
Abstract
In the shift from the balance of nature to the flux of nature paradigm, the concept of resilience has gained great traction in ecology. While it has been suggested that the concept of resilience does not imply a genuine departure from the balance of nature paradigm, I shall argue against this stance. To do so, I first show that the balance of nature paradigm and the related conception of a single-state equilibrium relies on what Eliot Sober has named the “Natural State Model (NSM)”, suggesting that the NSM has instead been dismissed in the flux of nature paradigm. I then focus on resilience as the main explanatory concept of the flux paradigm. After distinguishing between two main different understandings of “resilience”, namely engineering resilience and ecological resilience, I argue that the former is close to the concept of balance or stability and still part of the NSM, while the latter is not. Finally, I claim that ecological resilience is inconsistent with the NSM, concluding that this concept–being incompatible with the NSM–is not part of the balance of nature paradigm but rather a genuinely new explanatory tool.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
It is generally agreed that a change of paradigm has occurred in ecology due to the dismissal of the “balance of nature” as a background assumption (Simberloff, 1980; Pickett & Parker, 1992; Wu & Loucks, 1995). Far from being a rigorously defined principle, the balance of nature can rather be considered as a shared belief, an ideology (Blandin, 2009), a myth (Kricher, 2009), or even a religious perspective (Marsh 1964)Footnote 1, but certainly not a neat scientific concept. Nonetheless, it grounded and oriented classical ecological theories by means of a central presupposition: ecological systems tend to reach a state of equilibrium, and hence to maintain it, by going back to it after experiencing a shock (Egerton, 1973). The shift of paradigm–that took place around the half of the 20th century–went hand in hand with the gradual emergence of a different framework, commonly described by the metaphor of the “flux of nature” (Pickett & Ostfeld, 1995) in opposition to that of the “balance of nature” (Cuddington, 2001). The alternative framework acknowledges the influence that variation and fluctuation exert on all ecological systems and denies the traditional assumption of a stable, single-state equilibrium representing their reference state.Footnote 2 On the contrary, it recognizes that ecological systems can go through multiple equilibrium states and persistFootnote 3–alternating stable and unstable phases–in the face of disturbances and variation, whereby the latter are considered as the rule, and not the exception, in ecological dynamics. In virtue of this substantial difference, ecologists also talk about a “non-equilibrium” paradigm to distinguish it from the classical, equilibrium-based one (Pickett et al., 1994; Rhode, 2005). At present, theoretical ecology exhibits a quite remarkable divide between equilibrium and non-equilibrium advocates; however, some authors claim that this separation, despite being historically important, is now barely justified (Cooper, 2001; Eliot, 2011; Van Meerbeek et al., 2021).
Before proceeding, some terminological clarifications are in order. The first regards the expressions “flux of nature” and “non-equilibrium”, both commonly used to refer to the new paradigm in ecology. Notably, none of them manages to wholly grasp the distinctive features of the paradigm: while “flux of nature” stresses the importance of variability but leaves aside the different role that equilibrium plays compared to the balance of nature paradigm, “non-equilibrium” is potentially misleading in conveying the idea that the new paradigm denies the reality of equilibrium states in ecological systems, which is not the case, as it will be clarified in the course of this article. Still, since these expressions are of common use in the contemporary ecological debate, in this article I will use them interchangeably. As for the balance of nature idea, since it has never received a rigorous definition, many terms have been adopted in ecology to articulate it, including “equilibrium”, “stability”, and “resilience” (Delettre & Korniliou, 2022). However, as it was the case with the original expression, these notions present analogous interpretive problems generating confusion and indeterminacy. In an attempt to address these problems, some clarification attempts have been made to classify and better define stability-related concepts (Grimm & Wissel, 1997; Justus, 2007). In some cases, these attempts resulted in theoretical proposals suggesting that equilibrium and non-equilibrium theories–and the relative conceptual frameworks, including the concept of resilience–should be integrated under the umbrella-notion of ecological stability, which must be defined in comparison with the reference conditions of ecosystems (Justus, 2008; Van Meerbeek et al., 2021).Footnote 4
In what follows, I will introduce the leading question of my inquiry, i.e., whether resilience is just another way to name stability and, accordingly, whether the concept of resilience can still be considered to belong to the balance of nature paradigm (Sect. 1). To address this question, I will develop the following argument: first, I argue that the balance of nature paradigm and the related conception of a single-state equilibrium rely on what Sober (1980) characterized as Aristotelian Natural State Model (NSM) (Sect. 2). To do so, I provide a description of the NSM and some of its possible biological versions, thereby unearthing a possible analogy with the classical paradigm of ecology. I then describe the main features of the alternative paradigm, the flux of nature, showing the differences and the implications in relation to the traditional one, subsequently focusing on resilience as the main explanatory concept of the flux paradigm. After distinguishing between two different understandings, i.e., engineering resilience and ecological resilience, I argue that, while the former is close to the concept of balance or stability, the latter is not (Sect. 3). Finally, by comparing the main assets of the classical paradigm with the resilience explanatory framework, I show that ecological resilience is inconsistent with the NSM (Sect. 4). I therefore conclude that resilience–being incompatible with the NSM–is not part of the balance of nature paradigm and hence cannot be considered as the balance of nature under a new name.
2 Framing the question
According to some authors, the demise of the balance of nature paradigm in ecology is mainly terminological in nature and has not implied a genuine dismissal of the general idea that any ecological system possesses the “tendency […] to maintain itself in a certain state or to go back to that state in case of disturbance” (Delettre & Korniliou, 2022, p. 67). According to this interpretation, the concept of resilience is just a more sophisticated version of the balance of nature idea. As a matter of fact, the ways resilience is conceptualized often evoke the traditional, equilibrium-based framework. For example, Pimm (1991; but see also Gunderson, 2000 for other references) defines resilience as the time of return to equilibrium after disturbance, a definition also applying to the notion of “stability” as defined by Holling (1973). Following this line of reasoning, the introduction in ecology of the concept of resilience by Holling in 1973 has, on the one hand, fulfilled the task of tackling the conceptual indeterminacy affecting the non-scientific and metaphorical expression “balance of nature” and its common-use substitute, “stability”; on the other hand, it has met the conceptual needs emerged with the paradigm change in ecology by complexifying the notion of equilibrium in ecosystems (e.g. by integrating the role of perturbations and by identifying thresholds within which equilibrium is maintained despite fluctuations).
While agreeing with the general definition of the balance of nature paradigm given by Delettre and Korniliou (2022) and the importance they attribute to the concept of resilience in overcoming its conceptual indeterminacy, it remains nonetheless questionable whether the concept of resilience should be considered as belonging to the old balance of nature paradigm. More precisely, what I am going to suggest in the following sections is that the introduction of the concept of resilience in ecology implied a genuine change in the explanatory framework adopted in the field. Indeed, as a central concept of the non-equilibrium paradigm, it reflects a fundamentally different way of thinking and calls for the development of new modelling strategies, thus overcoming outdated, classical models of explanation in ecology. Hence, that of resilience should be considered a concept not in continuity but rather in opposition to the balance of nature paradigm.
Of course, a familiarity between (certain definitions of) resilience and other stability concepts cannot be denied. However, I think it is important to take seriously–even if still critically–the role of “paradigm-shifter” that the concept of resilience has been ascribed since its introduction in ecology (Holling, 1973). More in particular, on the one hand, it can be noted that the main problem investigated by the two ecological paradigms–i.e., the “balance of nature” and the “flux of nature”–is the same: the relationship between persistence and variability in natural systems (Pickett & Ostfeld, 1995). On the other, however, the way this problem is addressed, the models and measurements adopted, the theoretical and practical implications, as well as the solutions proposed by each side are different and not reducible to each other. I shall try to illustrate all this in the following sections.
