
Abstract
Citrus have been grown as a commercial fruit crop in Morocco for a decade. The area dedicated to growing and producing citrus has increased from 2006 to 2018 throughout the country. The rapid decline of citrus trees has been observed in many locations. Therefore, this study aimed to identify and characterize fungal pathogens associated with dry root rot (DRR) disease of citrus from diseased trees and soil collected from different citrus-growing plantations. Isolation and morphological identification of pathogens were performed using PDA-SC medium. Thirty-four Fusarium isolates were recovered from symptomatic plants. These isolates were identified based on morphological traits and the sequence of the internal transcribed spacer (ITS) region. Four Fusarium species were identified. Fusarium solani (Martius) was most frequently identified (70.60%), followed by F. oxysporum (17.65%), F. equiseti (8.82%) and F. brachygibbosum (2.94%). The two latter species have recently been associated with DRR in Morocco. Two separate pathogenicity trials were conducted to test the effect of the pathogens on young citrus seedlings. Results indicated that, with the exception of F. equiesti, Fusarium isolates were aggressive pathogens of citrange-Troyer and sour orange seedlings 2 months postinoculation. Citrus trees displayed key symptoms of DRR such as root rot, discolored and necrotic roots, purple wood discoloration, plant yellowing and general weakness. Further, all four species were able to induce lesions when they were used to inoculate citrus branches. This is the first detailed report of Fusarium species associated with DRR in Morocco.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Citrus are among the most highly cultivated fruit crops in Morocco and worldwide. Morocco is known as one of the important producers of citrus and has increased the area dedicated to their production to 126,600 ha, which has been used to produce 2.6 million tons of citrus fruits annually (Jaouad et al. 2020). Despite its wide distribution and economic importance, the crop is frequently threatened by pests and diseases. However, knowledge of citrus fungal diseases, particularly those caused by Fusarium species, is limited. The identification of pathogens imperative (Guarnaccia and Crous 2017). Fusarium spp. are known to affect a large number of crops and cause various serious diseases like fruit rot, root rot, seedling damping-off and plant wilt (Parikh et al. 2018b; Rivera-Jiménez et al. 2018; Ezrari et al. 2019).
Fusarium species are a major threat to citrus production worldwide and cause serious diseases including dry root rot (DRR), twig rot, decline dieback, twig blight, vascular wilt, feeder root rot, root and stem rot (Spina et al. 2008; Al-Sadi et al. 2014; Liu et al. 2019). To the best of our knowledge, a few studies have investigated the importance of these kinds of diseases in Mediterranean areas, including Morocco. In Tunisia, a preliminary study revealed the presence of F. oxysporum f. sp. citri in some orchards within the Cap-Bon region (Hannachi et al. 2014). As a result of this study, and observed declines in citrus production due to Fusarium spp., DRR is becoming an increasingly important global problem. In fact, during the last decade, citrus production has undergone severe decline, which was probably linked to climatic change.
The presence of DDR in Morocco was first reported by Vanderweyen and Serrhini (1981) and Fusarium spp. was identified its causative agent. Its incidence was enhanced when plants were subjected to biotic (interaction with Phytophthora species and the nematode Tylenchulus semipenetrans) or abiotic (drought, root asphyxiation due to over watering, and excessive fertilization) stresses (Adesemoye et al. 2011; Yaseen and D’Onghia 2012). Furthermore, DRR was reported to be caused by a complex of different Fusarium species, and F. solani (Neocosmospora solani) was shown to be most significantly associated with DRR (Yaseen and D’Onghia 2012; Sandoval-denis et al. 2018). F. solani is an opportunistic pathogen that affects citrus trees when they are stressed (Adesemoye et al. 2011). The combination of abiotic and biotic stressors and F. solani is destructive and might lead the sudden decline of trees (Sandoval-denis et al. 2018). The impact of the disease is most substantial in nurseries and orchards during spring and summer, and significantly damage new plantations. The first symptoms of disease typically initiate after spring temperatures warm, following the rainy season. DRR disease progression depends on weather conditions and decline are most rapid throughout hot summer periods (Adesemoye et al. 2011).
DRR Symptoms begin with the yellowing of leaves from the midrib, chlorosis and wilting, which are followed directly by an entire dieback of the plant. Regardless of rootstock, citrus trees of all ages are vulnerable to DRR (Adesemoye et al. 2011). Sometimes, hemiplegia can occur in diseased trees in the longitudinal direction (Malikoutsaki-Mathioudi et al. 1987). Infected trees lose their canopies as desication, leaf-fall, fruit drying on the branch occurs (Olsen et al. 2000; Kunta et al. 2015). The roots of symptomatic trees are blackened with discolored peels, necrosis and wood browning at the base of the trunk without gummosis (Malikoutsaki-Mathioudi et al. 1987).
The survival and persistence of Fusarium species in the soil and under stressed conditions are typically ensured by the presence of dormant chlamydospores and macroconidia structures. Under favorable environmental conditions, persistent forms of the pathogen germinate and give rise to mycelial filaments that penetrate the cortical cells of roots via natural openings and injuries. Under conditions of stress, the pathogen colonizes vascular tissues by degrading primary cell wall structures.
The genus Fusarium was introduced by Link in 1809 under the name “Fusisporium,” and is currently called Fusarium sensu Wollenweber (1913). It is one of the most studied genera the fungal kingdom. It is a filamentous fungus (Sordariomycetes: Hypocreales: Nectriaceae) that causes devastating diseases in plants (Nelson et al. 1983), and is one of the most diverse fungal species, both morphologically and phylogenetically, and with respect to host specificity. Molecular sequencing techniques have identified more than 300 species and 22 species complexes, which are comprised of more than 300 phytopathogenic species (Aoki et al. 2014). Currently, more than 1400 Fusarium names are listed in MycoBank databases (Sandoval-denis et al. 2018). However, after the introduction of modern molecular tools, the majority of phytopathogenic Fusarium species have been grouped into four species complexes, as follows: F. solani species complex (FSSC), F. solani sensulato, which the members are responsible for foot and root rot in many hosts; F. oxysporum species complex (FOSC), the species cause vascular wilt and rot associated with root rot and include more than 100 special formations (ff. Sp.); F. graminearum species complex (FGSC) and F. fujikuroi species complex (FFSC) (Aoki et al. 2014). Fusarium spp. are known to be a vascular pathogens and have been associated with several plant hosts, including citrus (Timmer 2000; Yaseen and D’Onghia 2012). They present serious threat to citrus production worldwide (Sandoval-denis et al. 2018).
