonal articular and chevron facets, similar to the middle caudal vertebrae of some styracosternans, such as Iguanodon bernissartensis and Mantellisaurus atherfieldensis (e.g. Norman, 1980, 1986). In addition, the proportions of MAP-4563 (Table 1) together with the presence of a ventral groove, which is occasionally present on the ventral side of some middle caudal centra in some iguanodontians (Verdú et al., 2017 and references therein), also suggest that MAP-4563 belongs to this part of the tail region, and possibly between the 28th and 30th positions. For its part, MAP-4566 might have belonged to any of the most posterior middle caudal vertebrae, possibly between the 35th and 37th vertebrae, because it is slightly smaller than MAP-4566 and lacks a ventral groove.
HADROSAURIFORMES Sereno, 1997 (sensu Madzia et al., 2021).
IGUANODONTOIDEA Norman, 2002 (sensu Verdú et al., 2018).
IGUANODON Mantell, 1825
IGUANODON GALVENSIS Verdú et al., 2015
4.2 Iguanodon cf. galvensis
4.2.1 Material from CM-8 site
A middle-to-anterior dorsal centrum (MAP-8044; Fig. 2c) and an anterior caudal vertebra (MAP-8043; Fig. 2d).
Geographical and geological location These remains originate from the CM-8 site in the Cabra de Mora municipality (Teruel, Spain; Fig. 1a). El Castellar Formation (Peñagolosa sub-basin, Maestrat Basin), upper Hauterivian-lowermost Barremian (Lower Cretaceous; Fig. 1b, c).
Description Dorsal vertebra. The middle-to-anterior dorsal centrum MAP-8044 is highly eroded and fragmented (Fig. 2c). This centrum is amphiplatyan with a sub-elliptic to sub-rectangular caudal facet. In the lateral view, it is rectangular, higher than long, and has craniocaudally concave and dorsoventrally convex lateral surfaces without parapophyses. The ventral surface is rounded and smooth. The neurocentral suture is sinuous and closed. MAP-8044 may have belonged to the 6th or 8th dorsal vertebra because it lacks parapophyses over the centrum, is amphiplatyan, and has sub-elliptical (higher than wide) articular facets, as in some large styracosternans such as Barilium dawsoni (Norman, 2011), Hypselospinus fittoni (Norman, 2015), Iguanodon galvensis (Verdú et al., 2020), Iguanodon bernissartensis (Norman, 1980), and Mantellisaurus atherfieldensis (Norman, 1986).
Caudal vertebra. The anterior caudal vertebra MAP-8043 is well-preserved and is one of the largest centra studied in this work (Fig. 2d). The centrum is platycoelous with sub-quadrangular articular facets. It is craniocaudally compressed and is higher than wide and long. The neural canal is reduced, and the transverse processes are very robust and laterally projected. The ventral surface is smooth and exhibits a wide chevron facet. The neurocentral suture is sinuous and fully closed. MAP-8043 may have belonged to any caudal vertebrae between the 2nd and the 5th because it is a highly platycoelous centrum with sub-quadrangular articular facets, craniocaudally compressed, and with a wide, developed chevron facet as observed in other styracosternans, for instance Hypselospinus fittoni (Norman, 2015), Magnamanus soriaensis (Fuentes-Vidarte et al., 2016), and Iguanodon bernissartensis (Norman, 1980).
4.2.2 Material from CM-3 site
Several centra comprising one middle-to-posterior cervical vertebra (MAP-8036; Fig. 3a), three anterior and middle dorsal vertebrae (MAP-8038, MAP-8035, MAP-8037; Fig. 3b–d), two posterior sacral vertebrae (MAP-4561; Fig. 4a), one middle caudal vertebra (MAP-8033; Fig. 4b), and the proximal epiphysis of the left fibula (MAP-8042; Fig. 4c).

Fossils of Iguanodon cf. galvensis from CM-3 site (I): a Posterior cervical centrum MAP-8036 in anterior (a1), posterior (a2), lateral (a3), dorsal (a4), and ventral (a5) views; b anterior dorsal centrum MAP-8038 in anterior (b1), left lateral (b2), right lateral (b3), and ventral (b4) views; c middle dorsal centrum MAP-8035 in anterior (c1), posterior (c2), left lateral (c3), right lateral (c4), and dorsal (c5) views; d Middle dorsal centrum MAP-8037 in anterior (d1), posterior (d2), left lateral (d3) and ventral (d4) views. k keel, nc neural canal, np neural pedicels, p parapophysis. The anatomical profiles have been redrawn and modified from Mantellisaurus atherfieldensis drawing in Norman (1986)

Fossils of Iguanodon cf. galvensis from CM-3 site (II): a Sacral vertebrae MAP-4561 in anterior (a1), posterior (a2), ventral (a3), and left lateral views (a4), and an interpretative drawing in lateral view (a5); b middle caudal centrum MAP-8033 in posterior (b1), left lateral (b2), and ventral (b3) views; and c proximal end of the left fibula MAP-8042 in distal (c1), medial (c2) views, and on the left fibula of Iguanacolossus fortis (c3), redrawn from McDonald et al. (2010). ac acetabulum, ca caudal vertebra, cf chevron facet, ct contact surface for the fibular process of tibia, g groove, ic intercentrum, il ilium, is ischium, nc neural canal, ns neural spine, s sacral vertebra, s7? possible sacral 7, s8? possible sacral 8, sr sacral ribs. The anatomical profiles have been redrawn and modified from the sacral and caudal region of Iguanodon bernissartensis drawing in Norman (1980)
Geographical and geological location These fossils come from the CM-3 site in the Cabra de Mora municipality (Teruel, Spain; Fig. 1a). El Castellar Formation (Peñagolosa sub-basin, Maestrat Basin), upper Hauterivian-lowermost Barremian (Lower Cretaceous; Fig. 1b, c).
Description Cervical vertebra. MAP-8036 is an almost complete middle-to-posterior cervical centrum (Fig. 3a). It is strongly opisthocoelous, with a convex and heart-shaped cranial facet and a concave caudal facet with a pentagonal outline. Both facets are slightly wider than high. The ventral view shows a wide longitudinal keel. The craniolateral parapophyses have great development. MAP-8036 may have corresponded to the 7th or the 8th cervical vertebrae due to the fact that it is a highly opisthocoelous centrum with wider rather than higher articular facets, and wide parapophyses, similar to those found in the middle-to-posterior cervical vertebrae of other styracosternans such as Iguanodon cf. galvensis (Verdú et al., 2020), Mantellisaurus atherfieldensis (Norman, 1986), and Eolambia caroljonesa (McDonald et al., 2012).
Dorsal vertebrae. The most anterior region of the dorsal vertebrae is represented only by the centrum MAP-8038 (Fig. 3b). This is slightly opisthocoelous; its higher-than-wide cranial facet is convex and heart-shaped, while the caudal facet is concave and highly eroded. The lateral surfaces have a certain degree of concavity craniocaudally, and they converge into a wide and rounded keel ventrally. In addition, this centrum shows a sinuous and completely closed neurocentral suture. MAP-8038 might have belonged to the 1st or 2nd dorsal vertebra based on the absence of parapophyses on the centrum, its degree of opisthocoely, and the slight development of the ventral keel, as observed in other styracosternans, for instance Iguanodon galvensis (Verdú et al., 2018), Iguanodon bernissartensis (Norman, 1980), Mantellisaurus atherfieldensis (Norman, 1986), and Eolambia caroljonesa (McDonald et al., 2012).
The middle region of the dorsal vertebrae is represented by dorsal centra MAP-8035 (Fig. 3c) and MAP-8037 (Fig. 3d). MAP-8035 is highly eroded, and only its anterior half is preserved. The cranial facet is elliptical, higher than wide, and flat. The lateral surfaces are craniocaudally concave. In the ventral view, this centrum does not have a longitudinal keel. In the parts where it is preserved, MAP-8035 exhibits a sinuous and closed neurocentral suture. In contrast, MAP-8037 is nearly complete and is the best-preserved dorsal centrum from the CM-3 site. This element, with a cylindrical morphology and rectangular profile, is taller than wide and long, amphiplatyan, and has elliptical to sub-quadrangular articular facets. The lateral surfaces are craniocaudally concave and dorsoventrally convex, causing the margins of the articular facets to protrude. The ventral surface is rounded and without a longitudinal keel. The neurocentral suture is sinuous and completely closed. Both MAP-8035 and MAP-8037 are middle dorsal vertebrae as they have flat and elliptical to sub-quadrangular articular facets, such as the middle dorsal centra of Barilium dawsoni (Norman, 2011), Iguanodon cf. galvensis (Verdú et al., 2020), Iguanodon bernissartensis (Norman, 1980), and Mantellisaurus atherfieldensis (Norman, 1986). While the higher rather than wider articular facet of MAP-8035 suggests that it may have occupied a more anterior position in the middle dorsal region, probably between the 5th and the 7th dorsal vertebrae, the proportions of MAP-8037 as well as the presence of everted margins over the centrum indicate that this centrum may have occupied a more posterior position in the middle dorsal region, possibly between the 9th, 10th, or 11th dorsal vertebrae.
Sacral vertebrae. MAP-4561 represents two fused sacral vertebrae (Fig. 4a). The most anterior vertebra is more eroded and fragmented, and only preserves its posterior region. However, the posterior vertebra of MAP-4561, significantly better preserved, has a wider than long platycoelous centrum with sub-circular articular facets. These vertebrae have the base of their neural spines completely fused each other. Besides, it has several sub-circular depressions and a closed neurocentral suture in the region where the neural spine meets the centrum. MAP-4561 may have been located in the most posterior region of the sacrum due to the presence of a longitudinal groove extending along the ventral side, likewise the posterior sacral vertebrae of Iguanodon bernissartensis (Norman, 1980), Iguanodon galvensis (unpublished specimens MAP-8048 and MAP-8116), and Equijubus normani (McDonald et al., 2014), and possibly with the 7th and 8th vertebrae because both centra are craniocaudally compressed, unlike the anterior sacral vertebrae of some styracosternans such as Iguanodon bernissartensis (Norman, 1980) and Mantellisaurus atherfieldensis (Norman, 1986).
Caudal vertebra. The caudal vertebrae are only represented by MAP-8033, the left half of a middle caudal centrum (Fig. 4b). It is amphiplatyan, sub-rectangular, and slightly longer than high in the lateral view. In addition, it has craniocaudally concave lateral surfaces. The ventral view shows a longitudinal groove extending along the concave ventral surface and an articular facet of the chevron. The neurocentral suture is sinuous and fully closed. MAP-8033 belongs to a middle caudal vertebra because it lacks transverse processes over the centrum, but it has a chevron facet as in other styracosternans, for instance Iguanodon bernissartensis and Mantellisaurus atherfieldensis (Norman, 1980, 1986). The individually variable ventral groove is also present in the middle caudal vertebrae of some ornithopods (Verdú et al., 2017 and references therein). According to its proportions, this centrum might have occupied a position between the 14th and 17th caudal vertebrae.
Fibula. MAP-8042 is the proximal end of the left fibula and is the single appendicular element preserved in CM-3 (Fig. 4c). This fragment is craniocaudally expanded and roughly crescent-shaped in the distal view. It has a flat medial surface with a certain degree of concavity and a flat-to-convex lateral surface. Moreover, MAP-8042 has two sub-circular protuberances. The lateral one is highly developed and has a medial projection, and the other, barely developed, is almost at a right angle with the shaft. They are separated by a slightly concave surface with an elliptical morphology, which corresponds to the articular surface that receives the fibular process of the left tibia.
5 Results of the multivariate analysis
Only the results of the PCA from the raw data are described here (Fig. 5). PCA from data after removing the component of the size resulted in a very noisy scatter plot, and it was difficult to establish any affinity for Cabra de Mora specimens (see Supplementary Information).

5.1 Cervical vertebrae
PC1 contributes 90.4% to the variance, while PC2 contributes 8.5%. PCA loading scores indicate that correlation with the length, height, and width is high with PC1, but not with PC2 (Table 2). The cervical centra of CM-3 appear in the sector with positive scores for both PC1 and PC2 (Fig. 5a), but are not included in the morphospace of any iguanodontian analysed. The morphospace occupied by the cervical centra of Iguanodon (i.e. Iguanodon bernissartensis, Iguanodon sp., and Iguanodon cf. galvensis) is mainly restricted to the sector with positive values of PC1 as CM-3, but with negative values of PC2. Unlike CM-3, the cervical centra of Mantellisaurus atherfieldensis are resolved mainly in the negative values of PC1, but extend into both negative and positive values of PC2.
5.2 Dorsal vertebrae
In this analysis, PC1 contributes 87% of the variation, while PC2 accounts for 10%. PC1 correlates highly with height and width and to a lesser extent with length, unlike PC2 (Table 2). Here, the resolved scatter plot is similar to that obtained by Verdú et al., (2019: Fig. 6c) with two differentiated groups (Fig. 5b): large-sized ornithopods, including Iguanodon and Magnamanus, occupying positive values of PC1, and small-to-medium-sized iguanodontians, including the styracosternans Mantellisaurus atherfieldensis, Morelladon beltrani, and the indeterminate styracosternan MOR2 (sensu Verdú et al., 2019) from the CT-16 and CT-17 sites. Both CM-3 and CM-8 overlap with the morphospaces of large-sized iguanodontians, in particular Magnamanus soriaensis and Iguanodon bernissartensis.

Natural tracks from Levels 1 and 2 of El Castellar Formation in the area of Cabra de Mora (Teruel, Spain). a Overview of both levels. b Sub-cylindrical infillings recorded on the underside of Level 2 (the scale equals 1 m). c Some of the greater tracks from Level 2. d Ex situ convex hyporelief and without a clear anatomical morphology (the scale bar equals 25 cm). e Tridactyl natural cast on the underside of rolled block from Level 2. f Interpretation of the tridactyl track (the scale equals 10 cm)
5.3 Caudal vertebrae
PC1 represents 90.4% of the variance, whereas PC2 represents only 8.5%. As in the case of cervical centra, loading scores indicate that PC1 is highly correlated with the length, height, and width, unlike PC2 (Table 2). There is no separate morphospace, and each overlaps with the other. The separation between small-to-middle-sized iguanodontians and the large-sized ones observed in the dorsal centra is not present here. This situation can be explained by the similar dimensions and proportions observed between the distal caudal centra in large iguanodontians and proximal centra in smaller ones (Verdú et al., 2019). Despite this, both CM-3 and CM-8 are resolved in or near the morphospace of large-sized ornithopods (Fig. 5c), consistent with the results observed in dorsal vertebrae. In particular, the caudal centra of CM-3 partially overlap with Magnamanus soriaensis, while CM-8 is included in the morphospace of Iguanodon sp. and is very close to the large styracosternan MOR1 (sensu Verdú et al., 2019) of CT-16. The caudal centra of CM-4 are different, being resolved in the morphospace of Valdosaurus canaliculatus, Camptosaurus altus, and the styracosternans Mantellisaurus atherfieldensis and Magnamanus soriaensis (Fig. 5c).
6 Palaeoichnology
6.1 Material
Several tracks of dinosaurs preserved as natural casts, among which one of tridactyl morphology stands out (Fig. 6).
6.2 Geographical and geological situation
The tracks studied here are located on the underside of the so-called Levels 1 and 2 of the El Castellar Formation (Peñagolosa sub-basin, Maestrat Basin), upper Hauterivian-lowermost Barremian (Lower Cretaceous; Figs. 1b, c; 6a). These layers overlie the CM-3, CM-4, and CM-8 sites in the Cabra de Mora municipality (Teruel, Spain).
6.3 Description
6.3.1 Layers with tracks
All convex hyporeliefs are developed on plastic and deformed marl-shales, and they are made up of limestone (those recorded at Level 1) and sandy limestones (those recorded at Level 2). These natural casts are generally deep, irregular, and massive, without an external structure or skin marks. The tracks located at Level 1 do not exceed 20 cm in depth, and their degree of conservation is moderately worse than those presented at Level 2. The latter have more defined and verticalized subcylindrical profiles (Fig. 6b) and a higher content of quartz grains. Furthermore, the infillings are deeper and longer, reaching depths of up to 40 cm and a maximum length of 35 cm (Fig. 6c). In some outcrops of Level 2, the overlapping of different hyporeliefs is common, which may distort the morphology and the original size of these tracks.
6.3.2 Ex situ hyporeliefs
In the surroundings of Levels 1 and 2 outcrops, there are some ex situ hyporeliefs and blocks that, when presenting a sandy matrix with a high presence of quartz grains, may come from Level 2. All these natural casts have a subcylindrical morphology profile and a poorly defined plantar surface (Fig. 6d). However, a convex hyporelief is slightly better preserved in a loose block (Fig. 6e). This track is tridactyl, mesaxonic, and subsymmetrical, wide or wider than longer, such as those pes tracks described to large ornithopod trackmakers and assigned to Iguanodontipodidae ichnofamily (Díaz-Martínez et al., 2015; Lockley et al., 2014; Vialov, 1988; references therein). In addition, it shows relatively short, robust digits with a similar length (slightly mesaxony), but with digit III being somewhat more developed anteroposteriorly, and an apparently rounded heel (Fig. 6f).
7 Discussion
7.1 Ontogenetic development and size
Fossil assemblages from each site, which show similar preservation features and are consistent in size (Table 1), represent three different individuals. In particular, two large individuals are distinguished in the CM-3 and CM-8 sites, and a smaller one in CM-4. This variation in size may be explained by ontogenetic (i.e. the smaller specimen is an immature individual of the larger one) or taxonomical reasons (i.e. large and smaller individuals represent different taxa). In fact, the presence of both medium and large-sized styracosternans has been demonstrated in the El Castellar Formation (Verdú et al., 2019). To discriminate between these two possibilities, it is necessary to determine the degree of ontogenetic development of these remains.
During the growth of ornithopods, several changes occurred in the axial skeleton that allow us to assess the degree of ontogenetic development of these vertebral remains. Among these, the degree of closure of the neurocentral suture (e.g. Griffin et al., 2020; Hübner, 2018; Verdú, 2017) and fusion of sacral vertebrae (Horner et al., 2004) stand out. The complete closure of the neurocentral suture in the dorsal centra of CM-3 and CM-8 indicates that these remains may belong to two sub-adult or adult individuals (Verdú, 2017 and references therein). In addition, the sacral centra in MAP-4561 is completely co-ossified, similar to the sacral vertebrae of other skeletally mature ornithopods (Horner et al., 2004).
In relation to their size, the centra of CM-3 and CM-8 are comparable with the vertebrae of other large ornithopods of the Lower Cretaceous of Europe. The anterior dorsal centrum MAP-8038 of CM-3 is longer (97 mm) than the first and second dorsal vertebrae of specimen RBINS 1551 of the medium-sized styracosternan Mantellisaurus atherfieldensis (68 and 70 mm respectively, Norman, 1986) estimated to be 6.5 m long (Norman, 2014). The middle dorsal centra MAP-8044 of CM-8 and MAP-8037 of CM-3 show a length (95 and 93 mm, respectively) similar to that of their homologous vertebrae in specimen RBINS 1534 of the large-sized styracosternan Iguanodon bernissartensis (6th and 10th dorsal, 93 and 98 mm respectively; Norman, 1980, 1986) which was roughly 8–10 m long (Norman, 2014; Paul, 2008), but both are slightly shorter than the equivalent vertebrae of a mature specimen of Iguanodon cf. galvensis, 9–10 m long (e.g. MAP-2067 and MAP-2065, 112 and 108 mm respectively, Verdú et al., 2020). Other vertebral centra such as the anterior caudal centrum MAP-8043 of CM-8 is almost as long (88 mm) and high (136 mm) as the anterior caudal vertebrae of the RBINS 1534 specimen of Iguanodon bernissartensis (e.g. 4th caudal vertebra, 87 and ~ 140 mm, respectively; Norman, 1980, 1986). Therefore, these observations on measurements of the vertebrae are consistent with observations in the pattern of closure of the neurocentral suture, and CM-3 and CM-8 are likely to belong to an adult individual of a huge styracosternan. Based on the size of CM-3 and CM-8 centra, their comparison with the vertebrae of more complete and articulated specimens of Iguanodon bernissartensis (Norman, 1980), Mantellisaurus atherfieldensis (Norman, 1986), and Iguanodon cf. galvensis (Verdú et al., 2020), and with the skeletal reconstructions of Norman (2004) and Paul (2008), we interpret the size of adult specimens of CM-3 and CM-8 as being very similar to that inferred by Paul (2008) for the Iguanodon bernissartensis holotype (RBINS 1534) of approximately 8–9 m long.
In the case of a CM-4 ornithopod, it is difficult to establish a concrete stage of development on the basis of the degree of closure of the neurocentral sutures. This follows a caudocranial pattern and occurs in early stages in the caudal region of some archosaurs (e.g. Brochu, 1996; Ikejiri, 2012, 2015), including ornithopod dinosaurs (Galton, 2012; Hübner, 2018) such as Iguanodon (Verdú, 2017), but alternatives to this sequence of closure have been also described in this clade (Griffin et al., 2020 and references therein). The middle caudal centrum MAP-4563 of CM-4 is larger (76 mm) than the 29th caudal vertebra of RBINS 1551 specimen of Mantellisaurus atherfieldensis (70 mm; Norman, 1986) and that of a sub-adult specimen of Iguanodon bernissartensis (RBINS 1730, 74 mm; Norman, 1980, 1986), which was approximately 7 m long (Verdú et al., 2017), but smaller than the 27th caudal vertebra of an adult specimen of this species (RBINS 1534, 98 mm; Norman, 1980, 1986) or its homologous vertebrae in an adult specimen of Magnamanus soriaensis (e.g. MNS2003/69.434b, 107 mm; Fuentes-Vidarte et al., 2016). Using only the size as a reference, the CM-4 remains belonged to a sub-adult of a large-sized species or an adult of a medium-sized species and, therefore, the difference in size observed between the CM-4 ornithopod and those of CM-3 and CM-8 was due to ontogeny, taxonomy, or both.
7.2 Major-group assignment of skeletal remains
The following characters (although they are not diagnostic of the group) demonstrate that the remains of CM-3, CM-4, and CM-8 belong to ornithopod dinosaurs: (1) vertebral centra without pleurocoeli, (2) dorsal and caudal centra slightly compressed between the articular facets, (3) middle caudal centra with hexagonal outline, and (4) the presence of sinuous neurocentral suture in some of them (Knoll, 2009; Norman, 2004; Pereda-Suberbiola et al., 2011; Verdú et al., 2019, 2020).
The ornithopods identified in the Hauterivian-Barremian section of the Iberian Peninsula are small non-iguanodontian ornithopods related to Hypsilophodon (Ruiz-Omeñaca et al., 2012), rhabdodontomorphans (Dieudonné et al., 2016), dryosaurids (e.g. Galton, 2009) and medium to large-sized non-hadrosaurid styracosternans (e.g. Fuentes-Vidarte et al., 2016; Gasulla et al., 2014, 2015; Sanguino & Buscalioni, 2018; Santos-Cubedo et al., 2021; Verdú et al., 2015, 2020). In relation to such diversity, the remains from CM-3, CM-4, and CM-8 much more closely resemble those of large styracosternans, as discussed in the following paragraphs.
7.2.1 CM-4
The middle caudal centra of CM-4 (Fig. 2a, b) belonged to Iguanodontia because they show hexagonal articular facets (Norman, 2004). However, their sizes differ from those of their homologous vertebrae in non-styracosternan iguanodontians. In fact, MAP-4563 is almost twice as long (76 mm) as its equivalent in the IWCMS 2013.175 specimen of the dryosaurid Valdosaurus canaliculatus (e.g. 20th caudal, 49 mm, Barrett, 2016) and much longer than the middle caudal vertebrae of a rhabdodontomorpha (e.g. MDS-VG.102, 10.9 mm, Dieudonné et al., 2016). Instead, MAP-4563 is more similar to the middle caudal vertebrae of some styracosternans, such as sub-adults of the large-sized Iguanodon bernissartensis and presumably adults of medium-sized Mantellisaurus atherfieldensis, as indicated in the previous section. Despite their significant size and the relative abundance of styracosternan remains in the El Castellar Formation, the isolated nature of CM-4 remains does not allow these fossils to be classified with more precision than cf. Styracosterna indet. because no diagnostic features of these clades have been identified in these specimens.
7.2.2 CM-8
In the case of CM-8, the anterior caudal vertebra MAP-8043 is craniocaudally compressed (Fig. 2d), similar to the anterior caudal vertebrae of some ornithopods such as the ankylopollexians Camptosaurus dispar (Gilmore, 1909), Uteodon aphanoecetes (Carpenter & Wilson, 2008; sensu McDonald, 2011), Barilium dawsoni (Norman, 2011), Hypselospinus fittoni (Norman, 2015), Magnamanus soriaensis (Fuentes-Vidarte et al., 2016), Iguanodon bernissartensis (Norman, 1980), Mantellisaurus atherfieldensis (Norman, 1986), Ouranosaurus nigeriensis (Bertozzo et al., 2017; Taquet, 1976), and Eolambia caroljonesa (McDonald et al., 2012), among others. Moreover, these remains are similar (amphiplatyan dorsal centrum and platycoelous anterior caudal centrum with sub-quadrangular articular facets) to the remains of CM-3 and those described in other medium to large-sized styracosternans (e.g. Fuentes-Vidarte et al., 2016; Norman, 1980, 1986; Verdú et al., 2015, 2020). Furthermore, in terms of absolute size, the vertebral centra of CM-8 are only comparable with the centra of large-sized styracosternan ornithopods at the current state of our knowledge of ornithopod diversity; therefore, they are considered to belong to this clade.
7.2.3 CM-3
The middle-to-posterior cervical vertebra MAP-8036 (Fig. 3a) has a strongly opisthocoelous centrum, like the cervical centra of other styracosternans such as Barilium dawsoni (Norman, 2011), Hypselospinus fittoni (Norman, 2015), Iguanodon cf. galvensis (Verdú et al., 2020), Iguanodon bernissartensis (Norman, 1980), Mantellisaurus atherfieldensis (Norman, 1986), Proa valdearinnoensis (AR-1/103), Ouranosaurus nigeriensis (Bertozzo et al., 2017; Taquet, 1976), and hadrosaurids (Horner et al., 2004). In fact, Norman (2015) recovered the opisthocoely in the cervical vertebrae as a character that diagnoses the Styracosterna clade in his phylogenetic analysis. This feature is not observed in non-styracosternan ornithopods such as Hypsilophodon foxii (Galton, 1974), rhabdodontomorphans (Ösi et al., 2012; Weishampel et al., 2003), or the basal ankylopollexian Camptosaurus dispar (Gilmore, 1909). In styracosternans, it is also common to have the anterior dorsal centrum moderately opisthocoelous (Norman, 2015) as in MAP-8038 (Fig. 3b) and other European taxa such as Hypselospinus fittoni (Norman, 2015), Iguanodon bernissartensis (Norman, 1980), Iguanodon galvensis (Verdú et al., 2018), and Mantellisaurus atherfieldensis (Norman, 1986). Therefore, the ornithopod remains of CM-3, similar to those of CM-8, are assigned to Styracosterna.
7.3 Comparison with Early Cretaceous styracosternans from Europe
7.3.1 CM-4
MAP-4563, assigned to Styracosterna indet., is an amphiplatyan (slightly platycoelous) middle caudal vertebrae similar to those of specimens of CM-3, perinates of Iguanodon galvensis (Verdú, 2017), and mature specimens of Mantellisaurus atherfieldensis (Norman, 1986), but unlike the amphicoelous of Barilium dawsoni (Norman, 2011) or the platycoelous of Magnamanus soriaensis (Fuentes-Vidarte et al., 2016) and Iguanodon bernissartensis (Norman, 1980). The middle caudal vertebrae of Morelladon beltrani are unknown (Gasulla et al., 2015). The lateral surfaces of MAP-4563 retain a slight ridge craniocaudally expanded, similar to the crested middle-to-posterior vertebrae of Barilium dawsoni (Norman, 2011), Hypselospinus fittoni (Norman, 2015), Magnamanus soriaensis (Fuentes-Vidarte et al., 2016), Iguanodon bernissartensis (Norman, 1980), and Mantellisaurus atherfieldensis (Norman, 1986). However, PCA analysis does not clarify the taxonomic affinity of CM-4, as it is included not only in the morphospace of the medium-sized styracosternan Mantellisaurus atherfieldensis, but also the large-sized Magnamanus soriaensis (Fig. 5c). Therefore, the CM-4 remains belonged to an iguanodontian, probably a styracosternan, with unclear affinities with other Early Cretaceous European species.
7.3.2 CM-8 and CM-3
Here, comparisons are made with both CM-8 and CM-3 specimens because we assume that they belong to the same or a very close species due to they share very similar dorsal vertebrae. The middle dorsal centra MAP-8035 and MAP-8037 of CM-3 and MAP-8044 of CM-8 are amphiplatyan and have elliptical to sub-rectangular articular facets (higher than wide; Fig. 7a, b), resembling the dorsal centra of Barilium dawsoni (Norman, 2011), Hypselospinus fittoni (Norman, 2015), Magnamanus soriaensis (Fuentes-Vidarte et al., 2016), Iguanodon cf. galvensis (Verdú et al., 2020), Iguanodon bernissartensis (Norman, 1980, 1986), Mantellisaurus atherfieldensis (Norman, 1986), and Brighstoneus simmondsi (Lockwood et al., 2021), but unlike the platycoelous dorsal centra of Morelladon beltrani (Gasulla et al., 2015). Additionally, these centra are dorsoventrally higher than craniocaudally long, like the middle dorsal vertebrae of the large-sized styracosternans Barilium dawsoni (Norman, 2011; Fig. 7c), Magnamanus soriaensis (Fuentes-Vidarte et al., 2016; Fig. 7d), Iguanodon cf. galvensis (Verdú et al., 2020; Fig. 7e), and Iguanodon bernissartensis (Norman, 1980; Verdú et al., 2020; Fig. 7f), but also the medium-sized styracosternan Brighstoneus simmondsi (Lockwood et al., 2021). They differ from the longer-than-high mid-to-posterior dorsal vertebrae of the medium-sized styracosternans Mantellisaurus atherfieldensis (Norman, 1986; Fig. 7g) and Morelladon beltrani (Gasulla et al., 2015; Fig. 7h). Furthermore, some styracosternans exhibit a ventral keel in their dorsal vertebrae, such as Barilium dawsoni (Norman, 2011), Hypselospinus fittoni (Norman, 2015), Iguanodon bernissartensis (Norman, 1980, 1986), Mantellisaurus atherfieldensis (Norman, 1986), Brighstoneus simmondsi (Lockwood et al., 2021), and Morelladon beltrani (Gasulla et al., 2015). In contrast, a ventral keel is absent in the dorsal centra of CM-3 (Fig. 7a) and CM-8 (Fig. 7b), similar to Iguanodon galvensis (Verdú et al., 2020; Fig. 7e) and Magnamanus soriaensis (Fuentes-Vidarte et al., 2016; Fig. 7d). In addition, the dorsal centra of CM-3 and CM-8 are moderately compressed between both articular facets, resembling those of Iguanodon cf. galvensis (Verdú et al., 2020) and Magnamanus soriaensis (Fuentes-Vidarte et al., 2016), but unlike the highly compressed centra of Iguanodon bernissartensis (Norman, 1986).

Comparison of some remains of CM-3 and CM-8 with other styracosternans of the Lower Cretaceous of Europe. a Dorsal centrum MAP-8037 in anterior (a1), lateral (a2) and ventral (a3) views. b Dorsal centrum MAP-8044 in anterior (b1), lateral (b2) and ventral (b3) views. c Middle dorsal vertebra NHMUK R798 of Barilium dawsoni in lateral view (c1) (redrawn from Norman, 2011). d Dorsal vertebra MNS 2001/122.576 of Magnamanus soriaensis in anterior view (d1) (redrawn from Fuentes-Vidarte et al., 2016). e Dorsal vertebra MAP-2074 of Iguanodon cf. galvensis in anterior (e1), ventral (e2) and lateral (e3) views (redrawn from Verdú et al., 2020). f Dorsal vertebrae of RBINS R352 specimen of Iguanodon bernissartensis in lateral (f1) and ventral (f2) views (redrawn from Verdú et al., 2020). g Dorsal vertebra of RBINS 1551 specimen of Mantellisaurus atherfieldensis in lateral (g1) and ventral (g2) views (redrawn from Norman, 1986). h Dorsal vertebra CMP-MS-03-10 of Morelladon beltrani in anterior (h1) and lateral (h2) views (redrawn from Gasulla et al., 2015). i Sacral vertebrae MAP-4561 in ventral view (i1). j Sacrum of 1535 RBINS specimen of Iguanodon bernissartensis in ventral view (j1) (redrawn from Norman, 1980). k Sacrum NHMUK R3789 of Barilium dawsoni in ventral view (k1) (redrawn from Norman, 2012). l Partial sacrum NHMUK R1635 of Hypselospinus fittoni in ventral view (l1) (redrawn from Norman, 2015). m Sacrum of Mantellisaurus atherfieldensis in ventral view (m1) (redrawn from Norman, 1986). n Sacrum of Morelladon beltrani in ventral view (n1) (redrawn from Gasulla et al., 2015). ac acetabulum, ca caudal vertebra, g groove, il ilium, is ischium, k keel, s sacral vertebra, sd sacrodorsal vertebra, sv, smooth ventral surface
The posterior sacral vertebrae of MAP-4561 (Fig. 7i) in CM-3 have a deep ventral groove similar to that observed in some posterior sacral vertebrae of Iguanodon galvensis (e.g. unpublished specimens MAP-8048 and MAP-8116), Iguanodon bernissartensis (Norman, 1980; Verdú et al., 2018; Fig. 7j), and Proa valdearinnoensis (unpublished specimen AR-1/103). However, this groove is not present in the sacrum of Barilium dawsoni (Norman, 2011; Fig. 7k), Hypselospinus fittoni (Norman, 2015; Fig. 7l), Mantellisaurus atherfieldensis (Norman, 1986; Fig. 7m), and Morelladon beltrani (Gasulla et al., 2015; Fig. 7n), which have a ventral keel instead (Fig. 7k–n). In addition, Brighstoneus simmondsi apparently shows a ventral keel on the first sacral vertebra, but the rest of the sacral vertebrae are flat (Lockwood et al., 2021). The sacrum is unknown in Magnamanus soriaensis (Fuentes-Vidarte et al., 2016). This ventral groove has been described in the sacrum of other non-European styracosternans such as Equijubus normani (McDonald et al., 2014), Bactrosaurus johnsoni (Godefroit et al., 1998), Gilmoreosaurus mongoliensis (Prieto-Márquez & Norell, 2010), and some hadrosaurids (Horner et al., 2004).
The anterior caudal centrum MAP-8043 of CM-8 is platycoelous with sub-quadrangular articular facets, a smooth ventral surface with wide chevron facet, and reduced neural canal, similar to the anterior caudal vertebrae of Hypselospinus fittoni (Norman, 2015), Magnamanus soriaensis (MNS 2000/132.2; Fuentes-Vidarte et al., 2016), and Iguanodon bernissartensis (Norman, 1980). The two first caudal vertebrae of Brighstoneus simmondsi are, or at least appear to be, slightly platycoelous (Lockwood et al., 2021). Other European styracosternans such as Barilium dawsoni (Norman, 2011), and perinates of Iguanodon galvensis (Verdú, 2017) or Mantellisaurus atherfieldensis (Norman, 1986) have anterior caudal vertebrae with amphiplatyan centra. The anterior caudal vertebrae of Morelladon beltrani and mature specimens of Iguanodon galvensis have not been described (Gasulla et al., 2015; Verdú, 2017).
The middle caudal centrum MAP-8033 of CM-3 is amphiplatyan as MAP-4563 in CM-4 (although they are not comparable in size and position), similar to those of Mantellisaurus atherfieldensis (Norman, 1986; Table 1), and perinates of Iguanodon galvensis (Verdú, 2017). It differs from the amphicoelous caudal vertebrae of Barilium dawsoni (Norman, 2011) or the platycoelous of Magnamanus soriaensis (Fuentes-Vidarte et al., 2016), and Iguanodon bernissartensis (Norman, 1980).
The proximal epiphysis of the left fibula MAP-8042 of CM-3 closely resembles (proximal end craniocaudally expanded with a concave medial side and a flat-to-convex lateral side, and with crescent shaped in distal view) those described for other European styracosternans such as the right fibula of one perinate of Iguanodon galvensis (MAP-4902, Verdú, 2017), Iguanodon bernissartensis (Norman, 1980), and Mantellisaurus atherfieldensis (NHMUK R6464, Norman, 1986), but also with other non-European iguanodontians as the styracosternan Iguanacolossus fortis (McDonald et al., 2010).
To summarise, CM-3 and CM-8 fossils resemble those of the Iberian styracosternans Magnamanus soriaensis, Iguanodon galvensis, and Iguanodon bernissartensis in having: (1) amphiplatyan middle dorsal centra with (2) elliptical (higher than wide) articular facets, (3) which are higher than long in lateral view; and (4) posterior sacral vertebrae with a ventral groove (latter unknown in Magnamanus soriaensis and CM-8), and (5) platycoelous anterior caudal centrum (unknown in CM-3) with (6) sub-quadrangular articular facets (unknown in CM-3). Given this combination of characters (some of them appearing in the diagnosis of Iguanodon and their species sensu Verdú et al., 2018), here we preliminarily assign these remains to Iguanodon genus, expecting that the anatomical similarities of Magnamanus with Iguanodon species—it potentially belongs to Iguanodon—pointed out by Verdú et al. (2020) to be clarified in the future. In the same way, the PCA results also support the similarity of the dorsal and caudal centra of CM-3 and CM-8 to those of the large-sized styracosternans Magnamanus or Iguanodon (Fig. 5b, c). Finally, these remains are attributed in open nomenclature to Iguanodon cf. galvensis, because dorsal vertebrae are moderately compressed between both articular facets and do not present a ventral keel as in Iguanodon galvensis (Verdú et al., 2020; condition present as well in Magnamanus, Fuentes-Vidarte et al., 2016), but unlike the highly compressed centra with a ventral keel of Iguanodon bernissartensis (Norman, 1980).
7.4 Diversity of large ornithopods in the El Castellar Formation of the Peñagolosa sub-basin
Verdú et al. (2019) proposed the existence of at least two morphotypes of large ornithopods, a large-sized styracosternan morphotype (MOR1) related to Magnamanus and Iguanodon, and another medium-sized styracosternan related to Morelladon beltrani (MOR2), after the study of some dorsal and caudal vertebrae from this lithostratigraphic unit.
As mentioned above, it is unknown if CM-4 (cf. Styracosterna indet.) corresponds to an adult of medium-size species or a sub-adult of a large one. The amphiplatyan middle caudal vertebra MAP-4563 of CM-4 is different to and shorter (76 mm) than those of the platycoelous or amphicoelous of the large MOR1 (e.g. MAP 4548, 100.8 mm; Verdú et al., 2019) or to the more anterior and amphicoelous of the medium MOR2, despite being similar in length (MAP-3906, 76.6 mm; Verdú et al., 2019). However, this vertebra is similarly amphiplatyan to those of the CM-3 (here as Iguanodon cf. galvensis) and perinates of Iguanodon galvensis, but also to the medium-sized styracosternan Mantellisaurus atherfieldensis. Therefore, the CM-4 specimen (cf. Styracosterna indet.) cannot be confidently ascribed to either medium or large-sized styracosternan morphotypes, given the available evidence as previously reported in comparison with European taxa.
CM-3 and CM-8 (both attributed to Iguanodon cf. galvensis) have more features in common with the genera Iguanodon and Magnamanus than with any other styracosternan described for the Hauterivian-Barremian range of the Iberian Peninsula, as is also the case for the large-sized morphotype (MOR1) described by Verdú et al. (2019). In fact, CM-8 and MOR1 share similar platycoelous anterior caudal vertebrae with sub-quadrangular articular facets, a unique element common to both specimens. These caudal vertebrae are different from the amphicoelous vertebrae of MOR2 in Verdú et al. (2019). The higher-than-long and amphiplatyan dorsal vertebrae of CM-3 and CM-8 (Iguanodon cf. galvensis), not recovered in MOR1, clearly differ from those of longer-than-high and platycoelous dorsal vertebrae of MOR2 (Verdú et al., 2019). Although the Lower Cretaceous fossils of Iguanodon and Magnamanus have not been described in the El Castellar Formation of the Peñagolosa sub-basin, they have been identified in other lithostratigraphic units, such as the presence of Iguanodon galvensis in the immediately overlying lower Barremian Camarillas Formation of the Galve sub-basin (Verdú et al., 2015, 2020) or Magnamanus soriaensis in the upper Hauterivian-lower Barremian Golmayo Formation of the Cameros Basin (Fuentes-Vidarte et al., 2016). Therefore, CM-3, CM-8, and MOR1 might represent different specimens of the same large-sized styracosternan, perhaps related to Iguanodon galvensis, as stated in this work for the two former specimens. The classification of part of this material to Iguanodon cf. galvensis corroborates the probable existence of this taxon in the sediments of the El Castellar Formation in the Peñagolosa sub-basin. This not only represents the potential oldest record for this species, but also allows us to eliminate the exclusive paleogeographic distribution for Iguanodon galvensis in the Galve sub-basin and extend it, as of now, to the Peñagolosa sub-basin.
Other remains, such as the proximal end of the left fibula MAP-8042 of CM-3, are very similar, both in size and in the previously mentioned characteristics, to that of an indeterminate styracosternan from the El Castellar Formation in the same-named municipality (Cobos et al., 2012) corresponding to the medium-sized morphotype (MOR2) in Verdú et al. (2019); therefore, the differences are not significant for this partial bone in both large and medium-sized morphotypes.
The PCA results of this study verified the hypothesis that two types of non-hadrosaurid styracosternans, one medium and other large-sized, are present in the upper Hauterivian-lowermost Barremian El Castellar Formation, as has also been reported for other Lower Cretaceous lithostratigraphic units in Europe (Verdú et al., 2019 and references therein). This is particularly evident in the case of the dorsal vertebrae, where MOR2 centra are resolved as overlapping by the morphospace occupied by medium-sized styracosternans such as Morelladon beltrani, and CM-3 and CM-8 (Iguanodon cf. galvensis) by the morphospace of large-sized styracosternans such as Magnamanus soriaensis and Iguanodon bernissartensis (Fig. 5b).
These styracosternans, represented by two morphotypes, may have been the dominant dinosaurs in the extensive coastal wetland system where the materials of the El Castellar Formation were deposited. In addition, these ornithopods share the ecosystem with other Barremian dinosaurs in this sub-basin and in the nearby Galve sub-basin (e.g. Cobos et al., 2012; Gasca, 2011; Gasca et al., 2008, 2018; Luque et al., 2006; Royo-Torres & Cobos, 2007; Verdú et al., 2019), as well as with testudines (e.g. Pérez-García et al., 2013, 2017), crocodyliforms, and fishes (Arribas et al., 2019; Guerrero & Cobos, 2017), among other vertebrates.
7.5 Palaeoichnology interpretation
In general, the natural casts from Level 1 are highly irregular and do not show a clear anatomical morphology. However, those from Level 2, with a fine-grained matrix and composed mainly of grains of quartz and ferruginous elements, show a higher degree of preservation. Although the anatomical definition is worse, these infillings resemble those described in the Río Alcalá site and assigned to large ornithopod trackmakers by Cobos et al. (2016). In addition, the best-preserved tridactyl track (Fig. 6e, f), found on the underside of a metric sandstone block from Level 2, shows similar features: (1) mesaxonic and tridactyl tracks, with (2) sub-symmetrical pes morphology that is as wide as or wider than long, to those described and assigned to the Iguanodontipodidae ichnofamily (Díaz-Martínez et al., 2015; Lockley et al., 2014; Vialov, 1988). Here, we assigned this track to Iguanodontipodidae indet. because of the absence of diagnostic characteristics. However, this natural cast resembles Caririchnium-like morphology (e.g. Díaz-Martínez et al., 2015; Figueiredo et al., 2022; Leonardi, 1984; Meyer & Thüring, 2003) for having: (1) wide heel impression (unlike the small, rounded heel of Iguanodontipus and the large, bilobed one of Hadrosauropodus), (2) relatively short and robust digits, with (3) rounded ends (unlike the narrow ones of Iguanodontipus). This ichnogenus is usually related to basal styracosternans (Díaz-Martínez et al., 2015; Lockley et al., 2014). Therefore, the assignment of the tracks to these potential trackmakers is consistent with these data and with the abundance of skeletal remains of these dinosaurs in the El Castellar Formation of the Peñagolosa sub-basin (Cobos et al., 2012; Guerrero & Cobos, 2017; Verdú et al., 2019; this work).
These ornithopod trackmakers may have a height to the acetabulum of 135 cm for the smallest (35 cm of track length) and 180 cm for the largest (45 cm of track length), according to Alexander (1976), or 190 and 250 cm according to Lockley et al. (1983). The disparity between these estimates suggests that they were produced by individuals of different sizes. Some of them denote, together with some bones described in this article (Table 1), the large size that this type of dinosaur might have reached during the Early Cretaceous. These data are congruent with the height up to the acetabulum inferred from the skeletal reconstruction of Paul (2008) for Iguanodon bernissartensis holotype (RBINS 1534 specimen), which is approximately 210 cm.
The large ornithopod-dominated ichnofauna in these facies of the El Castellar Formation corroborates that they frequented the coastal wetlands during the Hauterivian-Barremian of the current Peñagolosa sub-basin. In fact, Verdú et al. (2020) reported that Iguanodon and similar iguanodontians recurrently occupied areas near sources of fresh water in lacustrine, fluvial, and other depositional systems with more marine influence due to the association of several specimens of Iguanodon of different ages and tracks of large ornithopods in this type of facies.
8 Conclusions
This paper describes several ornithopods postcranial remains from the CM-3, CM-4, and CM-8 sites. All these fossils come from the upper Hauterivian-lowermost Barremian (Lower Cretaceous) of the El Castellar Formation of the Peñagolosa sub-basin and belong to three large ornithopods related to Styracosterna. The bones found in CM-3 and CM-8 belonged to two adult individuals, as evidenced by the presence of a closed neurocentral suture in the dorsal vertebrae and the size of the elements. In contrast, CM-4 was probably a sub-adult or adult based on its size.
The remains of CM-3 and CM-8 are assigned to Iguanodon cf. galvensis due to a combination of characters present in the dorsal, sacral, and caudal vertebrae, particularly the presence of dorsal centra which are moderately compressed between both articular facets and without a ventral keel, as in Iguanodon galvensis. In fact, the multivariate analysis (PCA) revealed a great affinity of the cervical and dorsal vertebrae of CM-3, and dorsal vertebra of CM-8, with the large-sized ornithopods Magnamanus and Iguanodon, though it is unclear for caudal vertebrae. This, in combination with their systematic study and the presence in this unit of other morphologically different dorsal vertebrae, verified the hypothesis of the presence of two morphotypes of non-hadrosaurid styracosternans in the El Castellar Formation: one medium-sized, related to Morelladon, and another large-sized, related to Iguanodon galvensis. CM-3 and CM-8 specimens represent the oldest record referable to Iguanodon galvensis and extend the distribution of this taxon to the Peñagolosa sub-basin. On the other hand, neither the multivariate analysis nor the anatomical features of the CM-4 vertebrae allow us to determine its taxonomic affinity with more precision than cf. Styracosterna indet. and to assign them to the medium or the large-sized morphotypes.
Finally, the tracks found in the upper section of the El Castellar Formation, which are all preserved as natural casts, are related to large ornithopod trackmakers. The unique presence of ornithopod tracks together with the large number of large-sized styracosternan remains in this unit and geological area provides new evidence that these dinosaurs may have been frequent in the coastal wetlands that represent the El Castellar Formation in the Peñagolosa sub-basin.
Availability of data and material
The remains studied here are deposited in a public institution (Museo Aragonés de Paleontología).
Code availability
Not applicable for that section.
References
Alcalá, L., Cobos, A., & Royo-Torres, R. (2018). Dinosaurios de la Península Ibérica. Revista PH, 94, 116–153. https://doi.org/10.33349/2018.0.4145
Alexander, R. M. (1976). Estimates of speeds of dinosaurs. Nature, 261(5556), 129–130. https://doi.org/10.1038/261129a0
Arribas, I., Buscalioni, A. D., Royo-Torres, R., Espílez, E., Mampel, L., & Alcalá, L. (2019). A new goniopholidid crocodyliform, Hulkepholis rori sp. Nov. from the Camarillas Formation (early Barremian) in Galve, Spain. PeerJ, 7, e7911. https://doi.org/10.7717/peerj.7911
Aurell, M., Bádenas, B., Gascá, J. M., Canudo, J. I., Liesa, C. L., Soria, A. R., et al. (2016). Stratigraphy and evolution of the Galve sub-basin (Spain) in the middle Tithonian-early Barremian: Implications for the setting and age of some dinosaur fossil sites. Cretaceous Research, 65, 138–162.
Barrett, P. M. (2016). A new specimen of Valdosaurus canaliculatus (Ornithopoda: Dryosauridae) from the Lower Cretaceous of the Isle of Wight, England. Memoirs of Museum Victoria, 74, 29–48. https://doi.org/10.24199/j.mmv.2016.74.04
Benito, M. I., Cobos, A., Mas, J. R., Alcalá, L., Campos-Soto, S., & Fernández-Donado, L., et al. (2015). “Walking on tides”: Sedimentary controls on dinosaur tracks formation and preservation in the Barremian coastal-alluvial to tidal-flat deposits of SE Spain (W Penyagolosa Sub-basin, Maestrat Basin). In: The 9th International Conference on Tidal Sedimentology (Ed.), Abstracts Tidalites (pp. 117–120). Puerto Madryn, Patagonia, Argentina: Asociación Argentina de Sedimentología y CONICET/CENPAT.
Bertozzo, F., Dalla-Vecchia, F. M., & Fabbri, M. (2017). The Venice specimen of Ouranosaurus nigeriensis (Dinosauria, Ornithopoda). PeerJ, 5, e3403. https://doi.org/10.7717/peerj.3403
Brochu, C. (1996). Closure of neurocentral sutures during crocodilian ontogeny: Implications for maturity assessment in fossil archosaurs. Journal of Vertebrate Paleontology, 16(1), 49–62.
Campos-Soto, S., Benito, M. I., Cobos, A., Caus, E., Quijada, I. E., Suarez-González, P., et al. (2019). Revisiting the age and palaeoenvironments of the Upper Jurassic-Lower Cretaceous? Dinosaur-bearing sedimentary record of eastern Spain: Implications for Iberian palaeogeography. Journal of Iberian Geology, 45, 471–510.
Campos-Soto, S., Cobos, A., Caus, E., Benito, M. I., Fernández-LabradorSuarez-González, L. P., et al. (2017). Jurassic Coastal Park: A great diversity of palaeoenvironments for the dinosaurs of the Villar del Arzobispo Formation (Teruel, eastern Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 485, 154–177. https://doi.org/10.1016/j.palaeo.2017.06.010
Carpenter, K., & Wilson, Y. (2008). A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of its forelimb. Annals of Carnegie Museum, 76(4), 227–263.
Cobos, A., & Gascó, F. (2012). Presencia del género Iguanodontipus en el Cretácico Inferior de la provincia de Teruel (España). Geogaceta, 52, 185–188.
Cobos, A., Gascó, F., Royo-Torres, R., & Alcalá, L. (2013). “Dinoichnofacies” barremienses en Teruel (España). In C. Álvarez-Vázquez & I. López-Rodríguez (Eds.), Libro de Resúmenes de las XXIX Jornadas de la Sociedad Española de Paleontología (pp. 65–66). Real Jardín Botánico de Córdoba.
Cobos, A., Gascó, F., Royo-Torres, R., Lockley, M. G., & Alcalá, L. (2016). Dinosaur tracks as “four-dimensional phenomena” reveal how different species moved. In P. L. Falkingham, D. Marty, & A. Richter (Eds.), Dinosaur tracks: The next steps (pp. 244–256). Indiana University Press.
Cobos, A., Royo-Torres, R., & Alcalá, L. (2012). Restos postcraneales de ornitópodo en el Cretácico Inferior de El Castellar (Subcuenca de Peñagolosa, Teruel). Geogaceta, 52, 181–184.
Cuesta, E., Vidal, D., Ortega, F., Shibata, M., & Sanz, J. L. (2021). Pelecanimimus (Theropoda: Ornithomimosauria) postcranial anatomy and the evolution of the specialized manus in ornithomimosaurs and sternum in maniraptoriforms. Zoological Journal of the Linnean Society. https://doi.org/10.1093/zoolinnean/zlab013
Díaz-Martínez, I., Pereda-Suberbiola, X., Pérez-Lorente, F., & Canudo, J. I. (2015). Ichnotaxonomic review of large ornithopod dinosaur tracks: Temporal and geographic implications. PLoS ONE, 10(2), e0115477. https://doi.org/10.1371/journal.pone.0115477
Dieudonné, P. E., Tortosa, T., Fernández-Baldor, F. T., Canudo, J. I., & Díaz-Martínez, I. (2016). An unexpected early rhabdodontid from Europe (Lower Cretaceous of Salas de los Infantes, Burgos Province, Spain) and a re-examination of basal iguanodontian relationships. PLoS ONE, 11(6), e0156251.
Dollo, L. (1888). Iguanodontidae et Camptonotidae. Comptes Rendus De Academie Des Sciences, Paris, 106, 775–777.
Fernández-Baldor, F. T., Canudo, J. I., Huerta, P., Montero, D., Suberbiola, X. P., & Salgado, L. (2011). Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontologica Polonica, 56(3), 535–552.
Figueiredo, S. D., Carvalho, I. S., Pereda-Suberbiola, X., Cunha, P. P., Antunes, V., & Díaz-Martínez, I. (2022). New ornithopod footprints from the Areia do Mastro Formation (Lower Cretaceous), Espichel Cape (Portugal, Western Iberia) and their context in the Iberian ichnological ornithopod record. Cretaceous Research. https://doi.org/10.1016/j.cretres.2021.105069
Fuentes-Vidarte, C., Meijide-Calvo, M., Meijide-Fuentes-Vidarte, F., & Meijide-Fuentes-Vidarte, M. (2016). Un nuevo dinosaurio estiracosterno (Ornithopoda: Ankylopollexia) del Cretácico Inferior de España. Spanish Journal of Paleontology, 31(2), 407–446.
Galton, P. M. (1974). The ornithischian dinosaur Hypsilophodon from the Wealden of the Isle of Wight. Bulletin of the British Museum (Natural History). Geology, 25, 1–152.
Galton, P. M. (2009). Notes on Neocomian (Lower Cretaceous) ornithopod dinosaurs from England—Hypsilophodon, Valdosaurus, “Camptosaurus”, “Iguanodon”—and referred specimens from Romania and elsewhere. Revue De Paléobiologie, 28, 211–273.
Galton, P. M. (2012). Hypsilophodon foxii and other smaller bipedal ornithischian dinosaurs from the Lower Cretaceous of Southern England. In P. Godefroit (Ed.), Bernissart dinosaurs and Early Cretaceous terrestrial ecosystems (pp. 225–281). Indiana University Press.
Gasca, J. M. (2011). Descripción de una tibia de dinosaurio ornitópodo de la Formación El Castellar (Cretácico Inferior, Teruel). In J. Marigó, M. Pérez de los Ríos, D. de Miguel, & A. Bolet (Eds.), Libro de Resúmenes de las XXVII Jornadas de la Sociedad Española de Paleontología (pp. 179–183). Sabadell: Institut Català de Paleontologia Miquel Crusafont.
Gasca, J. M., Canudo, J. I., & Moreno-Azanza, M. (2014a). On the diversity of Iberian iguanodont dinosaurs: New fossils from the lower Barremian, Teruel province, Spain. Cretaceous Research, 50, 264–272. https://doi.org/10.1016/j.cretres.2014.05.009
Gasca, J. M., Canudo, J. I., & Moreno-Azanza, M. (2014b). A large-bodied theropod (Tetanurae: Carcharodontosauria) from the Mirambel Formation (Barremian) of Spain. Neues Jahrbuch Für Geologie Und Paläontologie-Abhandlungen, 273(1), 13–23.
Gasca, J. M., Díaz-Martínez, I., Moreno-Azanza, M., Canudo, J. I., & Alonso, A. (2018). A hypertrophied ungual phalanx from the lower Barremian of Spain: Implications for the diversity and palaeoecology of Spinosauridae (Theropoda) in Iberia. Cretaceous Research, 84, 141–152.
Gasca, J. M., Moreno-Azanza, M., & Canudo, J. I. (2008). Dientes de dinosaurios terópodos espinosáuridos de la Formación El Castellar (Cretácico Inferior, Teruel). Paleontologica Nova, SEPAZ, 8, 233–234.
Gasulla, J. M., Escaso, F., Narváez, I., Ortega, F., & Sanz, J. L. (2015). A new sail-backed styracosternan (Dinosauria: Ornithopoda) from the Early Cretaceous of Morella, Spain. PLoS ONE, 10(12), e0144167. https://doi.org/10.1371/journal.pone.0144167
Gasulla, J. M., Escaso, F., Ortega, F., & Sanz, J. L. (2014). New hadrosauriform cranial remains from the Arcillas de Morella Formation (lower Aptian) of Morella, Spain. Cretaceous Research, 47, 19–24. https://doi.org/10.1016/j.cretres.2013.10.004
Gasulla, J. M., Ortega, F., Pereda-Suberbiola, X., Escaso, F., & Sanz, J. L. (2011). Elementos de la armadura dérmica del dinosaurio anquilosaurio Polacanthus Owen, 1865, en el Cretácico Inferior de Morella (Castellón, España). Ameghiniana, 48(4), 508–519.
Gilmore, C. W. (1909). Osteology of the Jurassic reptile Camptosaurus, with a revision of the species of the genus, and descriptions of two new species. Proceedings of the United States National Museum, 36, 197–332.
Godefroit, P., Dong, Z. M., Bultynck, P., Li, H., & Feng, L. (1998). New Bactrosaurus (Dinosauria: Hadrosauroidea) material from Iren Dabasu (Inner Mongolia, PR China). Bulletin De L’institut Royal Des Sciences Naturelles De Belgique, Sciences De La Terra, 68(Supplement), 3–70.
Griffin, C. T., Stocker, M. R., Colleary, C., Stefanic, C. M., Lessner, E. J., Riegler, M., et al. (2020). Assessing ontogenetic maturity in extinct saurian reptiles. Biological Reviews. https://doi.org/10.1111/brv.12666
Guerrero, A., & Cobos, A. (2017). Nuevos fósiles de dinosaurios cretácicos del yacimiento La Balsa 2 (El Castellar, Teruel, España). In S. Barrios de Pedro, C. Blanco-Moreno, A. de Celis, J. Colmenar, E. Cuesta, D. García-Martínez, et al. (Eds.), A Glimpse of the Past (pp. 15–19). Lisboa: Abstract book of the XV Encuentro de Jóvenes Investigadores en Paleontología.
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4(1), 1–9.
Horner, J. R., Weishampel, D. B., & Forster, C. A. (2004). Hadrosauridae. In D. B. Weishampel, P. Donson, & H. Osmólska (Eds.), The Dinosauria (2nd ed., pp. 438–463). University of California Press.
Hübner, T. (2018). The postcranial ontogeny of Dysalotosaurus lettowvorbecki (Ornithischia: Iguanodontia) and implications for the evolution of ornithopod dinosaurs. Palaeontographica Abteilung A, 310, 43–120. https://doi.org/10.1127/pala/2018/0072
Ikejiri, T. (2012). Histology-based morphology of the neurocentral synchondrosis in Alligator mississippiensis (Archosauria, Crocodylia). The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 295(1), 18–31.
Ikejiri, T. (2015). Modes of ontogenetic allometric shifts in crocodilian vertebrae. Biological Journal of the Linnean Society, 116, 649–670.
Isasmendi, E., Sáez-Benito, P., Torices, A., Navarro-Lorbés, P., & Pereda-Suberbiola, X. (2020). New insights about theropod palaeobiodiversity in the Iberian Peninsula and Europe: Spinosaurid teeth (Theropoda, Megalosauroidea) from the Lower Cretaceous of La Rioja (Spain). Cretaceous Research, 116, 104600. https://doi.org/10.1016/j.cretres.2020.104600
Knoll, F. (2009). A large iguanodont from the upper Barremian of the Paris Basin. Geobios, 42, 755–764. https://doi.org/10.1016/j.geobios.2009.06.002
Leonardi, G., et al. (1984). Le impronte fossili dei dinosauri. In J. F. Bonaparte, E. H. Colbert, P. H. Currie, A. de Ricqles, Z. Kielan-Jaworowska, & G. Leonardi (Eds.), Sulle Orme dei dinosauri (pp. 161–186). Erizzo Editrice.
Llandres, M., Vullo, R., Marugán-Lobón, J., Ortega, F., & Buscalioni, A. D. (2013). An articulated hindlimb of a basal iguanodont (Dinosauria, Ornithopoda) from the Early Cretaceous Las Hoyas Lagerstätte (Spain). Geological Magazine, 150(3), 572–576. https://doi.org/10.1017/S0016756813000095
Lockley, M. G., Xing, L., Lockwood, J. A. F., & Pond, S. (2014). A review of large Cretaceous ornithopod tracks, with special reference to their ichnotaxonomy. Biological Journal of the Linnean Society, 113, 721–736.
Lockley, M. G., Young, B. H., & Carpenter, K. (1983). Hadrosaur locomotion and herding behavior: Evidence from footprints in the Mesaverde Formation, Grand Mesa Coal Field, Colorado. The Mountain Geologist, 20(4), 5–14.
Lockwood, J. A. F., Martill, D. M., & Maidment, C. R. (2021). A new hadrosauriform dinosaur from the Wessex Formation, Wealden Group (Early Cretaceous), of the Isle of Wight, southern England. Journal of Systematic Palaeontology. https://doi.org/10.1080/14772019.2021.1978005
Luque, L., Espílez, E., Cobos, A., Alcalá, L., Royo-Torre, R., Aberasturi, A., et al. (2006). Iguanodóntidos y otros vertebrados cretácicos de Miravete de la Sierra (Teruel). Teruel, 91(1), 57–87.
Madzia, D., Arbour, V. M., Boyd, C. A., Farke, A. A., Cruzado-Caballero, P., & Evans, D. C. (2021). The phylogenetic nomenclature of ornithischian dinosaurs. PeerJ, 9, e12362. https://doi.org/10.7717/peerj.12362
Malafaia, E., Gasulla, J. M., Escaso, F., Narvaéz, I., & Ortega, F. (2020a). An update of the spinosaurid (Dinosauria: Theropoda) fossil record from the Lower Cretaceous of the Iberian Peninsula: Distribution, diversity, and evolutionary history. Journal of Iberian Geology, 46(4), 431–444.
Malafaia, E., Gasulla, J. M., Escaso, F., Narváez, I., Sanz, J. L., & Ortega, F. (2020b). A new spinosaurid theropod (Dinosauria: Megalosauroidea) from the upper Barremian of Vallibona, Spain: Implications for spinosaurid diversity in the Early Cretaceous of the Iberian Peninsula. Cretaceous Research, 106, 104221. https://doi.org/10.1016/j.cretres.2019.104221
Mantell, G. A. (1825). Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex. Philosophical Transactions of the Royal Society, 115, 179–186. https://doi.org/10.1098/rstl.1825.0010
Marsh, O. (1881). Classification of the Dinosauria. American Journal of Sciences, 3, 81–86.
Martín-Closas, C. (1989). Els caròfits del Cretaci Inferior de les conques perifèriques del Bloc de l’Ebre. PhD Thesis, Universidad de Barcelona, pp. 1–581.
McDonald, A. T. (2011). The taxonomy of species assigned to Camptosaurus (Dinosauria: Ornithopoda). Zootaxa, 2783(1), 52–68. https://doi.org/10.11646/zootaxa.2783.1.4
McDonald, A. T., Bird, J., Kirkland, J. I., & Dodson, P. (2012). Osteology of the basal hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah. PLoS ONE, 7, e45712.
McDonald, A. T., Kirkland, J. I., DeBlieux, D. D., Madsen, S. K., Cavin, J., et al. (2010). New basal iguanodonts from the Cedar Mountain Formation of Utah and the evolution of thumb-spiked dinosaurs. PLoS ONE, 5(11), e14075. https://doi.org/10.1371/journal.pone.0014075
McDonald, A. T., Maidment, S. C., Barrett, P. M., You, H. L., & Dodson, P. (2014). Osteology of the basal hadrosauroid Equijubus normani (Dinosauria, Ornithopoda) from the Early Cretaceous of China. In D. A. Eberth & D. C. Evans (Eds.), Hadrosaurs (pp. 3–34). Indiana University Press.
Meyer, C. A., & Thüring, B. (2003). The first iguanodontid dinosaur tracks from the Swiss Alps (Schrattenkalk Formation, Aptian). Ichnos, 10, 221–228.
Norman, D. B. (1980). On the ornithischian dinosaurs Iguanodon bernissartensis from the Lower Cretaceous of Bernissart (Belgium). Memoires De L’institut Royal Des Sciences Naturelles De Belgique, 178, 1–103.
Norman, D. B. (1986). On the anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda). Bulletin De L’institut Royal Des Sciences Naturelles De Belgique, 56, 281–372.
Norman, D. B. (2002). On Asian ornithopods (Dinosauria: Ornithischia). Probactrosaurus Rozhdestvensky, 1966. Zoological Journal of the Linnean Society, 136, 113–144.
Norman, D. B. (2004). Basal iguanodontia. In D. B. Weishampel, P. Donson, & H. Osmólska (Eds.), The Dinosauria (pp. 413–437). University of California Press.
Norman, D. B. (2011). On the osteology of the lower Wealden (Valanginian) iguanodontian Barilium dawsoni (Iguanodontia: Styracosterna). Special Papers in Palaeontology, Palaeontological Association, 86, 165–198.
Norman, D. B. (2012). Iguanodontian taxa (Dinosauria: Ornithischia) from the Lower Cretaceous of Britain and Belgium. In P. Godefroit (Ed.), Bernissart dinosaurs and Early Cretaceous terrestrial ecosystems (pp. 174–212). Indiana University Press.
Norman, D. B. (2014). Iguanodonts from the Wealden of England: Do they contribute to the discussion concerning hadrosaur origins? In D. A. Eberth & D. C. Evans (Eds.), Hadrosaurs (life of the past) (pp. 10–43). Indiana University Press.
Norman, D. B. (2015). On the history, osteology, and systematic position of the Wealden (Hastings group) dinosaur Hypselospinus fittoni (Iguanodontia: Styracosterna). Zoological Journal of the Linnean Society, 173(1), 92–189.
Ortega, F., Escaso, F., & Sanz, J. L. (2010). A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain. Nature, 467, 203–206.
Ösi, A., Prondvai, E., Butler, R., & Weishampel, D. B. (2012). Phylogeny, histology and inferred body size evolution in a new rhabdodontid dinosaur from the Late Cretaceous of Hungary. PLoS ONE, 7(9), e44318. https://doi.org/10.1371/journal.pone.0044318
Owen, R. (1842). Report on British fossil reptiles, Part II. In: Report of the Eleventh Meeting of the British Association Advancement of the Science Vol. 1841 (pp. 60–204).
Paul, G. S. (2008). A revised taxonomy of the iguanodont dinosaur genera and species. Cretaceous Research, 29, 192–216. https://doi.org/10.1016/j.cretres.2007.04.009
Pereda-Suberbiola, X., Fuentes, C., Meijide, M., Meijide-Fuentes, F., & Meijide-Fuentes, M., Jr. (2007). New remains of the ankylosaurian dinosaur Polacanthus from the Lower Cretaceous of Soria, Spain. Cretaceous Research, 28(4), 583–596.
Pereda-Suberbiola, X., Ruiz-Omeñaca, J. I., Canudo, J. I., Torcida, F., & Sanz, J. L. (2012). Dinosaur faunas from the Early Cretaceous (Valanginian-Albian) of Spain. In P. Godefroit (Ed.), Bernissart dinosaurs and early cretaceous terrestrial ecosystems (pp. 379–407). Indiana University Press.
Pereda-Suberbiola, X., Ruiz-Omeñaca, J. I., Fernández-Baldor, F. T., Maisch, M. W., Huerta, P., Contreras, R., et al. (2011). A tall-spined ornithopod dinosaur from the Early Cretaceous of Salas de los Infantes (Burgos, Spain). Comptes Rendus Palevol, 10, 551–558. https://doi.org/10.1016/j.crpv.2011.04.003
Pérez-García, A., Cobos, A., & Royo-Torres, R. (2017). The oldest evidence of a dortokid turtle (stem Pleurodira) from the uppermost Hauterivian-basal Barremian El Castellar Formation (Teruel, Spain). Journal of Iberian Geology, 43(2), 139–146.
Pérez-García, A., Scheyer, T. M., & Murelaga, X. (2013). The turtles from the uppermost Jurassic and Early Cretaceous of Galve (Iberian Range, Spain): Anatomical, systematic, biostratigraphic and palaeobiogeographical implications. Cretaceous Research, 44, 64–82.
Pérez-Moreno, B. P., Sanz, J. L., Buscalioni, Á. D., Moratalla, J. J., Ortega, F., & Rasskin-Gutman, D. (1994). A unique multitoothed ornithomimosaur dinosaur from the Lower Cretaceous of Spain. Nature, 370, 363–367.
Prieto-Márquez, A., & Norell, M. A. (2010). Anatomy and relationships of Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia. American Museum Novitates, 3694, 1–49.
Royo-Torres, R., & Cobos, A. (2007). Teeth of Oplosaurus armatus (Sauropoda) from El Castellar (Teruel, Spain). In: J. Le Loeuf, (Ed.), Abstract book of European Association of Vertebrate Paleontologists 5th anual meeting (pp. 52–55). Francia: Carcassonne.
Royo-Torres, R., Fuentes, C., Meijide, M., Meijide-Fuentes, F., & Meijide-Fuentes, M. (2017). A new brachiosauridae sauropod dinosaur from the Lower Cretaceous of Europe (Soria province, Spain). Cretaceous Research, 80, 38–55. https://doi.org/10.1016/j.cretres.2017.08.012
Ruiz-Omeñaca, J. I., Canudo, J. I., Cuenca-Bescós, G., Cruzado-Caballero, P. L., Gasca, J. M., & Moreno-Azanza, M. (2012). A new basal ornithopod dinosaur from the Barremian of Galve, Spain. Comptes Rendus Palevol, 11(6), 435–444. https://doi.org/10.1016/j.crpv.2012.06.001
Salas, R. (1987). El Malm i el Cretaci inferior entre el Massif de Garraf i la Serra d´Espadà. Anàlisi de conca. PhD Thesis, Universidad de Barcelona, pp. 1–365.
Salas, R., & Guimerà, J. (1996). Rasgos estructurales principales de la cuenca Cretácica Inferior del Maestrazgo (Cordillera Ibérica oriental). Geogaceta, 20, 1704–1706.
Salas, R., Guimerà, J., Mas, R., Martín-Closas, C., Meléndez, A., & Alonso, A. (2001). Peri-Tethys Memoir 6: Peri-Tethyan Rift/Wrench Basins and passive margins. In P. A. Ziegler, W. Cavazza, A. F. H. Robertson, & S. Crasquin-Soleau (Eds.), Mémoires du Muséum national d‘Histoire naturelle (pp. 145–185). Editions du Muséum.
Sanguino, F., & Buscalioni, Á. D. (2018). The Iguanodon locality of Barranco Pata la Mona (upper Barremian, Buenache de la Sierra, Cuenca) revisited. In E. Amayuelas, P. Bilbao-Lasa, O. Bonilla, M. del Val, J. Errandonea-Martin, I. Garate-Olave, et al. (Eds.), Life finds a way (pp. 103–106). Gasteiz: Abstract book of the XVI Encuentro de Jóvenes Investigadores en Paleontología.
Santos-Cubedo, A., de Santisteban, C., Poza, B., & Meseguer, S. (2021). A new styracosternan hadrosauroid (Dinosauria: Ornithischia) from the Early Cretaceous of Portell, Spain. PLoS ONE, 16(7), e0253599. https://doi.org/10.1371/journal.pone.0253599
Sanz, J. L., Casanovas, M. I., & Santafé, J. V. (1982). Paleontología. In J. V. Santafé, M. L. Casanovas, J. L. Sanz, & S. Calzada (Eds.), Geología y paleontología (dinosaurios) de las capas rojas de Morella (Castellón, España) (pp. 69–169). Diputación Provincial de Castellón y Diputación de Barcelona.
Schudack, U., & Schudack, M. (2009). Ostracod biostratigraphy in the Lower Cretaceous of the Iberian chain (eastern Spain). Journal of Iberian Geology, 35, 141–168.
Seeley, H. G. (1887). On the classification of the fossil animals commonly named Dinosauria. Proceedings of the Royal Society of London, 43, 165–171. https://doi.org/10.1098/rspl.1887.0117
Sereno, P. C. (1986). Phylogeny of the bird-hipped dinosaurs (Order Ornithischia). National Geographic Research, 2, 234–256.
Sereno, P. C. (1997). The origin and evolution of dinosaurs. Annual Review of Earth and Planetary Sciences, 25(1), 435–489.
Sigovini, M., Keppel, E., & Tagliapietra, D. (2016). Open Nomenclature in the biodiversity era. Methods in Ecology and Evolution, 7(10), 1217–1225.
Taquet, P. (1976). Geologie et paleontologie du gisement de Gadoufaoua (Aptien du Niger), Cahiers Paléontologie (pp. 1–191). es: Centre National de la Recherche Scientifiqu.
Torrente, M. Á., Verdú, F. J., Cobos, A., & Alcalá, L. (2018). Diente de un coelurosaurio (Theropoda, Dinosauria) del Barremiense (Cretácico Inferior) de El Castellar (Teruel, España). Geogaceta, 63, 79–82.
Verdú, F. J. (2017). Sistemática, filogenia y paleobiología de Iguanodon galvensis (Ornithopoda, Dinosauria) del Barremiense inferior (Cretácico Inferior) de Teruel (España). PhD Thesis, Universitat de València, pp. 1–445.
Verdú, F. J., Cobos, A., Royo-Torres, R., & Alcalá, L. (2019). Diversity of large ornithopod dinosaurs in the Upper Hauterivian-Lower Barremian (Lower Cretaceous) of Teruel, Spain: A morphometric approach. Spanish Journal of Palaeontology, 34(2), 269–288.
Verdú, F. J., Godefroit, P., Royo-Torres, R., Cobos, A., & Alcalá, L. (2017). Individual variation in the postcranial skeleton of the Early Cretaceous Iguanodon bernissartensis (Dinosauria: Ornithopoda). Cretaceous Research, 74, 65–86. https://doi.org/10.1016/j.cretres.2017.02.006
Verdú, F. J., Royo-Torres, R., Cobos, A., & Alcalá, L. (2015). Perinates of a new species of Iguanodon (Ornithischia: Ornithopoda) from the lower Barremian of Galve (Teruel, Spain). Cretaceous Research, 56, 250–264.
Verdú, F. J., Royo-Torres, R., Cobos, A., & Alcalá, L. (2018). New systematic and phylogenetic data about the early Barremian Iguanodon galvensis (Ornithopoda: Iguanodontoidea) from Spain. Historical Biology, 30, 437–474. https://doi.org/10.1080/08912963.2017.1287179
Verdú, F. J., Royo-Torres, R., Cobos, A., & Alcalá, L. (2020). Systematics and paleobiology of a new articulated axial specimen referred to Iguanodon cf. galvensis (Ornithopoda, Iguanodontoidea). Journal of Vertebrate Paleontology, 40(6), 8202. https://doi.org/10.1080/02724634.2021.1878202
Vialov, O. S. (1988). On the classification of dinosaurian traces. Ezhegodnik Vsesoyuznogo Paleontologicheskogo Obshchestva, 31, 322–325.
Villanueva-Amadoz, U., Sender, L. M., Royo-Torres, R., Verdú, F. J., Pons, D., Alcalá, L., et al. (2015). Palaeobotanical remains associated with dinosaur fossils from the Camarillas Formation (Barremian) of Galve (Teruel, Spain). Historical Biology, 27, 374–388.
Weishampel, D. B., Jianu, C. M., Cziki, Z., & Norman, D. B. (2003). Osteology and phylogeny of Zalmoxes (n. g.), an unusual ornithopod dinosaur from the latest Cretaceous of Romania. Journal of Systematic Paleontology, 1, 123–143. https://doi.org/10.1017/S1477201903001032
Acknowledgements
This research was funded by the Departamento de Educación, Cultura y Deporte (Gobierno de Aragón), Research Group E04_20R FOCONTUR financed by Departamento de Ciencia, Universidad y Sociedad del Conocimiento (Gobierno de Aragón), the Instituto Aragonés de Fomento, the Ministerio de Ciencia e Innovación (MCI), Agencia Estatal de Investigación (AEI), and Fondo Europeo de Desarrollo Regional (FEDER) from Spanish Government (PGC2018-094034-B-C22 and Unidad de Paleontología de Dinosaurios de Teruel). The authors thank Pascal Godefroit, Annelise Folie and all the RBINS (Royal Belgian Institute of Natural Sciences) staff for providing access to the Bernissart iguanodontian collection on 2016. We appreciate useful comments and suggestions made by the associate editor Laura Domingo, and the two reviewers Fernando Escaso and anonymous. Finally, we would like to express our gratitude to Mr. Gabriel Fuentes Bellido, neighbour of Cabra de Mora, for donating part of the material studied here to the Museo Aragonés de Paleontología (MAP).
Funding
This work was supported by the Departamento de Educación, Cultura y Deporte (Gobierno de Aragón), Research Group E04_20R FOCONTUR financed by Departamento de Ciencia, Universidad y Sociedad del Conocimiento (Gobierno de Aragón), the Instituto Aragonés de Fomento, the Ministerio de Ciencia e Innovación (MCI), Agencia Estatal de Investigación (AEI), and Fondo Europeo de Desarrollo Regional (FEDER) from Spanish Government (PGC2018-094034-B-C22 and Unidad de Paleontología de Dinosaurios de Teruel).
Author information
Authors and Affiliations
Contributions
Not applicable for that section.
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that they have no conflict of interest.
Ethics approval
Not applicable for that section.
Consent to participate
Informed consent was obtained from all individual participants included in the study. All authors consent to the publication of the study.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
García-Cobeña, J., Verdú, F.J. & Cobos, A. Abundance of large ornithopod dinosaurs in the El Castellar Formation (Hauterivian-Barremian, Lower Cretaceous) of the Peñagolosa sub-basin (Teruel, Spain). J Iber Geol 48, 107–127 (2022). https://doi.org/10.1007/s41513-021-00185-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41513-021-00185-w