3 The balance of nature and the natural state model
As I have anticipated in the introduction, I will now provide a description of the balance of nature paradigm as an ecological instantiation of the so-called Natural State Model. The idea that the natural sciences in general have been influenced by a theoretical model based on some notion of a natural state has been highlighted, among others,Footnote 5 by Sober (1980). According to him, the NSM is an explanatory model based on an Aristotelian conception of natural essences that still exerts influence on different domains of scientific thinking. While Sober’s inquiry mainly focuses on the influence of the NSM on genetics and biological accounts of development and evolution, I am going to argue that this model has been operative in ecology as well. To do so, I will first provide a characterization of the NSM by exposing its prototypical form, i.e., Aristotelian physics and biology. Then, I will show how it works in modern biology and, analogously, in ecology, specifically in the form of the balance of nature paradigm. I believe that reading the classical paradigm of ecology with the NSM lens could improve the understanding of the nature of the paradigm shift in ecology and of the role that the concept of resilience has played in such shift.
As anticipated, the metaphysical roots of the NSM may be found in Aristotle’s texts. Probably, a good starting point for exposing it is the Aristotelian theory of place. According to this theory, which is central to Aristotelian physics, the simple entities—namely earth, water, air, and fire, composing all animate and inanimate natural bodies–have a proper place representing the natural destination of their movement. It is important to note that, in this theory, movement is presented as the main kind of change since, after all, everything that happens in nature implies a change of place of the elements and of the objects–i.e., the substances–they compose. Aristotle conceives change as the actualization of a potentiality that is inherent to each substance (Bodnar, 2018). In the ever-changing world, movement can either follow a natural trajectory, i.e., toward the proper place of the element, or the substance, that is moving; or it can be interrupted or deviated by interfering forces. In other words, every natural body moves toward its proper place,Footnote 6 unless it is deflected or stopped by some other cause. This happens because, in Aristotelian physics, natural bodies–contrary to artificial ones–contain the principle of movement in themselves, i.e., they have the inherent tendency to follow fixed directions, leading to their natural state or, if they already are in their proper place, to remain there naturally, unless an external force causes their displacement. The implications of the theory of place for physics and biology lead to that way of explaining natural phenomena that Sober (1980) has labelled as “Natural State Model”.
The explanation of the reproduction of the organism given by Aristotle is an instance of the application of the NSM to biology. According to Aristotle’s account, each organism contains and expresses an eidos, a formal essence shared with all the other individuals of the same species (type). Any interfering force causally influencing the execution of the development programFootnote 7 leading to the eidos results in a deviation from the type and a failure to reach the natural state (Sober, 1980). According to Aristotle, the organism’s existence, indeed, doesn’t always lead to the manifestation of the organism’s essence, but it is rather subjected to accidents and contingencies that can cause a departure from the natural state (Kupiec, 1999). It is important to note that such deviation is viewed as a failure in reaching the proper essence. This implies a strongly normative connotation of the natural state, both in the Aristotelian thought–deviated organisms are terata, “monsters”–and in successive elaborations. Throughout the history of the natural science, what is considered natural sets the norm against what is not.
Due to the historical authority exercised by Aristotle’s natural philosophy, the NSM maintained a great influence throughout the history of the natural sciences. Accordingly, variants of the model provided the basis for many pre-Darwinian accounts of biological variability, inspiring both static and evolutionary accounts (Sober, 1980). Both kinds of theories presuppose indeed a set of natural regularities governing the normal path of organismal development (i.e., the path leading to only one possible outcome or natural phenotype) and a multitude of interferences (e.g., environmental variables) interrupting and deviating that process. Under the NSM, variability in nature was explained either as deviation from type or as interference with the natural plan of development and evolution. It was only after the emergence of Darwin’s theory of evolution that alternative models accounting for variability were proposed. The idea that variation is a constant of life, and that it has real causal efficacy in shaping biological processes–and that, as a consequence, is not merely noise, either produced by nature or discounted as causally irrelevant by the observer–significantly changed biologists’ way of thinking (Mayr, 1982).
However, even in contemporary biology, it is still possible to detect traces of the NSM. For instance, contemporary research in developmental biology is still typically based on the intentional idealization of “normal stages”, while variation (such as that generated by the processes underlying phenotypic plasticity) is ignored as noise (Love, 2010). While this reasoning strategy has many advantages for biological research–allowing experimental replication, unambiguous communication among researchers, and the elaboration of predictions and generalizations–it can engender actual blindness towards variability, for instance by underestimating its developmental and evolutionary role and neglecting research on its function. Moreover, despite the important contributions of alternative approaches (Gilbert & Epel, 2015; Jablonka & Lamb, 2005; Lewontin, 2000), the genome is still often conceived, both within a considerable part of the scientific community (Rosenberg, 1997; Wolff, 2013; Wolpert & Lewis, 1975) and among the general public (Gilbert, 2023; Maienschein, 2016), as a program directing development and determining the important features of the phenotype. This reductionist approach is considered to be no longer adequate for explaining the complexity of developmental and evolutionary processes (Keller, 2002) and the reason for this resides in its commitment to (a) a natural state of biological entities; (b) a fixed, pre-determined pathway of development; and (c) the characterization of mutation or environmental influence as an interfering force. That being said about ancient and modern biology, I shall now argue that something analogous has happened in ecology.
As a way to postulate a natural order underlying the variety of ecological phenomena, the balance of nature paradigm exhibits a striking familiarity with the NSM. This consonance underlies my suggestion that a version of the NSM operates in the classical paradigm of ecology as well. In what follows, I will motivate this claim by providing a description of the first comprehensive theory of ecology, which is also the very first attempt to render the balance of nature paradigm scientifically respectable (Simberloff, 1980; Tobey, 1981), namely, Frederic Clements’ theory of community succession. According to Clements (1916), ecological communitiesFootnote 8 are superorganisms whose development follows a pathway of fixed stages called succession. Clements gives the name “climax” to the last stage of succession, representing the ultimate and reference state of the community. The climax is characterized by an optimal fit between the biological organization of the community (e.g., species distribution, population regulation) and the external environment (mainly the climate). This implies the efficiency of community functions and–significantly–a long-lasting condition of stable equilibrium. Which climax type characterizes each community is mainly determined by the climate: according to the climatic conditions, the climax configuration will display a specific composition and structure. Failure to reach or maintain the climax is due to the interference of external agents, such as natural disruptive events or human activities, that within this framework are only considered as disturbances (McDonnell & Pickett, 1993).
In Clements’ theory, the climax formation, being conceived as the mature stage of the community–through which it succeeds in exemplifying its type–can be considered in analogy with the adult and reproductively successful organism in traditional developmental biology, and hence as the natural state of ecological communities. Succession is likewise considered as a progressive pathway of “normal stages”, reminiscent of the Aristotelian movement or the genetic program directing the development of the organism. Given such a predetermined pathway towards a single steady state, every disturbance–be it natural or human-induced–is conceived as an interfering force that could delay, deviate, or arrest the succession of the community toward its climax. Communities that did not reach their climax state are considered as incomplete or deviant communities, thus finding no place in the taxonomy of natural ecological units.