F. solani (Fusarium solani (Martius) Appel & Wollenweber emend. Snyder & Hansen) (teleomorph: Nectriahaematococca; Common Synonym. Nectriahaematococca) is the Fusarium species most frequently isolated from citrus (Adesemoye et al. 2011). It was described for the first time as Fusisporium solani by von Martius (1842), and then transferred named F. solani by Appel and Wollenweber (1910). The name Fusarium solani was attributed to species belonging to FSSC by plant pathologists and clinical microbiologists before the introduction of molecular techniques (Sandoval-Denis and Crous 2018). Lombard et al. (2015) and Sandoval-Denis et al. (2018) grouped species belonging to F. solani species complex (FSSC; O’Donnell 2000/sect. Martiella (Wollenweber 1913) into Neocosmospora. However, throughout the present work, the name Fusarium solani will be used (Liu et al. 2019).
F. solani (as N. solani) belonging to phylogenetic species 5 within FSSC clade 3 (Schroers et al. 2016), has a wide host range and has been reported to be associated with vegetables, humans and animals. It is a ubiquitous soil fungus, and has been associated with citrus tree roots worldwide, notably under conditions of stress (Nemec et al. 1980; Adesemoye et al. 2011). However, other species of Fusarium can cause the same type of disease. These species include F. oxysporum, which has been isolated from the roots and rhizosphere of citrus trees, and F. equiseti, which has been rarely isolated from citrus roots (Smith et al. 1989). F. proliferatum and F. sambucinum have also been isolated from citrus (Malikoutsaki-Mathioudi et al. 1987), but their impact on citrus health is still unknown. Therefore, the goal of the present investigation was to identify Fusarium species associated with severe declines in citrus in Morocco using morphological characteristics, genetic analyses of the ITS region of rDNA, and via the assessment of the capacity of isolates to induce DRR and twig lesions.
Materials and methods
Field sampling, pathogen isolation and purification
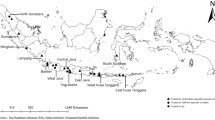
Throughout 2017 and 2018 growing seasons, citrus fields different locations throughout the country were surveyed (Fig. 1) for the presence of key symptoms of DRR and Fusarium dieback such as shrivelling and drying of the foliage, the presence of cracks at the base of the trunk and bark. Root samples from citrus trees showing typical symptoms of DRR, such as light purple coloration and vascular discoloration, were aseptically collected from orchards of key citrus-growing regions of Morocco (Fig. 2). Both the roots and soil were samples. Root and soil samples from each identified citrus tree were transported to the laboratory in separate plastic bags and stored at 4 °C until cultures were isolated from samples.

A map showing sampling fields containing plants examined for dry root rot symptoms. 1 Beau Séjour (Sidi Kacem); 2 Khnichet (Sidi Kacem); 3 Somapra (Khmisset); 4 Lac (Salé); 5 Benioukil (Beni-Mellal); 6 El-Kdima (Taroudant), prepared using ArcGIS software 10.3.1

Infected Trees displaying several symptoms of dry root rot in the field are shown (a–d): a and b indicate sparse canopy and wilted foliage, c indicates fruit persistent in the canopy with detached leaves, and d shows plants at an advanced stage of decline. Infected seedling sections (e) are reddish-purple to black and (f) shows an image of tree bark that was stained a dark color post-Fusarium solani infection
Symptomatic root tissues (roots showing vascular discoloration) were collected from the surveyed fields and rinsed using tap water to remove soil particles. Afterward, they were cut with a sterile scalpel into small pieces (5 × 5 mm) and surface-sterilized in a sodium hypochlorite solution (2%) for 30 s, followed by dipping in 70% ethanol for 30 s, and rinsed 3 times using sterile distilled water (SDW). Tissue pieces were deposited on sterilized Whatman filter paper, air-dried in a Laminar flow hood, and three pieces were placed onto a Petri dish (9 cm in diameter) containing 20 ml of potato dextrose agar (PDA-SC) medium supplemented with streptomycin (100 μg/mL) and chloramphenicol (100 μg/mL). All plates were incubated in the dark at 25 ± 1 °C for at least 4–7 days, and afterward fungal colonies growing on tissue pieces were transferred and subcultured onto new PDA media.
Soil samples (20 g) from each citrus field were suspended in 100 mL SDW, placed in a rotatory shaker set to 150 rpm for 30 min, and serially diluted 10-fold. Three Petri dish plates containing 20 mL PDA were inoculated with 0.1 mL of each diluted soil solution. All plates were incubated at 25 ± 1 °C until colonies of filamentous fungi were visible. Since our study focused on Fusarium species, fungal isolates containing Fusarium-like colonies were purified and transferred to fresh PDA plates for further morphological and molecular characterization.
Morphological identification
Fungal colonies resembling to Fusarium spp. were identified based on their morphological characteristics, as described previously (Leslie and Summerell 2006). Morphological traits including colony color, agar pigmentation on PDA, growth rate, macroconidia produced in sporodochia, and form and arrangement of both microconidia and chlamydospores were examined visually and using a light microscope. Microscopic images of both mycelium and spores, including micro-macroconidia and chlamydospores of each Fusarium species, were taken. The length of micro/macroconidia and chlamydospores was measured using a microscope equipped with a camera (Bx40 with a camera associated with TouchScope Integrated powerful software). Imaging results were compared with those reported in previous studies. Representative isolates of each sampled field were labelled, stored in tubes containing 25% glycerol, and then deposited into the ENA-Meknes (CMENA) microorganism collection.
DNA extraction, PCR amplification and molecular identification of Fusarium species
All suspected Fusarium fungal isolates were subcultured on PDA medium and incubated for 7 days at 25 °C in the dark. Afterward, the mycelium of each isolate was harvested and genomic DNA was extracted as described previously (Doyl and Doyle 1990). Briefly, a small amount of mycelium (~ 1 cm2) was gently scraped from the surface of each pure fresh-growing PDA-SC culture and placed into an Eppendorf tube (1.5 mL), and 500 μL extraction buffer was then added. The mixture was ground with a pestle, vortexed lightly, and incubated at 65 °C in a warm-water bath for 30 min before being briefly centrifuged for 5 min at 13,000 rpm. Supernatant (400 μL) was recovered, and an equivalent volume (400 μL) of chloroform/isoamyl-alcohol (24v/1v) was added. The resulting mixture was gently mixed for 5 min and centrifuged for 5 min at 14,000 rpm. Then, the supernatant (350 μL) was recovered and precipitated with 350 μL of isopropanol. The tubes were then centrifuged for 10 min at 14,000 rpm. The supernatant was then discarded and 500 μL of 70% ethanol was added to the pellet, which was vortexed and centrifuged for 5 min at 14,000 rpm. The obtained pellet (DNA) was dried in an incubator at 60 °C (30 to 45 min), resuspended in 50 μL SDW, and then stored at − 20 °C for further use.
