
Abstract
Testosterone (T) and cortisol (C) are hormones related to, respectively, dominance motivation and stress. Levels of both hormones increase in association with athletic competition. Estradiol (E) has been linked to dominance motivation in women, but has heretofore not been studied in the context of athletic competition. In this study, salivary levels of T, C, and E were determined for women soccer players to obtain a neutral-day baseline. Later, changing levels of these same hormones were tracked across the course of two intercollegiate matches, one played at home (a loss) and the other played away (a win). Before warm-up levels of C and E (but not T) were elevated relative to neutral-day baseline. T and E (but not C) increased during warm-up. T and C increased during the competition, while E declined to baseline. During the 30-min period after the end of competition, T decreased while C remained elevated. Pre-warm-up, warm-up, and competition-related changes in T, C, and E were not dependent on match venue (home vs away) or match outcome (win vs loss). T and C decreased more within the 30 min after the game that was won compared to the game that was lost. Over the course of athletic competition, levels of T, C, and E change differentially in relation to each other in remarkably reliable and predictable patterns across individuals and sports contexts. These findings should encourage study of the ways in which the interplay of these hormonal changes influence and/or is influenced by the psychological experience of competition.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Competition, in human social contexts, is a contest for status. What motivates the act of competing is, at its most fundamental level, a psychological drive to win or be better than one’s opponent. This drive appears to have a neuroendocrine basis. Testosterone (T) is a steroid hormone secreted in abundance from the testes and in lesser amounts by the ovaries and adrenal cortex. High levels of T are associated with high implicit power/dominance motivation and social status in men (for reviews, Eisenegger et al. 2011; Mazur and Booth 1998; Stanton and Schultheiss 2009), and social interactions in which status is contested can transiently increase T levels (e.g., Costa and Salvador 2012; van der Meij et al. 2010; for review, Carré and Olmstead 2015). Cortisol (C) is a glucocorticoid hormone, secreted from the adrenal glands, that is elevated in response to physical and psychological stress (for review, Dickerson and Kemeny 2004). Men who have relatively higher baseline T and/or higher implicit power motivation show increased C across a dominance contest that results in defeat (Mehta et al. 2008; Wirth et al. 2006). Additionally, relatively high levels of C prior to a dominance contest appear to reduce or eliminate the relationship between T and dominance-related behavior (Mehta and Josephs 2010) and status among teammates in women athletes (Edwards and Casto 2013). Estradiol (E) is a steroid hormone secreted from the testes in men and in larger quantities from the ovaries in women. E is primarily derived from the gonadal aromatization of T, but, aromatization of T to E also occurs in other peripheral and central locations, raising the possibility that many of the effects attributed to circulating T may actually result from the action of E derived from locally aromatized T (for review, Naftolin 1994). In women, baseline levels of E prior to engaging in a contest for status have been positively related to social status/implicit power motivation (Stanton and Schultheiss 2007).
Athletic competition is a formal contest for status requiring sport-dependent degrees of physical exertion and team coordination and offers an important, real-world setting for the study of the neuroendocrinology of social competition. For serious athletes, a competition is both meaningful and authentic – where performing well and match outcome are of great personal and team significance, often bearing consequence for status among one’s teammates and divisional or national team rankings. In a variety of sports, athletic competition is associated with a substantial increase in both T and C in men and women (e.g., Bateup et al. 2002; Casto et al. 2014; Edwards et al. 2006; Edwards and Kurlander 2010; Filaire et al. 2009; Gonzalez-Bono et al. 1999; Hamilton et al. 2009; Suay et al. 1999). How levels of E relate to athletic competition has never been studied. But, given the relationship between E and power motivation in women, this information would seem necessary for a comprehensive understanding of endocrine responses to competition. Further, a more thorough appreciation of the endocrine effects of competition would be aided by a study of how T, C, and E, hormones which are physiologically and mechanistically linked, change in relation to one another across the course of competition.
A number of factors merit special consideration in the discussion of hormone reactivity to competition. First, some previous studies have shown that T levels only increase across a competition that is won (“the winner effect,” e.g., Booth et al. 1989; Jiménez et al. 2012; Oliveira et al. 2009) and C levels only increase across a competition that is lost (e.g., Jiménez et al. 2012). Other research with men and women athletes shows a “competition effect” for T and C regardless of match outcome, provided the after-competition sample is taken immediately after the end of play (e.g., Casto et al. 2014; Edwards et al. 2006; Edwards and Kurlander 2010; Gonzalez-Bono et al. 1999). Inconsistencies may have to do with variability in when saliva samples are collected relative to the actual period of competition, ranging from 1 to 2 h before and after competition to immediately before and after. Given the multitude of psychological and physical stressors within the hour leading up to competition (e.g., pre-competition interactions with opponents, teammates and coaches and warm-up) and the hour following competition (e.g., the psychological effects of winning and losing, the dramatic decrease in the level of physical activity), it seems imperative that researchers specifically parse out separate effects due to the warm-up period, actual competition, and the after-competition period when the winners and losers have been officially determined. Indeed, initial studies on the matter indicate that warm-up and after-competition time periods have hormone changes outside of (but undoubtedly connected to) the actual competition. In apparently the only two studies of the matter, Edwards and Kurlander (2010) and Casto et al. (2014) show that for women athletes, the warm-up period is associated with an increase in T, but not C. Following the increase in both T and C during competition, levels of these hormones may quickly fall within the hour after competition (Filaire et al. 2009; Kivlighan et al. 2005). Given that the time within the hour after the end of a competition is when the psychological effects of winning and losing are most keenly felt – a time when the effects of victory/defeat on hormone levels should be most clearly evident – further study of the change in hormone levels during this period seems warranted.