As already mentioned, the rigid teleological structure theorized by Clements can be considered the first explicit scientific conceptualization of the balance of nature idea, as well as the first paradigm of ecology (Pickett & Ostfeld, 1995). While Clements’ theory of ecological succession applied to vegetational communities, other ecologists adopted a similar conceptual framework for the study of animal populations.Footnote 9 Notably, mathematical models describing population regulation dynamics–such as the Lokta-Volterra equations–gained huge importance in the 50s. Generally, they related the stability of population to their diversity or complexity (Justus, 2007). Ecosystem ecology as well has emerged within the balance of nature paradigm’s dominance. Indeed, when Arthur Tansley’s (1935) introduced the concept of ecosystem in ecology, he defined it in relation to equilibrium: the more stable its equilibrium, the more an ecosystem will persist (Golley, 1993). Later on, Eugene Odum’s (1953) classical characterization of the ecosystem was even more strongly positioned in line with a Clementsian version of the balance of nature paradigm: ecosystems developed following a progressive succession, with an inherent tendency to reach homeostasis, a steady state in which inputs and outputs of energy are balanced.Footnote 10 In light of the similarities with the features of the NSM exposed above, it is thus reasonable to argue that the NSM generally underpins ecological theories developed within the equilibrium paradigm (or that the paradigm can be conveniently interpreted in such terms). To synthetize, the ecological version of NSM can be generalized as follows: (i) ecological systems possess only one possible natural state, (ii) there is a fixed, pre-determined pathway of development that must be followed by ecological systems to reach their natural state, and (iii) those forces interfering with that pathway hinder the normal development of ecological systems and hence deviate them from their natural state.
Given that, starting from the 60 s, ecology has assumed a growing importance in guiding environmental policies (Worster, 1994), the implications of the NSM for resource policies and ecosystem management should be mentioned at this juncture. Despite the faith in the deterministic nature of succession (Simberloff, 1980) and in the climax (i.e., a single reference state resulting necessarily from the “natural” development of an ecological system), the pitfalls of the classical framework emerged during the first environmental management interventions. Clements’ climax theory revealed its poor predictive power, often resulting in the unsatisfactory or even harmful management of ecosystems (Botkin, 1990, Pickett et al., 1992, Mori, 2011). One iconic example is the case of the American Northern Plains, destroyed because of the combined effects of intensive agriculture and a series of aggressive dust storms that occurred during the 1930s, a phenomenon historically popularized as the “Dust Bowl”. On that occasion, restoration practices were planned according to Clements’ theory of succession. The intended goal was to restore the climax of the Plains–their “natural” configuration–by re-introducing the native species and waiting for the spontaneous development of succession. However, all efforts failed because the altered initial conditions of the Northern Plains ecosystemFootnote 11 couldn’t allow the establishment of what was considered the only possible final stage of succession. In general, during the second half of the 20th century, disagreement toward the environmental determinism implied by Clements’ theory–and its relative policy implications–gradually spread by hand of an emerging scholarship.
The main reason for the growing discontent with Clements’ theory was that it couldn’t withstand empirical scrutiny. Indeed, it is a general feature of equilibrium theories at any level of biological organization to adopt a somehow aprioristic and speculative approach to ecological dynamics. On the one hand, such an approach allows for the elaboration of sophisticated and mathematically sound models, allowing for relative predictive success. On the other, however, as components and interactions are added to increase models’ realism and adequacy, it gets more and more difficult to provide the relative empirical measurements (Justus, 2007). In other words, it gets difficult to corroborate empirical observations through such theoretical a priori models. A relevant example in ecology, described by Justus (2008), is the so-called Lyapunov stability. Named after the mathematician that elaborated the relative equations, this model for stability efficiently fits physical systems, as it accounts for perturbations affecting the values of the system’s variables. Accordingly, it has been used in ecosystem ecology as well for describing population fluctuations. However, Lyapunov stability fails in accounting for changes in the values of the system’s parameters: in other words, it does not capture the effects of perturbation at the level of the structure of the system, which in turn affects the behaviour of the variables. While this seldom happens in the case of controlled, close physical systems, it is quite common for complex, open systems such as ecosystems. Hence, as Justus (2008) concludes, Lyapunov stability is not an adequate theoretical tool for understanding ecosystem’s stability.
It was exactly this disconnection between the speculative and the empirical results in ecology that, by the end of the 20s, brought Henry Gleason and, later, an increasing number of ecologists to question Clements’ doctrinaire approach and make pleas for a more realistic–i.e., more empirically-based–framework for explaining ecological phenomena (Kingsland, 2005). As I will expose more broadly in the next section, this divergent attitude resulted in the advancement of a set of theories and methodologies bearing a stronger focus on the variability of environmental conditions, rather than on the natural balance.
4 The non-equilibrium paradigm and resilience
Henry Gleason, an American botanist contemporary to Clements who, at that time, was investigating the vegetation of Illinois, is generally considered the pioneer of the non-equilibrium paradigm (Kingsland, 2005; McIntosh, 1975). He devoted a consistent part of his work to elaborating a critique of Clementsian ideas. In particular, at the base of his disagreement with the colleague was the question of the nature of the ecological community. Gleason strongly refuted Clements’ organismal conception and advanced instead what he called an “individualistic” view (Gleason, 1926). The motivation for this proposal originates in the observations that Gleason himself collected on vegetation assemblages in Illinois, in the course of which he could note that the species distribution was not homogeneous in nature, contrarily to what was assumed at the time. He rather noted a random, fluctuating and heterogeneous distribution of vegetation, contrasting with the traditional conception of ecological communities as well-defined, integrated entities that could be classified by their climax. Consequently, in Gleason’s opinion the Clementsian organismal view of ecological communities was untenable. Ecological reality should not be seen as organized in organism-like communities, following an orderly, predictable and progressive development towards a self-perpetuating climax. Rather, Gleason claimed, a vegetation assemblage is but a “temporary and fluctuating phenomenon” (Gleason, 1926, p. 93) resulting from contingent conditions rather than from an internally determined development. From Gleason’s account of the nature of ecological entities naturally follows a novel view concerning the dynamics of their change. The variability that can be empirically observed in the dynamics of ecological assemblages could not be interpreted as mere noise or deviation from type. The accumulating evidence of fluctuation and retrogressiveFootnote 12 tendencies in the succession of ecological communities were in sharp contrast with the idea of a pre-determined progressive path toward a single final stable stage.
