
Abstract
In this study, biological features of Sphyraena chrysotaenia Klunzinger, 1884 determined in Iskenderun Bay (Turkey). A total of 560 individuals were examined by sampling monthly from September 2015 to August 2016. The examined samples were 225 (40.18%) females and 335 (59.82%) males. The total length distribution of S. chrysotaenia ranged from 14.0 to 34.5 cm and the weight distribution from 17.0 to 222.0 g. Growth equations for females and males were calculated as respectively W = 0.0117xL2.7922 and W = 0.0097xL2.8517. The growth of S. chrysotaenia was determined as negative allometric. The minimum and maximum ages determined based on otolith readings for females and males ranged from 1 to 6. The von Bertalanffy growth parameters were L∞= 58.682 cm, K = 0.095 year− 1, t0= -2.655 years for females, L∞= 58.907 cm, K = 0.090 year-1, t0= -2.686 years for males, and as L∞= 58.470 cm, K = 0.091 year− 1, t0= -2.647 years for both sexes. Fulton condition factor (CF) value was estimated as 0.6105 ± 0.028, for females, as 0.6092 ± 0.024 for males and as 0.6097 ± 0.018 for combined sexes. The total mortality rate (Z) was calculated as 0.503, the natural mortality rate (M) 0.222, and the fishing mortality rate (F) 0.281. Besides, the fishing mortality (F) was estimated higher than the biological reference points (Fopt =0.111 and Flimit =0.148). The estimated fishing mortality rate and exploitation rate (E = 0.55) obtained results in this study indicate a slightly high fishing pressure on the S. chrysotaenia stock in Iskenderun Bay. Besides, these data were discussed with the other geographical areas growth studies of S. chrysotaenia in the Mediterranean waters. To the best knowledge of the authors, this study presented the first reference on age, growth and mortality rates for this species; also this study will be useful for fishery biologists and managers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The yellowstripe barracuda, Sphyraena chrysotaenia Klunzinger 1884, is an Indo-Pacific species belonging to the Sphyraenidae family (De Sylvia & Williams, 1986; Froese & Pauly, 2021). S. chrysotaenia is distributed in pelagic and demersal zones and usually inhabits large schools in inshore waters often close to surface between 1 and 50 m depths and that it is not found in deeper than 50 m (Golani et al., 2006). This carnivorous species, feed on small fishes (clupeids and anchovy) and also crustaceans. Adult individuals are found in the open waters, while juveniles often occur in very shallow waters (Golani et al., 2002).
The yellowstripe barracuda, S. chrysotaenia has a widespread distribution from the Indo Pacific, to the Red Sea, the Persian Gulf, and East Africa throughout the Indian Ocean to Australia, Japan, southern Adriatic Sea, Libya, Malta, and also in the Eastern Mediterranean Sea (Ben-Tuvia, 1986; Froese & Pauly, 2021). Besides, this species was observed in the Aegean Sea, Ionian Sea, Italian coast, and Tunisian waters (Golani, 1998).
The first record of S. chrysotaenia as misidentification of Belone acus from the Eastern Mediterranean was reported by Spicer (1931) off Palestine in 1931 and then this species was recorded from Libya, Malta waters (Stirn, 1970; Lanfranco, 1993), and north to the southern Adriatic Sea (Pallaoro & Dulcic, 2001). S. chrysotaenia can easily be distinguished by other Levantine barracudas by the pectoral fin tip is reach the vertical of first dorsal fin origin and, second dorsal, pectoral, caudal fin yellowish (Golani et al., 2002).
S. chrysotaenia was first recorded in İskenderun on the northeastern Mediterranean coast of Turkey in 1957 by Akyuz (Akyuz, 1957). Other successive records of the species from the Turkish coast between Iskenderun Bay and Mersin were presented by Ben-Tuvia (1966) and Gücü et al. (1994). Later this species was successively recorded in the south Aegean Sea (Kusadasi Bay, Aegean Sea, Turkey and Diafani, Karpathos, Greece) (Golani & Ben-Tuvia, 1995).
Although S. chrysotaenia is a lessepsian migratory species that entered the Mediterranean from the Red Sea via the Suez Canal, this species is the most common species of barracuda caught by artisanal fisheries in the Eastern Mediterranean. Besides, this species for Turkish Seas has high economic value and, it is common in the Mediterranean coast of Turkey and frequently caught inshore waters using gillnets and by trawlers at depths down to 45 m.
There were many studies on the length and weight of S. chrysotaenia in the Mediterranean Sea (Taskavak & Bilecenoglu, 2001; Ceyhan et al., 2009; Erguden et al., 2009; Apostolidis & Stergiou, 2014; Bílge et al., 2014). However, little information is available on the age and growth of S. chrysotaenia in the Mediterranean (Allam et al. 2004; Zouari-Ktari et al., 2007; Zouari-Ktari et al., 2009; El Ganainy et al., 2017).
To date, there is no available data regarding comprehensive biology data of S. chrysotaenia for the northeastern Mediterranean, Turkey, and so the objectives of this paper are to present study provides the first information on age, growth, and mortality rates of the yellowstripe barracuda, S. chrysotaenia from the Iskenderun Bay (NE Mediterranean, Turkey).
Materials and methods
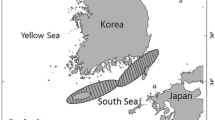
The yellowstripe barracuda, Sphyraena chrysotaenia were obtained from the İskenderun Bay between September 2015 and August 2016 using gillnet and commercial trawler (mesh size 44 mm) at depths ranging 12 to 44 m (Fig. 1).

The study area in the Iskenderun Bay (Northeastern Mediterranean), Turkey
All fish were put in crushed ice and transported to the laboratory immediately for analysis. A total of 560 specimens were measured to the nearest 1 mm (total length, TL) and weighed to the nearest 1 g (total weight, TW). Samples were dissected, and gonads were examined in order to determine their sex.
The length-weight relationship was calculated using the equation W = aLb (Ricker 1975). This equation can also be expressed in logarithmic form: log TW = log a + b log L, where W is the total weight, L is the total length, a and b parameters of the equation. The significance of the regression was tested by analysis of variance ANOVA. Students t-test was applied to determine the significance of differences (95% level) to verify if it was significantly different from 3 (Zar, 1999).
Age was determined by reading the sagittal otoliths. All otoliths were cleaned in ethanol and then immersed in glycerin for examination. A binocular microscope with reflected light was used for age determination. To minimize misreading, the number of translucent rings outside the nucleus was evaluated by three independent readers. Estimates of the precision of growth annuli (ring) count between readers were determined using the average percentage error (APE) of (Beamish & Fournier, 1981).
The von Bertalanffy growth equation was calculated according to Lt = Ls [1-e− k(t−t0)] for total length, TL, where Lt is fish length (cm), Ls is the ultimate length an average fish could achieve, t is the fish age (years), t0 (years) is the hypothetical time at which the fish length is zero, and k is the growth coefficient (years− 1) (Sparre & Venema, 1998).
Fulton’s coefficient of condition factor (CF) was calculated by CF = W/L3 × 100, where; L is length (cm) and W is weight (g) (Fulton, 1904; Le Cren, 1951; Sparre & Venema, 1998). The t-test was applied to confirm whether CF varied by gender and months.
The growth performance index (φ´) was used φ´=logK + 2logL∞ as suggested by Pauly and Munro (1984). The growth performance index was estimated to compare growth parameters obtained in this study with those reported by different authors.
The fishing mortality (F) was calculated following Pauly (1980): F = Z – M. Total mortality rate (Z) was calculated from the length converted catch curve Pauly (1983) using the FISAT program (Gayanilo et al., 1994). The natural mortality (M) was estimated from the empirical formula proposed for the Mediterranean fishes by Djabali et al. (1993): log M = 0.736–0.114 log L∞ + 0.522 log K + 0.583 log T, where the T value was taken as 22.6 °C as this value represented the mean water temperature in the Iskenderun Bay (Gücel and Sakalli, 2018).
The exploitation rate was estimated using this equation E = F/Z, where F: fishing mortality, Z: total mortality (Z), (Sparre & Venema, 1998).
The fishery resource status was assessed according to Patterson (1992) by the comparability of the current fishing mortality rate with the optimum or the target (Fopt) and limit (Flimit) biological reference points which were defined as; Fopt = 0.5 M and Flimit = 2/3 M. On the other hand, optimum exploitation rate (Eopt) was determined using Gulland (1971) equation: Eopt = Fopt (M + Fopt)−1.
Results
Fish composition and measurements
A total of 560 yellowstripe barracuda, S. chrysotaenia (335 males and 225 females), individuals were collected from the Iskenderun Bay. The population comprised 59.82% of males and 40.18% of females. The sex ratio for male and female individuals (M: F) was 1.00:0.67. The chi-square analysis (χ²) showed that the sex ratio was significantly different from the expected 1:1 ratio (χ², P < 0.05).
Total length in males ranged from 14.00 to 34.50 cm with a mean of 23.19 ± 4.087, and total length in females ranged from 15.50 to 33.60 cm with a mean of 23.15 ± 3.338 cm. The mean total weight of males and females values were 82.11 ± 43.020 and 79.71 ± 35.608 g, respectively (Table 1). The maximum total length and the total weight of S. chrysotaenia were 34.50 cm and 222.09 g. There was no significant statistical difference between sexes in overall total length and total weight values of male and female individuals (ttest, P > 0.05). The dominant length class in all individuals was 20.0–22.0 cm, TL. Besides, males dominated the all-length class (Fig. 2).

The length-frequency distributions for male, female, and both sexes of Sphyraena chrysotaenia from Northeastern Mediterranean
Length-weight relationship
The length-weight relationship of S. chrysotaenia was estimated as logW=-2.013 + 2.851logL (R2 = 0.982) for males; logW= -1.931 + 2.791logL (R2 = 0.975) for females and logW= -1.991 + 2.834logL (R2 = 0.980) for both sexes (Table 2). Length and weight relationships for the males, the females, and both sexes were shown in Fig. 3. The coefficient of determination (R2) was found to be > 0.97, a highly significant value of the result. The regression analysis has shown that fish length had a highly significant correlation with weight (P < 0.001).

Length–weight relations for male (A), female (B), and both sexes (C) of Sphyraena chrysotaenia from the Northeastern Mediterranean, Turkey
The exponent of the b parameter indicated negative allometry. The b values showed no significant difference for males, females, and both sexes (P > 0.05). The comparison of length-weight parameters calculated in this study as well as in previous studies is reported in Table 3.
Age and growth estimation
The age structures estimated by sagittal otoliths of S. chrysotaenia ranged from 1 to 6 years for males and females. The year class 2 (33.57%) was dominant, followed by year classes 3 (31.71%), 4 (15.89%), 5(8.7%5) and 6 (7.32%). The total length frequency analysis of S. chrysotaenia from the Northeastern Mediterranean Sea is given in Table 2.
The mean average percentage error (APE) was found as 7.1% by three independent readers for S. chrysotaenia. Three readers have estimated the same age in 526 of 560 samples. The agreement between three readers on all age assessments was 94%. The difference between readers in the accuracy of age assessment was found insignificant when repeated annuli counts were compared (paired t-test, P > 0.05).
The size of S. chrysotaenia to age data was fitted for each sex (male and female) by using von Bertalanffy growth function (VBGF) in order to estimate the growth pattern. The length of individuals assigned to each sex age group of the von Bertalanffy growth parameters was presented in Fig. 4. The von Bertalanffy growth function (VBGF) showed that the male and female fish had a similar growth rate (k = 0.09 y–1). The von Bertalanffy growth parameters were calculated as L∞= 58.47 cm, k = 0.091 year-1, t0 = -2.647 years for males; L∞= 58.90 cm, k = 0.090 year-1, t0 = -2.686 years for females; and L∞= 58.68 cm, k = 0.095 year-1, t0 = -2.655 years for both sexes. The growth performance index (φ´) was found to be 2.492 for males, 2.494 for females, and 2.514 for both sexes (Table 4). The comparisons of von Bertalanffy growth constants for different areas of S. chrysotaenia are given in Table 4.

von Bertalanffy growth curves for male and female of Sphyraena chrysotaenia by length at age data
Condition factor
In the study, the condition factor (CF) value was estimated as 0.6105 ± 0.028, for females, 0.6092 ± 0.024 for males, and 0.6097 ± 0.018 for both sexes. Condition factor values showed no significant variations (P > 0.05) of S. chrysotaenia between the sexes. Monthly condition factor values for females and males were presented in Fig. 5. The highest value of condition factor was shown in October 2015 for females and in March 2016 for males. The lowest value of condition factor was in May 2016 for females and males (Fig. 5). The monthly variations of the condition factor were not significantly different during all months (p > 0.05).

Monthly average condition factor values of Sphyraena chrysotaenia for females and males
Mortality rates
The total mortality rate (Z) for both sexes calculated was as 0.503 year-1. The natural mortality rate (M) computed was 0.222 year-1, and the fishing mortality (F) rate was 0.281 year-1. The exploitation ratio was estimated as E = 0.558 and the biological reference points were calculated as; the optimum fishing mortality, Fopt = 0.111; the fishing mortality limit, Flimit = 0.148, and also optimum exploitation rate was estimated as Eopt= 0.333.
Discussion
Fish composition and measurements
In this study, carried out in Iskenderun Bay (Northeastern Mediterranean, Turkey), the sex ratio of males to females was 1.00:0.67 (335:225) for S. chrysotaenia. Besides, it was also observed that males were more dominant in the population than females. Zouari-Kitari et al. (2019) similar reported to the male-dominant population of S. chrysotaenia from the central Mediterranean (Gulf of Gabes), Tunisian with a male to female ratio of 1.00:0.84. This small difference in sex ratio could be explained by the seasonal variations in populations, the age of sexual maturity, feeding, and also spawning grounds.
The total length and weight of both sexes ranged from 14.0 to 34.50 cm and from 17.03 to 222.09 g, respectively (Table 1). The size ranges (TLmin: 14.0 cm and TLmax: 34.50 cm) of S. chrysotaenia in the present study are close to those in the Mediterranean coast of Tunisia (Zouari-Ktari et al., 2007; Zouari-Ktari et al., 2009), Egypt (El Ganainy et al., 2017) and Turkey (Ceyhan et al. 2009; Bílge et al., 2014). On the other hand, the size range of S. sphyraena in the present study is not in accordance with those recorded in the Mediterranean coast of Turkey (Taskavak & Bilecenoglu, 2001). A detailed comparisons to other populations in the Eastern Mediterranean Sea according to length intervals was given in Table 3. However, our present study was first reported the larger specimen 34.50 cm in total length in the Mediterranean. These differences in overall total length can be explained by different sampling equipment, geographical areas, and depths.
Length-weight relationship
The age classes ranged from 1 to 6 years for males and females. The 2 age class was dominant entire the population (Fig. 4). The maximum length at age was determined as 34.5 cm at 6 age for individuals. The comparable maximum ages in previous studies include age 5+ (28.3 cm) by Zouari-Ktari et al. (2007) and age 5 (28.5 cm) by El Ganainy et al. (2017). Similarly, Wadie & Riskallah (2001) reported age 6 groups of S. chrysotaenia in the coast of the southeastern part of the Mediterranean Sea using size-frequency distribution.
The value of the b, showing the nature of growth in fish which varies between 2 and 4 (Tesch, 1971). The b value in fish may vary according to age and sex. In this study, the b (2.834) values were generally in agreement with results of S. chrysotaenia from other geographical areas (Taskavak & Bilecenoglu, 2001; Kulbicki et al., 1993; Zouari-Ktari et al., 2009; El Ganainy et al., 2017). However, Ceyhan et al. (2009), Erguden et al. (2009), and Bílge et al., (2014) reported different b values (positive allometry) than are presented in this paper for S. chrysotaenia from the Southern Aegean Sea and Eastern Mediterranean Sea, Turkey. These differences in the b value may be related to the combination of one or more factors such as sampling area, habitat, season, maturity, sex, and preservation techniques (Bagenal & Tesch 1978).