Dry root rot disease is commonly caused by Fusarium species; therefore, a polymerase chain reaction assay (PCR) was used to detect Fusarium spp., F. solani and F. oxysporum. Sets of genus specific primers were used (TEF-Fu3f:GGTATCGACAAGCGAACCAT/TEF-Fu3f:TAGTAGCGGGGAGTCTCGAA) (Arif et al. 2012) to amplify an 420 bp region of transcription elongation factor (TEF-1α) as follows: 5 min at 94 °C; and 40 cycles of 94 °C for 1 min, 58 °C for 1 min and 72 °C for 2 min; followed by a 10 min incubation at 72 °C (Arif et al. 2012). A second PCR assay was carried out to detect F. solani using specific primers (TEF-Fs4f: ATCGGCCACGTCGACTC/TEF-Fs4f: GGCGTCTGTTGATTGTTAGC) that amplified a 680 bp region. The PCR protocol for the amplification step was as follows: 5 min at 94 °C; 40 cycles of 94 °C for 1 min, 58 °C for 1 min and 72 °C for 2 min; followed by 10 min incubation step at 72 °C (Arif et al. 2012). To identify F. oxysporum, specific primers targeting an internal transcribed spacer region (ITS; FOF 1: 5′ ACATACCACTTGTTGCCTCG 3′ FOR1: 5′ CGCCAATCAATTTGAGGAACG 3′) were used to amplify 400 bp region. The PCR program for the amplification used was: 5 min at 95 °C; 35 cycles of 94 °C for 30 s, 55 °C for 30 s 72 °C for 1 min; followed by a 5 min incubation step at 72 °C (Mishra et al. 2003). Amplification of the internal transcribed spacer region was performed using ITS1 and ITS4 (White et al. 1990). The PCR amplifications were carried out using an Eppendorf thermal cycler using the following settings: 3 min at 95 °C; followed by 35 cycles of 35 s at 95 °C, 1 min at 55 °C and 2 min at 72 °C; with a final 10 min extension step at 72 °C.
A final volume of 25 μL was used for all PCR reactions. Reaction ingredients included 2.5 μL PCR buffer, 2 μL MgCl2 (50 mM), 0.25 μL dNTPs (10 mM), 0.25 µL of each primer (10 µΜ), 0.2 µL Taq DNA Polymerase (5 U/μL) (Invitrogen™ Taq DNA Polymerase), 2 μL of genomic DNA and SDW was added to reach a final 25 μL reaction volume. Amplified PCR products were visualized on a 1.5% agarose gel after by electrophoresis using tris borate EDTA buffer (TBE) (0.5×) and a 1-kb DNA ladder was used as a standard. Gels were stained with ethidium bromide and visualized using a UV trans-illuminator to assess the presence and size of amplicons. PCR products were then sequenced. The forward and reverse ITS sequences were edited and aligned using DNAMAN sequence analysis software (version 6.0, Lynnon Biosoft, Canada). The obtained sequences were verified using Blast search to identify the similar sequences in GenBank databases (National Center for Biotechnology Information) and then deposited in GenBank with the accession numbers as indicated in Table 1. Phylogenetic analysis was conducted using sequences of Fusarium spp. determined in the present study (Table 1) and those available in GenBank. The obtained ITS sequences were used to generate a phylogenetic tree, and isolates were rearranged to form major clusters using MEGA 5.0 software. The Rhizoctonia solani (HG934415) sequence from GenBank was selected as an out-group. The maximum likelihood (ML) method was used to estimate phylogenetic relationships. Support for each branch in the inferred tree was evaluated using 1000 bootstrap replications.
Pathogenicity tests
In this study, two separate pathogenicity trials were conducted. For all Fusarium isolates, the first trial was conducted on conditionally healthy one-year-old citrange-Troyer seedlings. Fusarium inoculums were prepared from 14-day-old PDA fungal cultures grown at 25 °C. Spores were harvested by adding a few millimeters of SDW to each Petri plate, and sporal concentrations were adjusted to 1 × 106 conidia/mL using a hemocytometer. Citrus seedlings were gently removed from their substrate, cleared of soils, and their roots were injured (Malikoutsaki-Mathioudi et al. 1987). Then, they were inoculated by dipping their roots into conidial suspensions of each isolate for 30 min and the rest of conidial suspension was added to the pot after replanting to make sure that the inoculum was in contact with the roots. Plants soaked in SDW served as controls. The inoculated seedlings were replanted into new pots containing a sterile soil substrate and kept in a greenhouse (18/26 °C). Plants were watered with normal tap water two to three times weekly and examined each week for disease symptoms. After 2 months, plants were removed from the greenhouse, examined for disease symptoms (Spina et al. 2008), and re-isolation of the pathogen was performed in accordance with Koch’s postulates (Leslie and Summerell 2006). Accordingly, plants were removed from pots, their root systems were separated from the aerials part, and plant height and roots length were assessed. To determine dry weight, aerial and root portions of the plants were cleaned, weighted and dried at 105 °C. Disease severity was assessed based on visual symptoms such as wilting, defoliation and the degree chlorosis observed using a 1–5 scale, which has previously been described (Spina et al. 2008), as follows: 1 (0%) was attributed to healthy plants, 2 (10%) indicated partially defoliated plants with interveinal chlorosis (1–20% of the foliage affected), 3 (35%) indicated partially defoliated plants with interveinal chlorosis (21–50% of the foliage affected), 4 (65%) indicated partially defoliated plants that was accompanied by leaf yellowing (51–80% of the foliage affected) and 5 (90%) indicated plants displaying total defoliation. Sour orange rootstocks have been demonstrated to be resistant to root rot diseases. Representative isolates of F. brachygibbosum K1, F. equiseti K2, F. Solani K3 and F. oxysporum K4 were used to test the pathogenicity of sour orange as described above. Another trial was performed with 8 representative isolates using the mycelial plug method as described in a recent citrus canker study (Sandoval-denis et al. 2018). Citrus seedlings were wounded on their stems and inoculated with a mycelial plug (5 mm in diameter) from the edge of 7-day-old, actively growing colonies. Pathogen-free PDA plugs were used as controls. To prevent desiccation, inoculated wounds were sealed with parafilm and plants were incubated in a greenhouse at 25 °C. Three replicates were performed for each treatment, and each plant was wounded six times. One month postinoculation, the aggressiveness of fungal isolates was assessed by measuring the length of wood discoloration (LWD) observed above and below inoculation sites. All experiments were repeated once.
Statistical analysis
All tests were carried out using a completely randomized design. All datasets were summarized as mean ± SD (standard deviation). To assess disease severity, datasets were transferred to arcsin values. All data obtained were tested for normality using the Shapiro–Wilk test. Dataset of each measured parameters from the first pathogenicity trial failed the normality test and variable transformation was carried out in order to improve normality or homogenize the variables, but with no success. Therefore, to assess the effect of inoculation with each isolate, the nonparametric statistical test Kruskal–Wallis was applied. When the effect was revealed to be significant, pairwise comparison was used to compare each isolate with the control. Dataset of the second pathogenicity trail was square-root transformation to improve normality and stabilize the variance before the two-way analysis of variance (ANOVA). When the effect was revealed to be significant, the Duncan test was performed and means determined to be different for values of p ≤ 0.05. All analyses were performed using SPSS statistical software (version 20, IBM SPSS Statistics 20).
