
Abstract
Stable carbon isotopes of dissolved inorganic carbon (δ13CDIC) in the ocean are generally not well understood as they are governed by a complex interplay of biological processes and air–sea exchange. In the Arctic Ocean, δ13CDIC values are prone to change in the near future with rapidly changing climate conditions. This study provides a baseline to assess the δ13CDIC of the Arctic Ocean with a focus on upper to intermediate waters (to ~500 m). Measured δ13CDIC values in the Arctic Ocean range from ~−0.6 to +2.2 ‰. In the Eurasian Basin, the δ13CDIC values lie between ~1 and 1.5 ‰ and exhibit little variation within the upper layers. In the Canada Basin, δ13CDIC values reach 2 ‰ in the surface layer, with lowest values of ~−0.6 ‰ found at ~200 m water depth. At greater depth, δ13CDIC values range from ~1 to 1.5 ‰ within both basins. In the Canada Basin, nutrient levels are higher than in the Eurasian Basin and associated variations in δ13CDIC are clearly related to biological processes. However, low δ13CDIC values in the Canada Basin are also strongly influenced by non-equilibrium air–sea exchange processes. The different δ13CDIC patterns between the Canada Basin and the Eurasian Basin appear to be linked to differences in transport processes within the Arctic Ocean halocline. The upper layers in the Canada basins have direct contributions of waters from the Laptev, East Siberian and Chukchi shelves, which contain elevated fractions of river waters and sea-ice related brines, whereas their counterparts, in the Eurasian Basin, are mostly formed by halocline waters from the Barents and Kara seas. River waters have low δ13CDIC of ~−8 ‰ on average, but in the Arctic basins this signal is mostly lost and δ13CDIC values show only a weak correlation to river water fractions contained in the water mass. No relation between δ13CDIC and sea-ice related brine contribution is apparent.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Stable carbon isotopes of dissolved inorganic carbon (δ13CDIC) in the Arctic Ocean are likely to change in the near future with rapidly changing climate conditions. Climate reconstructions of the recent geological past characterize our climate system under different preconditions and aid to better evaluate the modern climate change and its future projections. For paleoclimate reconstructions, stable isotopes of carbonate shells preserved within the sediments provide a powerful tool (e.g., [46]). While the factors controlling oxygen isotopes (δ18O) in carbonate shells in the Arctic are reasonably well understood (e.g., [51]), this is not the case for stable carbon isotopes (δ13C). Furthermore, we have only a rudimentary understanding of the complex interplay of biological and physical–chemical factors influencing the δ13CDIC that is recorded in the δ13C of carbonate shells (e.g., [27, 43]). With the ongoing climate change and the accompanying changes in Arctic hydrography as well as biological productivity and communities, it is important to assess and further understand the modern distribution of δ13CDIC in the Arctic Ocean. We note that the rapid pace of changes in the Arctic system dictates urgency in the baseline assessment of its components.
One of the most pronounced manifestations of the global climate change is the recent dramatic decline of the Arctic summer sea-ice extent (e.g., [16, 47]). This situation indicates that the Arctic environment is especially sensitive to climate change and that significant further changes may occur in this part of the Earth in the near future as feedback reactions (e.g., [13, 24]). Under present conditions, Arctic sea ice is mainly produced over the broad continental shelves that cover nearly half of the Arctic Ocean area and, to a large extent, are seasonally free of sea ice (e.g., [29]). Areas of seasonal ice distribution over the Arctic shelves can also have high biological productivity (e.g., [25, 41]. Alterations in the Arctic sea-ice cycle have a considerable hydrographic impact as changes in sea-ice extent and production affect the formation of brines and contribution of sea-ice melt to the water column (e.g., [8]) and all these changes may also impact air–sea exchange. Changes in sea-ice seasonality are also likely to have a dramatic effect on Arctic biological production and communities. Some biological aspects of the ongoing Arctic change have already been detected and more are expected to follow (e.g., [5, 21, 45, 50]).
An additional factor to consider for the evaluation of δ13CDIC is the emission of anthropogenic CO2 and the inherent isotopic decline in the atmosphere over the last 150 years (Suess effect). The bias toward lighter δ13CDIC in surface Arctic waters relative to deep waters, which are not affected by the Suess effect due to longer residence times, has been estimated from a comparison of water column and sediment surface data as ~−1 ‰ in the 1990s [7, 28] or somewhat smaller as ~−0.7 ‰ from model estimates (A. Schmittner pers. com., 2014). Here, we present data of δ13CDIC in the water column in the Eurasian and Amerasian basins of the Arctic Ocean from oceanographic summer expeditions taken between 1991 and 2008. In the upper 200 m, the residence time is within about 20 years [49], in the same range as the sampling period of our dataset. The δ13C change in the atmosphere during this period was about 0.5 ‰, with ~−8 and −8.5 ‰ in 1990 and 2010, respectively (based on NOAA ESRL GMD monthly data measured at Barrow, Alaska available at http://www.esrl.noaa.gov/gmd/). The difference in oceanic δ13CDIC is usually smaller compared to the atmosphere and considering the sampling time and upper water residence time may, therefore, be as large as 0.5 ‰ between the datasets and potentially also within the water column. With this potential bias in mind, our data can serve as a baseline for future investigations of changes to be expected with the progression of climate change. Such a baseline is also important for the interpretation of paleo-records from the Arctic region that is widely recognized in its importance for comprehending global paleoclimate processes (e.g., [37]. Beyond a description of δ13CDIC distribution in the Arctic Ocean, we evaluate the connection of the observed features with biological and transport processes within the Arctic Ocean as well as potential δ13CDIC sources such as low-δ13CDIC river water [2, 12, 20]. With the profound impact of climate change on sea ice and brine formation, we specifically address the question, whether sea-ice related processes also affect the distribution of δ13CDIC in the Arctic Ocean.