Summarizing what has been exposed so far, the individualistic conception of ecological units and the relevance attributed to environmental fluctuation were thus the main elements of Gleason’s critique toward Clements’ climax theory (McIntosh, 1975). Gleason’s ideas gave the impulse to a new wave of research in ecology, in which more emphasis was dedicated to aspects of contingency and variation in the study of ecological communities, thus refusing the natural state approach that was typical of the balance of nature paradigm. The classical ideas about a progressive succession leading to a stable climax were increasingly challenged (Whittaker, 1957), as they proved to be misleading hypotheses once they were tested on the field or confronted with empirical observation (Kingsland, 2005). Many alternative theories, measurements, and models emerged, both in population ecology (see Rhode, 2005 and Justus, 2007 for an overview) and, between the 60s and the 70s, in ecosystem ecology (Holling, 1973), resulting in the emergence of a non-equilibrium paradigm, in which change–and not equilibrium–is considered as the real driver of natural phenomena. The contrasting features of the equilibrium and the non-equilibrium paradigms have been summarized by Cooper (2001, p. 489) as follows:
[…] the importance of history over ahistorical equilibria, contingency over “laws” of population growth and community composition, the uniqueness of actual individuals over the homogeneity of the abstract individual, pluralism over monolithic theory, and chance over determinism.Footnote 13
It is important to specify that the kind of equilibrium that is rejected by non-equilibrium theories is the one that Cooper defines as “ahistorical”. Indeed, Clements’ characterization of the climax could be considered an instance of such an ahistorical conception of equilibrium, i.e., as a fixed and pre-determined reference state that ecological communities are “programmed” to reach–either as a result of a pre-determined development process or by following an intrinsic tendency–independently of the historical and contingent conditions. In other words, non-equilibrium ecology rejects the notion of equilibrium as a natural state. Therefore, in no way this means that the non-equilibrium paradigm denies the existence of equilibrium states in the dynamics of ecological units: rather, equilibrium must be considered as a relative and contingent condition, while the possibility of a multiplicity of possible equilibria of complex ecological systems must be recognized and, furthermore, unstable phases, disequilibria or non-linear dynamics are not to be interpreted as noise or deviations from type, but rather as part and parcel of typical ecosystems’ behaviour. At the same time, this alternative interpretation of ecological dynamics engendered a tension: how could the relationship between the persistence and relative stability of ecological systems and their constant variation be explained, given that in the new paradigm the notion of equilibrium has lost its privileged explanatory power? In order to solve that tension, non-equilibrium theories needed an alternative explanatory concept to the single-state equilibrium. What kind of concept could play this role?
To answer this question, let me summarize what has been said so far. From the perspective of the new flux of nature paradigm, the main shortcomings of the classical ecological paradigm–both for the understating of the actual ecosystem behaviour and for environmental management–derive, on the one hand, from the conception of equilibrium as a global, single-state reference point and, on the other, from the relegation of natural variability to the status of noise (Holling & Meffe, 1996; Pickett & Ostfeld, 1995). Resilience has been recognized by an increasing number of ecologists as a promising concept to deal with both issues, rapidly gaining a central relevance in the new paradigm. The concept of resilience has been introduced in ecology by J. C. Holling by transferring it from its original domain, i.e., mechanics, and readapting it to ecology (Alexander, 2013; Reid & Botterill, 2013). The main reason for introducing the concept was that, despite the new empirical evidence and the crisis that the balance of nature paradigm was undergoing, ecology was still focused on the idea that an ecosystem is defined by a global single-state equilibrium, i.e., a condition of stability thought to be its optimal configuration, characterized by efficiency of function (Holling, 1973). As a way to step out from such a persistent attitude toward ecosystem dynamics–and to avoid its deleterious implications for resource management (Holling & Meffe, 1996)–Holling inaugurates a new research framework in ecology by elaborating the original definition of resilience relevant for ecologyFootnote 14:
Resilience determines the persistence of relationships within a system and is a measure of the ability of these systems to absorb change of state variables, driving variables, and parameters, and still persist. (Holling, 1973, p. 17)
According to Holling (1998), a “transition in ecology” was needed in order to harmonize the contributions of ecological scientific inquiry and environmental policies in the practice of ecosystem management. In his view, ecologists must be sensitive to the practical implications of their scientific activity and should engage in guiding policy efforts to tackle the many problems affecting the interaction between people and nature in contemporary time. As another leading scientist in the resilience research has pointed out in a popular article (Folke, 2006), old perspectives postulating ecosystems’ self-regulation toward equilibrium are inadequate for understanding the role that disturbances have on ecosystems’ behaviour, thus impacting negatively on the possibility of sustainable environmental management. The primary reason for the introduction of the concept or resilience was, then, the development of an ecological science practically adequate to deal with environmental governance issues. Of course, this aim could only be achieved with the integration of a broader resilience-related conceptual framework–including, among others, the notions of perturbation, variation and stochasticity–in non-equilibrium ecological theories.
Unsurprisingly, this conceptual innovation inaugurated a phase of intense research which rapidly widened Holling’s seminal definition. A growing network of scientists focusing on the potentialities of resilience for the understanding of ecological systems and produced a variety of different interpretations and redefinitions of the concept (Brown, 2014; Desjardins, 2015). On the one hand, the emergence of such a varied and rich framework of “resilience thinking”Footnote 15 has eventually allowed to cross boundaries between sciences and to unify ecology and the social sciences under the umbrella of resilience thinking (Adger, 2000; Folke, 2006; Mathevet & Bousquet, 2014). On the other, however, justified worries have been raised that the acritical multiplication of definitions and uses may result in both terminological ambiguity and dilution of the scientific significance of the notion (Brand & Jax, 2007). Moreover, the concept is increasingly used with a normative connotation, making it a controversial discursive tool in the wake of its predecessor, the balance of nature idea. Indeed, environmental management, but also in other contexts like social and urban planning, disaster risk reduction, psychology, economics, and so onFootnote 16 increasingly equate resilience to a desirable property of any system in absolute terms (Klein et al., 2003), without contextualizing or operationalizing it. This state of affairs generates controversy: for example, both natural and social systems, e.g., a lake in a eutrophic state (Holling, 1973) and a totalitarian regime (Holling, 2001), can intuitively be resilient in undesirable ways, generating justified suspect over some of the current uses of the concept (Reghezza-Zitt et al., 2012). Furthermore, the fuzziness affecting the concept of resilience might lead to misapplications, especially when it is prescribed to individuals or collectives in the face of various kinds of distress (Davoudi, 2018). These concerns have been answered by efforts to distinguish the different frameworks in which the concept of resilience is embedded and by making pleas for better-informed uses (Alexander, 2013; Brand & Jax, 2007). While the disquiet around the problems affecting the broad field of resilience thinking is justified and deserves careful attention, it is a strand of analysis that would lead beyond the scope of this work. Therefore, in what follows I will limit my analysis to the explicatory role played by the concept of resilience as a scientific concept in the field of ecology.
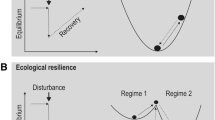
Elaborating on what has been said so far, it is important to note that, in the original definition of the concept of resilience, both persistence and change are included, a feature that remains traceable also in the subsequent definitions of the term. Explaining the relationship between the persistence and change of ecological units was also the role that the notion of single-state equilibrium played in the balance of nature paradigm. Contrary to the older paradigm, however, resilience is conceived as an “ability” or “capacity”–and not a “state”–that ecological systems display–to varying degrees and in various modalities–in their dynamic behaviour: as such, it implies the rejection of the postulation of equilibrium as a natural state resulting from the development of ecological systems or determining their dynamics in order to preserve their identity when confronted with interfering forces. However, the meaning that resilience is currently ascribed in ecology does not correspond to the original one (Reid & Botterill, 2013). In its original context, i.e., mechanics, the notion of resilience was used to describe the strength, elasticity and ductility of materials to which force is applied. The more resilient the material, the more rapid its return to the pre-disturbance conditions. Consequently, determining the resilience of an object allowed engineers and builders to know how much pressure could be applied to it before its rupture. As it has been observed (Bodin & Wiman, 2004), this way of intending resilience equates it with elasticity and considers it as a component of stability, which is usually the central focus when investigating the qualities of a system for technological reasons. Indeed, when considering a mechanical system, the point is usually to ensure its durability and efficiency by excluding the possibility of oscillations or non-linear behaviours. But the kind of systems ecology deals with are not the closed, simplified ones that are the object of mechanical science. On the contrary, ecological systems are complex systems, i.e., open systems presenting a high number of components, a high number of interactions among those components, a certain degree of disorder, as well as feedback mechanisms, all features generating emergent propertiesFootnote 17 that distinguish ecological systems from the closed, simplified ones that are the object of engineering science. As Holling noted, the mechanical definition of resilience, that he labels engineering resilience, cannot apply to ecological systems. An adapted ecological version is required. Holling hence provided a distinction between two possible meanings of resilience: the just mentioned engineering resilience and ecological resilience.
