
Abstract
The stratigraphic distribution of larger benthic foraminifera and other microfossils, as analysed in thin-sections, has been investigated in three stratigraphic sections from the island of Mallorca (Son Maina, S’Heretat and Cuevas de Artà). These stratigraphic sections represent deposition in a Lower Jurassic shallow-water carbonate platform succession (Es Barraca Member, Soller Formation) of the Llevant Mountains. The microfossil assemblages contain abundant benthic foraminifera and calcareous algae, including the typical Liassic dasycladalean alga Palaeodasycladus mediterraneus (Pia) and the microproblematica Thaumatoporella parvovesiculifera (Raineri), which provide good age constraint of the succession. Three consecutive biozones have been recognized based on the stratigraphic distribution of foraminifera, including the evolutionary bioseries Mesoendothyra—Lituosepta—Orbitopsella spp., which are documented for the first time in the Balearic Basin: Biozone A (interval Zone) is characterized by the occurrence of small Siphovalvulina spp., Mesoendothyra sp. and rare Lituosepta ancestors. Biozone B (lineage Zone), whose lower boundary is defined by the first occurrence of Lituosepta recoarensis Cati and its top by the first appearance of Orbitopsella aff. primaeva (Henson). The uppermost part of biozone B shows an interval with transitional morphotypes between Lituosepta and Orbitopsella, indicating that the change between biozones B and C1 is gradual (B/C1 transition). This interval is characterized by the presence of very primitive forms of Orbitopsella. Finally, Biozone C1 (lineage Zone) is defined by the first occurrence of Orbitopsella aff. primaeva together with L. recoarensis. The upper boundary of the Biozone C1, which is defined by the first occurrence of O. praecursor s.l., is not recorded in the studied sections of the Es Barraca Member. This biostratigraphic zonation is consistent with a Sinemurian age for the Es Barraca Member with its top most likely not extending into the Pliensbachian. The proposed biostratigraphic scheme is comparable with those established for other western Tethyan margins such as the High Atlas of Morocco, the Southern Italian Alps or the Dinaric Alps of Slovenia and Croatia. The reconstruction of a depositional transect across the studied sections shows the progressive loss of the upper biozones towards the northeast, demonstrating the existence of significant hiatuses in some sections of the Llevant Mountains domain. These hiatuses point to an intra-lower Pliensbachian (Carixian) early stage of platform fragmentation, with areas affected by erosion or non-deposition, and areas with marine marly limestone sedimentation followed by deltaic siliciclastic progradation on the north-western part of Mallorca.
Resumen
Se ha investigado, mediante el análisis de láminas delgadas, la distribución estratigráfica de foraminíferos bentónicos y otros microfósiles en tres secciones estratigráficas del Jurásico Inferior de la Sierra de Levante, en la isla de Mallorca (Son Maina, S’Heretat y Cuevas de Artà). Estas secciones representan el depósito, durante el Jurásico Inferior, de un sistema de plataforma carbonatada somera (Miembro Es Barraca de la Formación Soller). La asociación de microfósiles contiene abundantes foraminíferos bentónicos y algas calcáreas, entre las que se incluyen Palaeodasycladus mediterraneus (Pia) del Lías y la microproblemática Thaumatoporella parvovesiculifera (Raineri), lo que proporciona un buen control de edad de la sucesión. En base a la distribución estratigráfica de los foraminíferos y la bioserie evolutiva Mesoendothyra – Lituosepta – Orbitopsella spp., se han reconocido y documentado por primera vez, para el Miembro Es Barraca, tres biozonas consecutivas: Biozona A (Zona de intervalo), caracterizada por la existencia de pequeños ejemplares de Siphovalvulina spp., Mesoendothyra sp. y escasos ejemplares de Lituosepta ancestral. Biozona B (Zona de linaje), cuyo límite inferior está definido por la primera aparición de Lituosepta recoarensis Cati, mientras que su límite superior se caracteriza por la primera aparición de Orbitopsella aff. primaeva (Henson). La parte superior de la biozona B muestra un intervalo con morfotipos transicionales entre Lituosepta y Orbitopsella, indicando que el paso entre las biozonas B y C1 es gradual (transición B/C1). Este intervalo se caracteriza por la presencia de formas muy primitivas de Orbitopsella. Finalmente, la Biozona C1 (Zona de linaje) está definida por la primera aparición de Orbitopsella aff. primaeva junto con L. recoarensis. El límite superior de la Biozona C1, definido por la primera aparición de O. praecursor s.l., no se ha reconocido en las secciones estudiadas del Miembro Es Barraca. Esta zonación bioestratigráfica es compatible con una edad Sinemuriense para el Miembro Es Barraca y sugiere que el techo de la unidad muy probablemente no alcanza el Pliensbachiense. El esquema bioestratigráfico propuesto es comparable con el establecido para otros márgenes del Tethys occidental tales como el Alto Atlas en Marruecos, los Alpes del sur de Italia o los Alpes Dináricos de Eslovenia y Croacia. La reconstrucción de un transecto estratigráfico a lo largo de las secciones estudiadas muestra la pérdida progresiva de las biozonas superiores hacia el noreste, demostrando así la existencia de hiatos significativos en algunas secciones del dominio de Sierras de Levante. Estos hiatos apuntan a una fragmentación temprana de la plataforma (intra-Pliensbachiense inferior) con áreas afectadas por erosión o no-depósito, y otras áreas, al noroeste de Mallorca, con una sedimentación continua de margocaliza seguida de una progradación deltaica siliciclástica.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
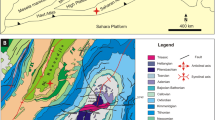
Shallow-marine carbonates developed widely on the western Tethyan margins during the Early Jurassic (Fig. 1). These shallow-water carbonate environments experienced important changes in platform architecture and sediment composition in the context of rifting related with the break-up of Pangea and are associated with important oceanic circulation and environmental changes, accompanied by eustatic sea-level rise (e.g. Zempolich 1993; Ruiz-Ortiz et al. 2004; Masetti et al. 2016; Preto et al. 2017; Rychlinski et al. 2018; Franceschi et al. 2019). These events also coincided with the Early Jurassic faunistic recovery after the Triassic–Jurassic (T/J) boundary mass extinction in the marine realm. The gradual recovery and diversification of the shallow-water benthic communities during the Early Jurassic was characterized by the appearance of the Jurassic larger benthic foraminifera (LBF), which underwent significant evolutionary developments. The poorly diversified benthic foraminifera associations of the Hettangian evolved during the early Sinemurian into more diversified Siphovalvulina-dominated assemblages, and finally into assemblages with more structurally complicated lituolids during the late Sinemurian–Pliensbachian, providing an important biostratigraphic tool (Hottinger 1967; Septfontaine 1984, 1988; Bassoullet 1997; Fugagnoli and Loriga Broglio1998; Kabal and Tasli 2003; Fugagnoli 2004; Mancinelli et al. 2005; BouDagher-Fadel and Bosence 2007; Velić 2007; Gale 2014). Several biostratigraphic schemes based on LBF have been proposed for the Early Jurassic (Fig. 2) (e.g. Septfontaine 1984; Kabal and Tasli 2003; Velić 2007; BouDagher-Fadel and Bosence 2007; Gale 2014). In this study, the biostratigraphic scale proposed by Septfontaine (1984) for the Sinemurian–Pliensbachian has been adopted (Fig. 2). The Septfontaine (1984) biostratigraphic scheme, based on data from Morocco calibrated with ammonite zones, established six biozones for the Sinemurian–Pliensbachian interval that in stratigraphic order are: A, B, C1, C2, D and E. The Sinemurian–Pliensbachian transition is included within the lower part of the Biozone C1 (Fig. 2). This biozonation was later recognised in the Dinarides by Velić (2007) and Gale (2014), in the Dinaric Alps of Croatia (e.g. Veseli et al. 2006), and more recently applied in the Southern Alps of Italy (Franceschi et al. 2019), confirming that this biozonation is still valid and useful for the Tethyan realm.