Data and methods
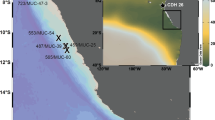
Samples were taken on various expeditions between 1991 and 2008 (Fig. 1) and are from published as well as previously unpublished data presented here as part of this study (Table 1). Precision for δ13CDIC ranges from ±0.02 to ±0.23 ‰ and is partly not known. Two entire datasets were excluded due to (1) little variation and consistently too low values within deep waters compared to all other datasets and (2) little variation and extremely low values within the upper 500 m that show no relation to variations in nutrient levels. We interpret these circumstances as signs for consistent atmospheric contamination probably acquired during inadequate handling of samples, e.g., bubbleling during sampling procedure.

a Index map with location of datasets. The different datasets are indicated by color coding. Single stations are highlighted by additional symbols as used in Figs. 4 and 5. Below b sections of δ13CDIC and δ13Cas of the upper 500 m. For an overview, all station data were projected to the 0°/180° meridian running from Fram Strait to the Chukchi Sea shelf
Calculation of river water and sea-ice meltwater contained within the water column
Marine water masses and freshwater fractions in each water sample are estimated using mass balance calculations based on δ18O and salinity (e.g., [6, 18, 32, 34, 52]). To distinguish also between Atlantic- and Pacific-derived waters, the ratios of nutrients with different nitrate to phosphate ratios (N/P) may be used [26]. It is assumed that each summer sample is a mixture between Atlantic-derived water (fmar), Pacific-derived water (fp), river runoff (fr), and sea-ice meltwater (fSIM). Technically fr refers to meteoric water, but as river runoff dominates in the Arctic Ocean we refer to runoff for simplicity. For further details on calculations and the selection of end members, refer to [9]. As N and/or P data needed for this analysis are not available at a number of stations, we also used a reduced 3-component calculation that assumes marine water to be Atlantic-derived water only and ignores Pacific-derived waters [6]. This approach may be safely applied, and is even more suitable in areas where Pacific-derived waters are absent such as in most parts of the Eurasian Basin. But in areas where Pacific-derived waters are present, such as in the Canada Basin, a 3-component approach considerably overestimates the river water component. Fractions of sea-ice meltwater (SIM), on the other hand, are nearly unaffected by different approaches [9]. Therefore, river water fractions were only calculated with the 4-component approach, while SIM fractions were calculated with the 3-component approach allowing for wider data coverage. All fractions are net values in each sample, and reflect the time-integrated effects (i.e., of SIM) on the sample volume over the residence time of the water. Negative SIM fractions (fSIM) reflect the amount of water removed by sea-ice formation and are proportional to the subsequent addition of brines to the water column. An uncertainty of ±0.3 to 1 %, resulting from analytical errors, associated with δ18O (0.05 and 0.15 ‰ for OM08), salinity and hydrochemical measurements, is estimated for each fraction [9]. An additional systematic error depends on the exact choice of end-member values. When end-member values are varied within the estimated uncertainties, both fractions are shifted by up to ~1 %, but results are always qualitatively conserved, even when tested with extreme end-member variations (see [9]).
Results
The δ13CDIC data compiled from the Arctic Ocean have an overall range between about −0.6 to +2.2 ‰ (Table 2). Deep waters are at ~1 to 1.5 ‰ in δ13CDIC within both basins (Fig. 2). The δ13CDIC within the upper layers of the Eurasian Basin is ~1 ‰ with a range of 0–1.5 ‰, and shows a relatively little variation compared to the Canada Basin. In Fram Strait and the adjacent Greenland Sea, δ13CDIC is predominantly above 1 ‰ and as high as 2.5 ‰ in the upper ~30 m (Fig. 2b). In the Canada Basin, δ13CDIC values range between −0.6 and 2 ‰ in the upper 200–250 m with strong regional differences (Fig. 2b). Within low-salinity surface waters, δ13CDIC reaches up to ~2 ‰ at salinities around 29-30 (Fig. 3). At higher salinities, δ13CDIC values decline to ~0 ‰ within the Upper Halocline Waters (UHW) at ~33.1 salinity and to ~−0.5 ‰ within the Lower Halocline Waters (LHW) between ~34 and 34.5 salinity (Table 3). In the Atlantic layer below ~200 m, δ13CDIC values are consistently between ~1 and 1.5 ‰ throughout the Arctic Ocean.

Profiles of δ13CDIC for the Eurasian and Canada basins of the Arctic Ocean. a Data for the entire water column. b Data for the upper 500 m of the water column. For the Fram Strait and Greenland Sea areas, only data from upper water column are available. Outliers are shown with open symbols

δ13CDIC versus salinity for the upper 500 m of the water column in the Eurasian and Canada basins as well as the Fram Strait and Greenland Sea. Note the inverted scale for salinity
Discussion
The broad Siberian Arctic shelf regions are areas of high biological productivity (e.g., [15, 25, 35, 41]). Therefore, it is not surprising to find high δ13CDIC within the shallow subsurface layer, where light δ13C is incorporated into organic material, and lower δ13CDIC values in the deeper layers of the halocline, where organic material is re-mineralized. This pattern, typical for productivity and decay of organic material, is clearly seen in δ13CDIC profiles from the Canada Basin, where low δ13CDIC values are found between ~150 and 250 m (Fig. 4a) and ~33 to ~34.5 salinity within UHW and LHW, while relatively higher values are seen near the surface (Fig. 5a). Low δ13CDIC values within the halocline layers are also found at the continental slope of the East Siberian Sea and, to a smaller extent, at the Laptev Sea slope (Figs. 4a, 5a). This distribution suggests that the low δ13CDIC values in the Amerasian Basin may originate from the shelves of the Chukchi, East Siberian, and Laptev seas. We hypothesize that low δ13CDIC signals from the Siberian shelves may propagate with the boundary current that transports halocline waters from west to east along the Eurasian continental margin [1, 11, 31]. Surprisingly, the δ13CDIC pattern related to organic matter cycling appears to be nearly absent in the Eurasian Basin, which poses the question, whether local productivity and decay of low-δ13C organic material are indeed so different between the Arctic basins? Alternatively, this dissimilarity may be controlled by different circulation patterns in the Canadian and Eurasian basins.

Depth profiles of δ13CDIC and δ13CDIC-as for the upper 500 m of the water column. The single light value at 175 m in the Canada Basin (open red dot) is considered to be an outlier. Detailed station locations are shown in Fig. 1 with corresponding symbols

Relation of δ13CDIC and δ13CDIC-as versus salinity. Note the inverted scale for salinity. The single light value at 33 salinity in the Canada Basin (open red dot) is considered to be an outlier. Detailed station locations are shown in Fig. 1 with corresponding symbols
In the following discussion, we will investigate what factors cause the low δ13CDIC values in the Canada Basin and the difference in δ13CDIC patterns between the Eurasian and the Canada basins. We will evaluate the impact to biological productivity on δ13CDIC within the two basins and analyze the non-biological factors. We will also review the Arctic Ocean halocline circulation in relation to δ13CDIC and analyze the influence of low δ13CDIC in river water as well as the potential effect of sea-ice melt and formation.
Biotic and non-biotic influences on δ13CDIC in the Arctic Ocean
To distinguish the influence of biological and non-biotic factors, δ13CDIC data can be adjusted by their nutrient levels. Phosphate- and nitrate-based adjustments have been applied by [27] and by [33], respectively. Adjusted δ13CDIC values are generally referred to as δ13Cas as all non-biotic factors are assumed to be caused by air–sea exchange processes. Calculation of δ13Cas may be based on:
-
1.
δ13Cas = δ13CDIC −(2.7 − 1.1 * PO4) after [27]
-
2.
δ13Cas = δ13CDIC −(2.2 − 0.06 * NO3) after [33]
Both these globally calibrated equations predict the δ13CDIC due to the biological activity from the measured nutrient concentrations and subtract the calculated value from the measured δ13CDIC. For the relation between δ13CDIC and nutrients, ideal biological processes with constant fractionation and constant Redfield nutrient ratios are assumed based on global or regional averages. Clearly, both corrections are not adjusted to local Arctic conditions, where, e.g., Pacific- and Atlantic-derived waters have different preformed nutrient levels. In our data from the Canada Basin and adjacent shelves, high nutrient levels are mostly related to low δ13CDIC from re-mineralized organic material, while low nutrient levels are related to high δ13CDIC reflecting nutrients fixed in organic material (Fig. 6a and b). In contrast, in the Eurasian Basin, δ13CDIC remains nearly constant, while the nitrate range is high (Fig. 6b) as biological fixation is probably limited by low phosphate levels (Fig. 6a). Atlantic-derived waters have relatively low P levels (Fig. 6c), which may limit biological production in the Eurasian Basin (Fig. 6a), while N levels remain relatively high (Fig. 6b; see blue diamond for Eurasian Basin waters). Pacific-derived waters have on average higher P (Fig. 6c), but both N and P levels are low for δ13CDIC values that are high, which reflect the fixation of carbon from the DIC pool in low-δ13C organic matter (Fig. 6a, b; see green triangles for Chukchi Sea and part of Canada Basin data shown by red squares). Due to the phosphate limitation in the Eurasian part of the Arctic Ocean, we have adjusted the δ13CDIC data for biological processes applying the phosphate-based correction [27], an approach that also allows us to analyze more of the δ13CDIC data due to the greater availability of phosphate measurements in our dataset. This result differs from the observations off Oregon, were [33] observed evidence for N-limitation. We will use the term δ13Cas for the adjusted δ13CDIC data, although this notation implies that all non-biotic factors are related to air–sea exchange, which is not strictly correct as river water and sea-ice processes may also have an impact.

Relation between δ13CDIC and a dissolved phosphate (P) and b nitrates (N). Indicated are lines of constant air–sea exchange according to equations given within text for phosphate- and nitrate-based adjustments. Also shown are the relation between c N and P and d the ratios NO/PO versus N/P. For further explanation, see text
In the southern Eurasian Basin and in Fram Strait, δ13Cas is relatively constant with values close to −1 ‰ (Figs. 4b, 5b) similar to North Atlantic values [27]. But in the Canada Basin and at the East Siberian and Chukchi slopes, δ13Cas values show a considerable variation and internal structure within the water column (Figs. 4b, 5b). In the upper 200 m in the Canada Basin and at 100-150 m at the ESS slope, δ13Cas is relatively high, between −0.5 and 0.5 ‰ (see yellow, orange and red profiles in Fig. 4b). Under equilibrium conditions, invading atmospheric CO2 at cold temperatures increases the δ13CDIC and thus leads to relatively high δ13Cas values [30]. But because the equilibration time between the atmosphere and even a shallow water column is relatively long, there are immediate impacts that lower or increase δ13CDIC when CO2 enters or leaves the DIC pool, respectively [27] and thus act initially into opposite direction than equilibrium processes. While chemical equilibrium occurs on timescales of about 1 year, isotopic equilibration takes on the order of 10 years or more for a shallow water column of ~50 m (see model calculation in [27]). As the surface residence time in the Arctic is well below 10 years and considerably shorter on the Arctic shelves [42], it may be concluded that air–sea processes are far from equilibrium within the entire Arctic Ocean and shelf regions and that invading atmospheric CO2 will lower δ13CDIC and δ13Cas values. Nevertheless, part of the δ13Cas variability may also be connected to variations in surface residence time and thus variations in incomplete isotopic equilibration. Therefore, the observed relatively high δ13Cas values may indicate that the invasion of isotopically light atmospheric CO2 in the upper Canada Basin waters is weaker than in the North Atlantic and Eurasian Basin, which likely arises from the inhibition of air–sea gas equilibration due to the more complete ice coverage in the Canada basin. Considerably, lighter δ13Cas values of ~−1.5 ‰ are found at ~150 m water depth or bottom depth at the continental slope of the ESS and on the Chukchi shelf (Fig. 4b). These light δ13Cas values potentially indicate enhanced invasion of light atmospheric CO2, likely associated with sinking of near surface water during winter cooling or brine rejection during sea-ice formation.
As an alternative, part of the observed structure in δ13Cas within the water column might be caused by regional deviations in rates of biological processes relative to the global δ13C–P relationship. For example, δ13C fractionation may be higher than the assumed global averages in high productivity areas at low temperatures [27]. Therefore, the δ13Cas values may be locally underestimated or overestimated, by the global relationship because the organic matter is formed or remineralized, respectively, with regionally varying biological processes. If that were the case, the relatively high δ13Cas in the upper 200 m of the Canada Basin and at 100–150 m at the ESS slope in comparison to the North Atlantic values may be caused by an underestimation of 13C fractionation during the production of organic matter or its respiration. Accordingly, low δ13Cas may partly result from an underestimation due to remineralization of organic matter with lower than average δ13C composition. A comparison of production between different regions is difficult but non-Redfield productivity may be estimated [17]. Away from the sea surface, the quasi-conservative tracers NO and PO (NO = 9*[NO3 −]+O2 and PO = 135*[PO4 −]+O2; [14]) account for the approximate stoichiometric ratios of nutrient and oxygen during consumption and production or regeneration. A plot of NO/PO versus N/P can provide an indication of differences in nutrient consumption or regeneration that deviates from Redfield ratios [17]. In the Arctic, variations in non-Redfield productivity can be seen between surface waters and halocline layers in comparison to deep waters within our datasets (Fig. 6d). No relationship between either the N/P ratio or the NO/PO preformed nutrient ratio to variations in δ13CDIC values is apparent (not shown). While relatively low NO/PO ratios within Pacific-derived waters indicate an enlarged N loss relative to P with slower N recycling relative to P [17], surface waters from the Eurasian Basin and all other locations show the opposite effect with high NO/PO relative to N/P ratios (Fig. 6d). Therefore, in addition to differences in fractionation due to different productivity also different non-Redfield behavior may influence our estimates of δ13Cas. With this caveats in mind, the high δ13Cas in the Canada Basin and low δ13Cas at the continental slope of the ESS and on the Chukchi shelf do suggest a possibility of reduced and enhanced air–sea exchange, respectively.
While a large part of the East Siberian Sea is overall an area of CO2 outgassing due to an excess of pCO2 derived from the degradation of organic carbon [2], the eastern part of the East Siberian Sea and the Chukchi Sea is a sink of CO2, especially at the end of the ice-free season [4, 36, 44]. This further confirms the notion that low δ13Cas values in these areas likely reflect the invasion of isotopically light atmospheric CO2 under non-equilibrium conditions on the shelf areas.
The distribution of elevated δ13Cas values of about 0.5 ‰ at ~32.5 to 34 salinity near the East Siberian Slope, on the Chukchi shelf and in the Canada Basin (Fig. 5) supports our hypothesis that high and low δ13Cas signals may originate from common water masses transported along the Eurasian continental slope and into the Canada Basin.
Oceanographic differences within the Arctic circulation with respect to δ13CDIC
Shelf waters in the Eurasian Basin are thought to recirculate in the Gakkel Gyre [23]. This concept is supported by a Ra/Th estimate of shelf water residence ages of at least 8 years, which is much older than ~3 years for waters in the Transpolar Drift over the Lomonosov Ridge [39]. This considerably longer residence time of shelf waters may be a factor for the missing variability in δ13CDIC in the Eurasian Basin due to a stronger degradation of any shelf-sourced biological signal in this region.
Halocline waters in the Eurasian Basin are mainly LHW, with a relatively high salinity of ~34.5, which are formed by the modification of Atlantic waters over the Barents and northern Kara seas [38]. The southern part of the Eurasian Basin is mainly influenced by Atlantic-derived waters, with only minor contributions from rivers. Waters from the large Siberian rivers, including the Ob and the Yenisey rivers, are carried eastward in the southern Kara Sea, but cross the shelf break only in the eastern Laptev Sea and the East Siberian Sea [9, 11]. Shelf waters with a considerable contribution from these rivers as well as sea-ice related brines then spread further eastward within the boundary current [1, 11, 31] into the Canada Basin or along the Lomonosov Ridge in the Transpolar Drift [10]. Due to these low-salinity contributions, halocline waters in the Canada Basin are much fresher than in the Eurasian Basin and UHW is found at ~33.1 and is also strongly influenced by Pacific water with high silicate and generally higher nutrient levels (e.g., [3, 10]).
According to the upper ocean circulation, the low δ13CDIC and high δ13Cas values in the Canada Basin may not be of local origin, but may fully or partly originate from the continental margin in the upper water column. Such a transport of light δ13CDIC signals is consistent with light δ13CDIC values and high δ13Cas in the bottom waters of the Chukchi shelf (Fig. 4a, black and yellow crosses) and at the upper slope of the East Siberian Sea at ~100–200 m water depth (Fig. 4a, yellow squares and brown triangles). Stations at the central Laptev Sea slope show a similar pattern, although with a smaller δ13CDIC depletion of 0.2 ‰ within the water column (Fig. 4a, blue crosses). Still the depletion is clearly visible relative to δ13CDIC values in the Eurasian Basin, which are higher on average by 0.5 ‰ (Fig. 4a, green bars). Due to the Suess effect, an anthropogenic-induced bias of up to ~0.5 ‰ between the upper layers and the observed depletion at ~100–200 m water depth may be possible. Therefore, the observed depletion within the Eurasian Basin stations might be solely due to the anthropogenic effect but cannot explain the depletions at the slope of the East Siberian Sea. Overall, this distribution indicates an accumulation of light δ13CDIC and high δ13Cas along the Siberian continental slope at ~100–200 m water depth (Fig. 4) and ~33 to 33.5 salinity (Fig. 5), possibly in connection to Pacific-derived waters and the export of low-salinity shelf water in the eastern part of the Laptev Sea and the East Siberian Sea [3, 10].
The pattern with high δ13CDIC at the surface and low δ13CDIC at an intermediate depth found in the Canada Basin is typical for biological processes. However, the δ13Cas adjusted for biological effects still shows a similar structure (Fig. 4b) with relatively high δ13Cas values at about 32.5 to 34 salinity (Fig. 5b). This supports the assumption that both light δ13CDIC and high δ13Cas values within the Canada Basin are transported in the Arctic Ocean halocline within the shelf-derived, eastward boundary current and may originate on the Laptev, East Siberian and Chukchi shelves and are thus partly influenced by Pacific-derived waters as well as Siberian shelf waters.
In contrast to the Canada Basin, δ13CDIC and δ13Cas in the Eurasian Basin remain relatively constant. The stratification between the surface, LHW, and Atlantic waters in the southern Eurasian Basin is relatively weak compared to waters flowing off the Laptev, East Siberian and Chukchi shelves into the Canada Basin. We can speculate that CO2 invasion is much smaller for the Eurasian Basin and adjacent shelve regions or that the structure due to CO2 invasion is simply lost in the Eurasian Basin due to the reduced stratification on the Barents Sea and northern Kara Sea shelves in comparison to the shelf regions further eastwards.
Influence of river δ13CDIC and brine enriched waters
Siberian rivers have low δ13CDIC values of about −8.5 ‰ in the Ob and Yenisey rivers [20] and about −8 ‰ in the Lena River [2, 12, 20]. Therefore, all water masses containing river water may potentially be influenced by low δ13CDIC from this source. However, a comparison between δ13CDIC and the fraction of river water (fr), derived from the 4-component δ18O and salinity mass balance (see methods), shows little or slight positive correlation (Fig. 7). Marine waters have higher DIC concentrations compared to meteoric waters, e.g., in the Laptev Sea and, therefore, mixing and potential dilution of δ13CDIC signals are not linear [19]. As river runoff is an important nutrient source for biological processes in the otherwise nutrient-depleted Arctic, the δ13CDIC values must be adjusted for biotic factors for a comparison with fr. This comparison shows considerable scatter, but the individual datasets show slight trends toward lower δ13Cas with increasing river influence (Fig. 7b). Shifts between and within datasets may be caused by strong variations in N/P consumption on the Siberian shelves that are reflected in strong variations in N/P ratios [11]. The applied adjustment for the calculation of δ13Cas may, therefore, only roughly remove the signal of biology in δ13CDIC. However, the slight trend of δ13Cas decrease with increasing river influence seen within some dataset suggests a minor but genuine remnant signal of low riverine δ13CDIC that is otherwise largely lost due to dilution.

Relation of δ13CDIC and δ13CDIC-as versus river fraction (fr; calculated with 4-component approach)
In addition to runoff, the shallow halocline carries a considerable fraction of shelf waters with brines introduced by sea-ice formation [9]. The signal from melting of sea-ice may also be transported in the upper halocline, but is usually found at the very surface in the upper 10–20 m of the water column only. We compare δ13CDIC and δ13Cas with the fraction of sea-ice meltwater (fSIM), where positive and negative fSIM values correspond to the actual sea-ice melt and sea-ice related brines, respectively (see methods) (Fig. 8). Overall, a weak trend toward higher δ13CDIC with increasing meltwater contribution is seen in some datasets. This pattern may be related to the notion that sea-ice meltwater adds nutrients to the water column and, thereby, enables biological production and also adds to the DIC pool [40]. However, there is no indication of an effect of sea-ice formation and thus sea-ice related brines on δ13Cas. High fractions of sea-ice formation are seen in the halocline layers of the Eurasian Basin (Fig. 8, see Oden 91 data with negative fSIM of up to ~−8 %) and the Canada Basin (Fig. 8, see ESS-08 data with fSIM of up to ~−3 %). But while the fractions of sea-ice formation (neg fSIM values) vary strongly, δ13CDIC as well as δ13Cas shows no trend.

Relation of δ13CDIC and δ13CDIC-as versus sea-ice meltwater fraction (fSIM; calculated with 3-component approach)
Summary and future directions
In the Canada Basin, high δ13CDIC values at the surface appear to reflect the biological productivity signal and removal of light δ13C from the DIC pool, whereas low δ13CDIC values deeper in the halocline (UHW and LHW) indicate the recycling of organic material. However, the evidence from δ13CDIC adjusted for global-scale, biological processes, δ13Cas, suggests a more complex picture, where δ13CDIC values may be largely transported to the Canada Basin from the adjacent shelf areas. Further, low δ13CDIC in the UHW and LHW at around 33.1 and 34.5 salinity, respectively, may not only be caused by the decay of organic material, but also by variable air–sea exchange processes under non-equilibrium conditions occurring primarily on the shelves under seasonal sea-ice conditions. Strong outgassing of CO2 and, thereby, enrichment of δ13CDIC has been demonstrated for much of the East Siberian Sea [2], and the resulting high δ13CDIC signal is transported within the halocline into the Canada Basin. However, the invasion of light atmospheric CO2 resulting in light δ13Cas values is also traceable along the continental slope and in the Canada Basin at higher salinities.
In the Eurasian Basin, δ13CDIC values show little variation. While biological productivity over the Barents Sea and northern Kara seas can be high, the signals of production and remineralization with high and low δ13CDIC, respectively, are apparently not transported into the Eurasian Basin. This may be connected to a weaker stratification in the Eurasian Basin and sufficient degradation of any shelf-sourced biological signal potentially present in this region as shelf waters recirculating in the Eurasian Basin have a relatively long residence time of at least 8 years [39].
In addition to air–sea interaction, river water, sea-ice meltwater, and sea-ice related brine waters are potentially important non-biotic effects on δ13CDIC in the Arctic Ocean. But while the extremely low-δ13CDIC river runoff signal is very prominent on the Siberian shelves [20], it does not significantly influence the overall δ13CDIC distribution within the Arctic basins and is only weakly visible—if at all—within the Canada basin. We also find no evidence for any direct influence of sea-ice production on δ13CDIC. While transport processes within the halocline clearly play an important role in the distribution of δ13CDIC within the Arctic Ocean, the fractions of river and sea-ice related waters, also transported within the halocline, show no significant effects on the δ13CDIC.
Our data are the first compilation of δ13CDIC from the Arctic Ocean and are meant to serve as an initial baseline for this region. As Arctic sea ice and related environments are especially sensitive to a rapidly changing climate, Arctic Ocean δ13CDIC values are prone to change in the near future. While our data show that sea-ice melt and formation processes have no strong direct influence on δ13CDIC, our data nevertheless suggest that increasing river runoff and changing sea-ice conditions in the Arctic Ocean and on the Arctic shelves may still significantly impact the δ13CDIC due to the indirect feedback mechanisms associated with changing biological activity and air–sea exchange.
When applied to paleo-climate studies, the presented δ13CDIC data need to be further evaluated in the context of the surface ocean Suess effect. The sampling period of nearly 20 years adds an increased uncertainty due to a decrease of about −0.5 ‰ in atmospheric δ13C values across the sampling interval. Within these limitations, our data may serve as a first useful δ13CDIC baseline due to a total lack of further data from the Arctic Ocean water column. For example, high δ13C in planktic foraminifers from surface sediments in the Canada Basin [51] may be explained by the import of biologically mediated δ13CDIC from the shelves, while pronounced low surface-sediment planktic δ13C along the shelf break of the Chuckchi and Beaufort seas [51] may be related to low δ13CDIC modified by air–sea exchange. Despite remaining substantial uncertainties with the application of surface-sediment stable isotope data to a more distant past, modern δ13CDIC and δ18O data from the water column constitute an essential prerequisite for paleoclimatic reconstructions.
References
Aksenov Y, Ivanov VV, Nurser AJG, Bacon S, Polyakov IV, Coward AC, Naveira-Garabato AC, Beszczynska-Moeller A (2011) The arctic circumpolar boundary current. J Geophys Res Oceans 116(C9):C09017. doi:10.1029/2010JC006637
Alling V, Porcelli D, Mörth C-M, Anderson LG, Sanchez-Garcia L, Gustafsson Ö, Andersson PS, Humborg C (2012) Degradation of terrestrial organic carbon, primary production and out-gassing of CO2 in the Laptev and East Siberian Seas as inferred from δ13C values of DIC. Geochim Cosmochim Acta 95:143–159. doi:10.1016/j.gca.2012.07.028
Anderson LG, Andersson PS, Björk G, Peter Jones E, Jutterström S, Wåhlström I (2013) Source and formation of the upper halocline of the Arctic Ocean. J Geophys Res Oceans 118(1):410–421. doi:10.1029/2012JC008291
Anderson LG, Jutterström S, Hjalmarsson S, Wåhlström I, Semiletov IP (2009) Out-gassing of CO2 from Siberian Shelf seas by terrestrial organic matter decomposition. Geophys Res Lett 36(20):L20601. doi:10.1029/2009GL040046
Arrigo KR, van Dijken GL (2011) Secular trends in Arctic Ocean net primary production. J Geophys Res Oceans 116(C9):C09011. doi:10.1029/2011JC007151
Bauch D, Schlosser P, Fairbanks RF (1995) Freshwater balance and the sources of deep and bottom waters in the Arctic Ocean inferred from the distribution of H 182 O. Prog Oceanogr 35:53–80
Bauch D, Carstens J, Wefer G, Thiede J (2000) The imprint of anthropogenic CO2 in the Arctic Ocean: evidence from planktonic δ13C data from watercolumn and sediment surfaces. Deep Sea Res II 47:1791–1808
Bauch D, Hölemann J, Willmes S, Gröger M, Novikhin A, Nikulina A, Kassens H, Timokhov L (2010) Changes in distribution of brine waters on the Laptev Sea shelf in 2007. J Geophys Res 115:C11008. doi:10.1029/2010JC006249
Bauch D, Gröger M, Dmitrenko I, Hölemann J, Kirillov S, Mackensen A, Taldenkova E, Andersen N (2011) Atmospheric controlled freshwater water release at the Laptev Sea Continental margin. Polar Res 30:5858. doi:10.3402/polar.v30i0.5858
Bauch D, Rutgers van der Loeff M, Andersen D, Torres-Valdes S, Bakker K, Abrahamsen EP (2011) Origin of freshwater and polynya water in the Arctic Ocean halocline in summer 2007. Prog Oceanogr 91:482–495. doi:10.1016/j.pocean.2011.07.017
Bauch D, Torres-Valdes S, Polyakov I, Novikhin A, Dmitrenko I, McKay J, Mix A (2014) Halocline water modification and along-slope advection at the Laptev Sea continental margin. Ocean Sci. 10(1):141–154. doi:10.5194/os-10-141-2014
Bauch HA, Erlenkeuser H, Bauch D, Mueller-Lupp T, Taldenkova E (2004) Stable oxygen and carbon isotopes in modern benthic foraminifera from the Laptev Sea shelf: implications for reconstructing proglacial and profluvial environments in the Arctic. Mar Micropaleontol 51(3–4):285–300. doi:10.1016/j.marmicro.2004.01.002
Bekryaev RV, Polyakov IV, Alexeev VA (2010) Role of polar amplification in long-term surface air temperature variations and modern Arctic warming. J. Climate 23(14):3888–3906. doi:10.1175/2010JCLI3297.1
Broecker WS (1974) “NO,” A conservative water mass tracer. Earth Planet Sci Lett 23:100–107
Carmack E, Wassmann P (2006) Food webs and physical–biological coupling on pan-Arctic shelves: unifying concepts and comprehensive perspectives. Prog Oceanogr 71(2–4):446–477. doi:10.1016/j.pocean.2006.10.004
Comiso JC, Parkinson CL, Gersten R, Stock L (2008) Accelerated decline in the Arctic sea ice cover. Geophys Res Lett 35:L01703. doi:10.1029/2007GL031972
Cooper LW, Cota GF, Pomeroy LR, Grebmeier JM, Whitledge TE (1999) Modification of NO, PO, and NO/PO during Flow across the Bering and Chukchi Shelves: Implications for Use as Arctic Water Mass Tracers. J Geophys Res 104:7827–7836
Ekwurzel B, Schlosser P, Mortlock R, Fairbanks R (2001) River runoff, sea ice meltwater, and Pacific water distribution and mean residence times in the Arctic Ocean. J Geophys Res 106(C5):9075–9092
Erlenkeuser H, TRANSDRIFT II Shipbord Scientific Party (1995) Stable carbon isotope ratios in the waters of the Laptev Sea/Sept. 94, Berichte zur Polarforschung, vol 176. AWI, Bremerhaven, Germany, pp 170–177
Erlenkeuser H, Cordt HH, Simstich J, Bauch D, Spielhagen RF (2003) DIC stable carbon isotope pattern in the surface waters of the southern Kara Sea, September 2000, in Siberian river run-off in the Kara Sea; characterisation, quantification, variability and environmental significance. In: Stein R, Fahl K, Fuetterer D, Galimov E, Stepanets O (eds) Proceedings in Marine Science. Elsevier, Amsterdam, pp 91–110
Grebmeier JM (2012) Shifting Patterns of Life in the Pacific Arctic and Sub-Arctic Seas, edited by C. A. Carlson and S. J. Giovannoni. Annu Rev Mar Sci 4:63–78. doi:10.1146/annurev-marine-120710-100926
Griffith DR, McNichol AP, Xu L, McLaughlin FA, Macdonald RW, Brown KA, Eglinton TI, Herndl G (2012) Carbon dynamics in the western Arctic Ocean: insights from full-depth carbon isotope profiles of DIC, DOC, and POC. Biogeosciences 9(3)
Gordienko PA, Laktionov AF (1969) Circulation and physics of the Arctic Basin waters. Ann Int Geophys Year Oceanogr 46:94–112
Holland MM, Bitz CM (2003) Polar amplification of climate change in coupled models. Clim Dyn 21(3–4):221–232. doi:10.1007/s00382-003-0332-6
Hunt GL, Blanchard AL, Boveng P, Dalpadado P, Drinkwater KF, Eisner L, Hopcroft RR, Kovacs KM, Norcross BL, Renaud P, Reigstad M, Renner M, Skjoldal HR, Whitehouse A, Woodgate RA (2013) The Barents and Chukchi Seas: comparison of two Arctic shelf ecosystems. J Mar Syst 109–110:43–68. doi:10.1016/j.jmarsys.2012.08.003
Jones E, Anderson L, Swift J (1998) Distribution of Atlantic and Pacific water in the upper Arctic Ocean: implications for circulation. Geophys Res Lett 25(6):765–768
Lynch-Stieglitz J, Stocker TF, Broecker W, Fairbanks RG (1995) The influence of air–sea exchange on the isotopic composition of oceanic carbon: observation and modeling. Global Biogeochem Cycles 9(4):653–665
Mackensen A (2013) High epibenthic foraminiferal δ13C in the Recent deep Arctic Ocean: implications for ventilation and brine release during stadials. Paleoceanography 28(3):574–584. doi:10.1002/palo.20058
Martin S, Cavalieri DJ (1989) Contributions of the Siberian Shelf polynyas to the Arctic Ocean intermediate and deep water. J Geophys Res 94(C9):12725–12738
Mook WG, Bommerson JC, Staverman WH (1974) Carbon isotope fractionation between dissolved bicarbonate and gaseous carbon dioxide. Earth Planatary Sci Lett 22:169–176
Newton R, Schlosser P, Martinson DG, Maslowski W (2008) Freshwater distribution in the Arctic Ocean: simulation with a high resolution model and model-data comparison. J Geophys Res 113(C05024). doi:10.1029/2007JC004111
Newton R, Schlosser P, Mortlock R, Swift J, MacDonald R (2013) Canadian Basin freshwater sources and changes: results from the 2005 Arctic Ocean Section. J Geophy Res Oceans 118(4):2133–2154. doi:10.1002/jgrc.20101
Ortiz JD, Mix AC, Wheeler PA, Key RM (2000) Anthropogenic CO2 invasion into the northeast Pacific based on concurrent δ13CDIC and nutrient profiles from the California Current. Global Biogeochem Cycles 14(3):917–929. doi:10.1029/1999GB001155
Östlund H, Hut G (1984) Arctic Ocean water mass balance from isotope data. J Geophys Res 89(C4):6373–6381
Pabi S, van Dijken GL, Arrigo KR (2008) Primary production in the Arctic Ocean, 1998–2006. J Geophys Res Oceans 113(C8):C08005. doi:10.1029/2007JC004578
Pipko I, Semiletov I, Tishchenko PY, Pugach S, Christensen J (2002) Carbonate chemistry dynamics in Bering Strait and the Chukchi Sea. Prog Oceanogr 55(1–2):77–94. doi:10.1016/S0079-6611(02)00071-X
Polyak L, Jakobsson M (2011) Quaternary sedimentation in the Arctic Ocean: recent advances and further challenges. Oceanography 24(3):52–64
Rudels B, Anderson LG, Jones EP (1996) Formation and evolution of the surface mixed layer and halocline of the Arctic Ocean. J Geophys Res Oceans 101(C4):8807–8821. doi:10.1029/96JC00143
Rutgers van der Loeff M, Cai P, Stimac I, Bauch D, Hanfland C, Roeske T, Moran SB (2012) Shelf-basin exchange times of Arctic surface waters estimated from 228Th/228Ra disequilibrium. J Geophys Res117(C03024). doi:10.1029/2011JC007478
Rysgaard S, Glud RN, Sejr MK, Bendtsen J, Christensen PB (2007) Inorganic carbon transport during sea ice growth and decay: a carbon pump in polar seas. J Geophys Res Oceans 112(C3):C03016. doi:10.1029/2006JC003572
Sambrotto RN, Goering JJ, McRoy CP (1984) Large yearly production of Phytoplankton in the Western Bering Strait. Science 225(4667):1147–1150. doi:10.1126/science.225.4667.1147
Schlosser P, Bauch D, Bönisch G, Fairbanks RF (1994) Arctic river-runoff: mean residence time on the shelves and in the halocline. Deep-Sea Res I 41(7):1053–1068
Schmittner A, Gruber N, Mix AC, Key RM, Tagliabue A, Westberry TK (2013) Biology and air–sea gas exchange controls on the distribution of carbon isotope ratios (δ13C) in the ocean. Biogeosciences 10(9):5793–5816
Semiletov IP, Pipko II, Repina I, Shakhova NE (2007) Carbonate chemistry dynamics and carbon dioxide fluxes across the atmosphere–ice–water interfaces in the Arctic Ocean: pacific sector of the Arctic. J Mar Syst 66(1–4):204–226. doi:10.1016/j.jmarsys.2006.05.012
Slagstad D, Ellingsen IH, Wassmann P (2011) Evaluating primary and secondary production in an Arctic Ocean void of summer sea ice: an experimental simulation approach. Prog Oceanogr 90(1–4):117–131. doi:10.1016/j.pocean.2011.02.009
Spielhagen RF, Erlenkeuser H (1994) Stable oxygen and carbon isotopes in planktic foraminifers from Arctic ocean surface sediments: reflection of the low salinity surface water layer. Mar Geol 119:227–250
Stroeve JC, Kattsov V, Barrett A, Serreze M, Pavlova T, Holland M, Meier WN (2012) Trends in Arctic sea ice extent from CMIP5, CMIP3 and observations. Geophys Res Lett 39 (16):L16502. doi:10.1029/2012GL052676
Volkmann R, Mensch M (2001) Stable isotope composition (δ18O, δ13C) of living planktic foraminifers in the outer Laptev Sea and the Fram Strait. Mar Micropaleontol 42:163–188
Wallace DWR, Schlosser P, Krysell M, Bönisch G (1992) Halocarbon ratio and tritium/3He dating of water masses in the Nansen Basin, Arctic Ocean, Deep Sea Research Part A. Oceanographic Research Papers 39(2, Part 1):S435–S458. doi:10.1016/S0198-0149(06)80015-4
Wassmann P, Reigstad M (2011) Future Arctic Ocean seasonal ice zones and implications for pelagic-benthic coupling. Oceanography 24:220–231. doi:10.5670/oceanog.2011.74
Xiao W, Wang R, Polyak L, Astakhov A, Cheng X (2014) Stable oxygen and carbon isotopes in planktonic foraminifera Neogloboquadrina pachyderma in the Arctic Ocean: an overview of published and new surface-sediment data. Mar Geol 352:397–408
Yamamoto-Kawai M, McLaughlin FA, Carmack EC, Nishino S, Shimada K (2008) Freshwater budget of the Canada Basin, Arctic Ocean, from salinity, δ18O, and nutrients. J Geophys Res 113(C01007). doi:10.1029/2006JC003858
Acknowledgments
We thank the captains and crews of the various ship-based expeditions and all other colleagues who assisted in sample collection and stable isotope measurements. We are thankful to A. Schmittner for commenting on an earlier version of the manuscript. Comments by two reviewers are gratefully acknowledged and helped to improve the study. The work was supported by the German Research Foundation DFG grant BA1638-1 to DB, and from the National Science Foundation NSF to LP and JDO (PLR-0520365).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bauch, D., Polyak, L. & Ortiz, J.D. A baseline for the vertical distribution of the stable carbon isotopes of dissolved inorganic carbon (δ13CDIC) in the Arctic Ocean. Arktos 1, 15 (2015). https://doi.org/10.1007/s41063-015-0001-0
Published:
DOI: https://doi.org/10.1007/s41063-015-0001-0