Engineering resilience measures the speed of return of a system to the previous equilibrium state, or “normal regime”, after disturbance. Usually, in ecology this understanding of resilience is used in management contexts where the focus is on the efficiency, constancy and predictability of ecosystems, which are conceived as mechanical systems, or–in the case, for instance, of croplands or grasslands–designed for single-operating objectives (Holling & Meffe, 1996). Here, disturbance is viewed as an undesirable event that must be limited as much as possible to prevent inefficiency.
Ecological resilience measures instead the magnitude of disturbance that an ecosystem can absorb before redefining its structure by changing the variables and processes determining its behaviour (Ibid.). In this understanding of resilience, the occurrence of multiple equilibrium states is recognized, with disturbance possibly influencing transitions between different states.
It has been noted that the engineering definition of resilience corresponds to the traditional notion of ecological stability, i.e., the ability of an ecosystem to return to an equilibrium state after a temporary disturbance (Holling, 1973).Footnote 18 However, this notion is implicitly committed to the assumption of a global and single-state equilibrium to which ecological systems strive to return (Gunderson, 2000). Therefore, by postulating such equilibrium, the concept of engineering resilience is still part of the balance of nature framework and its related commitment to a natural state. More in particular, it corresponds to the traditional notion of stability as it delivers its same explanatory role, i.e., to account for the persistence of ecological systems through change. Conversely, ecological resilience, by recognizing the possibility of different equilibrium states and the constitutive role played by disturbances in shaping ecosystem development, does not correspond to the traditional notion of stability. It rather seems to be a genuinely new concept, even if there is not, at present, consensus over its proper definition. According to some, resilience is a multi-dimensional (Desjardins, 2015) or polysemous (Strunz, 2012) concept, being conceived either as a set of properties (Carpenter et al., 2001) or in conjunction with other properties, like adaptability and transformability (Walker et al., 2004). According to others, it is a stability concept (Justus, 2008; Van Meerbeek et al., 2021). There is also who claims that resilience cannot be characterised independently of context but should, instead, be used hand in hand with the specification of the system (resilience of what?), the type of disturbance (resilience to what?) (Carpenter et al., 2001) and the type of identity that must be conserved in the system (Delettre, 2021), which are all specifications depending on the choice of the observer. In any case, because it departs from the traditional notion of stability in describing ecological systems’ behaviour, it is resilience understood sensu Holling, i.e., as ecological resilience, the version of the concept that should be used in ecological research (Brand & Jax, 2007).
While resilience theory gets more and more sophisticated, its measurement and practical applications remain underdeveloped (Folke, 2006). The insufficient operationalization of the concept can be explained by both the complexity of the property it denotes–including ecological thresholds and the response of a system to different disturbances, many of which are structural (Justus, 2008) or act at different spatial and temporal scales (Mori, 2011)–and by the lack of consensus over its meaning and interpretation in ecological theory. However, the difficulties in empirically assessing and measuring this property and, consequently, in applying the concept to practical projects contrasts with one of the central features of the non-equilibrium paradigm–at least in its “reasonable” versions (Cooper, 2001)–i.e., its strong focus on empirically-driven research. Nonetheless, efforts towards a more adequate quantification and measuring of resilience have been made (see Hodgson et al., 2015 for a programmatic overview). As a result, while they still have not turned into a uniform modelling strategy, formalization attempts have at least revealed some key differences with equilibrium models. First of all, perturbations and disturbance regimes are included in non-equilibrium models; this implies that they are not merely considered external interferences but, rather, as part and parcel of ecosystem dynamics. However, given the unpredictable nature of environmental shifts and the often-non-linear behaviour of disturbance regimes, it should also be highlighted that it is not easy to include environmental disturbance in the models (but see, for instance, Mumby et al. (2013) in the context of coral-reef resilience measurement study). Secondly, temporal and spatial scale effects must be included in modelling, as the resilience framework accounts for cross-scale effects in ecosystem dynamics in response to disturbances. For instance, Falk et al. (2019) provide a model of scaled resilience in relation to a case study disturbance, i.e., fire affecting forests. Disturbance and the ecosystem response it triggers are described on three axes: spatial extent of disturbance, recovery time and level of biological organization involved (individuals, populations, communities). The bigger the spatial and the temporal scale of the disturbance, and the greater its intensity, the higher the organizational level and the longer the temporal extent are required for the ecosystem to show resilience. These are just a couple of selected examples in the growing literature concerning resilience modelling. Much work is certainly still to be done before resilience becomes a truly operationalized property that could be used in management projects.
5 Ecological resilience and the natural state model
In the previous section, I argued that engineering resilience–corresponding to the notion of stability–is part of the traditional paradigm, while ecological resilience may be instead a truly new explanatory tool. In the remainder, I will critically address the question, introduced in Sect. 1, whether the concept of resilience is still committed to the balance of nature paradigm.
Before proceeding, I need to make a quick clarification: having distinguished between engineering and ecological resilience and individuated the latter as the relevant version of the term for ecological inquiry, from now on I will solely refer to the ecological understanding of the concept of resilience. In doing so, I will follow Holling’s inaugurated line of thought, and therefore refer to other, successive elaborations on the concept of resilience.
If resilience were a concept operating within a balance of nature framework, as Delettre and Korniliou (2022) have suggested, then–having individuated the NSM as the general way of reasoning underlying the balance of nature paradigm–it should exhibit some reference to–or at least be consistent with–some notion of a natural state, which in ecology is usually represented by the global, single-state equilibrium. In order to establish if this is the case, the concept of resilience will now be tested against the relevant aspects of the NSM presented in Sect. 2, namely (i) the existence of a single natural state (ii) the existence of fixed and necessary pathways to reach it and (iii) a negative conception of disturbances as external forces interfering with natural development.
(i) Does the concept of resilience make reference to or imply the existence of a natural state for ecological systems? Throughout Sect. 3, I have illustrated how ecological resilience fundamentally rejects any reference to a monistic conception of equilibrium. Actually, Holling’s formulation of the ecological meaning of resilience precisely aimed to provide an adequate analytic tool for multi-state fluctuating ecosystems. Ecological resilience indeed describes the amount of disturbance an ecosystem can deal with before shifting to an alternative regime of behaviour: this does not imply that, during and as a result of this shifting, the ecosystem fails to reach equilibrium, but rather that it can cross–without deviating from any natural state–a variety of possible stable states characterized by different structural, organizational and functional properties.Footnote 19 Stable states are thus possible, but they are temporary and contingent and they don't constitute natural reference conditions of ecological systems (Pickett et al., 1992).
(ii) Does the concept of resilience make reference to or imply the existence of a pre-determined path that ecological systems must follow to reach their natural state?