Sex of participants and competitive context likely have an important influence on hormone reactivity to competition. Studies involving athletic and non-athletic contests have shown that although T increases across competition in men, T may remain stable or even decrease across competition in women (e.g., Carré et al. 2013; Kivlighan et al. 2005; Mazur 1997). Kivlighan et al. (2005), for example, did not find a competition effect for T in women, but in this study, the competition (a 2000 m row ergometer race) was staged among members of women’s sports team rather than against an actual opponent. The complex dynamics and contextual factors of simulated competition, existing status structures among teammates, and competition among otherwise cooperative partners complicate the interpretation of the results of this study. Additionally, in other studies where women did not have a T response to competition (e.g., video game boxing and volleyball, Carré et al. 2013; video game ping-pong, Mazur 1997), competition was simulated in a laboratory and conducted through a computer interface against a virtual competitor. This environment or type of competition may not have been sufficient to motivate competitive drive in a random sample of undergraduate women and/or may not have been a situation that allowed these women to compete in a task in which they felt sufficiently self-efficacious.
Another factor relevant to understanding the endocrine response to athletic competition is match venue. For male athletes, pre-game T (Carré et al. 2006; Neave and Wolfson 2003) and C (Carré et al. 2006) levels have been shown to be higher when competing at home than when competing away from home. Additionally, Carré (2009) showed that the increase in T from before to after competition was greater when playing at home than away. But, the endocrine correlates of competing at home versus away have not been studied in women.
The use of oral contraceptives (OCs) may affect hormone reactivity to competition. Although OC use is associated with significantly lower T in women athletes at baseline (Bermon et al. 2014; Edwards and O’Neal 2009), both OC and non-OC using athletes show similar increases in T across competition (Edwards and O’Neal 2009). However, a recent report that tracked T change across several competitions and training sessions in the same women showed that OC users had a dampened T increase compared to non-OC users when values were pooled across all activities (Crewther et al. 2015). The effect of OC use on hormonal responses to the anticipation for competition, warm-up, and the after competition period have not been determined, but OC use could be an important moderating factor relevant to hormone reactivity to the different phases of athletic competition.
The present study was designed to explore changes in salivary levels of T, C, and E from a neutral-day baseline to pre-competition baseline and across warm-up, competition, and 30 min after competition for two intercollegiate intra-conference matches, one played at home and one played away from home. This study is the first to explore, together, T, C, and E reactivity to athletic competition and, additionally, allows for analysis of changes in hormone levels leading up to warm-up, during warm-up, and the 30 min following competition. By separating matches based on venue, this study is the first to explore potential differences for women in pre-match hormone levels related to competing at home and away. Data regarding OC use was collected and OC use was included as a potential moderator in all analyses.
Method
Participants
Participants for the study were the twenty-five consenting members (age 18–22) of the 2013 Emory University varsity women’s soccer team. This research was approved by Emory’s Institutional Review Board and athletes gave written informed consent prior to participation. As part of the consent procedure women were asked to respond “yes” or “no” to the question “Are you currently using an oral contraceptive?” and to one other: “Are you currently using any injected, implanted, or patch-delivered hormone contraceptive?”
Procedure
Nine saliva samples were obtained from each participant. Participants gave one baseline sample at the time of consent on a neutral day, 3 days before the first match. Then, participants gave four samples in association with each of two intra-conference NCAA soccer competitions, 1 week apart. All samples were collected between 2 and 4 PM. The first of the two matches (a 0–1 loss) was played at the participants’ home field against Washington University and the second game (a 2-0 victory) was played away from home against the University of Chicago. For each match, participants gave a saliva sample 10–15 min before the start of an hour-long warm-up, another sample immediately after warm-up/a few minutes before the start of the match, a third sample immediately after match completion, and a final sample 30 min later.
Saliva Samples and Hormone Assays
Participants were provided with a piece of sugar-free gum (Trident®, original flavor) to stimulate saliva production and they gave samples by passive drool exactly according to the protocol used for athletes in other sports (e.g., Edwards and Casto 2013).