Results
Symptoms of DRR in surveyed citrus orchards
Infected citrus exhibited specific symptoms of DRR, including sudden decline and grayish-brown discoloration of the large wood and collar. This had the potential to lead to the complete decline and death of the tree (Fig. 2). In the orchard, different symptoms of dieback were noted, including mild canopy symptoms (early wilting) (Fig. 2a), sparse canopies, withered foliage, severe twig dieback (Fig. 2b), defoliation with persistence of mature fruit on the canopy (Fig. 2c) and advanced dieback and plant death (Fig. 2d).
Morphological characterization of the causal pathogen of DRR
Based on morphological traits, the results indicated that the fungal pathogen responsible for citrus DRR citrus belongs to the genus Fusarium spp. Four Fusarium species were distinguished from infected citrus roots, which included F. solani, F. oxysporum, F. equiseti and F. brachygibbosum. These different species belong to species complexes a (Fig. 3), b (Fig. 4), c (Fig. 5) and d (Fig. 6), respectively.

Morphological characters of Fusarium solani. a Colonies grown on PDA for 7 days at 25 °C in the dark are shown. b–d Sporodochial conidiophores and phialides, e chlamydospores, f aerial conidia (microconidia) and g sporodochial conidia (macroconidia) are shown (scale bars = 10 μm)

Morphological characters of Fusarium oxysporum. a Colonies grown on PDA for 7 days at 25 °C in the dark are shown. b Chlamydospores, c, d aerial phialides, aerial conidia (microconidia), f sporodochial conidia (macroconidia) are shown (scale bars = 10 μm)

Morphological characters of Fusarium brachygibbosum. a Colonies grown on PDA for 7 days at 25 °C in the dark are shown. b Microscopic images of colonies, c chlamydospores, d–g conidia, d aerial conidia (microconidia) and f–g sporodochial conidia (macroconidia) are shown (scale bars = 10 μm)

Morphological characters of Fusarium equiseti. a Colonies grown on PDA for 7 days at 25 °C in the dark are shown. Chlamydospores are shown in (b), and sporodochial conidia (macroconidia) are shown in (c) (scale bars = 10 μm)
Fungal isolates belonging to FSSC displayed cultural and morphological variability. Cultured colonies of F. solani K3 were white to cream, with sporodochia that were often produced in colors that varied from cream, blue to green. Macroconidia are wide, straight to slightly curved and characterized by a short, blunt and rounded apical cell, and are arranged in shorter and branched conidiophores. Basal cells are straight to almost cylindrical, and usually display a notched or a rounded end. The macroconidia have 3–5 septa and measure 26.2–47.8 × 3.23–6.56 µm. Microconidia are oval, ellipsoid, reniform and produced on elongated and scarified conidiophores. They had 0–3 septae and measured 9.1–23.151 × 3.11–5.03 µm. Chlamydospores appeared after 14 days of incubation. They were abundant, rounded, terminal or interleaving, occurred in singular form or in rough or smooth pairs, were globose or subglobose in shape, and measured 5.56–9.60 µm.
Fungal isolates of FOSC were white with intense aerial mycelium. The inverted surface of most colonies was pink or light to dark violet. Macroconidia range from short to medium length, sickle- to nearly straight-shaped, and are thin-walled with three septa. The apical cell is tapered and curved, short and attached. The basal cell is foot-shaped or notched and measured 19.78–35.91 × 4.24–7.32 µm. Observed microconidia were arranged on short monophialides and were generally observed to be abundant in elliptical, oval or reniform forms and have 0–1 septa that measure 5.38–20.44 × 2.34–6.57 µm. Chlamydospores are produced after 14 days of incubation and measure 8–10 µm in diameter.
Fusarium isolates of FEISC had fast growing colonies, and abundant mycelium that were initially white and later turned brown. Chlamydospores measured 5.6–12.07 μm, and were either isolated or formed short chains on PDA after 14 days of incubation. Macroconidia had strong dorsiventral curvature and 5–7 sections measuring 25.8–48.03 × 3.47–6.03 μm, with a distinctly foot-shaped basal cell and the apical cells were tapered. F. brachygibbosum isolates had intercalary and terminal chlamydospores measuring 5.31–9.484 μm. Macroconidia were slightly curved with three to five septa measuring 25.87–40.4 × 4.5–5.05 μm with slightly hook-shaped apical cells and slightly foot-shaped basal cells. Microconidia were oval-shaped, sometimes ovoid, usually with one septum, 11.5–20.87 × 3.62–5.02 μm.
ITS amplification and phylogenetic analysis
In addition to morphological characterization, the PCR assays were used to assess 34 isolates. Primers specific detection of the Fusarium spp. genus confirmed that all isolates were indeed Fusarium. Several isolates were identified as F. solani and F. oxysporum and the identity of 29 Fusarium isolates out of 34 collected were further confirmed by molecular sequencing of the rDNA internal transcribed spacer (ITS) region using ITS1 and ITS4 primers (Ezrari et al. 2020). Based on morphological traits and phylogenetic analysis, we were able to confirm that Fusarium spp. fungal pathogens are responsible for citrus DRR. Four different Fusarium species were isolated from the citrus roots. Phylogenetic relationships between Fusarium isolates from citrus clustered into four species complexes; F. solani (Martius) Appel and Wollenweber emend. Snyder and Hansen belonging to Fusarium solani species complex (FSSC); F. oxysporum Schlechtendahl emend. Snyder and Hansen belonging to Fusarium oxysporum species complex (FSSC); and two new species found to be associated with decline disease in Morocco, F. brachygibbosum Padwick belonging to the Fusarium sambucinum species complex (FSAMSC) and F. equiseti (Corda) Saccardo belonging to Fusarium incarnatum-equiseti species complex (FIESC) (Fig. 7). Fusarium solani was the most frequently identified species, followed by F. oxysporum, two isolates of F. equiseti and one isolate of F. brachygibbosum.

Phylogenetic tree comprised of Fusarium isolates associated with citrus dry root rot created using an ITS rDNA dataset. The maximum likelihood method was implemented using a Kimura two-parameter model in MEGA X software. The analyses were performed using a heuristic search. The tree was evaluated via 1000 bootstrap replications and was rooted using Rhizoctonia solani (HG934415)
Pathogenicity
The pathogenicity of all isolates against citrange-Troyer plants was assessed. Results showed that Fusarium spp. are pathogenic when wounded plants are inoculated, and produced variable symptoms (Fig. 8). Inoculated seedlings showed root rot, vascular discoloration and wilting. Two month later, plants presented shorter shoots, started wilted leaves and stem drying. Their root systems were reduced in size and had root hairs of reduced length when compared with non-inoculated citrus seedlings. Furthermore, skin desquamation of the epidermis was observed in some plants with severe DRR (Fig. 8a–c). Some plants are survived the infection, however, they displayed necrotic spots in their secondary roots. Not all citrus seedlings inoculated by isolates were killed, and the ultimate stage of infection in which leaf desiccation occurs was not always reached. As a result, the dehydrated plants harbored the fungus only in their root tissues and all root levels were colonized by the pathogenic fungus. In general, symptoms described above were not found in the root systems of uninoculated control seedlings.