From the negative answer to (i), it logically follows a negative answer to (ii): since no natural state is implied in the concept of resilience, it is impossible that it could imply the existence of a unique, pre-determined succession like the one theorized by Clements for ecological communities. If an ecosystem can cross among multiple equilibrium states, and if the transition between states can be influenced by disturbance events, then the conception of an entirely internally-determined development–assumed in the NSM as the necessary path to reach the natural state–can only be seen as an idealization. Therefore, resilience does not manifest itself through sequences of “normal stages” that can be predicted or prescribed to ecological systems. On the contrary, many possible trajectories are possible according to the specific historical conditions of each ecosystem, which are unique and irreproducible. Change is determined both by disturbance coming from outside the ecosystem and by interactions among its internal components at different scales (Pickett & Ostfeld, 1995). Thus, ecosystems are not predictable in their behaviours and should be studied in their context and complex organization. Indeed, their processes occur at different spatial and temporal scales creating non-linear effects (Holling & Meffe, 1996). Resilience is hence only observable at specified temporal and spatial scales in ecosystem (Walker et al., 2004) and in no way presupposes a global end state to be reached as a result of a pre-determined linear succession.
(iii) Are disturbances just disruptive interferences?
Disturbance assumes, in relation to ecological resilience, an important causal role. Not only disturbance is granted a full ontological status; in fact, disturbance events are the rule, and not the exceptions, in a nature in constant “flux” (Pickett & Ostfeld, 1995). Moreover, its agency in shaping ecosystem structure and processes is integrated as a “key component” of ecosystem behaviour (Turner, 2010) and can as a consequence no longer be relegated to the status of external interference. As a result, the contrast with the NSM is flagrant again, since the assessment of resilience necessarily requires specific consideration of disturbances as fundamental agents of the persistence and change of ecological systems (Mori, 2011).Footnote 20 Another consequence of the acknowledgment of the role of disturbance is the abandonment of a normative stand towards it: at the ecosystem scale, and with the lack of a reference natural state, a disturbance is described as one kind of ecological process among others, and not as something diminishing the “naturalness” (and henceforth the health) of ecosystems. As such, disturbance events and regimes add to the internal dynamics of ecological systems in influencing their resilience, and disturbance regimes of cyclical recurrence are often considered as part of ecosystem dynamics (Burton et al., 2020). Moreover, human activities are framed as qualitatively equivalent disturbances vis-à-vis natural ones (McDonnell & Pickett, 1993). Thus, within a resilience framework, humans are recognized as part of ecosystems rather than external perturbators, as it was the case in Clements’ account. Finally, both natural and anthropogenic disturbances can have non-obvious, long-term and cumulative effects affecting ecosystem resilience, which can be augmented or diminished accordingly. This means that resilience can be reduced or increased by–intentional or unintentional–human activities (Mori, 2011). In this sense, it provides ecosystem management a conceptual tool to avoid simplistic approaches and to appropriately consider the role of disturbance in assessment and planning.
Clearly, the concept of resilience is inconsistent with all three main elements of the NSM–together with their deterministic and teleological implications. No natural state is assumed when looking at an ecosystem through the lens of resilience, and hence no fixed succession is recognized, while disturbances–in the form of natural or anthropogenic forces–are recognized a full causal role in the development of ecosystems. Hence, resilience, when understood as ecological resilience, seems to provide a genuinely alternative explanatory framework to account for the persistence and variability of ecosystems compared to the balance of nature perspective.
6 Conclusion
The classical paradigm in ecology, based on the balance of nature idea, relied on stability, defined as return to a single-equilibrium state, for explaining ecosystem successful response to disturbance. This framework has been put into question by the alternative, non-equilibrium paradigm. However, one of the main explanatory concepts of the new paradigm, resilience, has been interpreted by some as just a new version of the traditional stability concept, henceforth situating it within the old paradigm. In this article, I formulated an answer to the question concerning whether or not the concept of resilience is subsumed by the old balance of nature paradigm. In doing so, I suggested that the balance of nature paradigm is pivoted, in analogy with classical explanatory frameworks in developmental biology, on a particular model of explanation, namely the Aristotelian Natural State Model. This model postulates (i) the existence of a natural state (ii) the existence of fixed and necessary pathways to reach it and (iii) a negative conception of disturbances as external forces interfering with natural development. My analysis aimed to show that the main elements of the concept of resilience are at odds with the NSM, leading to the conclusion that resilience, when understood as ecological resilience, is not just the balance of nature by a new name. As a result, I concluded that the concept is not only incompatible with the balance of nature paradigm but, even more so, it stands in opposition to it.
Actually, the concept of resilience captures the ability of an ecosystem to persist in time despite change without referring to a defining single-state equilibrium (i.e., natural state); it relinquishes the linear and deterministic conception of succession in ecology; finally, by including disturbance in its definition, it entails a proper consideration of the causal role of variability in ecosystem behaviour. In virtue of these features, the concept of resilience exemplifies the change of thinking that occurred in ecology with the advent of the non-equilibrium paradigm and seems to be more consistent with the now broadly adopted flux of nature paradigm.
Resilience hence seems to be a genuine alternative to stability, both for ecological theory and practice, entailing an anti-deterministic and nonteleological approach to ecosystem behaviour. Environmental management undoubtedly benefited from the insight that this concept allowed since its introduction in ecology. In particular, the young discipline of restoration ecology–and the related practice, ecological restoration–emerged as an alternative to, on the one hand, the established framework of conservation and, on the other, wilderness preservation (Martins, 2022). In very general terms, while conservation ecology aims at maintaining ecosystems as reliable furnishers of ecosystem services and resources by ensuring their biodiversity and integrity, wilderness preservation takes on a “hands-off” approach by creating protected areas in which the human impact is erased, so that their “naturalness” is preserved. Therefore, the basic assumption underlying restoration ecology is different from the preceding environmental management frameworks: as there is no optimal nor natural state to be preserved in ecosystems, and as natural systems do not self-recover to their previous states after disturbance, restoration practices should aim to recover something else. As Anna Tsing (2015) points out, we should refrain from endorsing the misleading assumption that perturbations come to alter a previously harmonious state; rather, constantly changing environmental conditions inevitably correspond to constantly changing ecosystems, resulting in the constant “flux of nature” that I evoked throughout this article. Moreover, as the current environmental crisis and the past and present anthropogenic disturbance often exclude the possibility of both defining and restoring past “untouched” states of degraded ecosystems, their recovery should integrate–historical and present–human activities as well as those of other species. As it is increasingly suggested, the aim of environmental interventions should be to build novel, more liveable and durable systems compatibly with the current historical conditions (Martins, 2022), rather than maintaining or re-creating the allegedly “natural” ecosystems of the past. To succeed in reaching such goal, an active management of ecosystems focusing on enhancing resilience–rather than reaching or preserving equilibrium–may be key (Ren & Coffman, 2023). Indeed, what Palmer (2016) has called “restoration for resilience” would challenge the assumptions of the NSM. In alternative, it would disclose a different approach to the uncertain trajectories of global change and the complex dynamics of ecosystems,Footnote 21 taking on the challenge of modelling restored ecosystems on the basis of future–and not past–conditions (Palmer et al., 2016).