Samples collected at baseline and during the home game were chilled on ice after collection and then stored at -80 °C within the hour. Samples collected at the field in Chicago were packed and frozen in dry ice (~-80 °C) on site and delivered to storage with the home game samples at -80 °C within 7 h after the match. Samples were assayed in duplicate for T, C, and E on a single thaw by the Biomarkers Core laboratory of the Yerkes Primate Center (Atlanta, GA) using competitive enzyme immunoassay kits from Salimetrics (State College, PA). The average intra-assay CV percents for T, C, and E were 6.1 and 7.2 and 8.65 %, respectively. The average inter-assay CV percents for T, C, and E were 8.1, 6.26 and 7.4 %, respectively.
Statistical Analyses
The SPSS statistical package was used for calculation of independent and repeated measures t-tests (two-tailed) and mixed ANOVA with within-subjects post-hoc repeated contrasts for the changes in hormone levels across time by OC use for each match. In all cases, p ≤ 0.05 was required for statistical significance. Effect sizes, Cohen’s d (d) and partial eta squared (η2), were calculated for all t-tests and ANOVAs, respectively. There were no apparent outliers in the data and, except for one woman whose baseline E sample was non-detectable, all data are included for analysis. Log-transformed data produce the same results as the raw data. Thus, in keeping with procedures in other descriptive reports of hormone levels associated with athletic competition (e.g. Casto et al. 2014; Edwards et al. 2006; Edwards and Kurlander 2010), raw data are used for all figures and statistical analyses in this report.
Results
Although 25 participants gave saliva samples at neutral-day baseline, only 17 would go on to play in the home game and only 15 would go on to play in the away game. For the home game, the eight participants who did not play warmed up with the team, but stayed on the sidelines throughout competition. For the away game, six athletes who gave saliva at baseline did not travel to the game with the team (and thus, did not provide any additional saliva samples). Four participants traveled to the game and warmed up with the team, but stayed on the sidelines throughout competition. Although non-playing athletes (those who watched the match from the sidelines) showed increases in T and E associated with warm-up (33 and 32 %, respectively), average levels of both hormones decreased from before to after competition (Fig. 1). Levels of C in these athletes did not change throughout the course of warm-up or competition (Fig. 1). Due to the small and unequal (relative to those who played) number of athletes that did not play in each game, these athletes were excluded from subsequent statistical analyses.

Testosterone, Cortisol, and Estradiol levels at baseline, before warm-up, after warm-up, and after competition by OC use for two games, a home loss and an away win. Errors bars represent 95 % confidence intervals
Of the 17 women that would go on to play in one or both matches, 11 were OC users and 6 were not using any form of hormonal contraceptive (non-OC users). At neutral-day baseline, the OC users had significantly lower T (M = 47.43 pg/ml, SE = 3.50) than the non-OC users (M = 62.28 pg/ml, SE = 5.74), t(15) = 2.35, p = .033, d = 1.15. C and E levels were not significantly different between OC users and non-OC users at baseline.
Hormone Levels by Time and OC Use for Each Game
A 5 × 2 mixed ANOVA was used to compare hormone levels from neutral-day baseline to before warm-up, from before to after warm-up, from after warm-up to after competition, and from after competition to 30 min. after competition for OC users and non-users. The model included the within-subjects factor of Time (baseline, before warm-up, after warm-up, after competition, and 30 min. after competition), the between-subjects factor of OC use, and the Time x OC use interaction predicting T, C, and E, separately. Post-hoc repeated contrasts for time were conducted and all post-hoc statistical results are shown in Table 1. These analyses were conducted for each of the two matches (Home-Loss and Away-Win) separately. Levels of T, C, and E by time, OC use, and match are shown in Fig. 1.
Testosterone
For T, there was a significant main effect for Time in both matches (Home-Loss: F (4, 60) = 12.81, p < .001, η2 = .46; Away-Win: F (4, 52) = 24.26, p < .001, η2 = .65). Post-hoc repeated contrasts revealed that, across both matches, before warm-up T levels were not significantly changed from neutral day baseline. In both matches, however, T levels significantly increased from before to after warm-up (Table 1) and from after warm-up to immediately after competition (Table 1). The magnitude of the warm-up and competition effects on T are quite large – increases in T across warm-up occurred in 97 % of the instances in both matches with an average increase of 27 % and continued to increase across competition in 81 % of the instances across both matches with an average additional increase of 18 % from after warm-up level. Although the percent increase in T during the Home-Loss and Away-Win are equivalent (19 and 18 %, respectively), the effect size for T is more than double for the Away-Win (Table 1). Increases in T during competition were not correlated with the number of minutes an athlete played or with her playing position. After competition, T levels significantly decreased during the 30 min after the end of play in both matches (Table 1). T levels were lower in OC users than non-users in both matches at every time point sampled (Fig. 1). Across all time points associated with the Away-Win, OC users had significantly lower T levels than non-users, F (1, 13) = 5.15, p = .041, η2 = .28. OC users did not have significantly lower T than non-users across time points associated with the Home-Loss, F (1, 15) = 2.37, p = .144, η2 = .14. Post-hoc independent t-tests for the Away-Win revealed that OC users had significantly lower T than non-users for the after-warm up (t(13) = 2.26, p = .041, d = 1.06) and the 30-min-after-competition (t(13) = 2.48, p = .028, d = 1.43) comparisons and although the after competition difference was not statistically significant, the effect size was quite large (t(13) = 1.89, p = .081, d = 1.0). The OC use by Time interaction was not significant for either match.
Cortisol
For C, there was a significant main effect for Time in both matches (Home-Loss: F (4, 60) = 13.84, p < .001, η2 = .48; Away-Win: F (4, 52) = 7.26, p < .001, η2 = .36). Post-hoc repeated contrasts showed that before-warm-up C levels were significantly elevated relative to neutral-day baseline for the home match (Table 1) and elevated, but not significantly, for the away match (Table 1), although the magnitude of the effect is large to moderate in both cases. From neutral-day baseline to before warm-up for both games, C levels increased in 80 % of the athletes who would go on to play, with the average increases being 55 % and 53 % for the home and away matches, respectively. C levels remained relatively stable from before to after warm-up. But in both matches, C levels substantially increased during the period of competition (Table 1). For the Home-Loss, but not the Away-Win, OC users showed a greater increase in C from before to after competition compared to non-users (F (1, 15) = 4.38, p = .054, η2 = .23). Competition-related increases occurred in 91 % of the instances, with an average increase of 108 % across both matches. With the exception of the goal-keeper whose C levels decreased during the period of competition for both matches, the percent increases in C levels were not related to player position or number of minutes played. C levels did not significantly change from immediately after the end of competition to 30 min after competition following either match. There was no overall difference in C levels based on OC use.
Estradiol
For E, there was a significant main effect for Time in both matches (Home-Loss: F (4, 56) = 5.01, p = .002, η2 = .26; Away-Win: F (4, 44) = 10.86, p < .001, η2 = .50). Post-hoc repeated contrasts showed that before-warm-up E levels were significantly elevated relative to neutral-day baseline for the Home-Loss (Table 1). For the Away-Win, before warm-up E levels were also elevated, but the effect was not significant. Despite that, effects sizes for pre-warm-up increases for both matches were identical (Table 1). In both matches, levels of E were significantly elevated from before to after warm-up (Table 1). This warm-up-associated increase in E occurred in 91 % of the instances across both matches with an average increase of 27 %. In both matches, E levels were significantly decreased from after warm-up to after competition (Table 1), and remained stable at this new level for the 30 min following competition. There was no overall difference in E levels based on OC use. Additionally, the interaction between Time and OC use was not statistically significant.
Between Game Comparisons
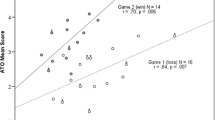
Although 17 women played in the Home-Loss and 15 women played in the Away-Win, only 14 women played in both games. For these 14 women, mean levels of T, C, and E, at any time point sampled, were not significantly different for the Home-Loss compared to the Away-Win. However, when expressed as a percent change, after competition differences between the Home-Loss and Away-Win in T and C were significant. Specifically, although T levels declined after both matches, the percent decrease in T from immediately after competition to 30 min. after competition was significantly greater for the Away-Win (M = -26 %, SE = 2 %) than the Home-Loss (M = −17 %, SE = 3 %), t(13) = 3.27, p = .006, d = 0.91. C levels remained relatively stable within the 30 min. after the Home-Loss (M = 3 %, SE = 6 %), but decreased after the Away-Win (M = −13 %, SE = 6 %). The mean difference between these after-competition percent changes in C is significant, t(13) = 3.86, p = .002, d = 1.03. Changes in T, C, or E associated with the stage from baseline to before warm-up, warm-up, and competition were not statistically different between the Home-Loss and Away-Win.
Discussion
Consistent with previous reports (e.g., Alix-Sy et al. 2008; Bateup et al. 2002; Filaire et al. 2009; Salvador et al. 2003), pre-warm-up levels of C on the day of competition were higher relative to neutral-day baseline. Additionally, this study offers new evidence of a similar, match-day anticipatory elevation effect in E. Although thought to reflect athlete performance anxiety (e.g., Eubank et al. 1997), levels of pre-competition C have not previously been found to be correlated with levels of anxiety or negative emotions (Alix-Sy et al. 2008; Filaire et al. 2009). Higher pre-warm-up, pre-competition C and E levels could represent an adaptive anticipatory physiological response to the physical and or psychological stress of the competition to come. Although other reports have provided evidence for a game-day anticipatory rise in T for men (Booth et al. 1989; Salvador et al. 2003), before warm-up levels of T for women athletes in the present study were not elevated relative to neutral-day baseline.
Warm-up prepares the athlete’s mind and body for the competition to come. In this study, levels of T and E (but not C) significantly increased during warm-up for both competitions. The increase in T appears to begin with warm-up; the increase in E continues the process begun some time earlier. These results provide the first demonstration of a warm-up effect on E in women athletes preparing for competition, and confirm, for women soccer players, findings of a “warm-up effect” for T in women cross country runners, volleyball, and tennis players (Casto et al 2014; Edwards and Kurlander 2010). The warm-up period often provides an opportunity for athletes to watch and be watched by their opponents and is time when athletes often engage in behaviors that signal strength and confidence. Warm-up related changes in levels of gonadal hormones may influence or be influenced by both overt and covert competitive/dominant behaviors prior to competition and/or feelings of self-confidence. Additionally, social interactions among teammates and coordinated efforts during warm-up may influence or be influenced by T and/or E increases associated with warm-up.
Competition was associated with a significant increase in salivary levels of C and T (but not E) for each of the two matches. This “competition effect” for T and C is, by now, well-demonstrated in a variety of men’s and women’s sports (e.g., Bateup et al. 2002; Edwards et al. 2006; Edwards and Kurlander 2010; Casto et al. 2014; Suay et al. 1999). Importantly, it matters that an athlete actually participate in the competition – athletes who do not play, but watch the game from the sidelines, do not show increases in T or C associated with competition. From the already substantially elevated T levels after warm-up, the additional percent increase in T was virtually the same for the Home-Loss (19 %) compared to the Away-Win (18 %). Although the percent increase was equivalent in both matches, the effect size of the T increase was more than double for the match that was won. However, if there were additional T increases due to winning, they were short-lived – in both the game that was lost and the game that was won, levels of T significantly declined within the 30 min after competition. In fact, T levels decreased significantly more after the match that was won than the match that was lost. Previous studies that sample before and after competition T at variable lengths from the actual period of competition have suggested that T levels only increase across a competition that is won (“the winner effect,” e.g., Booth et al. 1989; Jiménez et al. 2012; Oliveira et al. 2009). However, provided that saliva samples are collected immediately after the end of competition, we find that competition is associated with substantial elevations in T in victory and defeat. Unlike T, C levels remained relatively elevated for at least 30 min after both a win and a loss. But, when expressed as a percent change, levels of C were significantly decreased during the 30 min following victory compared to the match that ended in defeat. A decrease in C levels after an athletic victory compared to defeat has been found in other studies (e.g., Filaire et al. 2009; Jiménez et al. 2012) and may reflect a relief or reduction in psychological stress or anxiety associated with winning compared to losing.
Previous studies with men athletes have reported that T and C may be higher before competing at one’s home venue compared to competing away from home (e.g., Neave and Wolfson 2003; Carré et al. 2006) and that the change in T associated with competition may be greater when competing at home (Carré 2009). In the present study, for women, levels of T, C, and E were no different before warm-up, after warm-up, or after competition when competing at home compared to away. Additionally, the percent changes in T, C, and E from neutral-day baseline and those associated with warm-up and competition were not significantly different when playing at home compared to away. Although this study is limited in that it only samples one home and one away match, we do not find evidence of a venue effect on women athletes.
Causes and Effects of Sport-Related Hormone Change
Increases in T and E during warm-up and T and C during competition could reflect the psychological anticipation for or experience of competition, the physical exertion of sport, or some combination of the two. Non-competitive exercise can increase T, C, and E in blood (Copeland et al. 2002; Farrell et al. 1983; Wilkerson et al. 1980). Changes in blood or salivary hormone concentrations are often thought to reflect new production and secretion of these hormones via activation of the hypothalamic-pituitary-gonadal (HPG) or hypothalamic-pituitary-adrenal (HPA) axes. However, exercise can produce a decrease in blood and saliva fluid volume as water shifts to interstitial and intracellular compartments of muscles and tissues and is lost through mechanisms of dehydration (Kargotich et al. 1998). Thus, exercise-related increases in hormone levels in blood could represent only an inherent increase in hemoconcentration. In this case, vascular fluid shifts/dehydration should affect levels of T, C, and E equally. However, warm-up, which includes at least moderate levels of physical exertion, resulted in dramatic increases in T, moderate to large increases in E, and no change in C. Further, T and C increased during competition by vastly different amounts (T = 17 % and C = 108 %) and E actually decreased. Additionally, the changes in T and C associated with competition in this study were not correlated with playing position or to the number of minutes an athlete played in either match. That T, C, and E change differently across the various phases of athletic competition and that changes are not related to indirect metrics of physical exertion suggests that there are multiple, hormone-specific, psychological and physical factors that affect competition-related changes in hormone levels. Regardless of the direct cause, changes in salivary concentrations of hormones presumably reflect increased delivery of these physiologically and psychologically relevant hormones to muscle and other tissues including the central nervous system (Vining and McGinley 1987).
A rise in levels of T, C, or E during warm-up or athletic competition may have physiological and/or psychological benefits for athletes. Transient changes in endogenous T may increase feelings of dominance, power, or competitiveness (for review, Carré and Olmstead 2015; Eisenegger et al. 2011). In men, increased T in response to watching a four-minute aggressive or training motivational video was positively related to heavier lifting during a resistance training exercise (Cook and Crewther 2012). T changes associated with a competitive task may also increase willingness to engage in a subsequent competition and/or a person’s dominance motivation (Carré and McCormick 2008; Mehta and Josephs 2006).
It is likely that increases in C associated with competition reflect the physiological and/or psychological stress of sport. Short-term elevations in C increase glucose levels in blood (thereby supplying muscle tissue with energy) and reduce inflammation in muscle and other tissues (Coutinho and Chapman 2011). Administration of C has been shown to increase positive affect as well as decrease anger, anxiety, and fatigue following a cognitive task (Schlotz et al. 2008). Although beneficial for physiology during physical stress, relatively high C after contests for status may reflect the stress of losing in power motivated men (Mehta et al. 2008; Wirth et al. 2006) and relatively high before-competition C may reduce or eliminate the relationship between T and dominance/status seeking behavior (Mehta and Josephs 2010) and T reactivity to athletic competition (Edwards and Casto 2015).
Short-term increases in levels of E activate serotonin systems responsible for inducing positive affect, increase cell excitability in various brain regions, increase vasodilation of coronary arteries, and stimulate insulin production (for review, Gruber et al. 2002; Lasiuk and Hegadoren 2007; Woolley 2007). E may moderate the effect of oxytocin, a hormone known for its association with social bonding (for review, McCarthy 1995). Thus, E could play a role in enabling the social affiliation effects of oxytocin, augmenting the feelings of social connectedness among teammates prior to a competition in which their coordinated efforts will be essential for optimum team performance.
OC Use
In agreement with previous literature (Edwards & O’Neal, 2009), OC users had generally lower salivary levels of T than non-users at neutral-day baseline and at all game-day points of saliva collection, but showed similar T reactivity to competition. Additionally, C levels increased more across competition for OC users than for non-users for one of the two matches. Despite the limited and inconsistent reports of the effects of OC use on actual athletic performance (for review, Burrows and Peters 2007), some researchers have actually recommended OC use in women athletes as a means of having a “predictable hormonal milieu for training and competition” (Frankovich and Lebrun 2000, p. 267). However, whether or not OC use compromises the psychological experience associated with warming-up or competition is not known. The associations between T and status, dominance motivation, and competitive drive (e.g., Edwards and Casto 2013; Mehta and Josephs 2006; Stanton and Schultheiss 2009) indeed raise concern for the use of hormone contraceptives in competitive sports. Lower T in preparation for or during competition could present a psychological as well as physiological disadvantage for athletes using hormonal contraceptives. More research on this matter is both critical and obligatory.
Conclusions
Collectively, results from this study provide new information regarding the differential changes in levels of psychologically-relevant hormones as women anticipate, prepare for, and compete in athletic settings. Specifically, findings from this study support previous reports of an anticipatory rise in C prior to warming-up for competition. Further, results of this study provide new evidence for a “warm-up effect” for E in women and provide confirming evidence of a similar effect for T, both of which appear independent of match venue. Results also confirm a “competition effect” for T and C in women regardless of outcome or match venue, in a competitive context in which the competitors are sufficiently motivated and skilled. T levels declined within the 30 min following competition, whereas C levels remained relatively high. When expressed as a percent change, T and C decreased more after a competition that was won compared to a competition that was lost.
Increases in T associated with warm-up and/or competition (a contest for both physical and social status) may motivate competitive drive and dominance seeking behaviors as well as prepare an athlete for the physical demands of athletic competition. Increases in C before warm-up relative to neutral day baseline and increases in C associated with competition itself could recruit important physiological resources required for optimum performance. By observing T, C, and E across the pre-warm-up anticipatory phase, warm-up, competition, and after competition we identify four temporally, physically, and hormonally distinct phases/stages of competition. The relationships between specific hormonal changes associated with each of these phases and relevant psychological experiences remain to be determined.
References
Alix-Sy, D., Le Scanff, C., & Filaire, E. (2008). Psychophysiological responses in the pre-competition period in elite soccer players. Journal of Sports Science and Medicine, 7, 446–454.
Bateup, H. S., Booth, A., Shirtcliff, E. A., & Granger, D. A. (2002). Testosterone, cortisol, and women’s competition. Evolution and Human Behavior, 23, 181–192.
Bermon, S., Garnier, P. Y., Lindén Hirschberg, A., Robinson, N., Giraud, S., Nicoli, R., & Ritzen, M. (2014). Serum androgen levels in elite female athletes. Journal of Clinical Endocrinology and Metabolism, 99, 4328–4335.
Booth, A., Shelly, G., Mazur, A., Tharp, G., & Kittok, R. (1989). Testosterone, and winning and losing in human competition. Hormones and Behavior, 23, 556–571.
Burrows, M., & Peters, C. E. (2007). The influence of oral contraceptives on athletic performance in female athletes. Sports Medicine, 37(7), 557–574.
Carré, J. M. (2009). No place like home: testosterone responses to victory depend on game location. American Journal of Human Biology, 21, 392–394.
Carré, J. M., & McCormick, C. M. (2008). Aggressive behavior and change in salivary testosterone concentrations predict willingness to engage in a competitive task. Hormones and Behavior, 54, 403–409.
Carré, J. M., & Olmstead, N. A. (2015). Social neuroendocrinology of human aggression: examining the role of competition-induced testosterone dynamics. Neuroscience, 286, 171–186.
Carré, J. M., Muir, C., Belanger, J., & Putnam, S. K. (2006). Pre-competition hormonal and psychological levels of elite hockey players: relationship to the “home advantage.”. Physiology & Behavior, 89, 392–398.
Carré, J. M., Campbell, J. A., Lozoya, E., Goetz, S. M. M., & Welker, K. M. (2013). Changes in testosterone mediate the effect of winning on subsequent aggressive behavior. Psychoneuroendcrinology, 38, 2034–2041.
Casto, K. V., Elliott, C. M., & Edwards, D. A. (2014). Intercollegiate cross country competition: effects of warm-up and racing on salivary levels of cortisol and testosterone. International Journal of Exercise Science, 7(4), 318–328.
Cook, C. J., & Crewther, B. T. (2012). Changes in salivary testosterone concentrations and subsequent voluntary squat performance following the presentation of short video clips. Hormones and Behavior, 61, 17–22.
Copeland, J. L., Consitt, L. A., & Tremblay, M. S. (2002). Hormonal responses to endurance and resistance exercise in females aged 19–69 years. Journal of Gerontology. Series A, Biological Sciences and Medical Sciences, 57(4), 158–165.
Costa, R., & Salvador, A. (2012). Associations between success and failure in a face-to-face competition and psychobiological parameters in young women. Psychoneuroendocrinology, 37, 1780–1790.
Coutinho, A. E., & Chapman, K. E. (2011). The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights. Molecular and Cellular Endocrinology, 335, 2–13.
Crewther, B. T., Hamilton, D., Casto, K., Kilduff, L. P., & Cook, C. J. (2015). Effects of oral contraceptive use on salivary testosterone and cortisol responses to training sessions and competitions in elite women athletes. Physiology & Behavior, 147, 84–90.
Dickerson, S. S., & Kemeny, M. E. (2004). Acute stressors and cortisol responses: a theoretical integration and synthesis of laboratory research. Psychological Bulletin, 130, 355–391.
Edwards, D. A., & Casto, K. V. (2013). Women’s intercollegiate athletic competition: cortisol, testosterone, and the dual-hormone hypothesis as it relates to status among teammates. Hormones and Behavior, 64, 153–160.
Edwards, D.A, & Casto, K.V. (2015). Baseline cortisol moderates testosterone reactivity to women’s intercollegiate athletic competition. Physiology & Behavior, 142, 48--51.
Edwards, D. A., & Kurlander, L. S. (2010). Women’s intercollegiate volleyball and tennis: effects of warm-up, competition, and practice on saliva levels of cortisol and testosterone. Hormones and Behavior, 58, 606–613.
Edwards, D. A., & O’Neal, J. L. (2009). Oral contraceptives decrease saliva testosterone but do not affect the rise in testosterone associated with athletic competition. Hormones and Behavior, 56(2), 195–198.
Edwards, D. A., Wetzel, K., & Wyner, D. R. (2006). Intercollegiate soccer: saliva cortisol and testosterone are elevated during competition, and testosterone is related to status and social connectedness with teammates. Physiology & Behavior, 87, 135–143.
Eisenegger, C., Haushofer, J., & Fehr, E. (2011). The role of testosterone in social interaction. Trends in Cognitive Neuroscience, 15(6), 263–271.
Eubank, M., Collins, D., Lovell, G., Dorling, D., & Talbot, S. (1997). Individual temporal differences in pre-competition anxiety and hormonal concentration. Personality and Individual Differences, 23(6), 1031–1039.
Farrell, P. A., Garthwaite, T. L., & Gustafson, A. B. (1983). Plasma adrenocorticotropin and cortisol responses to submaximal and exhaustive exercise. Journal of Applied Physiology, 55, 1441–1444.
Filaire, E., Alix, D., Ferrand, C., & Verger, M. (2009). Psychophysiological stress in tennis players during the first single match of a tournament. Psychoneuroendocrinology, 34, 150–157.
Frankovich, R. J., & Lebrun, C. M. (2000). The athletic woman: menstrual cycle, contraception, and performance. Clinics in Sports Medicine, 19(2), 251–271.
Gonzalez-Bono, E., Salvador, A., Serrano, M. A., & Ricarte, J. (1999). Testosterone, cortisol, and mood in a sports team competition. Hormones and Behavior, 35, 55–62.
Gruber, C. J., Tschugguel, W., Schneeberger, C., & Huber, J. C. (2002). Production and action of estrogens. New England Journal of Medicine, 346(5), 340–352.
Hamilton, L. D., van Anders, S. M., Cox, D. N., & Watson, N. V. (2009). The effect of competition on salivary testosterone in elite female athletes. International Journal of Sports Physiology and Performance, 4, 538–542.