(modified from Dercourt et al. 2000)
Paleogeographical map of the western Tethys for the Sinemurian–earliest Pliensbachian showing the paleogeographical location of the studied area. Yellow star: Location of the studied area (BB Balearic Basin). Red dots: Location of other neighbouring basins (HA High Atlas, IbR Iberian Range, BC Betic Cordillera, ApCP Apennine carbonate platform, AdCP Adriatic carbonate platform)

Comparison between the standard ammonite biozones and various benthic foraminiferal biostratigraphic schemes for the Sinemurian–Pliensbachian interval proposed for different carbonate platforms of the Western Tethys. Age (Ma.) after Ogg et al. (2016)
The Lower Jurassic succession of the Balearic Islands (Fig. 3) is a good representation of a wide shallow-marine carbonate platform that was fragmented into depositional highs and lows during the Pliensbachian and afterwards submerged and drowned (Álvaro et al. 1989; Rosales et al. 2018; Sevillano et al. 2019). A comprehensive study of facies and platform evolution of the shallow-water carbonates (Es Barraca Member, Soller Formation) has recently been provided by Sevillano et al. (2019). The microfossil content of the Es Barraca Member was previously studied by Fallot (1922), Colom (1966, 1970, 1980), and Colom and Dufaure (1962), but no detailed stratigraphic distribution of the LBF or biozonation was completed. The new micropaleontological study presented here has three aims: (1) to develop an LBF-based biostratigraphy for Early Jurassic of Mallorca; (2) to follow changes in the studied samples of the gradual evolution of the mesoendothyrids with the aim of using this as a biostratigraphic tool (Septfontaine 1984, 1988); (3) to use the new biostratigraphic information to obtain a better understanding of the evolution of the carbonate platform and stratigraphy during the Early Jurassic. In addition, this study improves the knowledge of the paleobiogeographical distribution of the Lower Jurassic foraminiferal species in the western Tethys of the eastern Iberian Margin, allowing for a better comparison with other Tethyan regions.

Modified from Álvaro et al. (1989)
General chronostratigraphic chart of the Lower Jurassic of Mallorca with indication of the defined lithostratigraphic units.
2 Geological setting
The Jurassic sedimentary succession of Mallorca was deposited in the Balearic Basin, located on the eastern margin of the Iberian Plate. This paleogeographic position was adjacent to the emerged Ebro High (Fig. 1; Thierry 2000; Scotese and Schettino 2017). The Jurassic sedimentary evolution of the Balearic Basin related to the opening of the Central Atlantic Ocean and the tectonic evolution of the western Tethyan rift (Dewey et al. 1973; Dercourt et al. 2000). Although the Balearic archipelago has traditionally been considered as the north-eastern prolongation of the Betic Cordillera (BC in Fig. 1), new studies suggest that the geodynamic evolution of the area also seems to be related to the early opening of the Gulf of Valencia. In this area recent offshore seismic surveys have highlighted the existence of a thick submerged Mesozoic rift basin, whose south-eastern border is represented by the Balearic archipelago (Etheve et al. 2018). The symmetrical north-western basin margin would be localised on the Iberian margin. The Balearic segment of this Mesozoic rift basin margin formed part of a larger rift system related to the opening of the Alpine-Ligurian Tethys.
On Mallorca, the largest island of the archipelago, the rifting evolution during the Jurassic recorded the following main sedimentary events (Barnolas and Simó 1984; Álvaro et al. 1989; Barnolas et al. 2010): (1) Development of shallow-water platform successions (Hettangian–lower Pliensbachian). (2) Rifting with formation of blocks and a drowning succession that includes hardgrounds (upper Pliensbachian–lower Bajocian). (3) Localised hemipelagic sedimentation (Toarcian– Bajocian). (4) Typical ammonitico rosso-type facies, radiolarites, and mass-wasting deposits indicative of deeper water slope, base of slope and basin environments (Middle and Late Jurassic).
The depositional setting of the Lower Jurassic shallow-water succession deposited on Mallorca includes (Álvaro et al. 1989; Fig. 3): (1) Hettangian sabkha to restricted platform dolomites of the Mal Pas Formation; (2) Sinemurian to lower Pliensbachian shallow-platform carbonates and siliciclastics of the Soller Formation; (3) upper Pliensbachian open platform calcarenites and marly limestones of the Es Cosconar Formation. The Soller Formation is in turn subdivided into three members (Fig. 3): Es Barraca Member, dominated by shallow-water peritidal and lagoonal limestones; Sa Moleta Member, consisting of outer platform shales-siltstones and marly limestones; and Es Racó Member, consisting of deltaic siliciclastics.
In the Es Barraca Member, the focus of this study, previous studies (Colom 1970; Álvaro et al. 1989) reported Palaeodasycladus mediterraneus (Pia), Thaumatoporella parvovesiculifera (Raineri), Lituosepta recoarensis Cati, and Orbitopsella praecursor (Gümbel). Based on these micropaleontological occurrences and a deduced lateral relationship with the Sa Moleta Member (lower Carixian), Álvaro et al. (1989) established the age of the Es Barraca Member as Sinemurian-lower Pliensbachian (lower Carixian; Fig. 3). The Sa Moleta Member is attributed to the Jamesoni and Ibex ammonite Zones of the early Pliensbachian (early Carixian), based on brachiopods and a scarce ammonite fauna (Colom 1942; Álvaro et al. 1989), whereas the Es Racó Member has been ascribed to the upper part of the early Pliensbachian (late Carixian).
3 Methods and materials
Three stratigraphic sections of the Es Barraca Member (the Son Maina, S’Heretat and Cuevas de Artà sections) located in the northeastern and southeastern sectors of Mallorca (Llevant Mountains, Fig. 4) were logged bed-by-bed and sampled. The logged deposits (Fig. 5) consist of limestones and dolostones deposited on tidal flat, subtidal inner platform (including lagoon, internal bars and shoals), muddy open lagoon and muddy outer platform environments (Sevillano et al. 2019). The sedimentological information of depositional facies and facies associations, along with the stratigraphic location of the studied samples were represented in stratigraphic logs (Fig. 5). Eight depositional facies (F1 to F8 in Fig. 5) have been distinguished in the studied sections based on variations in lithology and carbonate microfacies as well as principal biotic components. The studied micropaleontological material comes mainly from the subtidal inner platform facies.

Simplified geological map of Mallorca with the location of the three studied stratigraphic sections (SH S’Heretat, CA Cuevas de Artà, MAI Son Maina). The present-day distance between stratigraphic sections is shown without restoring the tectonic shortening

Detailed stratigraphic logs of the Es Barraca Member in the Son Maina, S’Heretat and Cuevas de Artà sections, with location of the studied samples, distribution of benthic foraminifera and algae, and interpretation of facies associations and facies types (F1 to F8, after Sevillano et al. 2019)
The micropaleontological content (foraminifera and other microfossils) was analysed in thin sections. A total of 56 samples in thin sections were examined (see location in Fig. 5). The stratigraphic distribution of the recognized taxa was represented in the stratigraphic logs (Fig. 5). The determination of foraminiferal biozones follows the standard biozonation proposed by Septfontaine (1984), which is widely accepted for the Mediterranean area. The comparison of this biostratigraphical scheme with other schemes based also on LBF and with the lower Jurassic standard ammonite biozones is given in Fig. 2.
4 Description of the sections and microfossil assemblages
4.1 Son Maina section
The Son Maina section is located southeast of the town of Manacor (coordinates: 39°30′49″ N, 3°15′4″ E; Fig. 4) in the mid-southern Llevant Mountains. It crops out in the inverted flank of a NW–SE fold (Fornós et al. 1984). This section is the most representative of those studied, showing the thickest and most complete stratigraphic record. The Es Barraca Member consists of about 219 m of tabular limestones overlaying a dolomite succession possibly belonging to the Mal Pas Formation (Hettangian). The top of the succession is located at a thin ferruginous crust overlaid by crinoidal limestone of the Es Cosconar Formation (Fig. 5). Depositional facies in this section include the facies F1–F8 (Fig. 5). In the basal 126.5 m, facies consist mainly of flat-pebble breccias, microbial laminites/stromatolites and intraclastic-peloidal-oolitic grainstone deposited in peritidal environments (facies F1, F2 and F3 respectively; Fig. 5), interbedded with abundant lime mudstone with a rare restricted fauna of ostracods and miliolids (facies F4) and fossiliferous wackestone-packstone (facies F6), deposited in shallow restricted and open lagoonal environments respectively. Some thin levels of oolitic-peloidal grainstone (facies F5) representative of internal bars are also present (Sevillano et al. 2019). The succession evolves upward, in the following 88 m (up to meter 214.5; Fig. 5), to a predominance of skeletal mudstone-wackestone (facies F6, F8), peloidal-oncolitic-foraminiferal wackestone-packstone (facies F6, F7) and oolitic-oncolitic grainstone (facies F5), interpreted as subtidal facies. They are interbedded with a minor amount of fenestral mudstone deposited in peritidal environments (facies F2), which are noticeably less developed in this upper part of the succession (Fig. 5). This interval ends with the development of a thin ferruginous hardground surface (HG-0 in Fig. 5). Above this hardground (HG-0), from 214.5 to 219 m, the facies consist mainly of silty, peloidal-oolitic packstone to grainstone containing larger foraminifera and large coarse oncoids, suggesting still deposition in a shallow protected carbonate shelf. This interval finishes, at 219 m, with the development of an irregular ferruginous hardground surface (HG-1 in Fig. 5) containing pockets of rounded, centimetric quartzite pebbles on top, suggesting the by-pass of coarse siliciclastics. Above this hardground surface at meter 219 m (HG-1), the succession continues with a partly covered interval of crinoidal grainstone of the Es Cosconar Formation, deposited in an open shelf. This interval contains some internal ferruginous hardgrounds with accumulations of ammonites and belemnites attributable to the upper Pliensbachian (Fig. 5).
The observed microfossil assemblage (Figs. 5, 6, 7) contains, throughout the whole section, abundant specimens of Thaumatoporella parvovesiculifera (Raineri) (Fig. 6a–c) and Palaeodasycladus mediterraneus (Pia) (Fig. 6d–f). In the first 106 m of the section, the foraminifera are represented by: Siphovalvulina (Fig. 6g–i), Siphovalvulina evolving to primitive Pseudopfenderina and in transition to Valvulina (?), Pseudopfenderina, Everticyclammina praevirguliana Fugagnoli (Fig. 6o, p), Meandrovoluta asiagoensis Fugagnoli & Rettori (Fig. 7o), Duotaxis (Fig. 6q), Biokovina (Fig. 7a), Mesoendothyra (Fig. 6j), evolved uncoiled Mesoendothyra with an important planispiral nepionic stage and some ancestors of Lituosepta (Fig. 6k–n). The later are uncoiled cylindrical Mesoendothyrids, very small forms, with multiple apertures (Fig. 6m, n) but no internal subdivisions (radial partitions). The first occurrence of Lituosepta recoarensis Cati with radial partitions (Fig. 7b) is at 106 m and it is recorded upward for the rest of the succession (Fig. 7c–e) along with Siphovalvulina-Valvulina (?), Everticyclammina and Mesoendothyra. Approximately 160 meters from the base of the succession abundant flat morphotypes of Planisepta (Fig. 7i) appear with less common Haurania deserta Henson (Fig. 7g, h red arrow). The first appearance of primitive forms of Orbitopsella occurs in the upper part of the succession (Fig. 7j–l), from 196 to 202 m, showing within this interval a clear transition of Lituosepta to Orbitopsella, with small size (2–3 mm) flabelliform to reniform tests but no true discoidal forms. Orbitopsella aff. primaeva (Henson) first occurs at 202 m (small diameter up to 4–5 mm) (Fig. 7m, n), along with cylindrical L. recoarensis, flat Planisepta, Siphovalvulina and uncoiled Mesoendothyra. The last interval of the succession, between hardgrounds HG-0 and HG-1 (214.5–219 m, Fig. 5), contains Orbitopsella primaeva (Henson) (diam. 6 mm), small primitive forms of O. primaeva (diam. 1.5–3 mm), Everticyclammina sp., Siphovalvulina (thick wall) and Valvulina (?). The described Orbitopsella corresponds to microspheric (B) forms, and megalospheric (A) forms have not been found in this section. From 219 m, above the hardground surface HG-1 (Fig. 5), the microfacies changes to sandy crinoidal packstone with lagenids, but lacking larger complex foraminifera, indicating an abrupt change to open marine conditions (Es Cosconar Formation).

Representative microfossils from the studied sections. aPalaeodasycladus mediterraneus (Pia). bPalaeodasycladus mediterraneus (Pia) (yellow arrow) and lituolids (red arrow). cPalaeodasycladus mediterraneus (Pia). d–fThaumatoporella parvovesiculifera (Raineri). g–iSiphovalvulina sp. jMesoendothyra sp. k–n illustrations of the transition between Mesoendothyra (adult stage rectilinear) and a primitive Lituosepta with multiple apertures (m–n). Specimens from the Son Maina section. Radial partitions are not yet developed, they appear higher in the section (see Fig. 7b). o–pEverticyclammina praevirguliana Fugagnoli. qDuotaxis sp.

Representative microfossils from the studied sections. aBiokovina sp. morphotype with pseudokeriotheca and filament bacterial symbionts (Vachard et al. 2004). b–fLituosepta recoarensis Cati (yellow arrow in f). Note the presence of radial partitions. gHaurania deserta Henson. hHaurania sp. (red arrow) and Lituosepta recoarensis Cati (yellow arrow). iPlanisepta morphotype (yellow arrow). j–l transitional morphotypes to Orbitopsella or primitive Orbitopsella. mOrbitopsella sp. nOrbitopsella aff. primaeva Henson. oMeandrovoluta asiagoensis Fugagnoli & Rettori
4.2 S’Heretat section
The S’Heretat section is located east of the town of Artà (coordinates: 39°40′47″ N, 3°25′55″ E; Fig. 4) and is 131.5 m thick. Its lower boundary with the Mal Pas Formation is not exposed, whereas the top boundary is represented by an unconformity surface with the overlying crinoidal limestone of the Es Cosconar Formation. The first basal 45 m of the succession is composed mainly of microbial laminites and laminated stromatolites (facies F2, Fig. 5) deposited in peritidal environments, alternating with less abundant lime mudstone with scarce fauna (facies F4) and oolitic-peloidal grainstone (facies F5) interpreted to be deposited in subtidal restricted lagoons and internal bars respectively (Sevillano et al. 2019; Fig. 5). Above 45 m and up to 106 m, the deposits are dominated by subtidal facies consisting of skeletal mudstone–wackestone and peloidal–oncolitic wackestone–packstone with abundant foraminifera (facies F6–F8), deposited in shallow open lagoons to open platform environments, interbedded with subordinated oolitic–peloidal–oncolitic grainstone (facies F5) deposited in internal bars and shoals (Sevillano et al. 2019; Fig. 5). In this interval, a few thin intercalations of peritidal microbial laminites (facies F2) also occur. Finally, from 106 m up to 131.5 m, the succession follows with oolitic–peloidal–oncolitic grainstone (facies F5) alternating with peloidal–oncolitic–foraminiferal wackestone (facies F6) deposited in subtidal inner platform environments (Fig. 5). In this interval, only one thin intercalation of peritidal laminites with flat pebbles (facies F1) is present at approximately 114 m. Above this interval, the succession is characterised by some levels of echinoderm grainstone attributed to the Pliensbachian Es Cosconar Formation (Fig. 5).
The microbial-dominated lower part of the succession (up to 45 m) contains very rare microfossils. In samples from some basal grainy layers P. mediterraneus has been identified. From meter 46 upward, the microfossils are notably more diversified and include: Siphovalvulina, transitional morphotypes of Siphovalvulina to Pseudopfenderina, Mesoendothyra, Everticyclammina sp., L. recoarensis (Fig. 7f), P. mediterraneus and T. parvovesiculifera. From 87 m upward flat morphotypes of Planisepta appear, along with evolved flabelliform forms of L. recoarensis, varied forms of Siphovalvulina, and evolved Mesoendothyra. This assemblage is observed up to 131.5 m. An absence of Orbitopsella is notable. From 131.5 m upwards, the microfacies changes sharply to crinoidal wackestone-packstone with bryozoans and lagenids but lacking complex larger foraminifera, reflecting an abrupt change to completely different open shelf conditions (Es Cosconar Formation).
4.3 Cuevas de Artà section
The Cuevas de Artà section is located close to the Cap Vermell (coordinates: 39°39′55″ N, 3°27′8″ E; Fig. 4). At this locality, the measured thickness of the Es Barraca Member is 110 m. The lower boundary is the contact with a basal dolomite body attributed to the Hettangian Mal Pas Formation. The upper boundary is an unconformity represented by a ferruginous hardground, which is overlain by red nodular limestones with “filaments” of pelagic–benthic thin-shelled bivalves attributed to Bositra, and ammonites of Bajocian age (Álvaro et al. 1989). Therefore, at this locality, a hiatus spanning at least from the Pliensbachian to the Bajocian occurs. The whole succession of the Es Barraca Member is made mainly of peritidal deposits, consisting of an alternation of black-pebble conglomerates (facies F1 in Fig. 5), microbial laminites-stromatolites (facies F2), intraclastic-peloidal-oolitic grainstone with pedogenic features (facies F3) deposited in intertidal-supratidal environments, and lime mudstone (facies F4) deposited in restricted lagoons (Fig. 5). A few thin oolitic grainstone intercalations (facies F5), interpreted as internal bars, are also present.
The Cuevas de Artà section contains a low-diversity microfossil content with no LBF. It includes abundant P. mediterraneus and T. parvovesiculifera, along with rare specimens of Siphovalvulina and Mesoendothyra, with some transitional forms to primitive Lituosepta in the uppermost part of the succession. The rarity of foraminifera in this section may be due in part to the restriction imposed by the depositional environment, which is dominated by peritidal conditions with frequent emersion episodes and by the stacking of metre-thick parasequences dominated by intertidal and supratidal facies (Sevillano et al. 2019). Such restricted depositional environments might have been unfavourable for the proliferation of foraminifera (Gale and Kelemen 2017).
5 Discussion
5.1 Biostratigraphy and age constraints of the Es Barraca member
The analysed thin section samples from the three studied sections show microfacies indicative of shallow subtidal, lagoonal, inner platform environments (following the microfacies analysis of Sevillano et al. 2019), except in two samples belonging to the Es Cosconar Formation at the top of the Son Maina and S´Heretat sections (Fig. 5). These two samples represent a completely different environment rich in echinoderm remains and lagenids, indicative of open marine conditions. For the rest of the studied samples (Es Barraca Member), the allochems are made mainly of ooids, micro-oncoids, bioclasts, muddy lithoclasts (lithic peloids), benthic foraminifera, calcareous algae, calcimicrobes and large oncoids.
The dasycladalean alga Palaeodasycladus occurs throughout all the studied sections associated with the microproblematica Thaumatoporella, both characteristic of the Lower Jurassic protected marine settings of the western Tethys (e.g. BouDagher-Fadel and Bosence 2007). The widespread occurrence of these taxa in the Early Jurassic of the western Tethys was favoured by the wide continuity of shallow-carbonate platform environments along the entire region. Palaeodasycladus was confined to the lower and middle part of the early Jurassic (Hettangian to Pliensbachian; Sokač 2001), whereas Thaumatoporella had a much longer stratigraphic range, from the Middle Triassic to the Paleocene (Schlagintweit and Velić 2011).
The analysed samples of the Es Barraca Member show that larger foraminifera were taxonomically diverse in the Balearic region, showing affinities with those reported in Slovenia, Croatia, Italy, southern Iberia and Morocco (Septfontaine 1984; Fugagnoli and Loriga Broglio 1998; Azerêdo et al. 2003; Veseli et al. 2006; BouDagher-Fadel and Bosence 2007; Velić 2007; Gale 2014; Gale and Kelemen 2017). The biostratigraphy and the chronostratigraphic interpretation of the studied successions are shown in Fig. 8. According to the observed foraminiferal assemblages and stratigraphic distribution (Fig. 5), three biozones A, B and C1 have been identified (Fig. 8). These biozones could be correlated with those proposed by Septfontaine (1984) for the lower Jurassic of the High Atlas of Morocco (Figs. 2, 8). The Es Barraca Member corresponds mainly to the biozones A, B and the base of C1. The biozone C1 was observed only in the uppermost part of the unit in the Son Maina section (Fig. 8).

Distribution of benthic foraminiferal biozones of the Es Barraca Member in the Son Maina, S’Heretat and Cuevas de Artà sections (Llevant Mountains domain, Mallorca), based on the biozonation scheme proposed by Septfontaine (1984). The steps 1 to 3 (phylozones) represent evolutionary stages of the larger lituolids based on morphological transitions from Mesoendothyra to orbitopsellinids s.l. The pink striped area in the Son Maina section indicates the interval of uncertainty in the limit between biozones A and B due to the absence of samples
The biozone A (Siphovalvulina sp. and Mesoendothyra sp. Interval Zone), identified in the three studied sections, is characterized by the presence of Siphovalvulina and Mesoendothyra along with advanced individuals interpreted as belonging to an ancestor morphotype of Lituosepta. The upper boundary of this biozone is marked by the first occurrence of L. recoarensis. In the Son Maina section, the most complete of the three sections, biozone A spans from the base of the section up to 106 m. In the S’Heretat section, biozone A spans from the base up to 46 m. In the Cuevas de Artà section, the entire succession of the Es Barraca Member up to the hardground overlaying the shallow-water limestones corresponds to biozone A.
The biozone B (Lituosepta recoarensis Lineage Zone, Septfontaine 1984) is bounded by the first occurrence of L. recoarensis up to the first occurrence of O. primaeva. In the Son Maina section, the biozone B starts at 106 m and spans up to 202 m, where the first forms of Orbitopsella aff. primaeva appear. In this biozone L. recoarensis co-occurs with Siphovalvulina and Mesoendothyra persisting from the underlying biozone. In the upper part of the biozone flat morphotypes of Planisepta and H. deserta also occur. In the uppermost part of the biozone B, the interval 196 m to 202 m are characterized by the appearance of small primitive forms of Orbitopsella (transitional morphotypes), suggesting a transitional change between biozones B and C1 (B/C1 transition, Fig. 8) as a result of the transitional character of one lineage zone to the next (as defined by Septfontaine 1984). The base of this transitional interval is marked by the first occurrence of ancestral Orbitopsella and the top by the first occurrence of Orbitopsella aff. primaeva (Figs. 5, 7, 8). The biozone C1 (Orbitopsella primaeva Lineage Zone, Septfontaine 1984), characterized in its lower part by the first appearance of Orbitopsella aff. primaeva (Henson), is present at the uppermost part of the Son Maina section, from 202 m upwards, and extends above the hardground surface HG-0 up to HG-1 (Fig. 8).
In the S’Heretat section, biozone B starts at 46 m and spans up to the hardground at top of the succession of the Es Barraca Member, since no Orbitopsella specimens have been found. In the Cuevas de Artà section, biozone B is absent (Fig. 8).
Fugagnoli (2000) established the stratigraphic position of Everticyclammina praevirguliana in northern Italy as being from the late Sinemurian to the Pliensbachian. Later, BouDagher-Fadel and Bosence (2007) set the first appearance of E. praevirguliana in the middle Sinemurian for the western Mediterranean region, whereas according to Velić (2007) it first appeared in the late Hettangian. Fugagnoli and Bassi (2015), in their revision of L. recoarensis, recognised a variable stratigraphic range for this taxon in different Tethyan areas, between the Sinemurian and the Pliensbachian. There is, however, a good consensus that the first appearance of L. recoarensis is in the early late Sinemurian (Septfontaine 1984; BouDagher-Fadel and Bosence 2007; Velić 2007; Gale 2014; Fugagnoli and Bassi 2015), this species having a relatively well-defined lowest biostratigraphic position in this part of the stage. The first occurrence of Orbitopsella is established in the latest Sinemurian, with O. primaeva spanning from the latest Sinemurian to the early Pliensbachian (Septfontaine 1984; Fugagnoli and Loriga Broglio 1998; Velić 2007; BouDagher-Fadel 2008). According to these data, it can be concluded that the Es Barraca Member (biozones A, B and base of C1) of Mallorca has mainly a Sinemurian age. At Son Maina, the most complete section, the upper boundary of the Es Barraca Member is represented by the hardground surface HG-0. In this section, the uppermost part of the Es Barraca Member (the last 12.5 m that correspond to biozone C1 below the hardground surface HG-0) contains only small primitive forms of O. primaeva. This suggests an age not younger than the latest Sinemurian for the top of the Es Barraca Member. This contrasts with earlier studies (Álvaro et al. 1989) that suggested the Es Barraca Member could be as young as Pliensbachian (Fig. 9). The interval of shallow-water limestones containing Orbitopsella aff. primaeva between the hardgrounds HG-0 and HG-1 (214.5–219 m, above the top of the Es Barraca Member; Figs. 5 and 8) could range in age from the latest Sinemurian to the early Pliensbachian (early Carixian), biozone C1. However, the Orbitopsella aff. primaeva specimens appearing in this interval are still very small (diam. 1.5–3 mm) and primitive B-forms compared to the typical O. primaeva (4–6 mm for microspheric forms; Hottinger 1967; BouDagher-Fadel 2008) and are also rare, which could point to a likely latest Sinemurian age for this interval. However, the absence of more complex and evolved larger orbitopsellinids in these beds could be in part biased by sorting effects in the carbonate sands (which could lead to the absence of larger particles), implying, therefore, that an early Pliensbachian age cannot be excluded for this interval. More additional information of this interval from other sections is needed.

Stratigraphical scheme established by Álvaro et al. (1989) for the Lower Jurassic succession of Mallorca (left) and comparison with the new proposal for the Sinemurian–lower Pliensbachian in this work (right)
5.2 Evolutionary considerations
The Jurassic lituolids underwent significant evolutionary changes during the Sinemurian. The most important is the evolution of most of the lower Jurassic lituolids from Mesoendothyra sp. (Septfontaine 1984, 1988). One of the proposed evolutionary lines evolving from this form in the late Sinemurian gave rise to the orbitopsellinids, according to the evolutionary bioseries: Mesoendothyra sp.—Lituosepta recoarensis—Orbitopsella spp. (Septfontaine 1984, 1988). Their phyletic relationships in terms of morphological characteristics and internal microstructures were already introduced by Hottinger (1967) and later illustrated and redefined in Septfontaine (1984, 1988). This evolutionary bioseries has been accepted by other authors (e.g. Bassoullet 1997; Azerêdo et al. 2003; BouDagher-Fadel 2008) and seems to be confirmed in Mallorca. In the studied samples, this gradual evolution of the orbitopsellinids s.l. during the Sinemurian could be observed in three steps (step 1–3 in Fig. 8), which represent relatively stable evolutionary morphogenetic (architectural) innovations with many transitions, as observed in the Son Maina section. The biozonation proposed in this study is based on these steps (phylozones), and their equivalence with the biozones is shown in Fig. 8.
Taking the Son Maina section as the reference section of this study (Fig. 8), the evolutionary steps are observed as follows: Step 1, 24–106 m, characterized by an ancestor to Lituosepta (uncoiled cylindrical Mesoendothyra) with no internal subdivisions, multiple apertures, large planispiral nepionic stage, and very small size, less than 500 μm in diameter or test length (only B-forms found). Radial partitions are not yet developed. Step 2, 106–161 m, characterized by the first occurrence of vertical radial partitions, true cylindrical L. recoarensis with rare flat “Planisepta-like” morphotypes. Step 3, 161–214.5 m, characterized by the diversification of the shape and external contour of the test, advanced Lituosepta to Orbitopsella with reniform to discoidal test, vertical radial partitions in the marginal zone of the chambers and pillars in the central zone (sensu Septfontaine 1980). The flat “Planisepta-like” morphotypes become frequent. Many transitions between the different morphotypes were observed, but with a general trend to increase the chamber volume, a known evolutive feature in foraminifera developed for hosting more symbionts (Hottinger 2000).
The characteristics of the steps 2 and 3 can also be observed in the S’Heretat section (Fig. 8). To further understand the mode of evolution (punctuated versus gradual), more data from bed-by bed sampling are needed.
5.3 Stratigraphic implications
The biostratigraphic results presented in this study have some significant implications for the stratigraphy and paleogeographic reconstruction of the Lower Jurassic of Mallorca. The current stratigraphic scheme by Álvaro et al. (1989) attributes the Es Barraca Member to a Sinemurian to early Pliensbachian age, showing a lateral transition of the upper part of the shallow-water limestones of the Es Barraca Member in the Llevant Mountains paleogeographic domain, to the marls and marly limestones with brachiopods of the Sa Moleta Member (Fig. 9; Álvaro et al. 1989). The latter developed only in the paleogeographic domain of the Tramuntana range (Fig. 4). Because the Sa Moleta Member has been dated with ammonites and brachiopods as lower Pliensbachian (Carixian), within the Jamesoni and Ibex standard ammonites zones (Álvaro et al. 1989), these authors inferred an early Pliensbachian age also for the upper part of the Es Barraca Member. However, the regional lateral correlation between the upper part of the Es Barraca Member and the Sa Moleta Member is not well established, since the lateral relationships between these two units have not yet been directly observed in the field. In the Son Maina section, the most complete section of the Llevant Mountain domain, the top of the Es Barraca Member is represented by the hardground surface HG-0 (Fig. 8). The biostratigraphic data presented in this study do not support the early Pliensbachian age for the upper Es Barraca Member, because below the hardground HG-0 only the very base of the biozone C1 is present, which is dated upper late Sinemurian (Figs. 8, 9). This implies the lack of a lateral correlation between these two stratigraphic units. However, just above of the Es Barraca Member in the Son Maina section, the thin interval of shallow-water limestones between hardground surfaces HG-0 and HG-1 with specimens of Orbitopsella aff. primaeva (biozone C1), could be dated as latest Sinemurian–earliest Pliensbachian. These beds overlie the Es Barraca Member, being separated from that unit by a hiatus of probably short duration (hardground HG-0). These shallow-water limestones are separated from the overlaying open sea calcarenites of the upper Pliensbachian Es Cosconar Formation by another hiatus (hardground HG-1) of probably longer duration (Fig. 8). Therefore, this thin interval of biozone C1 shallow-water limestones between the two unconformities (HG-0 and HG-1) could not be attributed to any existing lithostratigraphic unit, and should be regarded as a new stratigraphic unit. The age of this new unit could range from the latest Sinemurian to the early Pliensbachian, but its lateral correlation with the Sa Moleta and Es Racó Members of the Tramuntana range remains unclear (Fig. 9). On Cabrera Island Arbona et al. (1984) documented, above the Es Barraca Member, the existence of a ~ 10 m thick succession of shallow-water carbonates dated with ammonites as early Pliensbachian (Carixian). It is probable that it corresponds to the same unit. More stratigraphic and biostratigraphic information from this new unit are needed.
Another significant stratigraphic implication of this study is the existence of an intra-early Pliensbachian phase of platform fragmentation and erosion or, alternatively, of local non-deposition during the latest Sinemurian–earliest Pliensbachian. The reconstruction of the depositional transect across the three studied profiles shows the northward progressive loss of the upper biozones of the Sinemurian (Fig. 8). This would suggest the existence of sedimentary hiatuses in some sections of the Llevant Mountains domain that become more significant toward the northeast. Thus, in the Son Maina section located to the south, the complete record of the Es Barraca Member is preserved, and at least part of a new carbonate unit dated as biozone C1, which overlies the Es Barraca Member. This new carbonate unit, latest Sinemurian to the earliest Pliensbachian in age, is topped by an unconformity with a hiatus spanning most of the early Pliensbachian up to the upper Pliensbachian (Domerian) Es Cosconar Formation. Toward the north, in the S´Heretat section, there is a stratigraphic gap on top of the Es Barraca Member, spanning from the upper part of the late Sinemurian up to the early Pliensbachian (Carixian), before the onset of sedimentation of the upper Pliensbachian (Domerian) calcarenites of the Es Cosconar Formation (Figs. 8, 9). Finally, to the northeast, in the Cuevas de Artà section (Fig. 8), only the biozone A of the early-middle Sinemurian was preserved or registered below the unconformity at the top of the Es Barraca Member. In the Cuevas de Artà section, the unconformity, evidenced by a ferruginous hardground, represents a hiatus spanning from the late Sinemurian biozone B up to the Bajocian (Álvaro et al. 1989). This period of platform fragmentation and erosion was likely coeval with the formation of intraplatform basins during which lower Pliensbachian marls, marly limestones and deltaic siliciclastics (Sa Moleta and Es Racó Members) deposited in the north-western part of Mallorca island (Tramuntana range) (Álvaro et al. 1989). Later, the upper Pliensbachian sedimentation was discontinuous and characterized by open sea carbonate facies (Es Cosconar Formation) with notable changes of thickness. While some sedimentary successions were expanded and show continuous sedimentation (Rosales et al. 2018), others were more condensed and containing several internal disconformities and hardgrounds.
6 Conclusions
The Sinemurian shallow-water carbonate succession (Es Barraca Member) cropping out in the Llevant Mountains of Mallorca (Balearic Islands) has been investigated to create a biostratigraphic framework based on LBF assemblages. Benthic foraminifera and algae identified from randomly oriented thin sections have allowed the identification of a gradual evolution of the orbitopsellinids s.l. in three steps (phylozones): Step 1, characterized by small forms of a Lituosepta ancestor; Step 2, typified by the occurrence of true cylindrical L. recoarensis and rare “Planisepta-like” morphotypes; and Step 3, characterized by advanced Lituosepta to Orbitopsella and frequent “Planisepta-like” morphotypes. Based on these evolutionary steps the biostratigraphic setting of the Es Barraca Member is proposed. The Es Barraca Member is ascribed to the biozones A, B and base of C1, although some of the zones may be missing in different sections due to the unconformity surface on top of the Es Barraca Member. This study restricts the age of the Es Barraca Member to the Sinemurian and gives an age not younger than the late Sinemurian for its top.
A new shallow-water carbonate stratigraphic unit belonging to the biozone C1 (likely latest Sinemurian–earliest Pliensbachian) and deposited above the Es Barraca Member was identified. This unit is observed only in the most complete section (Son Maina) because in the other sections it was eroded during Pliensbachian and/or later unconformities. The biostratigraphic correlation across the studied stratigraphic transect in the Llevant Mountains shows the progressive loss of the upper biozones towards the northeast. This loss suggests the existence of significant hiatuses and allows to infer an early Pliensbachian phase of platform fragmentation, which resulted in some areas with non-deposition or erosion during the early Pliensbachian, and in other areas with deposition of marine marly limestone and deltaic siliciclastics (Sa Moleta and Es Racó Members, respectively), as observed in the north-western part of Mallorca (Tramuntana range).
References
Álvaro, M., Barnolas, A., Cabra, P., Comas-Rengifo, M. J., Fernández-López, S. R., Goy, A., et al. (1989). El Jurásico de Mallorca (Islas Baleares). Cuadernos de Geología Ibérica,13, 67–120.
Arbona, J., Fontbote, M., González-Donoso, J. M., Linares, A., Olóriz, F., Pomar, L., et al. (1984). Precisiones bioestratigráficas y aspectos sedimentológicos del Jurásico-Cretácico basal de la isla de Cabrera (Baleares). Cuadernos de Geología,12, 169–186.
Azerêdo, A. C., Manuppella, G., & Ramalho, M. M. (2003). The late Sinemurian carbonate platform and microfossils with Tethyan affinities of the Algarve Basin (South Portugal). Facies,48, 49–60.
Barnolas, A., & Simó, J. A. (1984). Sedimentología del Jurásico de Mallorca. In A. Barnolas (Ed.), Grupo Español del Mesozoico (pp. 73–120). Palma de Mallorca: IGMECGS S.A.
Barnolas, A., Simò, J. A., Rosales, I., Gil-Peña, I., Sevillano, A., & Armendáriz, M. (2010). Revisión de la información paleogeográfica del Jurásico de Mallorca basada en datos sedimentológicos y en el análisis de cuenca. In J. I. Ruiz-Omeñaca, L. Piñuea, & J. C. García-Ramos (Eds.), Comunicaciones del V Congreso del Jurásico de España (p. 163). Colunga: Museo del Jurásico de Asturias.
Bassoullet, J. P. (1997). Les grands foraminifères. In: Cariou, E. and Hantzpergue, P. (coords.). Biostratigraphic du Jurassique Ouest-Européen et Méditerranéen. Groupe Francais d´étude du Jurassique. Bulletin du Centre Recherches Elf Exploration Production Mém,17, 293–304.
BouDagher-Fadel, M. K. (2008). Evolution and geological significance of larger benthic Foraminifera. Developments in paleontology and stratigraphy (Vol. 21). Amsterdam: Elsevier.
BouDagher-Fadel, M. K., & Bosence, D. W. J. (2007). Early Jurassic benthic foraminiferal diversification and biozones in shallow-marine carbonates of western Tethys. Senckenbergiana Lethaea,87, 1–39.
Colom, G. (1942). Sobre nuevos hallazgos de yacimientos fosilíferos del Lias medio y superior en la Sierra Norte de Mallorca. Boletín de la Real Sociedad Española de Historia Natural,11, 221–265.
Colom, G. (1966). Dos niveles micropaleontológicos interesantes en el Lias inferior del Sur de España y Baleares. Acta Geologica Hispanica,1(3), 15–18.
Colom, G. (1970). Estudio litológico y micropaleontológico del Lías de la Sierra Norte y porción central de la isla de Mallorca. Memorias de la Real Academia de la Ciencias exactas, físicas y naturales de Madrid, Tomo XXIV, Mem 2.
Colom, G. (1980). Estudios sobre las litofacies y micropaleontología del Lías inferior de la Isla de Cabrera (Baleares). Revista Española de Micropaleontología,12, 47–64.
Colom, G., & Dufaure, P. (1962). Présence de la zone à Palaeodasycladus mediterraneus (Pia) dans le Lias moyen du Pla de Cuber (Majorque). Comptes Rendus de l’Académie des Sciences de Paris,12, 2617–2619.
Dercourt, J., Gaetani, M., Vrielynck, B., Barrier, E., Biju-Duval, B., Brunet, M. F., et al. (Eds.). (2000). Atlas Peri-Tethys, Palaeogeographical maps, I–XX. Paris: CCGM/CGMW.
Dewey, J. F., Pitman, W. C., Ryan, W. B. F., & Bonnin, J. (1973). Plate tectonics and the evolution of the Alpine system. Geological Society of America Bulletin,84, 3137–3180.
Etheve, N., Mohn, G., Frizon de Lamotte, D., Roca, E., Tugend, J., & Gómez-Romeu, J. (2018). Extreme mesozoic crustal thinning in the Eastern Iberia Margin: the example of the Columbrets Basin (Valencia Trough). Tectonics,37(2), 1–27.
Fallot, P. (1922). Étude geologique de la sierra de Majorque. Thése détat. Libr. Polytechnique Ch. Béranger, Paris i Liège.
Fornós, J., Rodriguez-Perea, A., & Sabat, F. (1984). El mesozoico de la Serra de Son Amoixa (Serres de Llevant, Mallorca). I Congreso Español de Geología. Tomo,1, 173–185.
Franceschi, M., Dal Corso, J., Cobianchi, M., Roghi, G., Penasa, L., Picotti, V., et al. (2019). Tethyan carbonate platform transformation during the Early Jurassic (Sinemurian–Pliensbachian, Southern Alps): comparison with the Late Triassic Carnian Pluvial Episode. Geological Society of America Bulletin,131, 1255–1275.
Fugagnoli, A. (2000). First record of Everticyclammina Redmond 1964 (E. Praevirguliana n. sp.; Foraminifera) from the early Jurassic of the Venetian Prealps (Calcari Grigi, Trento platform, northern Italy). Journal of Foraminiferal Research,30, 126–134.
Fugagnoli, A. (2004). Trophic regimes of benthic foraminiferal assemblages in Lower Jurassic shallow water carbonates from northeastern Italy (Calcari Grigi, Trento Platform, Venetian Prealps). Palaeogeography, Palaeoclimatology, Palaeoecology,205, 111–130.
Fugagnoli, A., & Bassi, D. (2015). Taxonomic and biostratigraphic reassessment of Lituosepta recoarensis Cati, 1959 (Foraminifera, Lituolacea). Journal of Foraminiferal Research,45, 402–412.
Fugagnoli, A., & Loriga Broglio, C. (1998). Revised biostratigraphy of Lower Jurassic shallow water carbonates from the Venetian Prealps (Calcari Grigi, Trento Platform, Nothern Italy). Studi Trentini di Science Naturali Acta Geologica,73(1996), 35–73.
Gale, L. (2014). Lower Jurassic foraminiferal biostratigraphy of Podpeč Limestone (External Dinarides, Slovenia). Geologija,57, 119–146.
Gale, L., & Kelemen, M. (2017). Early Jurassic foraminiferal assemblages in platform carbonates of Mt. Krim central Slovenia. Geologija,60(1), 99–115.
Guex, J. (2016). Retrograde evolution during major extinction crises. Springer, Berlin: SpringerBriefs in Evolutionary Biology.
Hottinger, L. (1967). Foraminifères imperforés du Mesozoïque marocain. Notes et Mémoires Service Géologique,209, 1–168.
Hottinger, L. (2000). Functional morphology of benthic foraminiferal shells, envelopes of cells beyond measure. Micropaleontology,46, 57–86.
Kabal, Y., & Tasli, K. (2003). Biostratigraphy of the Lower Jurassic carbonates from the Aydinick area (Central Taurides, S. Turkey) and morphological analysis of Lituolipora termieri (Hottinger, 1967). Journal of Foraminiferal Research,33, 338–351.
Mancinelli, A., Chiocchini, M., Chiocchini, R. A., & Romano, A. (2005). Biostratigraphy of Upper Triassic–Lower Jurassic carbonate platform sediments of the central-southern Apennines (Italy). Rivista Italiana di Paleontologia e Stratigrafia,111, 271–283.