Notwithstanding the innovative and promising trajectories of restoration ecology, the majority of current environmental interventions keep ignoring the specific ecological dynamics of ecosystems. Successional models based on single-state equilibrium and predictable pathways still ground many environmental practices. While such models can prove successful in some cases–usually, in the absence of factors constraining succession out of its predictable trajectories–in most cases re-establishing an ecosystem’s desired historical condition by fast-forwarding its natural succession remains a myth (Hilderbrand et al., 2005). Moreover, as resilience becomes an increasingly leading concept in restoration practice, it may not deliver its potential benefits. As a matter of fact, while the actual (usually, degraded) and the past states of ecosystems are assessed by ecologists on a scientific basis, the design of future states as objectives of restoration is usually a shared process, in which different stakeholders bearing competing socioeconomic interests intertwine with ecological values (Zellmer & Gunderson, 2008). The reality of ecosystem management, also in a restoration frame, is that of a trade-off among a variety of factors encompassing the ecological and the social sphere. In such a complex scenario, it gets very difficult to operationalize resilience, i.e., to say the “resilience of what, to what” is pursued (Brand & Jax, 2007). As a result, even if resilience could get a clearer characterization–an aim towards which the present article aims to contribute–in the theory of ecology, the practice shows a much more nebulous use of the concept, which is often used in its “engineering” meaning, i.e., as an analytical tool to enhance the ecosystem’s efficiency of production by establishing a single steady state and avoiding others (Holling, 1996).
The application issues affecting resilience just described reflect the more general struggle affecting restoration ecology in gaining actual relevance for guiding environmental policy (Jørgensen et al., 2014). Moreover, as the concept is increasingly adopted in different contexts than ecology, the danger of conceptual confusion, dilution of scientific significance and misuse is even more real (Brand & Jax, 2007). It is an open question whether and how the concept of resilience will deliver its promises both in the theoretical and the practical sphere. Today, the field of ecology is but one of the many contexts of use of the concept. As a normative and non-rigorous connotation of the meaning of resilience spreads in many branches of the public discourse, its technical and more rigorous connotation struggles to establish in the crossing of the strictly scientific field. As a result, the potential interdisciplinary benefits of the concept of resilience may rather result in mere acritical usage of a fashionable term. Despite these perils, uncovering the philosophical frameworks underlying the usage of scientific concepts–as I attempted to do in this article–is a necessary step for grasping their role both in scientific and in extra-scientific discourse.
Notes
Marsh’s natural theology played an important role in shaping the modern conservation movement. According to him, the balance of nature is created by God, and humans must take responsibility for its destruction and repair it with wise ecological practices. See Kingsland’s (2005) historical reconstruction for a more detailed description of the different roles that the balance of nature idea has played in ecology.
The reference state is the configuration of the ecosystem against which change and variation are identified or, in other words, its “normal” and stable state (Grimm & Wissell, 1997). Different criteria can be pointed at in the effort to determine a reference state such as, for instance, the set of natural conditions in the absence of human disturbance, or the state of the ecosystem at some point in the past. As Jax et al. (1998) suggest, reference states are usually invoked as natural or absolute states of ecological units, thereby implying a strong ontological commitment concerning the nature of ecological objects. An alternative would be the individuation of criteria for self-identity allowing for the comparison of different ecological units according to certain values: in this case, which reflects an epistemological stance, ecological units are considered as abstractions extrapolated from reality according to historical and social criteria, with the implication that there would be no natural reference state for ecosystems. For an overview of this debate, see Stoddard et al. (2006).
Here I follow Delettre’s (2021) account of persistence defined as the maintenance of identity of the system.
Whether resilience, in its actual usage, should be considered as a component of the contemporary conception of ecological stability or as an independent concept is a matter of on-going debate in ecology. To overcome the conceptual vagueness affecting resilience will surely require taking a position on this matter. While I will not attempt that in the present article, since it would lead me beyond the circumscribed research question that inspired it, I genuinely thank Revisor 2 for making the importance of this question clear to me.
Following Morison (2002, p. 51), Aristotle’s place has causal, explanatory, and defining power over the elements: "[Aristotle] has in mind the power that places have to define what the elements are, for the elements would not be that way they are if their own places were not related in the way they are. This tells us something about how places play an explanatory role in, or make a difference to, the world".
See Cooper (2001) for an analysis of the balance of nature in population ecology.
Retrogressive succession occurs when an ecological community changes following opposite directions compared to progressive succession (Shugart 2001). Hence, if progressive succession is usually intended as a change in the direction of increasing species diversity, structural complexity and stability, retrogressive succession implies a decrease in these factors.
To this description could added the distinction between equilibrium and non-equilibrium theories given by Pickett et al. (2007). According to these authors, theoretical ecology thus illustrates the distinction between equilibrium and non-equilibrium theories: in the case of the former “interactions [of the ecological system] are considered to be stationary, or the same through time”, while in the case of the latter “episodic and rare events may sometimes alter the dynamics of the system, and understanding the current state of a system is not sufficient to understand the trajectory through time” (p. 118).
More precisely, the notion of resilience was already widely used in engineering and construction disciplines as a property of building materials (Hassler & Kohler, 2014).
By the turn of the twentieth century, resilient thinking eventually converged in the institutionalization of a specific journal, Ecology and Society, and of the Resilience Alliance interdisciplinary scientific network.
For a detailed review of the meanings and contexts of the use of the term “resilience”, see Reid and Botterill (2013).
This list of features characterising complexity is taken from Wiesner and Ladymann (2020); the emergent properties these authors argue to be the result of complexity are nonlinearity, self-organisation, robustness of order, nestedness, robustness of function, adaptive behaviour, modularity and memory. All of them are studied in ecosystems and in socio-ecological systems.
The basic question underlying the possibility of alternative stability regimes is how the persistence through change can be explained: how can an ecosystem remain “the same” while shifting across different states? Answering this question would require, first, determining the ontological status of ecosystems, and then endorsing a specific theory of identity of biological objects. It is beyond the scope of this article to make a contribution in this sense. Possible accounts for an investigation in this direction are Jax et al. (1998) and Huneman (2011, 2014a, b).
As Suding et al. (2016, p. 28) point out in their inquiry on the relation between ecological theory and restoration practice, “the dynamics of ecological systems, particularly of a degraded system undergoing restoration, is a function of many factors, some deterministic and some stochastic, working at multiple temporal and spatial scales”.
References
Adger, W. N. (2000). Social and ecological resilience: Are they related? Progress in Human Geography, 24(3), 347–564. https://doi.org/10.1191/030913200701540465
Alexander, D. E. (2013). Resilience and disaster risk reduction. An etymological journey. Natural Hazards and Earth System Sciences, 13, 2070–2716. https://doi.org/10.5194/nhess-13-2707-2013
Blandin, P. (2009). De la protection de la nature au pilotage de la biodiersité. Éditions Quae. https://doi.org/10.3917/quae.bland.2009.01
Bodin, P., & Wiman, B. (2004). Resilience and other stability concepts in ecology: Notes on their origins, validity and usefulness. The Earth System Science Bulletin, 2(2), 33–43.
Bodnar I. (2018). Aristotle's natural philosophy. In Edward N. Zalta (Ed.), The Stanford encyclopaedia of philosophy (Spring edition). https://plato.stanford.edu/archives/spr2018/entries/aristotle-natphil/
Botkin, D. (1990). Discordant harmonies. Oxford University Press.
Brand, S. F., & Jax, K. (2007). Focusing the meaning(s) of resilience: Resilience as a descriptive concept and a boundary object. Ecology and Society, 12(1), 1–16. https://doi.org/10.5751/ES-02029-120123
Brown, K. (2014). Global environmental change I: A social turn for resilience? Progress in Human Geography, 38(1), 107-117. https://doi.org/10.1177/0309132513498837
Burton, J. P., Jentsch, A., & Walker, L. R. (2020). The ecology of disturbance interactions. BioScience, 70(10), 854–870. https://doi.org/10.1093/biosci/biaa088
Carpenter, S., Walker, B., Anderies, J. M., & Abel, N. (2001). From metaphor to measurement: Resilience of what to what? Ecosystems, 4, 765–781. https://doi.org/10.1007/s10021-001-0045-9
Clements, F. (1916). Plant succession. An analysis of the development of vegetation. Carnegie Institution of Washington. https://doi.org/10.5962/bhl.title.56234
Cooper, G. (2001). Must there be a balance of nature? Biology and Philosophy, 16, 481–506.
Cuddington, K. (2001). The ‘Balance of Nature’ metaphor and equilibrium in population ecology. Biology and Philosophy, 16, 463–479. https://doi.org/10.1023/A:1011910014900
Davoudi, S. (2018). Just resilience. City Community, 17(1), 3–7. https://doi.org/10.1111/cico.12281
Delbrück, M. (1971). Aristotle-totle-totle. Of Microbes and Life, 50, 5.
Delettre, O. (2021). Identity of ecological systems and the meaning of resilience. Journal of Ecology, 109(9), 3147–3156. https://doi.org/10.1111/1365-2745.13655
Delettre, O., & Korniliou, A. (2022). From balance of nature to stability and resilience: Disuse and persistence. Philosophia Scientiae, 26(1), 53–72. https://doi.org/10.4000/philosophiascientiae.3346
Desjardins, E. (2015). Promoting resilience. The Quarterly Review of Biology, 90(2), 147–165. https://doi.org/10.1086/681439
Egerton, F. N. (1973). Changing concepts of the balance of nature. The Quarterly Review of Biology, 48(2), 322–350. https://doi.org/10.1086/407594
Eliot, C. (2011). The legend of order and chaos: Communities and early community ecology. Philosophy of Ecology, 49, 107.
Falk, D., Watts, A., & Thode, A. (2019). Scaling ecological resilience. Conservation and Restoration Ecology, 7, 275. https://doi.org/10.3389/fevo.2019.00275
Folke, C. (2006). Resilience: The emergence of a perspective for social-ecological systems analysis. Global Environmental Change, 16, 253–267. https://doi.org/10.1016/j.gloenvcha.2006.04.002
Gilbert, S. F. (2023). Pseudo-embryology and personhood: How embryological pseudoscience helps structure the American abortion debate. Natural Science. https://doi.org/10.1002/ntls.20220041
Gilbert, S. F., & Epel, D. (2015). Ecological developmental biology. Oxford University Press.
Gleason, H. (1926). The individualistic concept of the plant association. Bulletin of the Torrey Botanical Club, 53(1), 7–26. https://doi.org/10.2307/2479933
Golley, F. B. (1993). A history of the ecosystem concept in ecology. Yale University Press.
Gottlieb, P., & Sober, E. (2017). Aristotle on ‘Nature does nothing in vain.’ Hopos, 7(2), 246–271. https://doi.org/10.1086/693422
Grimm, V., & Wissel, C. (1997). Babel, or the ecological stability discussions: An inventory and analysis of terminology and a guide for avoiding confusion. Oecologia, 109, 323–334. https://doi.org/10.1007/s004420050090
Gunderson, L. H. (2000). Ecological resilience–in theory and application. Annual Review of Ecology and Systematics, 31, 425–439. https://doi.org/10.1146/annurev.ecolsys.31.1.425
Hassler, U., & Koher, N. (2014). Resilience in the built environment. Building Research and Information, 42(2), 119–129. https://doi.org/10.1080/09613218.2014.873593
Hilderbrand, R. H., Watts, A. C., & Randle, A. M. (2005). The myths of restoration ecology. Ecology and Society, 10(1).
Holling, C. S. (1998). Two cultures of ecology. Conservation Ecology, 2(2), 4. http://www.consecol.org/vol2/iss2/art4/
Hodgson, D., McDonald, J., & Hosken, D. (2015). What do you mean, ‘resilient’? Trends in Ecology Evolution. https://doi.org/10.1016/j.tree.2015.06.010
Holling, C. S. (1973). Resilience and stability in ecological systems. Annual Review of Ecology and Systematics, 4, 1–23. https://doi.org/10.1146/annurev.es.04.110173.000245
Holling, C. S. (1996). Engineering resilience versus ecological resilience. Engineering within Ecological Constraints, 31, 32.
Holling, C. S. (2001). Understanding the complexity of economic, ecological, and social systems. Ecosystems, 4, 390–405. https://doi.org/10.1007/s10021-001-0101-5
Holling, C. S., & Meffe, G. K. (1996). Command and control and the pathology of natural resource management. Conservation Biology, 10(2), 328–337. https://doi.org/10.1046/j.1523-1739.1996.10020328.x
Huneman, P. (2011). About the conceptual foundations of ecological engineering: Stability, individuality and values. Procedia Environmental Sciences, 9, 72–82. https://doi.org/10.1016/j.proenv.2011.11.013
Huneman, P. (2014a). Individuality as a theoretical scheme. I. Formal and material concepts of individuality. Biological Theory, 9(4), 361–373. https://doi.org/10.1007/s13752-014-0192-9
Huneman, P. (2014b). Individuality as a theoretical scheme. II. About the weak individuality of organism and ecosystems. Biological Theory, 9(4), 374–381. https://doi.org/10.1007/s13752-014-0193-8
Jablonka, E., & Lamb, M. J. (2005). Evolution in four dimensions. Genetic, epigenetic, behavioral, and symbolic variation in the history of life. Bradfrod Books/MIT Press
Jackson, L. (1992) The role of ecological restoration in conservation biology. In Fiedler, P. L., & Jain, S. K. (Eds), Conservation biology: The theory and practice of nature conservation, preservation, and management. (pp. 433-451). Chapman & Hall.
Jax, K., Jones, C. G., Pickett, J., & Pickett, S. (1998). The self-identity of ecological units. Oikos, 82(2), 253–264. https://doi.org/10.2307/3546965
Jørgensen, D., Nillson, C., Hof, A., Hasselquist, E., Baker, S., Chapin, S., Eckerberg, K., Hjaltén, J., Polvi, L., & Meyerson, L. (2014). Policy language in restoration ecology. Restoration Ecology, 22(1), 1–4.
Justus, J. (2007). Complexity, diversity, and stability. In S. Sarkar & A. Plutynski (Eds), A companion to the philosophy of biology. Wiley.
Justus, J. (2008). Ecological and Lyapunov stability. Philosophy of Science, 75(4), 421–436. https://doi.org/10.1086/595836
Keller, E. F. (2002). Making sense of life: Explaining biological development with models, metaphors and machines. Harvard University Press. https://doi.org/10.4159/9780674039445
Kingsland, S. (2005). The evolution of American ecology 1890–2000. John Hopkins University Press.
Klein, R. J. T., Nicholls, R. J., & Thomalla, F. (2003). Resilience to natural hazards: How useful is this concept? Environmental Hazards, 5, 35–45. https://doi.org/10.1016/j.hazards.2004.02.001
Kricher, J. (2009). The balance of nature. Princeton University Press. https://doi.org/10.1515/9781400830268
Kupiec, J. (1999). L’influence de la philosophie d’Aristote sur l’élaboration de la théorie de l’évolution et sur la génétique. Revue Européenne des Sciences Sociales, 37(115), 89–116.
Lewontin, R. (2000). The triple helix. Gene, organism, environment (pp. 10–24). Harvard University Press.
Love, A. C. (2010). Idealization in evolutionary developmental investigation: A tension between phenotypic plasticity and normal stages. Philosophical Transactions of the Royal Society B, 365(1540), 679–690. https://doi.org/10.1098/rstb.2009.0262
Maienschein, J. (2016). Embryos, microscopes, and society. