
Abstract
Drought stress is a major concern for current agriculture as it limits plant growth and yield. An experiment was conducted with a drought susceptible rice (Oryza sativa L. cv. BRRI dhan29) to explore the potential functions and possible mechanisms of exogenous glutathione (GSH)-induced drought stress tolerance. Fifteen-day-old seedlings, GSH-pretreated or non-pretreated (0.2 mM GSH for 72 h), were subjected to PEG-induced (15% polyethylene glycol 6000) drought stress for 4 days. Rice plants exposed to drought stress displayed reduced growth, which was correlated with reduced chlorophyll content, water balance, antioxidant enzymes activities and mineral contents (K+, Ca2+ and Mg2+), as evidenced by principle component analysis (PCA) and heatmap clustering. The PCA also revealed that hydrogen peroxide (H2O2) and malondialdehyde (MDA) accumulations were strongly linked with the declined growth and development of rice plants under drought stress. Importantly, pretreatment with GSH improved the growth responses of rice plants to drought stress. GSH strengthened numerous physio-biochemical processes which were coupled with increased uptake of K+, Ca2+ and Mg2+, enhanced accumulation of soluble sugars, higher activities of enzymatic and non-enzymatic antioxidants. Moreover, GSH-pretreated seedlings under drought stress conditions had lower levels of O·−2 , H2O2 and MDA which indicates successful adaptation of rice seedlings to drought stress. The results of PCA strongly supported that GSH conferred drought tolerance mostly by enhancing antioxidant activities and mineral homeostasis. Collectively, our findings provide an important prospect for use of GSH in modulating drought tolerance in rice plants. Breeding strategies should therefore be undertaken to increase the GSH content of rice plants as a means to increase drought tolerance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Drought stress is a pivotal constraints to global agricultural production, especially in warm, arid and semi-arid regions (Wang et al. 2016; Barik et al. 2019). By 2080, most of the world’s population will directly or indirectly be affected by the reduction in crop yields caused by drought stress (Schmidhuber and Tubiello 2007). In recent years the incidence and harshness of drought have increased, most likely due to the global heating (IPCC 2014; Liu et al. 2018). For instance, in Bangladesh, the temperature has increased by up to 2.48 °C in the last four decades, and it will continue to rise in the future (Rahman and Lateh 2017). Moreover, annual rainfall has been estimated to reduce by 153 mm between 2010 and 2020 (Rahman and Lateh 2017). Thus, drought stress is considered as a serious threat for agriculture in Bangladesh, especially in the northwest part of the country (Habiba et al. 2012).
Water deficiency limits crop productivity by affecting plant growth and can also deteriorate the nutritional value of crops. Primarily, drought instigates osmotic imbalance in plants which interrupts many physiological aspects, such as enzymatic and non-enzymatic processes, stomatal conductance, membrane electron transport chain, photosynthesis and the production and scavenging of reactive oxygen species (ROS) (Lisar et al. 2012; Osakabe et al. 2014). ROS, a group of molecules that include the superoxide radical (O·−2 ), singlet oxygen (1O2), hydrogen peroxide (H2O2) and hydroxyl radical (OH·), are produced at a low level in different cell organelles during normal physiological state, whereas under conditions of environmental stress, it’s production increase by several folds (Demidchik 2015; Hossain et al. 2015; Mignolet-Spruyt et al. 2016; Czarnocka and Karpiński 2018). Lower amount of H2O2 accumulation in plant cells activates defensive signaling pathways in plants, whereas higher concentrations are detrimental for cells and cause programmed cell death (Dat et al. 2000; Hossain et al. 2015), and can inactivate Calvin cycle (Halliwell 2006) and other vital enzymes (Das and Roychoudhury 2014). Overproduction of ROS can also result in damage to membrane lipids (Das and Roychoudhury 2014) and leads to the generation of malondialdehyde (MDA), which can cause protein degradation, disruption of membrane fluidity and inhibition of ion transport. Moreover, MDA is often considered as a pivotal hallmark of the level of oxidative damage in plants (Li et al. 2013; Awasthi et al. 2017; Tahjib-Ul-Arif et al. 2018a, b, c).
Plants have evolved mechanisms to restrict the detrimental consequence of drought-induced oxidative burst by scavenging excessive ROS through enzymatic, such as catalase (CAT), ascorbate peroxidase (APX), guaiacol peroxidase (POX), glutathione peroxidase (GPX), glutathione S-transferase (GST), etc., and non-enzymatic antioxidants, such as ascorbic acid (AsA), glutathione (GSH), carotenoids (Car), phenolic compounds (total phenolic compounds; TPC) and proline (Pro) (Gill and Tuteja 2010; Das and Roychoudhury 2014; Czarnocka and Karpiński 2018; Tahjib-Ul-Arif et al. 2019a, b). Under water deficit condition, plants also accumulate osmolytes, such as Pro and soluble sugars, that help to maintain cellular water balance and regulate essential ions (e.g. K+, Ca2+, Mg2+, etc.) uptake (Ashraf and Foolad 2007; Krasensky and Jonak 2012). Otherwise, drought-induced water loss can cause changes to the cation–anion ratio that lead to a loss of cell turgor, intervention of cell division, reduction in photosynthesis, and imbalances in carbon metabolism (Liu et al. 2005; Miller et al. 2010).
Glutathione (GSH), a tripeptide (γ-Glu-Cys-Gly) which can function as an antioxidant and rescue plant cells from ROS-induced oxidative injury (Gill and Tuteja 2010), has numerous roles in plant cells including stress tolerance, regulating cellular redox balance, modulating the expression of stress-related genes, xenobiotic detoxification, thiol group protection, influencing enzyme activity, protein and nucleotide synthesis, heavy metal chelation, plant growth and senescence (Gill et al. 2013; Gietler and Nykiel 2017; Hasanuzzaman et al. 2017). Moreover, transgenic plants over-expressing GSH biosynthetic genes displayed better salinity tolerance (Bae et al. 2013; Choe et al. 2013). These multiple roles of GSH in plants indicate that the manipulation of GSH levels and metabolism has the potential to enhance stress tolerance of plants (Gill and Tuteja 2010). Recently, exogenous application of GSH has received considerable attention by the scientific community due to its involvement in the enhancement of abiotic stresses tolerance by restricting the entry of toxic ions such as mercury, cadmium, excess sodium, etc. (Ding et al. 2017; Kim et al. 2017; Zhou et al. 2018), enhancing antioxidant defences, and by altering other physio-biochemical attributes (Ding et al. 2016; Zhou et al. 2017).
Rice is the second most important staple food worldwide and the most important food in Bangladesh, where drought stress can cause 30–60% yield losses (Ouk et al. 2006; Khoury et al. 2014; BBS 2018). Among the rice cultivars in Bangladesh, BRRI dhan29 is consider as a popular mega variety due to its acreage and yield potentiality. However, BRRI dhan29 is susceptible to drought stress and the county frequently experience significant yield loss due to drought stress (Islam et al. 2017). It is well established that drought stress impaired essential mineral nutrient uptake and translocation from root to shoot due to reduced transpiration and limited transporter activity (Silva et al. 2011; Bista et al. 2018; Chen et al. 2019), however, the roles of GSH on ion accumulation and translocation remains elusive. To the best of our knowledge, this is the first report describing the possible intriguing roles of exogenous GSH on ion homeostasis and ROS metabolism in rice plants under drought stress.
2 Materials and Methods
2.1 Plant Growth Conditions and Treatments
Rice (cv. BRRI dhan29) seeds were sterilized with 2.5% sodium hypochlorite and 2% tween-20 solution for 15 min and were further washed four times with deionized water. The seeds were then imbibed in deionized water for 24 h. Following imbibition, the seeds were placed in petri dishes and kept for 2 days at 28 ± 2 °C in the dark to allow germination. After germination, 150 germinated rice seeds were sown on floating nets in 350 mL plastic pots, containing distilled water. From the 3rd day of sowing, the rice seedlings were grown in modified hydroponic Cooper’s nutrient solution (Cooper 1988) as described in Tahjib-Ul-Arif et al. (2019a) in a growth chamber (temperature of 25 ± 2 °C and relative humidity of 65–70%). Used solution was changed at 3-day intervals for the duration of the experiment.
Twelve-day-old rice seedlings were pretreated with 0.2 mM GSH, in the nutrient solution, for three days. After pretreatment, 15-day-old GSH-pretreated and untreated rice seedlings were subjected to drought stress (induced by 15% polyethylene glycol 6000; PEG) for a duration of 4 days. GSH-pretreated seedlings were also grown in nutrient solution without PEG. In summary, the study comprised a total of four treatments; (1) Control (C): 0% PEG + 0 mM GSH, (2) Drought (D): 15% PEG + 0 mM GSH, (3) GSH: 0% PEG + 0.2 mM GSH and (4) D + GSH: 15% PEG + 0.2 mM GSH. Randomized complete block design with five independent replicates for each treatment was used to carry out the experiment. Different morphological and biochemical parameters of the rice seedlings were measured after 4 days of drought stress treatment.
2.2 Determination of Plant Growth Parameters, Relative Water Content (RWC), Pro, Photosynthetic Pigments and Carotenoid (Car) Contents
Plant growth parameters and RWC were measured according to Tahjib-Ul-Arif et al. (2018b, c). The methods of Zhang and Huang (Zhang and Huang 2013), with minor modifications, were used to measure the Pro content of rice leaves and standard curve were produced with analytical grade Pro.
Chlorophylls (Chl a and Chl b) and Car were extracted from the 2nd leaves (0.5 g) of the rice seedlings, by soaking in acetone (10 mL and 80% acetone) for 7 days inside the dark chamber. The absorbance of the acetone extracts was measured at 645, 663 and 470 nm, using a UV–Vis spectrophotometer (Shimadzu, UV-1201, Kyoto, Japan) and Chl a, Chl b and Car contents were determined according to the method of Lichtenthaler (1987).
2.3 Visual Detection of Superoxide and Determination of H2O2 and MDA Contents
To visualize \({\text{O}}_{2}^{ - }\) in rice leaves, the 3rd leaves of rice seedlings were soaked in 0.1% nitroblue tetrazolium (NBT) reagent and stored in the dark for 12 h, according to the method of Tahjib-Ul-Arif et al. (2019a). MDA content (product of lipid peroxidation) was measured in the 3rd leaves of the seedlings according to the method of Heath and Packer (Heath and Packer 1968), using an extinction coefficient of 155 mM−1 cm−1.
2.4 Determination of Activities of Antioxidant Enzymes
Three antioxidant enzymes activities were determined in extracts from the 3rd leaves of rice seedlings. Fresh leaf samples (0.05 g) were homogenized with 1 mL of 50 mM potassium-phosphate buffer (pH 8.0), using pre-chilled mortars and pestles. The homogenates were centrifuged at 11,500 × g for 10 min, and the resultant supernatants were collected to analyze the activity of CAT (EC 1.11.1.6) (Aebi 1984), APX (EC 1.11.1.11) and POX (EC: 1.11.1.7) (Nakano and Asada 1981). All procedures were performed at 0–4 °C, and all of the spectrophotometric assays were performed using a UV–Vis spectrophotometer (Shimadzu, UV-1201, Kyoto, Japan).
2.5 Determination of Total Soluble Sugars, Total Phenolic Compounds and Ascorbate
Total soluble sugars (TSS) were determined using the modified anthrone method as previously reported by Ciha and Brun (1978). Dried shoot samples (0.3 g of finely powdered sample) were extracted with 10 mL of 80% ethanol, and the resultant extract was filtered through Whatman filter paper (11 µm pore size) to remove particulates. 1 mL of 1:10 diluted extract was then mixed with 4 mL of 2% anthrone solution, and the mixture was then heated for 10 min and then cooled on ice. The absorbance of the solution was then measured at 620 nm using a UV–Vis spectrophotometer (Shimadzu, UV-1201, Kyoto, Japan). The TSS contents of the samples were determined from the standard curve prepared using glucose solutions of different concentrations (1.0 mg mL−1, 0.8 mg mL−1, 0.6 mg mL−1, 0.4 mg mL−1 and 0.2 mg mL−1).
The TPC was determined according to the method of Singleton and Rossi (1965), using the Folin-Ciocalteu reagent. A standard curve was constructed using gallic acid as a standard, and the results were expressed as µg of gallic acid equivalents (GAE) per gram of DW (µg GAE g−1 DW). The AsA content was determined according to the method of Jagota and Dani (1982). AsA was used to construct a standard curve, and the results are expressed as nmol g−1 FW.
2.6 Determination of Na+, K+, Ca2+ and Mg2+ Ion from Shoot Tissues
After harvesting, rice seedlings were washed rigorously with deionized water to eliminate surface contaminants. Seedlings were then oven dried at 70 °C for 4 days, and the finely powdered plant material was digested with HNO3: HClO4 (2:1v/v) mixture at 220 °C for 2 h according to the method of Tahjib-Ul-Arif et al. (2018b). Na+ and K+ contents were quantified by flame photometry (Jencon PFP 7, JENCONS-PLS, UK). Ca2+ and Mg2+ contents were measured by titration, using ethylenediaminetetraacetic acid disodium salt according to the methods of Schmid and Reilley (1957).
2.7 Statistical Analysis
A one-way analysis of variance was performed using Minitab 17.0. Different letters denote the statistically significant differences between treatments at P < 0.05, according to the Tukey’s honest significant differences test. The heatmap and Euclidean complete linkage-based cluster analyses were performed using MetaboAnalyst 4.0 (www.metaboanalyst.ca) and the principal component analysis (PCA) was constructed using R 3.5.3.
3 Results
3.1 GSH-Pretreatment Improved Plant Growth and Development Under Drought Stress
A significant (P < 0.05) reduction in shoot length (SL), root length (RL), shoot fresh weight (SFW), root fresh weight (RFW), shoot dry weight (SDW) and root dry weight (RDW) (38.65, 32.86, 67.61, 44.05, 43.23 and 56.06%, respectively) was found in rice seedlings in response to drought stress as compared to unstressed control (Table 1). The pretreatment of unstressed seedlings with exogenous GSH did not influence SL, RL, SFW, RFW, SDW and RDW as compared to control plants. However, GSH-pretreated drought-stressed seedlings showed significant increases in SL, RL, SFW, RFW, SDW and RDW (42.68, 34.27, 182.50, 25.20, 30.68 and 130.17%, respectively) when compared to seedlings subjected to drought stress only (Table 1).
3.2 GSH-Pretreatment Protected Photosynthetic Pigments Under Drought Stress
Under drought stress, Chl a, Chl b and total Chl contents were decreased significantly (55.97, 20.61 and 49.73%, respectively) compared with control seedlings (Table 1). In contrast, GSH-pretreated drought-stressed seedlings showed a marked increase in Chl a, Chl b, and total Chl levels (83.27, 25.39 and 67.14%, respectively) as compared to seedlings under drought stress only. GSH pretreatment in well-watered rice seedling enhanced the Chl a, Chl b, and total Chl contents by 36.54, 117.37 and 50.81%, respectively, when compared with untreated control seedlings (Table 1).
3.3 GSH-Pretreatment Elevated RWC and TSS Whereas Declined Pro Contents Under Drought Stress
The RWC and Pro content are important parameters for assessing the physiological water status of plants. RWC and Pro showed an inverse relationship in drought-stressed rice seedlings. In response to drought stress, the RWC decreased by 62.97% whereas the Pro content showed a significant (P < 0.05) increase of 69.69% when compared to control seedlings (Fig. 1a, b). In contrast, GSH-pretreated drought-stressed seedlings showed an increase in RWC content of 145.75% when compared to only drought-stressed seedlings, while the Pro content showed a significant decrease of 41.44% when compared to control seedlings (Fig. 1a, b). In addition, RWC and Pro in unstressed control and GSH-pretreated unstressed seedlings showed no significant changes.

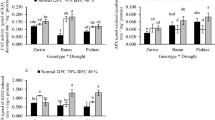
Effects of exogenous glutathione (GSH) on a relative water content (RWC); b proline content and c total soluble sugars (TSS) content of rice seedlings. Plotted data represent the average of five individual replicates for each treatment (n = 5, three leaves per replicate). Vertical bar indicates the standard error. The letter on top of each bar denotes a statistically significant difference at P < 0.05 (Tukey’s honest significant differences test). Control (C): 0% PEG + 0 mM GSH; Drought (D): 15% PEG + 0 mM GSH; GSH: 0% PEG + 0.2 mM GSH; D + GSH: 15% PEG + 0.2 mM GSH
Drought stress significantly (P < 0.05) increased the shoot TSS content of 53.75% when compared to control seedlings. In contrast, GSH-pretreated seedlings displayed further increase in TSS of 27.61% when compared to drought-stressed seedlings. GSH-pretreated unstressed seedlings showed a significant escalation in TSS content of 69.83% when compared to untreated control seedlings (Fig. 1c).
3.4 GSH-Pretreatment Reduced ROS and MDA Contents in the Shoots and Root Tissues Under Drought Stress
Drought-stressed rice seedlings showed the highest O·−2 accumulation (observed as dark blue spots within the leaf blade) as compared to controls. In contrast, GSH-pretreated drought-stressed seedlings showed less O·−2 accumulation as compared to seedlings without drought stress (Fig. 2a).

Effects of exogenous glutathione (GSH) on deposition of superoxide in leaves (a); H2O2 content in shoot (b), root (c); and MDA content in shoot (d), root (e) of rice seedlings. Data represented in figures are the mean of five replicates for each treatment (n = 5, three leaves per replicate). Vertical bar indicates the standard error. Different letters represent significant differences at P < 0.05 (Tukey’s honest significant differences test). Control (C): 0% PEG + 0 mM GSH; Drought (D): 15% PEG + 0 mM GSH; GSH: % PEG + 0.2 mM GSH; D + GSH: 15% PEG + 0.2 mM GSH
Drought stress led to a significant (P < 0.05) increase in MDA and H2O2 contents of 83.20 and 37.2% in shoots, respectively, and 47.60 and 68.05% in roots, respectively, when compared to control seedlings (Fig. 2b–e). In contrast, GSH-pretreated drought-stressed seedlings showed a decrease in MDA and H2O2 contents (22.25 and 14.40% in the shoot and 18.98 and 31.80% in root tissues, respectively) as compared to seedlings treated with drought stress only (Fig. 2b–e). Furthermore, only GSH-pretreated seedlings showed no significant changes in MDA and H2O2 contents when compared with untreated control seedlings.
3.5 GSH-Pretreatment Improved Antioxidant Metabolism in Shoots and Roots of Rice Seedlings Under Drought Stress
The activities of CAT, APX and POX in the leaves and roots of rice seedlings were significantly affected by drought stress and GSH pretreatment. Drought-stressed seedlings showed a marked decline in the activities of CAT, APX and POX (30.31, 25.60 and 40.72%, respectively) in shoots, compared to untreated control seedlings. In root tissues, CAT and POX activities decreased by 60.53, and 71.21%, respectively, but APX activity increased by 109.67%, compared to untreated control seedlings (Fig. 3). In contrast, exogenous pretreatment of seedlings with GSH resulted in increases in CAT, APX and POX activities by 66.54, 63.35 and 30.60%, respectively, in shoots and by 352.70, 84.20 and 86.81%, respectively, in the roots of seedlings under drought stress, compared to the seedlings exposed to drought stress without GSH pretreatment. Moreover, GSH treatment of seedlings did not significantly alter the activities of CAT, APX and POX in the absence of drought stress (Fig. 3).

Modulation of enzymatic antioxidants activity catalase (CAT) activity in shoot (a) and root (b); ascorbate peroxidase (APX) activity in shoot (c) and root (d) and peroxidase (POX) activity in shoot (e) and root (f) of rice seedlings by exogenously applied glutathione (GSH). Data represented in figures are the mean of five replicates for each treatment (n = 5, three leaves per replicate). The vertical bar indicates the standard error. Letter on top of the bar denotes the statistically significant difference at P < 0.05 (Tukey’s honest significant differences test). Control (C): 0% PEG + 0 mM GSH; Drought (D): 15% PEG + 0 mM GSH; GSH: 0% PEG + 0.2 mM GSH; D + GSH: 15% PEG + 0.2 mM GSH
3.6 GSH Positively Affected Non-enzymatic Antioxidants Under Drought Stress
TPC levels showed a significant (P < 0.05) increase of 44.17% in drought-stressed seedlings, compared to controls. GSH-pretreated unstressed or drought-stressed seedlings also showed a significant (P < 0.05) increase in TPC contents of 39.26 and 38.65%, respectively, compared to control seedlings. However, there was no significant difference in TPC levels observed for GSH-pretreated and untreated drought-stressed seedlings (Fig. 4c). Likewise, AsA and Car contents decreased significantly (P < 0.05) in response to drought stress by 20.76 and 54.81%, respectively, compared to control seedlings (Fig. 4a, b). In contrast, GSH-pretreated drought-stressed seedlings showed a significant (P < 0.05) increase in AsA and Car contents of 12.41 and 152.43%, respectively, compared to the seedlings treated with drought stress only. Furthermore, GSH-pretreated seedlings in the absence of drought showed no significant change in AsA content, but significant change of 42.14% in Car content when compared to control seedlings (Fig. 4a, b).

Effects of exogenous glutathione (GSH) on a ascorbic acid (AsA) content; b carotenoids content and c total phenolic compounds (TPC) content of rice seedlings. Plotted data represent the average of five replicates for each treatment (n = 5, three leaves per replicate). Vertical bar indicates the standard error. Letter on top of bar denotes the statistically significant difference at P < 0.05 (Tukey’s honest significant differences test). Control (C): 0% PEG + 0 mM GSH; Drought (D): 15% PEG + 0 mM GSH; GSH: 0% PEG + 0.2 mM GSH; D + GSH: 15% PEG + 0.2 mM GSH
3.7 GSH-Pretreatment Maintained Mineral Homeostasis of Rice Shoots Under Drought Stress
The Na+, K+, Ca2+ and Mg2+ ion contents decreased by 34.91, 27.45, 19.02 and 13.00%, respectively, in drought-stressed seedlings relative to control (Table 2). However, GSH-pretreated drought-stressed seedlings showed enhance Na+, K+, Ca2+ and Mg2+ ion contents of 22.92, 20.33, 56.44 and 20.48%, respectively, when compared to the seedlings subjected to drought stress without GSH pretreatment. Moreover, GSH-pretreated non-stressed seedlings showed no significant differences in ion contents relative to control seedlings (Table 2).
3.8 Hierarchical Clustering and PCA Analysis
The morpho-physiological and biochemical data from seedlings (both drought stress and stress-free control conditions) were undergone hierarchical clustering, heat map analysis and PCA (Fig. 5). In terms of variables, three clusters (cluster-A, B and C) were formed in hierarchical clustering (Fig. 5a). The variables, POX shoot, POX root, RFW, Na+, SDW, AsA, K+, SL, RL, SFW and RWC, were clustered into cluster-A and Chl b, Car, Chl a, total Chl, RDW, CAT shoot, Mg2+, Ca2+, APX shoot, and CAT root were clustered into cluster-B. In the heat map, all the parameters of cluster-A and B displayed a significantly decreasing trend in drought-stressed rice seedlings. However, the application of exogenous GSH to drought-stressed seedlings (D + GSH) upregulated the parameters in cluster-A and B. All parameters in cluster-A and B were positively related to growth and tolerance of rice seedlings to drought stress. Thus, it can be inferred that exogenous GSH counteracted drought-induced damage. Cluster-C was comprised of H2O2 shoot, H2O2 root, MDA shoot, MDA root, Pro, APX root, TSS and TPC. With respect to stress-free control seedlings, these parameters exhibited increasing trends under drought stress. However, application of GSH to drought-stressed rice seedlings resulted in differential responses with respect to cluster-C parameters such as lower H2O2 shoot, H2O2 root, MDA shoot, MDA root, Pro values and increased values for APX activity and TSS, with no change for TPC. With respect to treatments, three clusters, cluster-X, Y and Z, were found by hierarchical clustering (Fig. 5a) and usually similar attributes clustered together. GSH application to seedlings prior to drought stress improved drought stress tolerance of rice seedlings, with ‘D + GSH’ being in the cluster-Y. In contrast, the only drought ‘D’ treatment fitted into the cluster-Z and the cluster-X consisted of the ‘C’, and ‘GSH’ treatments. It clearly shows that the application of GSH to unstressed seedlings did not have a significant effect on the different variables when compared with control seedlings. Afterward, this analysis was carried out to reveal the association among different variables with other treatment groups (Fig. 5b). The PCA biplot reveals clear segregation of the treatments to three groups (C and GSH; D; and D + GSH) and their biological replicates. The two components of PCA (PC1 and PC2) collectively explained 79.8% of data variability. The results displayed that some variables of cluster-A were intimately associated with ‘C’ and ‘GSH’ treatments, whereas other variables of cluster-A also closely associated with ‘D + GSH’ treatment. The cluster-B variables showed negative correlation with growth parameters and were interlinked with only ‘D’ treatment.

Hierarchical clustering and heatmap (a) to elucidate the treatment variable relationships in rice plants under both drought-free and drought conditions. The mean values of the various parameters obtained in this study were normalized and clustered. The color scale displays the intensity of normalized mean values of different parameters. The entire dataset was analyzed using principal component analysis (PCA) (b). The lines which is originating from the central point of biplots, display negative or positive correlations of different variables. The nearness of the lines indicates correlation strength with a particular treatment. Catalase-root (CAT.R), catalase-shoot (CAT.S), ascorbate peroxidase-root (APX.R), ascorbate peroxidase-shoot (APX.S), guaiacol peroxidase-root (POX.R), guaiacol peroxidase-shoot (POX.S) malondialdehyde-root (MDA.R), malondialdehyde-shoot (MDA.S), hydrogen peroxide-root (H2O2.R), hydrogen peroxide-shoot (H2O2. S), chlorophyll a (Chl.a), chlorophyll b (Chl.b), total chlorophyll (total chl), carotenoid (Car), ascorbate (AsA), proline (Pro.S), total soluble sugar (TSS), total phenolic contents (TPC), shoot length (SL), root length (RL), shoot dry weight (SDW), root dry weight (RDW), shoot fresh weight (SFW), root fresh weight (RFW), relative water content (RWC), sodium (Na+), potassium (K+), magnesium (Mg2+), calcium (Ca2+). Control (C): 0% PEG + 0 mM GSH; Drought (D): 15% PEG + 0 mM GSH; GSH: 0% PEG + 0.2 mM GSH; D + GSH: 15% PEG + 0.2 mM GSH
4 Discussion
The importance of GSH for modulating physiological responses of plants under unfavorable environments has been described in a diverse plants species (Cao et al. 2017; Sabetta et al. 2017), but has not been studied so far in rice under drought stress. In the present study, GSH was found to play a critical role in enhancing drought stress tolerance by modulating physiological and biochemical traits or processes, such as root and shoot growth characteristics, photosynthetic pigments, enzymatic and non-enzymatic antioxidants, secondary metabolites and regulation of ion fluxes.
Drought stress severely inhibits plant growth and development (Salehi-Lisar and Bakhshayeshan-Agdam 2016; Fahad et al. 2017). In the present study, drought stress caused a considerable reduction in seedling growth, as reflected in reduced SL, RL, SFW, RFW, SDW and RDW (Table 1). Similar reduction in shoot and root traits was also demonstrated in rice (Sohag et al. 2020a), wheat (Triticum durum) (Boutraa et al. 2010) and barley (Hordeum vulgare) (Hellal et al. 2018). This growth inhibition might be due to lower photosynthetic pigment levels, cell dehydration, osmotic imbalance, ROS production and improper nutrient uptake (Tables 1, 2; Figs. 1a–c, 2) (Forni et al. 2017). However, GSH-pretreated drought-stressed seedlings displayed mitigation of the negative effects of drought, with improved SL, RL, SDW and RDW (Table 1). The positive influence of exogenous GSH in modulating growth and yield under drought or salinity stress has also been observed in Arabidopsis (Chen et al. 2012; Cheng et al. 2015), mung bean (Vigna radiata) (Nahar et al. 2015) and in soybean (Glycine max) (Akram et al. 2017). The PCA also indicated that GSH-pretreated drought-stressed rice plants showed positive relationship with growth-related parameters (Fig. 5b), suggesting protective roles of GSH in diminishing the toxic results of drought on rice seedling growth and development.
Additionally, drought-induced growth inhibition of rice plants (Table 1) might be connected with unfavorable ionic homeostasis, as water availability is directly related to mineral uptake (Stagnari et al. 2016). Our result of PCA demonstrated a strong and negative correlations between drought treatment and uptake of minerals, such as Na+, K+, Ca2+ and Mg2+ (Fig. 5b). Specifically, in this study, drought stress reduced Ca2+ and K+ levels in rice shoots, which in turn probably hampered growth-related features of rice plants (Table 2), because Ca2+ requires in cell division, cell enlargement and cell wall formation, and K+ involves in enzyme activation and photosynthesis (Hepler 2005; Silva et al. 2011). In accordance with our results, a similar decrease in essential ions in response to drought stress was reported in rice (Andrade et al. 2018) and maize (Zea mays) (Kaya et al. 2006). Low K+ reduces the osmotic pressure of the guard cells, thereby lowering their ability to swell, and can lead to partial stomatal closure (Battie-Laclau et al. 2013). Hence, the lower K+ levels observed in rice seedlings in the present study could lead to stomatal dysfunction, poor stomatal regulation of water loss and reduced photosynthesis, and subsequently, this could result in poor growth of rice seedlings under drought (Table 1). A recent study suggests that optimum levels of Na+ can have a positive role in plants, as Na+ can perform some metabolic functions in plants under low K+ conditions due to the structural similarities (Maathuis 2014). Our results also showed that drought stress lowered Na+ content in rice shoots (Table 2), which was coincided with the previous studies in sunflower (Helianthus annuus) (Gunes et al. 2008) and tomato (Solanum lycopersicum) (Nahar and Gretzmacher 2002) plants under drought stress. However, pretreatment of seedlings with GSH prior to drought stress imposition enhanced Na+, K+, and Ca2+ levels in drought-stressed rice seedlings (Table 2) and partly ameliorated the drought-induced reduction in growth of rice seedlings (Table 1). Enhancement of K+ and Ca2+ levels was also reported for GSH-treated salt-stressed tomato (Zhou et al. 2018) and cadmium-stressed rice (Wang et al. 2011) plants. The enhanced level of essential nutrient ions in GSH-treated drought-stressed rice seedling might be due to the modulation of genes regulating nutrient uptake and transport (Ding et al. 2017; Gheshlaghi et al. 2020).
Leaf osmotic adjustment is crucial for the survival of plants under osmotic stress which is usually regulated by the amassing compatible solutes, such as Pro and TSS (Ashraf and Foolad 2007). Thus, elevation of Pro levels is regarded as an adaptive response of drought stress (da Silva Folli-Pereira et al. 2016), and increased Pro levels were observed in the seedlings grown under drought stress only (Fig. 1b). This finding is in agreement with similar studies on rice (Dien et al. 2019). In contrast, Pro content was much lower in GSH-pretreated drought-stressed rice seedlings (Fig. 1b), which suggested that GSH might have contributed to reduce the osmotic stress. A similar finding was also reported for mung bean (Nahar et al. 2015) under salt stress. Moreover, in the present study, TSS levels in the shoots increased significantly in response to drought stress and pretreatment with GSH and subsequent imposition of seedlings to drought stress further enhanced its contents (Fig. 1c) which is in agreement with similar studies on Haloxylon persicum and Haloxylon aphyllum (Arabzadeh and Khavari-Nejad 2013) plants. In line with these findings, the RWCs of rice seedlings declined under drought stress (Fig. 1a), which is in agreement with other research findings (Kaya et al. 2006; Meher et al. 2018). However, higher RWC was observed in GSH-pretreated rice seedlings under drought stress (Fig. 1a), a similar finding was also reported for mung bean (Nahar et al. 2015). Importantly, the results of PCA revealed that TSS showed a strong positive correlation and Pro showed a negative correlation with GSH-pretreated drought-stressed plants (Fig. 5b), which let us conclude that GSH pretreatment in drought-stressed plants enhanced TSS accumulation and maintained higher RWC, thereby reduced higher Pro accumulation in plant cells (Fig. 1).
Like osmotic stress, drought also induces oxidative stress in plants, mainly by interrupting electron flow during photosynthesis (Fahad et al. 2017). In the present study, drought stress substantially elevated O·−2 , H2O2 and MDA content. Similar increase in ROS levels in response to drought and other stresses in rice also reported by others (Andrade et al. 2018; Afrin et al. 2019; Wang et al. 2019; Sohag et al. 2020a, b). ROS directly attack membrane lipids and disorganize photosynthetic pigments (Gill and Tuteja 2010), which was indicated by the increased levels of MDA content and decreased levels of Chl pigments under drought stress (Fig. 2d, e; Table 1). Collectively, these findings could result in reduced plant growth and development as observed in the present study (Table 1). Particularly, our results have proven that exogenous GSH pretreatment inhibited the accumulation of ROS in plant cells (Fig. 2) and thereby maintained cell-membrane integrity and prevented Chl degradation in drought-exposed rice plants (Table 1). In agreement with our results, several recent studies were also reported the protective roles of GSH against oxidative stress in mung bean under drought stress (Nahar et al. 2015), in tomato and soybean under salinity stress (Akram et al. 2017; Zhou et al. 2017) and in cucumber (Cucumis sativus) under high temperature stress (Ding et al. 2016). Consequently, the PCA biplot projected that drought-stressed only rice plants had a stronger positive correlation with ROS and MDA contents in shoots and roots than GSH-pretreated drought-stressed rice plants (Fig. 5b). Besides, down-regulating the ROS-induced Chl breakdown, GSH pretreatment also upregulated the uptake of divalent cations, such as Mg2+ (Table 2), which might contributed to augment the Chl biosynthesis, thus maintains higher photosynthetic capacity leading to improved growth (Table 1, 2).
The severity of ROS-induced oxidative damage can be mitigated by upregulating the enzymatic and non-enzymatic antioxidants such as CAT, APX, POX, AsA, GSH, Car, TPC etc. These antioxidants exert a synergic effect to eliminate excessive drought-induced ROS accumulation and hence oxidative damage to cells (Gill and Tuteja 2010; Caverzan et al. 2016). In the present study, CAT, POX (in shoots and roots) and APX (in shoots) activity decreased, whereas the APX in roots increased in rice plants grown under drought stress (Fig. 3), which is in compliance with previous findings on rice, maize and barley plants (Sharma and Dubey 2005; Guo et al. 2006; Salekjalali and Jafari 2011; Xin et al. 2011). The non-synchronous activity of APX in root and shoot tissues of rice plants under drought stress has been reported previously (Guo et al. 2006). However, it is likely that CAT, APX and POX activities in drought-stressed seedling did not sufficient to neutralize the overproduced ROS, as also evident by the higher amounts of H2O2 and MDA observed in drought-exposed rice plants leading to diminished seedling growth and development (Fig. 2, Table 1). In contrast, GSH pretreatment augmented the activities of these antioxidant enzymes (Fig. 3), and resulted in a lower H2O2 and MDA accumulation under drought stress (Fig. 2b–e). Moreover, our results of PCA showed that GSH-treated drought-stressed plants had a greater positive correlations with antioxidants enzymes than those of the plants treated with drought stressed only, clearly suggesting that GSH upregulate antioxidants enzymes (Fig. 5b). A number of earlier studies have claimed that exogenous GSH application stimulates the antioxidants capacity and stress response gene expression of plants under different abiotic stresses (Chen et al. 2012; Hussain et al. 2016; Akram et al. 2017; Hasanuzzaman et al. 2018; Pei et al. 2019).
Plant secondary metabolites including phenolics exhibits several protective mechanism against stresses (Akula and Ravishankar 2011). Our data showed that drought stress decreased the examined non-enzymatic antioxidants except TPC (Fig. 4c), thus reduced the antioxidant capacity of the rice plants and eventually plant growth hampered due to oxidative stress (Table 1; Fig. 2). It was also evidenced in this study that exogenous GSH contributed to the removal of excess ROS by enhancing the levels of non-enzymatic antioxidants like AsA, TPC and Car (Fig. 4c). In support of this finding, GSH pretreatment showed higher levels of AsA and Car in some plant species like mung bean (Nahar et al. 2015), soybean (Seminario et al. 2017) and maize (Taïbi et al. 2016). However, in the present study, the level of TPC increased when exposed to drought stress or pretreated with GSH (Fig. 4c) and these findings are in accordance with previous reports conducted with Hypericum pruinatum (Caliskan et al. 2017) and garden cress (Lepidium sativum) (Ahmed et al. 2012) under drought and salt stress, respectively. The elevation of the TPC would increase the ROS and lipid free radicals scavenging capacity of rice plants (Gill and Tuteja 2010; Waśkiewicz et al. 2013; Das and Roychoudhury 2014).
Overall, the heatmap revealed that cluster-A and cluster-B variables showed a decreasing trend and cluster-C variables showed an increasing trend in drought-stressed seedlings, when compared with control, but displayed an inverse trend in GSH-pretreated drought-stressed seedlings, compared to that of seedlings treated with drought stress only (Fig. 5a). This provides a strong evidence that GSH pretreatment could enhance drought stress tolerance in rice plants. Moreover, the PCA revealed that the antioxidant enzymes (CAT, APX and POD) activities and mineral contents were closely and positively associated with the ‘D + GSH’ treatment (Fig. 5b), which suggests that the exogenous GSH-mediated growth improvement and drought stress tolerance exclusively conferred by enhanced antioxidant activities and mineral homeostasis.
5 Conclusion
Based on the above findings, it can be concluded that drought stress causes severe oxidative stress in rice seedlings, as indicated by higher ROS and ROS-induced damage, as well as disruption of essential ion homeostasis and reduced seedling growth (Fig. 6). Importantly, pretreatment with GSH can help to limit the ROS-induced damage, maintain ion homeostasis and water balance, and hence results in improved growth of rice seedlings under drought through the modulation of different morpho-physiological and biochemical processes (Fig. 6). However, further studies are required to find out how GSH regulates ion homeostasis in rice seedlings under drought stress.

Schematic representation of drought-induced growth inhibition and improvement of growth by exogenously GSH pretreatment. Drought exerts its harmful effects on plants by reducing the Chl content resulting in a decreased rate of photosynthesis. It also imbalances nutrient uptake by roots influencing Mg2+, K+ and Ca2+ ion concentrations. Lower Mg2+ levels disrupt Chl formation, low K+ levels imbalance stomatal regulation and lower Ca2+ inhibits cell division. Water deficit increases superoxide which in turn increases H2O2 content, contributing to excessive production of ROS. Drought induces lower levels of Car and AsA which facilitates increased superoxide levels. Lower levels of antioxidant enzymes, such as CAT, APX and POX, are also caused by drought, resulting in an increase in H2O2. RWC also decreases in plants under drought limiting gas exchange and photosynthesis. Lipid peroxidation is also enhanced by drought, which damages cell membrane and promotes lipid peroxidation. In contrast, GSH pretreatment can restore the growth of drought-stressed plants and reduce oxidative damage. GSH can maintain Chl contents, help to maintain photosynthesis, and also help maintain nutrient uptake. Pretreated GSH induced increase in CAT, APX, POX, Car and AsA maintain the optimum level of ROS. GSH also increase RWC and maintained higher gas exchange and photosynthesis. GSH reduces the cell membrane damage by lowering lipid peroxidation which was confirmed through reduced MDA. Chl, Chlorophyll; H2O2, hydrogen peroxide; ROS, reactive oxygen species; Car, carotenoids; AsA, ascorbic acid; CAT, catalase; APX, ascorbate peroxidase; POX, guaiacol peroxidase; RWC, relative water content; MDA, malondialdehyde; and GSH, glutathione
References
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
Afrin S, Tahjib-Ul-Arif M, Sakil M, Sohag A, Polash M, Hossain M (2019) Hydrogen peroxide priming alleviates chilling stress in rice (Oryza sativa L.) by enhancing oxidant scavenging capacity. Fundum Appl Agric 4:713–722
Ahmed AR, Gabr AM, AL-Saye HM, Smetanska I (2012) Effect of drought and salinity stress on total phenolic, flavonoids and flavonols contents and antioxidant activity in vitro sprout cultures of garden cress (Lepidium sativum). J Appl Sci Res 8:3934–3942
Akram S, Siddiqui MN, Hussain BMN, Al Bari MA, Mostofa MG, Hossain MA, Tran LSP (2017) Exogenous glutathione modulates salinity tolerance of soybean [Glycine max (L.) Merrill] at reproductive stage. J Plant Growth Regul 36:877–888
Akula R, Ravishankar GA (2011) Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal Behav 6(11):1720–1731
Andrade FR, da Silva GN, Guimarães KC, Barreto HBF, de Souza KRD, Guilherme LRG, Faquin V, dos Reis AR (2018) Selenium protects rice plants from water deficit stress. Ecotoxicol Environ Saf 164:562–570
Arabzadeh N, Khavari-Nejad RA (2013) Effect of drought stress on superoxide dismutase activity in two species of Haloxylon aphyllum and Haloxylon persicum. Pak J Biol Sci 16:351–361
Ashraf M, Foolad MR (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exp Bot 59:206–216
Awasthi JP, Saha B, Regon P, Sahoo S, Chowra U, Pradhan A, Roy A, Panda SK (2017) Morpho-physiological analysis of tolerance to aluminum toxicity in rice varieties of North East India. PLoS ONE 12:e0176357
Bae MJ, Kim YS, Kim IS (2013) Transgenic rice overexpressing the Brassica juncea gamma glutamylcysteine synthetase gene enhances tolerance to abiotic stress and improves grain yield under paddy field conditions. Mol Breed 31:931–945
Barik SR, Pandit E, Mohanty SP, Pradhan SK, Mohapatra T (2019) Genetic mapping of morpho-physiological traits involved during reproductive stage drought tolerance in rice. bioRxiv 590075
Battie-Laclau P, Laclau JP, de Cassia Piccolo M, Arenque BC, Beri C, Mietton L, Muniz MRA, Jordan-Meille L, Buckeridge MS, Nouvellon Y, Ranger J, Bouillet JP (2013) Influence of potassium and sodium nutrition on leaf area components in Eucalyptus grandis trees. Plant Soil 371:19–35
BBS (Bangladesh Bureau of Statistics) (2018) Yearbook of agricultural statistics of Bangladesh. Statistics Division, Ministry of Planning, Government of the People’s Republic of Bangladesh, Dhaka
Bista DR, Heckathorn SA, Jayawardena DM, Mishra S, Boldt JK (2018) Effects of drought on nutrient uptake and the levels of nutrient-uptake proteins in roots of drought-sensitive and -tolerant grasses. Plants (Basel) 7(2):28
Boutraa T, Akhkha A, Al-Shoaibi AA, Alhejeli AM (2010) Effect of water stress on growth and water use efficiency (WUE) of some wheat cultivars (Triticum durum) grown in Saudi Arabia. J Taibah Univ Sci 3:39–48
Caliskan O, Radusiene J, Temizel KE, Staunis Z, Cirak C, Kurt D, Odabas MS (2017) The effects of salt and drought stress on phenolic accumulation in greenhouse-grown Hypericum pruinatum. Ital J Agron 12:271–275
Cao F, Fu M, Wang R, Diaz-Vivancos P, Hossain MA (2017) Exogenous glutathione-mediated abiotic stress tolerance in plants. In: Hossain MA, Mostofa MG, Diaz-Vivancos P, Burritt DJ, Fujita M, Tran LSP (eds) Glutathione in plant growth, development, and stress tolerance, 1st edn. Springer, Basel, pp 171–194
Caverzan A, Casassola A, Brammer SP (2016) Antioxidant responses of wheat plants under stress. Genet Mol Biol 39:1–6
Chen JH, Jiang HW, Hsieh EJ, Chen HY, Chien CT, Hsieh HL, Lin TP (2012) Drought and salt stress tolerance of an Arabidopsis Glutathione S-transferase U17 knockout mutant are attributed to the combined effect of glutathione and abscisic acid. Plant Physiol 158:340–351
Chen J, Qi T, Hu Z, Fan X, Zhu L, Iqbal MF, Yin X, Xu G, Fan X (2019) OsNAR2.1 Positively regulates drought tolerance and grain yield under drought stress conditions in rice. Front Plant Sci 10:197
Cheng MC, Ko K, Chang WL, Kuo WC, Chen GH, Lin TP (2015) Increased glutathione contributes to stress tolerance and global translational changes in Arabidopsis. Plant J 83:926–939
Choe YH, Kim YS, Kim IS, Bae MJ, Lee EJ, Kim YH, Park HM, Yoon HS (2013) Homologous expression of gamma-glutamylcysteine synthetase increases grain yield and tolerance of transgenic rice plants to environmental stresses. J Plant Physiol 170:610–618
Ciha AJ, Brun WA (1978) Effect of pod removal on nonstructural carbohydrate concentration in soybean tissue. Crop Sci 18:773–776
Cooper A (1988) 1. The system. 2. Operation of the system. In: The ABC of NFT. Nutrient film technique. Grower Books, London, England, pp 3–123. ISBN 0901361224
Czarnocka W, Karpiński S (2018) Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic Biol Med 122:4–20
da Silva Folli-Pereira M, Ramos AC, Bertolazi AA, Passamani LZ, Eutrópio FJ, da Conceição JM, Rasool N (2016) Water stress and higher plants: an overview. In: Ahamed P (ed) Water stress and crop plants: a sustainable approach, 1st edn. Wiley, Singapore, pp 422–451
Das K, Roychoudhury A (2014) Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front Environ Sci 2:53
Dat J, Vandenabeele S, Vranová E, Van Montagu M, Inzé D, Van Breusegem F (2000) Dual action of the active oxygen species during plant stress responses. Cell Mol Life Sci 57:779–795
Demidchik V (2015) Mechanisms of oxidative stress in plants: from classical chemistry to cell biology. Environ Exp Bot 109:212–228
Dien DC, Mochizuki T, Yamakawa T (2019) Effect of various drought stresses and subsequent recovery on proline, total soluble sugar and starch metabolisms in rice (Oryza sativa L.) varieties. Plant Prod Sci 22:530–545
Ding X, Jiang Y, He L, Zhou Q, Yu J, Hui D, Huang D (2016) Exogenous glutathione improves high root-zone temperature tolerance by modulating photosynthesis, antioxidant and osmolytes systems in cucumber seedlings. Sci Rep 6:35424
Ding S, Ma C, Shi W, Liu W, Lu Y, Liu Q, Luo ZB (2017) Exogenous glutathione enhances cadmium accumulation and alleviates its toxicity in Populus × canescens. Tree Physiol 37:1697–1712
Fahad S, Bajwa AA, Nazir U, Anjum SA, Farooq A, Zohaib A, Sadia S, Nasim W, Adkins S, Saud S, Ihsan MZ (2017) Crop production under drought and heat stress: plant responses and management options. Front Plant Sci 29:1147
Forni C, Duca D, Glick BR (2017) Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 410:335–356
Gheshlaghi Z, Khorassani R, Abadía J, Alvarez-Fernández A, Luis-Villarroya A, Fotovat A, Kafi M (2020) Glutathione supplementation prevents iron deficiency in Medicago scutellata grown in rock sand under different levels of bicarbonate. Plant Soil 446:43–63
Gietler M, Nykiel M (2017) Involvement of thiol-based mechanisms in plant growth, development, and stress tolerance. In: Hossain MA, Mostofa MG, Diaz-Vivancos P, Burritt DJ, Fujita M, Tran LSP (eds) Glutathione in plant growth, development, and stress tolerance, 1st edn. Springer, Basel, pp 59–98
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Gill SS, Anjum NA, Hasanuzzaman M, Trivedi DK, Ahmad I, Pereira E, Tuteja N (2013) Glutathione and glutathione reductase: a boon in disguise for plant abiotic stress defense operations. Plant Physiol Biochem 70:204–212
Gunes A, Pilbeam DJ, Inal A, Coban S (2008) Influence of silicon on sunflower cultivars under drought stress, I: growth, antioxidant mechanisms, and lipid peroxidation. Commun Soil Sci Plant Ana 39:1904–1927
Guo Z, Ou W, Lu S, Zhong Q (2006) Differential responses of antioxidative system to chilling and drought in four rice cultivars differing in sensitivity. Plant Physiol Biochem 44:828–836
Habiba U, Shaw R, Takeuchi Y (2012) Farmer’s perception and adaptation practices to cope with drought: perspectives from Northwestern Bangladesh. Int J Disaster Risk Reduct 1:72–84
Halliwell B (2006) Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol 141:312–322
Hasanuzzaman M, Nahar K, Anee TI, Fujita M (2017) Glutathione in plants: biosynthesis and physiological role in environmental stress tolerance. Physiol Mol Biol Plant 23:249–268
Hasanuzzaman M, Nahar K, Rahman A, Mahmud JA, Alharby HF, Fujita M (2018) Exogenous glutathione attenuates lead-induced oxidative stress in wheat by improving antioxidant defense and physiological mechanisms. J Plant Interact 13:203–212
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Hellal FA, El-Shabrawi HM, Abd El-Hady M, Khatab IA, El-Sayed SAA, Abdelly C (2018) Influence of PEG induced drought stress on molecular and biochemical constituents and seedling growth of Egyptian barley cultivars. J Genet Eng Biotechnol 16:203–212
Hepler PK (2005) Calcium: a central regulator of plant growth and development. Plant Cell 17:2142–2155
Hossain MA, Bhattacharjee S, Armin SM, QianP Xin W, Li H-Y, Burritt DJ, Fujita M, Tran LSP (2015) Hydrogen peroxide-priming modulates abiotic oxidative stress tolerance: insights from ROS detoxification and scavenging. Front Plant Sci 6:420
Hussain BN, Akram S, Burritt DJ, Hossain MA (2016) Exogenous glutathione improves salinity stress tolerance in rice (Oryza sativa L.). Plant Gene Trait 8:1–17
IPCC (2014) Climate change 2014: impacts, adaptation, and vulnerability. https://www.ipcc.ch/report/ar5/wg2/ Accessed 15 Nov 2018
Islam AR, Shen S, Hu Z, Rahman MA (2017) Drought hazard evaluation in boro paddy cultivated areas of western Bangladesh at current and future climate change conditions. Adv Meteorol 2017, 3514381
Jagota SK, Dani HM (1982) A new calorimetric technique for the estimation of Vitamin C using Folin phenol reagent. Anal Biochem 127:178–182
Kaya C, Tuna L, Higgs D (2006) Effect of silicon on plant growth and mineral nutrition of maize grown under water-stress conditions. J Plant Nutr 29:1469–1480
Khoury CK, Bjorkman AD, Dempewolf H, Ramirez-Villegas J, Guarino L, Jarvis A, Rieseberg LH, Struik PC (2014) Increasing homogeneity in global food supplies and the implications for food security. Proc Natl Acad Sci 111:4001–4006
Kim YO, Bae HJ, Cho E, Kang H (2017) Exogenous glutathione enhances mercury tolerance by inhibiting mercury entry into plant cells. Front Plant Sci 8:683
Krasensky J, Jonak C (2012) Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J Exp Bot 63:1593–1608
Li Y, Zhang S, Jiang W, Liu D (2013) Cadmium accumulation, activities of antioxidant enzymes, and malondialdehyde (MDA) content in Pistia stratiotes L. Environ Sci Pollut Res 20:1117–1123
Lichtenthaler HK (1987) Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol 148:350–382
Lisar YSS, Motafakkerazad R, Hossain MM, Rahman IMM (2012) Water stress in plants: causes, effects and responses. In: Rahman IMM (ed) Water stress, 1st edn. IntechOpen, Croatia, pp 1–14
Liu F, Jensen CR, Andersen MN (2005) A review of drought adaptation in crop plants: changes in vegetative and reproductive physiology induced by ABA-based chemical signals. Aust J Agric Res 56:1245–1252
Liu W, Sun F, Lim WH, Zhang J, Wang H, Shiogama H, Zhang Y (2018) Global drought and severe drought-affected populations in 1.5 and 2°C warmer worlds. Earth Syst Dyn 9:267–283
Maathuis FJM (2014) Sodium in plants: perception, signaling, and regulation of sodium fluxes. J Exp Bot 65:849–858
Meher Shivakrishna P, Reddy KA, Rao DM (2018) Effect of PEG-6000 imposed drought stress on RNA content, relative water content (RWC), and chlorophyll content in peanut leaves and roots. Saudi J Biol Sci 25:253–258
Mignolet-Spruyt L, Xu E, Idänheimo N, Hoeberichts FA, Mühlenbock P, Brosché M, Van Breusegem F, Kangasjärvi J (2016) Spreading the news: subcellular and organellar reactive oxygen species production and signalling. J Exp Bot 67:3831–3844
Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ 33:453–467
Nahar K, Gretzmacher R (2002) Effect of water stress on nutrient uptake, yield and quality of tomato (Lycopersicon esculentum Mill.) under subtropical conditions. Die Bodenkultur 53:45–51
Nahar K, Hasanuzzaman M, Alam MM, Fujita M (2015) Exogenous glutathione confers high temperature stress tolerance in mung bean (Vigna radiata L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environ Exp Bot 112:44–54
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22:867–880
Osakabe Y, Osakabe K, Shinozaki K, Tran LSP (2014) Response of plants to water stress. Front Plant Sci 5:86
Ouk M, Basnayake J, Tsubo M, Fukai S, Fischer KS, Cooper M, Nesbitt H (2006) Use of drought response index for identification of drought tolerant genotypes in rainfed lowland rice. Field Crop Res 99:48–58
Pei L, Che R, He L, Gao X, Li W, Li H (2019) Role of exogenous glutathione in alleviating abiotic stress in maize (Zea mays L.). J Plant Growth Regul 38:199–215
Rahman MR, Lateh H (2017) Climate change in Bangladesh: a spatio-temporal analysis and simulation of recent temperature and rainfall data using GIS and time series analysis model. Theor Appl Climatol 128:27–41
Sabetta W, Paradiso A, Paciolla C, de Pinto MC (2017) Chemistry biosynthesis, and antioxidative function of glutathione in plants. In: Hossain MA, Mostofa MG, Diaz-Vivancos P, Burritt DJ, Fujita M, Tran LSP (eds) Glutathione in plant growth, development, and stress tolerance, 1st edn. Springer, Basel, pp 1–27
Salehi-Lisar SY, Bakhshayeshan-Agdam H (2016) Drought stress in plants: causes, consequences, and tolerance. In: Tran LS, Burritt DJ, Bhattacharjee S, Wani SH, Hossain MA (eds) Drought stress tolerance in plants, 1st edn. Springer, Basel, pp 1–16
Salekjalali M, Jafari B (2011) Analysis of antioxidant enzyme activity during reproductive stages of barley under drought stress. J Ecobiotechnol 3:40–47
Schmid RW, Reilley CN (1957) New complexon for titration of calcium in the presence of magnesium. Anal Chem 29:264–268
Schmidhuber J, Tubiello FN (2007) Global food security under climate change. Proc Natl Acad Sci 104:19703–19708
Seminario A, Song L, Zulet A, Nguyen HT, González EM, Larrainzar E (2017) Drought stress causes a reduction in the biosynthesis of ascorbic acid in soybean plants. Front Plant Sci 8:1–10
Sharma P, Dubey RS (2005) Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul 46:209–221
Silva EC, Nogueira RJMC, Silva MA, Albuquerque M (2011) Drought stress and plant nutrition. Plant Stress 5(1):32–41
Singleton VL, Rossi JR (1965) Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic 16:144–158
Sohag AA, Tahjib-Ul-Arif M, Brestič M, Afrin S, Sakil MA, Hossain MT, Hossain MA, Hossain MA (2020a) Exogenous salicylic acid and hydrogen peroxide attenuates drought stress in rice. Plant Soil Environ 66:7–13
Sohag AAM, Tahjib-Ul-Arif M, Afrin S, Khan MK, Hannan MA, Skalicky M, Mortuza MG, Brestic M, Hossain MA, Murata Y (2020b) Insights into nitric oxide-mediated water balance, antioxidant defence and mineral homeostasis in rice (Oryza sativa L.) under chilling stress. Nitric Oxide. https://doi.org/10.1016/j.niox.2020.04.001
Stagnari F, Galieni A, Pisante M (2016) Drought stress effects on crop quality. In: Ahmad P (ed) Water stress and crop plants: a sustainable approach, 1st edn. Wiley, Singapore, pp 422–451
Tahjib-Ul-Arif M, Sayed MA, Islam MM, Siddiqui MN, Begum SN, Hossain MA (2018a) Screening of rice landraces (Oryza sativa L.) for seedling stage salinity tolerance using morpho-physiological and molecular markers. Acta Physiol Plant 40:1–12
Tahjib-Ul-Arif M, Siddiqui MN, Sohag AA, Sakil MA, Rahman MM, Polash MA, Mostofa MG, Tran LS (2018b) Salicylic acid-mediated enhancement of photosynthesis attributes and antioxidant capacity contributes to yield improvement of maize plants under salt stress. J Plant Growth Regul 37:1318–1330
Tahjib-Ul-Arif M, Roy PR, Sohag AAM, Afrin S, Rady MM, Hossain MA (2018c) Exogenous calcium supplementation improves salinity tolerance in BRRI dhan28; a salt-susceptible high-yielding Oryza sativa cultivar. J Crop Sci Biotechnol 21:383–394
Tahjib-Ul-Arif M, Afrin S, Polash MAS, Akter T, Ray SR, Hossain MT, Hossain MA (2019a) Role of exogenous signaling molecules in alleviating salt-induced oxidative stress in rice (Oryza sativa L.): a comparative study. Acta Physiol Plant 41(5):69
Tahjib-Ul-Arif M, Sohag AAM, Afrin S, Bashar KK, Afrin T, Mahamud AGM, Polash MAS, Hossain M, Sohel M, Taher A, Brestic M (2019b) Differential response of sugar beet to long-term mild to severe salinity in a soil–pot culture. Agriculture 9:223
Taïbi K, Taïbi F, Abderrahim LA, Ennajah A, Belkhodja M, Mulet JM (2016) Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defense systems in Phaseolus vulgaris L. S Afr J Bot 105:306–312
Wang F, Chen F, Cai Y, Zhang G, Wu F (2011) Modulation of exogenous glutathione in ultrastructure and photosynthetic performance against Cd stress in the two barley genotypes differing in Cd tolerance. Biol Trace Elem Res 144:1275–1288
Wang W, Ertsen MW, Svoboda MD, Hafeez M (2016) Propagation of drought: from meteorological drought to agricultural and hydrological drought. Adv Meteorol 2016, 6547209
Wang X, Liu H, Hu B, Jia Y, Sha H, Zhao H (2019) Diferential activity of the antioxidant defence system and alterations in the accumulation of osmolyte and reactive oxygen species under drought stress and recovery in rice (Oryza sativa L.) tillering. Sci Rep 9:8543
Waśkiewicz A, Muzolf-Panek M, Goliński P (2013) Phenolic content changes in plants under salt stress. In: Ahmad P, Azooz MM, Prasad MN (eds) Ecophysiology and responses of plants under salt stress, 1st edn. Springer, New York, pp 283–314
Xin ZL, Mei G, Shiqing L, Shengxiu L, Zongsuo L (2011) Modulation of plant growth, water status and antioxidantive system of two maize (Zea mays L.) cultivars induced by exogenous glycinebetaine under long term mild drought stress. Pak J Bot 43:1587–1594
Zhang Z, Huang R (2013) Analysis of malondialdehyde, chlorophyll proline, soluble sugar, and glutathione content in arabidopsis seedling. Bio-Protocol 3:e817
Zhou Y, Wen Z, Zhang J, Chen X, Cui J, Xu W, Liu HY (2017) Exogenous glutathione alleviates salt-induced oxidative stress in tomato seedlings by regulating glutathione metabolism, redox status, and the antioxidant system. Sci Hortic 220:90–101
Zhou Y, Diao M, Cui JX, Chen XJ, Wen ZL, Zhang JW, Liu HY (2018) Exogenous GSH protects tomatoes against salt stress by modulating photosystem II efficiency, absorbed light allocation and H2O2-scavenging system in chloroplasts. J Integr Agric 17:2257–2272
Acknowledgements
The authors gratefully acknowledge the constructive suggestions of Professor Md. Abdul Hannan, Department of Biochemistry and Molecular Biology, Bangladesh Agricultural University, during manuscript preparation. The authors also thankful to Md Kawsar Khan, Department of Biochemistry and Molecular Biology, Shahjalal University of Science and Technology, Sylhet-3114, Bangladesh for his valuable time for making PCA graph.
Author information
Authors and Affiliations
Contributions
Conceptualization: MT-U-A, MAH; Methodology: AAMS, MT-U-A; Formal analysis and investigation: AAMS, MT-U-A, MASP, SA; Writing-original draft preparation: AAMS, MBC; Writing-review and editing: MT-U-A, DJB, YM, MAH; Funding acquisition: MAH, MT-U-A; Resources: MT-U-A; Supervision: MT-U-A.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Sohag, A.A.M., Tahjib-Ul-Arif, M., Polash, M.A.S. et al. Exogenous Glutathione-Mediated Drought Stress Tolerance in Rice (Oryza sativa L.) is Associated with Lower Oxidative Damage and Favorable Ionic Homeostasis. Iran J Sci Technol Trans Sci 44, 955–971 (2020). https://doi.org/10.1007/s40995-020-00917-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40995-020-00917-0