Age and growth estimation
The von Bertalanffy growth parameters derived in the present study are generally different from previous studies (Table 4). In our computations, the asymptotic length of S. chrysotaenia was L∞= 58.90 cm for females and L∞= 58.47 cm for males. Kulbicki et al. (1993) estimated L∞= 26.50 cm for New Caledonia, while according to Allam et al. (2004), the von Bertalanffy growth parameters for the Egyptian Mediterranean waters (Egypt) were L∞= 27.10 cm and k = 0.390 year− 1. Besides, similar growth for S. chrysotaenia from the Gulf of Gabes (Eastern Mediterranean) was L∞= 28.3 cm and k = 0.319 year− 1. Apostolidis & Stergiou (2014) and El Ganainy et al. (2017) reported growth parameters for S. chrysotaenia from Mediterranean waters and Gulf of Suez (Egypt) as L∞= 25.0 cm and k = 0.585 year− 1 and L∞= 29.3 cm and k = 0.480 year− 1 respectively.
Our estimates of the growth parameters from the Iskenderun Bay (northeastern Mediterranean) are higher than those of the others, and these differences with previous studies can be explained partly by the maximum recorded length values for S. chrysotaenia individuals in this study. Wotton (1990) stated that fish populations of the same species from different geographical areas may exhibit highly variable, individual growth rates. On the other hand, some differences between growth characteristics may probably be from the quality of food and related to the water temperature (Santic et al., 2002).
The growth performance index (φ´) takes into account the correlation between L∞ and k. The growth performance index for the S. chrysotaenia in Iskenderun Bay was 2.514 for both sexes. The overview of growth parameters and growth performance index obtained from previous studies for S. chrysotaenia are given in Table 4. This obtained value (φ´) is quite similar to that estimated values for the same species from the Gulf of Suez (Egypt), Egyptian and Tunisian Mediterranean waters (Allam et al., 2004; Zouari-Ktari et al., 2009; El Ganainy et al., 2017) and also Eastern Mediterranean (Apostolidis & Stergiou, 2014). These results are indicating that the environmental conditions are well suitable for the growth of S. chrysotaenia in the Mediterranean waters.
Condition factors
Condition factor values were also calculated, where the lowest and highest estimations were found to be 0.5296 and 0.7274, respectively, with a mean value of 0.6105 ± 0.028 for females and 0.5180 and 0.7326, respectively, with a mean value of 0.6092 ± 0.024 for males. Zouari-Ktari et al. (2009) reported the condition of S. chrysotaenia from the central Mediterranean waters for females (0.9866 − 0.9670) and males (1.0173–0.9372). If the condition factor is less than 1.0, this value indicates slow growth, possibly due to stress, overcrowding, diseases, low productivity. According to Le Cren (1951) and Ricker (1975) condition factor of fish populations may reveal variations with gonad development, feeding activity, and seasonal changes in growth. The obtained results showed that the condition factor (< 1) of S. chrysotaenia in Iskenderun Bay was indicated slow growth.
Mortality rates
In this study, the total mortality rate (Z) of S. chrysotaenia individuals estimated from the catch curve was 0.50 year− 1. The fishing mortality rate (F) was 0.28 year− 1 and the natural mortality rate (M) was 0.22 year− 1 for all S. chrysotaenia individuals. Allam et al. (2005) are found fairly higher the total mortality of S. chrysotaenia according to the Egyptian Mediterranean coast (Z = 1.05 year− 1). However, the natural mortality rates for S. chrysotaenia (M = 0.39), for S. flavicauda (M = 0.32), and S. sphyraena (M = 0.20) from the Egyptian Mediterranean coast were reported similar to our study by Allam et al. (2005). Besides differently, the mean instantaneous natural mortality of the 4 different methods (Taylor’s, Pauly’s, Ralston’s, and King’s) of S. chrysotaenia from the Gulf of Gabes (South Tunisian coast) was stated as 0.652 by Zouari-Ktari et al. (2007).
Allam et al. (2005) reported of the fishing mortality rates (F) for S. chrysotaenia (F = 0.65), S. flavicauda (F = 0.49) and S. sphyraena (F = 0.58). The fishing mortality rate (F = 0.28) of S. chrysotaenia in the present study is lower than the values obtained (F = 0.65) in the Egyptian Mediterranean coast. According to these reported values, the total mortality (Z), the fishing mortality rate (F), and the natural mortality rate (M) in this study were found comparatively lower than in previous studies. These differences may be explained by fishing effort, biological features, behavior characteristics as well as ecological factors in the different geographical regions.
Pauly (1980) claimed that natural mortality for marine species of 175 fish stocks showed overall modal mortality of M = 0.2–0.3 year− 1. According to this value, the natural mortality rate of S. chrysotaenia gives similar results to the Pauly (1983). Therefore, the natural mortality rate seems to be optimum for S. chrysotaenia. On the other hand, Sparre & Venema (1998) also indicated values of natural mortality coefficients for the same species may be different in distinct areas, depending on the density of predators and competitors whose abundance is other influenced by fishing activities.
Gulland (1971) declared that the exploitation ratio should not be more than 0.5 as optimum exploitation. Gulland (1983) also suggested that in an optimally exploited stock the fishing mortality rate (F) is equal to the natural mortality rate (M) or the rate of exploitation (E) is equal to 0.5.
In the present study, the exploitation ratio was estimated as E = 0.55. Allam et al. (2005) reported the exploitation ratio to a higher value for S. chrysotaenia (E = 0.62) in the Egyptian Mediterranean waters. Although the exploitation rate (E = 0.55) was close to Gulland (1983). This value indicated that fishing pressure on S. chrysotaenia in Iskenderun Bay is slightly high. However, the fishing mortality estimated was lower than in the Egyptian Mediterranean waters. These differences probably are appeared to be area-specific and may be related to the density of fishing pressure for the Northeastern Mediterranean (Iskenderun Bay).
Estimating parameter of the fishing mortality rate (F = 0.28) was higher than the biological reference points derived from the equation of Patterson (1992) as follows: (Fopt = 0.11 and Flimit = 0.14); additionally the exploitation rate (0.55) was found to be higher than the optimum exploitation rate (Eopt = 0.33). Thus, S. chrysotaenia in Iskenderun Bay appears to be over-exploited as the fishing mortality (F) is higher than the defined values of reference points Fopt and Flimit. All these results indicate that the S. chrysotaenia stock in the Iskenderun Bay is in a situation of overexploitation and also the current exploitation rate should be reduced to below the level of E = 0.50 for the fisheries management.
Lessepsian species are highly influential on Northeast Mediterranean biodiversity and fisheries. Determining the biological features of the lessepsian migrant fish species such as S. chrysotaenia has vital for reflecting the status of established populations and fisheries in the Mediterranean. Although its presence in the region since it has been reported, there has been little or no information about the age growth and mortality parameters of this lessepsian migrant in Iskenderun Bay.
To the best knowledge of the authors, this study presented the first reference on age, growth, and mortality rates for this species; also this study will be very useful for fisheries biologists and managers to know the population structure of this species.
References
Akyuz E (1957) Observations on the Iskenderun red mullet (Mullus barbatus) and its environment. General Fisheries Council for the Mediterranean, Proceedings and Technical Papers 4, FAO, Rome
Allam SM, Faltas SN, Ragheb E (2004) Reproductive biology of Sphyraena species in the Egyptian Mediterranean waters off Alexandria. Egypt J Aquat Res 30(B):255–270
Allam SM, Faltas SN, Ragheb E (2005) Stock assessment of barracuda, genus Sphyraena, along the Egyptian Mediterranean coast. Egypt J Aquat Res 31(2):281–292
Apostolidis C, Stergiou KI (2014) Estimation of growth parameters from published data for several Mediterranean fishes. J Appl Ichthyol 30:189–194. https://doi.org/10.1111/jai.12303
Bagenal TB, Tesch FW (1978) Age and growth. In: Bagenal T (ed) Methods for assessment of fish production in fresh waters. IBP Handbook No. 3, 3rd edn. Blackwell Scientific Publications, Oxford, UK, pp 101–136
Beamish RJ, Fournier DA (1981) A method for comparing the precision of a set of age-determinations. Can J Fish Aquat Sci 38(8):982–983. https://doi.org/10.1139/f81-132
Ben-Tuvia A (1966) Red sea fishes recently found in the Mediterranean. Copeia 2:254–275. https://doi.org/10.2307/1441133
Ben-Tuvia A (1986) Sphyraenidae. In: Whitehead PJP, Buchot ML, Hureau JC, Nielsen J, Tortonese E (eds) Fishes of the North-eastern Atlantic and the Mediterranean. Volume III, Unesco, Paris, pp 1015–1473
Bílge G, Yapici S, Fíliz H, Cerím H (2014) Weight-length relationshions for 103 fish species from the southern Aegean Sea, Turkey. Acta Ichthyol Piscat 44:263–269. https://doi.org/10.3750/AIP2014.44.3.11
Ceyhan, Akyol O, Erdem M (2009) Length-weight relationships of fishes from Gökova Bay, Turkey (Aegean Sea). Turk J Zool 33:69–72. https://doi.org/10.3906/zoo-0802-9
De Sylvia DP, Williams F (1986) Sphyraenidae. In: Smith MM, Heemstra PC (eds) Smiths’ sea fishes. Springer-Verlag, Berlin, pp 721–726
Djabali F, Mehailia A, Koudil M, Brahmi B (1993) Empirical equations for the estimation of natural mortality in Mediterranean teleosts. Naga ICLARM Q 16:35–37
El Ganainy A, Amal A, Abdallah A, Hanan O (2017) Age and growth of two barracuda species Sphyraena chrysotaenia and S. flavicauda (Family: Sphyraenidae) from the Gulf of Suez, Egypt. Egypt J Aquat Res 43:75–81. https://doi.org/10.1016/j.ejar.2016.09.002
Erguden D, Turan C, Gurlek M (2009) Weight-length relationships for 20 Lessepsian fish species caught by bottom trawl on the coast of Iskenderun Bay (NE Mediterranean Sea, Turkey). J Appl Ichthyol 25:133–135. https://doi.org/10.1111/j.1439-0426.2008.01198.x
Froese R, Pauly D (eds) (2021) FishBase. http://www.fishbase.org. World Wide Web Electronic Publication. Accessed 05 May 2021
Fulton TW (1904) The rate of growth of fishes. 22nd Ann Rep Fis. Board Scotl Part III 1904(3):141–240
Gayanilo FC, Sparre P, Pauly D (1994) The FAO ICLARM stock assesment tools (FISAT) user’s guide. In: FAO computerized information series (Fisheries). FAO Press, Rome
Golani D, Ben-Tuvia A (1995) Lessepsian migration and the Mediterranean fisheries of Israel. In: Armantrout NB (ed) Proceedings of the World Fisheries Congress, Theme 1, Conditions of the world’s aquatic habits, Oxford & IBH Pub. Co. Pvt. Ltd, New Delhi, pp 279–289
Golani D (1998) Distribution of Lessepsian migrant fish in the Mediterranean. Italian J Zool 65(Sup 1):95–99. https://doi.org/10.1080/11250009809386801
Golani D, Orsi-Relini L, Massutí E, Quignard JP (2002) CIESM Atlas of exotic species in the Mediterranean. Fishes, vol 1. CIESM Publications, Monaco
Golani D, Öztürk B, Başusta B (2006) Fishes of the Eastern Mediterranean. Publication No. 24, Turkish Marine Research Foundation (TUDAV), Istanbul, Turkey
Gulland JA (1971) The fish resources of the ocean. Fishing news (Books) West Byfleet, Surrey, UK for FAO, Rome
Gulland JA (1983) Fish stock assessment. A manual of basic method. John Wiley & Sons, Chichester
Gücel MU, Sakalli A (2018) Analysing sea surface temperature change in Gulf of Iskenderun from 1982 to 2015. NESciences 3:159–168. https://doi.org/10.28978/nesciences.424665
Gücü AC, Bingel F, Avsar D, Uysal N (1994) Distribution and occurrence of Red Sea fish on the Turkish Mediterranean coast northern Cilician basin. Acta Adriat 34:103–113
Kulbicki M, Moutha G, Thollot P, Wantiez P (1993) Length weight relationship of fish from lagoon of New Caledonia. Naga ICLARM Q 16:26–30
Lanfranco GG (1993) The fish around Malta (Central Mediterranean). Progress Press Co. Ltd, Malta
Le Cren ED (1951) The Length-weight relationship and seasonal cycle in gonad weight and condition in Perch, Perca fluviatilis. J Anim Ecol 20:201-219
Pallaoro A, Dulcic J (2001) First record of the Sphyraena chrysotaenia (Klunzinger, 1884) (Pisces, Sphyraenidae) from the Adriatic Sea. J Fish Biol 59:179–182. https://doi.org/10.1111/j.1095-8649.2001.tb02349.x. )
Pauly D (1980) On the inter-relationships between natural mortality, growth parameters and mean environmental temperature in 175 fish stocks. ICES J Mar Sci 39(2):175–192. https://doi.org/10.1093/icesjms/39.2.175
Pauly D (1983) Length-converted catch curves: a powerful tool for fisheries research in the tropics (Part I). Fishbyte 1:9–13
Pauly D, Munro J (1984) Once more on the comparison of growth in fish and invertebrates. ICLARM Fishbyte 2:1–21
Patterson K (1992) Fisheries for small pelagic species: an empirical approach to management targets. Rev Fish Biol Fish 2:321–338
Ricker WE (1975) Computation and interpretation of biological statistics of fish populations. Bull Fish Res Board Canada 191:1–382
Santic M, Jardas I, Pallaoro A (2002) Age, growth and mortality rate of horse mackerel Trachurus trachurus (L.) living in the eastern Adriatic. Period Biol 104:65–173
Sparre P, Venema SC (1998) Introduction to tropical fish stock assessment. Part 2: Manual. FAO Fisheries Technical Paper. 306/1, Rev, vol 2. FAO, Rome
Spicer IJ (1931) Fisheries. Report of the department of agriculture and forests for the years 1927-30. Printing Office. Russian Building, Jerusalem
Stirn J (1970) Some notes on western trends of Lessepsian migration. Journées Ichthyologiques. CIESM, Monaco, Rome
Taskavak E, Bilecenoglu M (2001) Length-weight relationships for 18 Lessepsian (Red Sea) immigrant fish species from the eastern Mediterranean coast of Turkey. J Mar Biol Assoc UK 81:895–896. https://doi.org/10.1017/S0025315401004805
Tesch FW (1971) Age and growth. In: Ricker WE (ed) Methods for assessment of fish production in freshwaters, 1st edn. Blackwell Scientific Publications, Oxford, UK, pp 98–103
Wadie WF, Riskallah SI (2001) Fisheries for the genus Sphyraena (Perciformes, sphyraenidae) in the southeastern part of the Mediterranean Sea. Pakistan J Mar Sci 10:21–34
Wotton RJ (1990) Ecology of teleost fishes. Fish and Fisheries Series 1. Chapman and Hall, London: England
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice Hall, New Jersey
Zouari-Ktari R, Ennejjar S, Bradai MN, Ghorbel M, Bouain A (2007) Age and growth of the lessepsian migrant Sphyraena chrysotaenia Klunzinger, 1884 from the Gulf of Gabes (Eastern Mediterranean). Rev Fish Sci 15:169–181. https://doi.org/10.1080/10641260701484259
Zouari-Ktari R, Bradai MN, Bouain A (2009) Reproduction and growth of the yellowstripe barracuda Sphyraena chrysotaenia Klunzinger, 1884, Central Mediterranean. Rev Fish Sci 17:485–493. https://doi.org/10.1080/10641260903082471
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
There is no ethical issue concerning this article.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Erguden, D., Ozdemir, O. Age, Growth and Mortality Rate of Yellowstripe Barracuda, Sphyraena Chrysotaenia Klunzinger 1884 Living in the Northeastern Mediterranean. Thalassas 38, 1165–1174 (2022). https://doi.org/10.1007/s41208-022-00431-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41208-022-00431-7