Jiménez, M., Aguilar, R., & Alvero-Cruz, J. R. (2012). Effects of victory and defeat on testosterone and cortisol response to competition: evidence for same response patterns in men and women. Psychoneuroendocrinology, 37, 1577–1581.
Kargotich, S., Goodman, C., Keast, D., & Morton, A. R. (1998). The influence of exercise-induced plasma volume changes on the interpretation of biochemical parameters used for monitoring exercise, training and sport. Sports Medicine, 26(2), 101–117.
Kivlighan, K. T., Granger, D. A., & Booth, A. (2005). Gender differences in testosterone and cortisol response to competition. Psychoneuroendcrinology, 30, 58–71.
Lasiuk, G. C., & Hegadoren, K. M. (2007). The effects of estradiol on central serotonergic systems and its relationship to mood in women. Biological Research Nursing, 9(2), 147–160.
Mazur, A. (1997). Sex difference in testosterone response to a video game contest. Evolution and Human Behavior, 18, 317–326.
Mazur, A., & Booth, A. (1998). Testosterone and dominance in men. Behavioral and Brain Sciences, 21, 353–397.
McCarthy, M. M. (1995). Estrogen modulation of oxytocin and its relation to behavior. Advances in Experimental Medicine and Biology, 395, 235–245.
Mehta, P. H., & Josephs, R. A. (2006). Testosterone change after losing predicts the decision to compete again. Hormones and Behavior, 50(5), 684–692.
Mehta, P.H., Josephs, R.A. (2010). Testosterone and cortisol jointly regulate dominance: Evidence for a dual-hormone hypothesis. Hormones and Behavior, 58, 898--906.
Mehta, P. H., Jones, A., & Josephs, R. A. (2008). The social endocrinology of dominance: basal testosterone predicts cortisol changes and behavior following victory and defeat. Journal of Personality and Social Psychology, 94, 1078–1093.
Naftolin, F. (1994). Brain aromatization of androgens. Journal of Reproductive Medicine, 39(4), 257–261.
Neave, N., & Wolfson, S. (2003). Testosterone, territoriality, and the “home advantage.”. Physiology & Behavior, 78, 269–275.
Oliveira, T., Gouveia, M. J., & Oliveira, R. F. (2009). Testosterone responsiveness to winning and losing experiences in female soccer players. Psychoneuroendocrinology, 34, 1056–1064.
Salvador, A., Suay, F., González-Bono, E., & Serrano, M. A. (2003). Anticipatory cortisol, testosterone and psychological responses to judo competition in young men. Psychoneuroendocrinology, 28(3), 364–375.
Schlotz, W., Kumsta, R., Layes, I., Entringer, S., Jones, A., & Wust, S. (2008). Covariance between psychological endocrine responses to pharmacological challenge and psychosocial stress: a question of timing. Psychosomatic Medicine, 70, 787–796.
Stanton, S. J., & Schultheiss, O. C. (2007). Basal and dynamic relationships between implicit power motivation and estradiol in women. Hormones and Behavior, 52(5), 571–580.
Stanton, S. J., & Schultheiss, O. C. (2009). The hormonal correlates of implicit power motivation. Journal of Research in Personality, 43, 942–949.
Suay, F., Salvador, A., González-Bono, E., Sanchís, C., Martínez, M., Martínez-Sanchis, S., Simón, V. M., & Montoro, J. B. (1999). Effects of competition and its outcome on serum testosterone, cortisol and prolactin. Psychoneuroendocrinology, 24, 551–566.
van der Meij, L., Buunk, A. P., Almela, M., & Salvador, A. (2010). Testosterone responses to competition: the opponent’s psychological state makes it challenging. Biological Psychology, 84, 330–335.
Vining, R. F., & McGinley, R. A. (1987). The measurement of hormones in saliva: possibilities and pitfalls. Journal of Steroid Biochemistry, 27, 81–94.
Wilkerson, J. E., Horvath, S. M., & Gutin, B. (1980). Plasma testosterone during treadmill exercise. Journal of Applied Physiology, 49, 249–253.
Wirth, M. M., Welsh, K. M., & Schultheiss, O. (2006). Salivary cortisol changes in humans after winning or losing a dominance contest depend on implicit power motivation. Hormones and Behavior, 49, 346–352.
Woolley, C. S. (2007). Acute effects of estrogen on neuronal physiology. Annual Reviews in Pharmacological Toxicology, 47, 657–680.
Acknowledgments
We thank the Emory University varsity women’s soccer coach, Sue Patburg, assistant coach Rachel Moreland, and the 2013 varsity women’s team for their participation in the research described in this article.
Ethical Statement
This study complies with the laws for research with human subjects in the United States and was approved by Emory University’s IRB. All persons gave written informed consent prior to participation in this study.
Conflict of Interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Casto, K.V., Edwards, D.A. Before, During, and After: How Phases of Competition Differentially Affect Testosterone, Cortisol, and Estradiol Levels in Women Athletes. Adaptive Human Behavior and Physiology 2, 11–25 (2016). https://doi.org/10.1007/s40750-015-0028-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40750-015-0028-2