Masetti, D., Figus, B., Jenkyns, H. C., Barattolo, F., Mattioli, E., & Posenato, R. (2016). Carbon-isotope anomalies and demise of carbonate platforms in the Sinemurian (Early Jurassic) of the Tethyan region: evidence from the Southern Alps (Northern Italy). Geological Magazine,154, 625–650.
Ogg, J. G., Ogg, G., & Gradstein, F. M. (2016). A concise geologic time scale (1st ed.). Amsterdam: Elsevier.
Preto, N., Breda, A., Dal Corso, J., Franceschi, M., Rocca, F., Spada, C., et al. (2017). The Loppio Oolitic Limestone (Early Jurassic, Southern Alps): a prograding oolitic body with high original porosity originated by a carbonate platform crisis and recovery. Marine and Petroleum Geology,79, 394–411.
Rosales, I., Barnolas, A., Goy, A., Sevillano, A., Armendáriz, M., & López-García, J. M. (2018). Isotope records (C–O–Sr) of late Pliensbachian-early Toarcian environmental perturbations in the westernmost Tethys (Majorca Island, Spain). Palaeogeography Palaeoclimatololy Palaeoecology,497, 168–185.
Ruiz-Ortiz, P. A., Bosence, D. W., Rey, J., Nieto, L. M., Castro, J. M., & Molina, J. M. (2004). Tectonic control of facies architecture, sequence stratigraphy and drowning of a Liassic carbonate platform (Betic Cordillera, Southern Spain). Basin Research,16, 235–257.
Rychliński, T., Uchman, A., & Gaździcki, A. (2018). Lower Jurassic Bahamian-type facies in the Choč Nappe (Tatra Mts, West Carpathians, Poland) influenced by palaeocirculation in the Western Tethys. Facies,64, 15.
Schlagintweit, F., & Velić, I. (2011). Foraminiferan test and dasycladalean thalli as cryptic microhabitats for thaumatoporellacean algae from Mesozoic (Late Triassic–Late Cretaceus) platform carbonates. Facies,58, 79–94.
Scotese, C. R., & Schettino, A. (2017). Late Permian–Early Jurassic paleogeography of Western Tethys and the World. Permo-Triassic Salt Provinces of Europe, North Africa and the Atlantic Margins. Amsterdam: Elsevier.
Septfontaine, M. (1980). Les Foraminifères imperforés des milieux de plateforme au Mésozoïque: détermination pratique, interprétation phylogénétique et utilisation biostratigraphique. Revue de Micropaléontologie,23, 169–203.
Septfontaine, M. (1984). Biozonation (à l’aide des Foraminifères imperforés) de la plate-forme interne carbonatée liasique du Haut Atlas (Maroc). Revue de Micropaléontologie,27, 209–229.
Septfontaine, M. (1988). Vers une classification évolutive des lituolidés (Foraminifères) jurassiques en milieu de plate-forme carbonatée. Revue Paléobiologie,2(86), 229–256.
Sevillano, A., Rosales, I., Bádenas, B., Barnolas, A., & López-García, J. M. (2019). Spatial and temporal facies evolution of a Lower Jurassic carbonate platform, NW Tethyan margin (Mallorca, Spain). Facies,65, 3. https://doi.org/10.1007/s10347-018-0545-0.
Sokač, B. (2001). Lower and middle Liassic calcareous algae (Dasycladales) from Mt. Velebit (Croatia) and Mt. Trnovski Gozd (Slovenia) with particular reference to the genus Palaeodasycladus (Pia, 1920) 1927 and its species. Geologia Croatica,54(2), 133–257.
Thierry, J. (2000). Late Sinemurian (193–191 Ma). In J. Dercourt, M. Gaetani, B. Vrielynck, E. Barrier, B. Biji-Dubal, M. F. Brunet, J. P. Cadet, S. Crasquin, & M. Sandulescu (Eds.), Atlas Peri-Tethys. Palaeogeographical Maps explanatory notes (pp. 49–59). Paris: Commission for the Geologic Map of the World.
Vachard, D., Munnecke, A., & Servais, T. (2004). New SEM observations of keriothecal walls: implications for the evolution of Fusulinida. Journal of Foraminiferal Research,34, 232–242.
Velić, I. (2007). Stratigraphy and palaeobiogeography of Mesozoic benthic foraminifera of the Karst Dinarides (SE Europe). Geologia Croatica,60, 1–113.
Veseli, V., Velić, I., Vlahovic, I., Tisljar, J., & Stankovic, D. (2006). Biozonation of Sinemurian and Pliensbachian larger benthic foraminifera (Velebit Mt., Croatia). FORAMS 2006. Anuário do Instituto de Geociências UFRJ,29(1), 368–369.
Zempolich, W.G. (1993). The drowning succession in Jurassic carbonates of the Venetian Alps, Italy, a record of supercontinent breakup, gradual eustatic rise, and eutrophication of shallow water environments. In R.G. Loucks, J.F. Sarg (Eds.), Carbonate Sequence Stratigraphy: Recent Developments and Applications (pp. 63– 105). AAPG Memoir, 57.
Acknowledgements
This paper is a contribution to the research projects RTI2018-093613-B-100 and CGL2017-85038-P funded by the Spanish Ministry of Science, Innovation and Universities. The authors are grateful to two anonymous reviewers for their critical reading and useful comments that improved the manuscript. We also include a special acknowledgment to Mike Simmons (Halliburton) for his help with the English revision of the final version of the manuscript and to Associate Editor Juan Carlos Braga for handing the manuscript.
Author information
Authors and Affiliations
Corresponding author
Glossary
- Advanced or ancestor form (morphotype)
-
This term, usual in paleontological and evolutionary studies, permits a rough estimation of the degree of morphological evolution in a bioseries (evolutionary trend) through time, such as the schematic morphological steps proposed by Septfontaine (1988), Fig. 1, which seems to fit for the orbitopsellinids and many other discoidal larger foraminifera taxa of Mesozoic carbonate shelves around the Tethyan domain. Logically (Depéret’s Law) ancestral forms are smaller (less than 0.5 mm) and have a simpler morphology, whereas the advanced forms display a larger test with complex inner microstructures and modifications of the shape and size of the adult chambers. The term “inner microstructures” means endoskeletal (pillars) and exoskeletal (vertical radial partitions) elements. This can be a response to better and larger symbiotic hosting (“greenhouse structures”) and changes (see Hottinger 2000).
- Morphotype
-
This practical and informal term designates the particular morphology of one individual which could belong to a new taxon not yet introduced. In a bioseries it can be a missing link connecting morphologically two chronospecies already defined (see Septfontaine 1988, Fig. 1, and Guex 2016 for a current use of the term).
Rights and permissions
About this article

Cite this article
Sevillano, A., Septfontaine, M., Rosales, I. et al. Lower Jurassic benthic foraminiferal assemblages from shallow-marine platform carbonates of Mallorca (Spain): stratigraphic implications. J Iber Geol 46, 77–94 (2020). https://doi.org/10.1007/s41513-019-00117-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41513-019-00117-9
Keywords
- Sinemurian-earliest Pliensbachian
- Benthic foraminifera
- Biozones
- Carbonate platform
- Es Barraca Member
- Balearic Basin