Different symptoms caused by inoculation of citrus trees with Fusarium isolates are shown. a–c Symptoms of dry root rot generated by Fusarium spp. on one-year-old citrange-Troyer 60-day postinoculation are shown. Black discoloration of roots (a), desiccation of leaves (b), complete drying (c) and discoloration (d) are shown. d–k Symptoms of dry root rot generated by Fusarium spp. infection in 6-month-old citrus seedlings recorded 60-day postinoculation with F. oxysporum and F. solani (d and e, respectively) are shown. Control plants (f), and yellowed of seedlings inoculated with F. brachygibbosum (g, h), and black discoloration, desquamation and necrotic root spots (i–k) are shown
Table 2 includes information regarding disease severity, plant height, root length, fresh and dry root weight per citrange-Troyer plant 60-day postinoculation. Statistical analyses revealed highly significant differences (p = 0.000) in aggressiveness among the isolates. Furthermore, a significant effect (p < 0.05) of all Fusarium isolates was observed for all evaluated parameters. The average plant height of non-inoculated plants was 101.17 ± 1.09 cm, which differed highly significantly from plant inoculated with F. oxysporum L6 (75.33 ± 0.67) and L4 (76.00 ± 6.00), F. brachygibbosum k1 (76.00 ± 7.09) and F. solani (79.3 ± 3.3). The average of root length (RL) for control plants was 21.17 ± 1.69, which highly significantly differed from 9.33 ± 1.20 to 13.67 ± 2.03 cm values produced by inoculating with K3, L4, L6, BSJ2, L5 and SMP2-1. Plant dry weight (PDW) and root dry weight (RDW) were significantly reduced in plants inoculated with Fusarium spp. relative to uninoculated plants. Observed PDW and RDW values ranged from 5.41 ± 0.37 to 16.43 ± 1.57 g and from 2.53 ± 0.33 to 5.83 ± 0.87 g, respectively, while the average PDW and RDW of control plants was 31.80 ± 0.76 g and 8.03 ± 1.39 g, respectively. Interestingly, our results revealed a significant effect of Fusarium isolate type on disease severity. The most highly aggressive isolates were K3, K5, L3, L7 and SMP2-2 belonging to the Fusarium solani group, and K4, L6 and BM4 belonging to F. oxysporum group.
The representative isolates of all Fusarium species, except F. equiseti, were determined to be pathogenic to sour orange. Under greenhouse conditions, inoculation of plants induced wilting, a significant degree of root browning, root hair loss and the presence of necrotic spots (Fig. 8d–k). Isolated fungal species were re-isolated from the root tissues of inoculated plants and confirmed, in accordance with Koch’s postulates. Moreover, the mycelial plug inoculation method showed that all tested Fusarium spp. isolates were pathogenic and induced visible lesions in citrus seedlings. Within inoculated wounds, the internal discoloration of tissues could be observed (Fig. 8l–p). Conversely, no apparent symptoms were observed in control plants. Interestingly, a very highly significant difference was observed between the aggressiveness of isolates was observed (p = 0.0001). The greatest degree of lesion formation was observed in plants inoculated with F. brachygibbosum, whereas F. oxysporum L1 was the most aggressive among F. oxysporum isolates. Further, no differences were observed among between rootstocks and the grafts tested. F. solani K3 induced larger lesions on rootstocks than grafts. However, F. equiseti K2 was weakly aggressive on rootstocks relative to grafts (Table 3, Fig. 8l–p).
Discussion
For the first time, this study detailed the morphological and the molecular identification of Fusarium species associated with DRR in citrus plantations in Morocco, and tested the pathogenicity of isolates used to inoculate greenhouse-grown plants. Isolates were successfully identified based on morphological traits and molecular characterization of the ITS region of the genome. Obtained results confirmed the existence of a complex of four Fusarium species Fusarium solani, F. oxysporum, F. equiseti and F. brachygibbosum within diseased citrus roots. The disease was detected in a variety of citrus-growing areas including El-Kdima (Taroudante), Lac (Salé) Beau séjour and Khnichet (Sidi Kacem), Somapra (Khmisset) and Benioukil (Beni-Mellal). Similar symptoms to those outlined in this study have previously been described by many authors (Adesemoye et al. 2011; Kunta et al. 2015; Sandoval-denis et al. 2018). In Morocco, citrus plantations are continually under stress as a result of climactic changes, which will likely influence the appearance of disease and intensify the threat of DRR to citrus plantations. This phenomenon might explained the current results in which F. solani, F. oxysporum, F. equiseti and F. brachygibbosum are the most frequently identified Fusarium species associated with citrus decline in Morocco. With the exception of F. brachygibbosum, these species were previously reported to be associated with diseased citrus trees worldwide (Smith et al. 1989; Yaseen and D’Onghia 2012; Sandoval-denis et al. 2018). Species of Fusarium were also reported to be the causal agent of many citrus diseases including wilt, canker damping-off and root rot. F. solani was the species most frequently isolated from diseased plants in studies previously published (Yaseen and D’Onghia 2012; Kunta et al. 2015).
Fusarium species, particularly F. solani and F. oxysporum, have been demonstrated to be present in all citrus orchards worldwide, which probably facilitates the rapid spread of disease as stressful conditions occur (Adesemoye et al. 2011). F. solani has been reported to be present in citrus plantations of Brazil (Paguio 1984), Greece (Malikoutsaki-Mathioudi et al. 1987), South Africa (Labuschagne and Kotze 1988; Van Janse Rensburg et al. 2001), Iles Canaries (Rodríguez 1989), Morocco (El Guilli and Chafai 1995), Italy (Ippolito et al. 1990; Spina et al. 2008; Sandoval-denis et al. 2018), India (Ramakrishnan 1954; Chandran and Kumar 2012), Egypt (El-Mohamedy and Hasabo 2005), Oman (Al-Sadi et al. 2014), Pakistan (Rehman et al. 2012), Iraq (Al-Karboli and Kuthair 2016), China (Liu et al. 2019) and USA, in particular in California (Klotz and Sokoloff 1943; Bender et al. 1982), Arizona (Allen 1962), Florida (Nemec 1975; Graham et al. 1985), and Texas (Kunta et al. 2015; Kunta et al. 2015). While, F. oxysporum was isolated from citrus orchards in Florida (Sherbakoff 1953), California (Nemec 1989), South Africa (Labuschagne et al. 1987) and Tunisia (Khouja et al. 2008). Fusarium equiseti was only found in citrus plantations in Florida (Smith et al. 1989). Recently, Sandoval-Denis et al. (2018) reported the presence of a several-organism complex of Fusarium species in citrus plantations of southern Italy.
This study is the first report the presence of Fusarium equiseti in citrus trees in Morocco. Nevertheless, this pathogenic fungus has been isolated from other diseased crops including zucchini (Ezrari et al. 2019), bread and durum wheat (El Yacoubi et al. 2012; Ennouari et al. 2018). Moreover, the species has also been isolated from citrus roots in Florida, but its role in inducing and affecting the development of disease is not known (Smith et al. 1989). The species is commonly found in soil and plant tissues, and acts as a saprophytic colonizer and invader (Leslie and Summerell 2006). There have been no previous reports regarding an association between F. brachygibbosum and citrus plantations. However, this study revealed for the first time that the species is present in citrus fields surveyed. This finding indicates that the species may be involved in DRR disease progression. This fungus was first described by Padwick in 1945 and was reported to be associated with many other plants. It was identified as a pathogenic fungus, causing disease in crops including sugar beet (Cao et al. 2018), sunflower (Xia et al. 2018), maize (Shan et al. 2016), onion (Tirado-Ramírez et al. 2019), olive, almond trees (Stack et al. 2016), Euphorbia larica (Al-Mahmooli et al. 2013), date palm (Al-Sadi et al. 2012) and legumes (Tan et al. 2011).
In accordance with Koch’s postulate, the pathogenicity of the four Fusarium species isolated from citrus samples against healthy citrus seedlings was assessed using two different inoculation protocols. Results showed that Fusarium isolates were able to induce a variety of disease symptoms, which had the potential to lead to plant death. However, there was significant variation regarding the pathogenicity of Fusarium isolates. In this study, Fusarium isolates produced symptoms of DRR that included browning and wilting. In contrast, numerous studies have reported that reproducing Fusarium disease via artificial inoculation is difficult, since disease onset depends on interactions between biotic and abiotic factors (Graham et al. 1985; Dandurand 1993). In accordance with results reported here, several studies shown that F. solani and F. oxysporum have the capacity to aggressively cause disease in citrus plantations (Nemec et al. 1980; Bender et al. 1982; Malikoutsaki-Mathioudi et al. 1987; Kunta et al. 2015). Since Fusarium spp. induces disease under conditions of stress, several authors have not been able to reproduce symptoms of the disease under experimental conditions (Bender et al. 1982; Strauss and Labuschagne 1995). The disease caused severe root rot in citrus roots, but there was a possibility that it appeared in the epidermis and was triggered by interactions with other citrus soilborne pathogens of citrus, Phytophthora species, CTV viruses, rodents and insects (Bender et al. 1982; Dandurand and Menge 1992; Adesemoye et al. 2011; Al-Sadi et al. 2014). Therefore, the population density of Fusarium spp. in the soil could play a crucial role in triggering disease. The increased initial concentration of F. solani in soil contributes substantially to the predisposition of citrus plants to disease (Nemec et al. 1980; Khouja et al. 2008). In contrast, less virulent Fusarium isolates are able to colonize healthy citrus roots and establish asymptomatic infections (Nemec 1978).
It was observed that symptoms recorded at the end of the pathogenicity test were associated with reduced seedling growth and vigor. These results are in agreement with those of previous studies in which citrus seedlings inoculated with Fusarium species have been shown to be decreased in height, root length and weight relative to healthy plants (Olsen 2000; Spina et al. 2008; Kunta et al. 2015). Similarly, Spina et al. (2008) showed that reduced stem height, root length and weight of citange troyer seedlings was observed in soil infested with F. solani (Nemec and Zablotowicz 1981; Dandurand 1993). Graham et al. (1985) were able to show that F. solani infected only weakened citrus trees and reduced both root growth and vigor. The fungus induced root rot and increased vessel plugging in both the roots and above ground parts of blighted trees (Nemec et al. 1980). In addition, other studies showed that F. solani infection can cause significant reductions in seedling growth, reducing seedling dry weight by about 33% (Olsen 2000). Infection by the fungus is chronic, and significantly decreases the capacity of root to nourish plants (Strauss and Labuschagne 1995).
Fusarium spp. have been shown to be pathogenic invaders of sour orange rootstocks. Furthermore, the availability of rootstocks resistant to F. solani is low (Krueger and Bender 2015). However, the use of grafted citrus plants might provide information regarding the impact of Fusarium spp. infection on the aerial parts of plants and the resulting inhibition of plant development. These findings are line with our results, which in which four Fusarium species induced lesions in inoculated citrus seedlings. Moreover, these species can be added to the list of other Fusarium spp. responsible or citrus canker worldwide (Sandoval-denis et al. 2018).
The management of soilborne diseases caused by pathogenic fungi is a difficult task for farmers. Therefore, increasing our knowledge of the identity of fungal species involved in DRR and the environmental factors that affect its development is essential if we want to adequately control disease. Currently, there is no known means to cure this devastating disease. Consequently, an integrated management approach that is based on a combination of sanitation, culture, use of biological control agents, chemical substances and resistant rootstock varieties has been reported to be the best method for combatting DRR (Abd-Elgawad et al. 2010; Adesemoye et al. 2011; Marais 2015; Khanzada et al. 2016; Parikh et al. 2018a).
Conclusions
Taken together, findings reported here enhance our understanding of the biodiversity of Fusarium species associated with citrus plantations in Morocco. Four species including F. solani, F. oxysporum, F. equiseti and F. brachygibbosum were observed to be associated with DRR disease in citrus, based on morphological and molecular characterization of isolates. Furthermore, F. solani were the species that were most frequently isolated in this study. However, results also showed variability between the four species isolated, with regard to their characteristics in culture, morphological traits and capacity to induce DRR disease in pathogenicity tests. Researchers observed reductions in plant height, root length and the fresh and dry weights of both plants and roots inoculated with fungal strains relative to uninoculated plans. To the best of our knowledge, this is the first detailed report of an association between both F. brachygibbosum and F. equiseti and citrus trees in Morocco.
References
Abd-Elgawad M, El-Mougy N, El-Gamal N, Abdel-Kader M, Mohamed M (2010) Protective treatments against soilborne pathogens in citrus orchards. J Plant Prot Res 50:477–484
Adesemoye A, Eskalen A, Faber B, Bender G, Connell NO, Kallsen C, Shea T (2011) Fusarium dry rot of citrus. Citrograph 2:29–33
Al-Karboli M, Kuthair W (2016) Isolation and pathogenicity of the fungus, Fusarium solani a causal of dry root rot on sour orange in Baghdad province, Iraq. Int J Agric Technol 12:927–938
Allen RM (1962) Dry root rot of citrus induced by ammonia excesses. In: Phytopathology. American Phytopathological Society, St. Paul, MN
Al-Mahmooli IH, Al-Bahri YS, Al-Sadi AM, Deadman ML (2013) First report of Euphorbia larica Dieback caused by Fusarium brachygibbosum in Oman. Plant Dis 97:687
Al-Sadi AM, Al-Hilali SA, Al-Yahyai RA, Al-Said FA, Deadman ML, Al-Mahmooli IH, Nolasco G (2012) Molecular characterization and potential sources of Citrus tristeza virus in Oman. Plant Pathol 61:632–640
Al-Sadi AM, Al-Ghaithi AG, Al-Fahdi N, Al-Yahyai R (2014) Characterization and pathogenicity of fungal pathogens associated with root diseases of citrus in Oman. Int J Agric Biol 16:371–376
Aoki T, O’Donnell K, Geiser DM (2014) Systematics of key phytopathogenic Fusarium species: current status and future challenges. J Gen Plant Pathol 80:189–201
Arif M, Chawla S, Zaidi MW, Rayar JK, Variar M, Singh US (2012) Development of specific primers for genus Fusarium and F. solani using rDNA sub-unit and transcription elongation factor (TEF-1α) gene. Afr J Biotechnol 11:444–447
Bender G, Menge J, Ohr H (1982) Dry root rot of citrus. Its meaning for the grower. Citrograph 67:249–254
Cao S, Yang N, Zhao C, Liu J, Han C, Wu X (2018) Diversity of Fusarium species associated with root rot of sugar beet in China. J Gen Plant Pathol 84:321–329
Chandran MR, Kumar MR (2012) Studies on cultural, morphological variability in isolates of Fusarium solani (Mart.) Sacc., incitant of dry root-rot of Citrus. Curr Biot 6:152–162
Dandurand LM (1993) Influence of Fusarium solani on citrus root growth and population dynamics of Phytophthora parasitica and Phytophthora citrophthora. Phytopathology 83:767–771
Dandurand LM, Menge JA (1992) Influence of Fusarium solani on citrus root rot caused by Phytophthora parasitica and Phytophthora citrophthora. Plant Soil 144:13–21
Doyl JJ, Doyle JL (1990) Isolation of plant DNA from fresh tissue. Focus (Madison) 12:13–15
El Guilli M, Chafai N (1995) Symptomes et etiologie de la pourritture sèche des agrumes au Maroc. Alawamia 83:103–110
El Yacoubi H, Hassikou R, Badoc A, Rochdi A, Douira A (2012) Complexe Fongique De La Pourriture Racinaire Du Ble Tendre Au Nord-Ouest. Bull la Société 592 Pharm Bordeaux 151:35–48
El-Mohamedy RSR, Hasabo SA (2005) Response of some citrus rootstocks to infection with Fusarium solani and citrus nematode Tylenchulus semipenetrans under greenhouse conditions. Egypt J Phytopathol 33:11–25
Ennouari A, Sanchis V, Rahouti M, Zinedine A (2018) Isolation and molecular identification of mycotoxin producing fungi in durum wheat from Morocco. J Mater Environ Sci 9:1470–1479
Ezrari S, Lahlali R, Radouane N, Tahiri A, Lazraq A (2020) First report of Fusarium equiseti causing pre-and postharvest fruit rot on zucchini in Morocco. J Plant Pathol 102(1):251
Graham JH, Brlansky RH, Timmer LW, Lee RF, Marais LJ (1985) Comparison of citrus tree declines with necrosis of major roots and their association with Fusarium solani. Plant Dis 69(12):1055–1058
Guarnaccia V, Crous PW (2017) Emerging citrus diseases in Europe caused by species of Diaporthe. IMA Fungus 8:317–334
Hannachi I, Rezgui S, Cherif M (2014) First report of mature citrus trees being affected by Fusarium wilt in Tunisia. Plant Dis 98:566
Ippolito A, De Cicco V, Salerno M (1990) Role of Phytophthora spp. in citrus decline in Apulia and Basilicata, Italy 1. EPPO Bull 20:91–94
Jaouad M, Moinina A, Ezrari S, Lahlali R (2020) Key pests and diseases of citrus trees with emphasis on root rot diseases: an overview. Mor J Agri Sci 1(3):149–160
Khanzada MA, Tanveer M, Maitlo SA, Hajano J, Ujjan AA, Syed RN, Lodhi AM, Rajput AQ (2016) Comparative efficacy of chemical fungicides, plant extracts and bio-control agents against Fusarium solani under laboratory conditions. Pak J Phytopathol 28:133–139
Khouja HR, Yaseen T, Cherif M, Ippolito A (2008) Etiological and epidemiological aspects of dry root rot in nurseries and orchards in Tunisia. Proc Int Soc Citric
Klotz LJ, Sokoloff VP (1943) The possible relation of injury and death of small roots to decline and collapse of citrus and avocado. Calif Citrogr 28:86–87
Krueger RR, Bender GS (2015) Screening a core collection of citrus genetic resources for resistance to Fusarium solani. Acta Hortic 1065:155–164
Kunta M, Salas B, Gonzales M, da Graça JV (2015) First report on citrus dry rot in sour orange rootstock in Texas. J Citrus Pathol 2
Labuschagne N, Kotze JM (1988) Factors affecting feeder root rot of citrus caused by Fusarium solani. In: Citric Proc Sixth Int Citrus Congr Middle-East, Tel Aviv, Isr March 6-11, 1988/Sci Ed R Goren K Mendel, Ed N Goren
Labuschagne JF, Kotzé JM, Putterill JF (1987) Incidence of Fusarium solani and F. oxysporum in citrus roots and infection by F. solani. Phytophylactica 19:315–318
Leslie JF, Summerell BA (2006) The Fusarium laboratory manual. Iowa Blackwell Publ CrossRef Google Sch, Ames
Liu HF, Zhou J, Liao J, Yi JP, Ma DF, Deng JX (2019) Grafted twig rot on Citrus sinensis caused by a member of the Fusarium solani species complex. Can J Plant Pathol 42:133–139
Lombard L, van der Merwe NA, Groenewald JZ, Crous PW (2015) Generic concepts in Nectriaceae. Stud Mycol 80:189–245
Malikoutsaki-Mathioudi M, Bourbos VA, Skoudridakis MT (1987) La pourriture sèche des racines—une maladie très grave des agrumes en Grèce. EPPO Bull 17:335–340
Marais LJ (2015) Efficacy of water soluble silicon in managing Fusarium dry root rot of citrus. Acta Hortic 1065:993–1000
Mishra PK, Fox RTV, Culham A (2003) Development of a PCR-based assay for rapid and reliable identification of pathogenic Fusaria. FEMS Microbiol Lett 218:329–332
Nelson PE, Toussoun TA, Marasas WFO (1983) Fusarium species: an illustrated manual for identification
Nemec S (1975) Vessel blockage by myelin forms in citrus with and without rough-lemon decline symptoms. Can J Bot 53:102–108
Nemec S (1978) Symptomatology and histopathology of fibrous roots of rough lemon (Citrus limon) infected with Fusarium solani. Mycopathologia 63:35–40
Nemec S (1989) Effects of dihydrofusarubin and isomarticin from Fusarium solani on carbohydrate status and metabolism of rough lemon seedlings. Phytopathology 79:700
Nemec S, Zablotowicz RM (1981) Effect of soil temperature on root rot of rough lemon caused by Fusarium solani. Mycopathologia 76:185–190
Nemec S, Baker R, Burnett H (1980) Pathogenicity of Fusarium solani to citrus roots and its possible role in blight etiology. Proc Fla State Hort Soc 93:36–41
Olsen M (2000) Diseases of citrus in Arizona. Coll Agric Life Sci Univ Arizona (Tucson, AZ), pp 1–15
Olsen M, Matheron M, McClure M, Xiong Z (2000) Diseases of Citrus in Arizona. Coll Agric Life Sci Univ Arizona (Tucson, AZ):1–15
Paguio OR (1984) Citrus declinio in the State of Brazil: occurrence and responses to blight diagnostic tests. In: International organization of citrus virologists conference proceedings (1957–2010)
Parikh L, Eskelson MJ, Adesemoye AO (2018a) Relationship of in vitro and in planta screening: improving the selection process for biological control agents against Fusarium root rot in row crops. Arch Phytopathol Plant Prot 51:156–169
Parikh L, Kodati S, Eskelson MJ, Adesemoye AO (2018b) Identification and pathogenicity of Fusarium spp. in row crops in Nebraska. Crop Prot 108:120–127
Ramakrishnan TS (1954) The deterioration of mandarin orange in Madras state. South Indian Hortic 2:52–56
Rehman A, Rehman AU, Javed N, Ullah Malik A, Mehboob S (2012) Toxin production by Fusarium solani from declining citrus plants and its management. Afr J Biotechnol 11:2199–2203
Rivera-Jiménez MN, Zavaleta-Mancera HA, Rebollar-Alviter A, Aguilar-Rincón VH, García-de-los-Santos G, Vaquera-Huerta H, Silva-Rojas HV (2018) Phylogenetics and histology provide insight into damping-off infections of ‘Poblano’ pepper seedlings caused by Fusarium wilt in greenhouses. Mycol Prog 17:1237–1249
Rodríguez RR (1989) Resultados preliminares de la aplicación de métodos, de reconocimiento del “tizón” de los cítricos (Citrus Blight), a árboles en Gran Canaria. Levante Agrícola Rev Int cítricos 208–213
Sandoval-Denis M, Crous PW (2018) Removing chaos from confusion: assigning names to common human and animal pathogens in neocosmospora. Persoonia Mol Phylogeny Evol Fungi 41:109–129
Sandoval-denis MP, Guarnaccia V, Polizzi G, Crous PW (2018) Symptomatic citrus trees reveal a new pathogenic lineage in Fusarium and two new neocosmospora species. Persoonia Mol Phylogeny Evol Fungi 40:1–25
Schroers HJ, Samuels GJ, Zhang N, Short DPG, Juba J, Geiser DM (2016) Epitypification of Fusisporium (Fusarium) solani and its assignment to a common phylogenetic species in the Fusarium solani species complex. Mycologia 108:806–819
Shan LY, Cui WY, Zhang DD, Zhang J, Ma NN, Bao YM, Dai XF, Guo W (2016) First report of Fusarium brachygibbosum causing maize stalk rot in China. Plant Dis 101:837
Sherbakoff CD (1953) Fusaria associated with citrus feeder roots in Florida. Phytopathology 43:395–397
Smith GS, Nemec S, Gould AB, Sonoda RM (1989) Effect of deep-tillage and soil amendments on growth of rough lemon citrus and root and soil microflora population densities
Spina S, Coco V, Gentile A, Catara A, Cirvilleri G (2008) Association of Fusarium solani with rolABC and wild type troyer citrange. J Plant Pathol 90:479–486
Stack AJ, Yaghmour MA, Kirkpatrick SC, Gordon TR, Bostock RM (2016) First report of Fusarium brachygibbosum causing cankers in cold-stored, bare-root propagated almond trees in California. Plant Dis 101:390
Strauss J, Labuschagne N (1995) Pathogenicity of Fusarium solani isolates on citrus roots and evaluation of different inoculum types. Appl Plant Sci 92:48–52
Tan DC, Flematti GR, Ghisalberti EL, Sivasithamparam K, Chakraborty S, Obanor F, Barbetti MJ (2011) Mycotoxins produced by Fusarium species associated with annual legume pastures and ‘sheep feed refusal disorders’ in Western Australia. Mycotoxin Res 27:123–135
Timmer LW (2000) Fusarium wilt. Compend Citrus Disease
Tirado-Ramírez MA, López-Orona CA, Díaz-Valdés T, Velarde-Félix S, Martínez-Campos AR, Romero-Gómez SJ, Retes-Manjarrez JE (2019) First report of basal rot of onion caused by Fusarium brachygibbosum in Sinaloa, Mexico. Plant Dis 103:582
Van Janse Rensburg JC, Labuschagne N, Nemec S (2001) Occurrence of Fusarium-produced naphthazarins in citrus trees and sensitivity of rootstocks to isomarticin in relation to citrus blight. Plant Pathol 50:258–265
Vanderweyen A, Serrhini M. (1981) La pourriture seche des racines d’ agrumes au Maroc [Fusarium sp.]. Comptes Rendus des Seances l’Academie d’Agriculture Fr v. 67
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Prot Guid Methods Appl 18:315–322
Wollenweber HW (1913) Studies on the Fusarium problem. Phytopathology 3:24–50
Xia B, Hu JY, Zhu XF, Liang Y, Ren X, Wu YH, Chen DX (2018) First report of sunflower broomrape wilt caused by Fusarium brachygibbosum in China. Plant Dis 102:2372
Yaseen T, D’Onghia AM (2012) Fusarium spp. associated to citrus dry root rot: an emerging issue for Mediterranean citriculture. Acta Hortic 940:647–656
Acknowledgments
This study was supported by the Phytopathology Unit of Ecole Nationale d’Agriculture de Meknes. We are thankful for Providence Verte group-Louata and Mr. Tijani Bihi (Domaine Dar Esslaem) for providing us with citrus seedlings. The first author wishes to acknowledge the CNRST 'Centre National pour la Recherche Scientifique et Technique' for a scholarship grant.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Ezrari, S., Lahlali, R., Radouane, N. et al. Characterization of Fusarium species causing dry root rot disease of citrus trees in Morocco. J Plant Dis Prot 128, 431–447 (2021). https://doi.org/10.1007/s41348-020-00392-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41348-020-00392-0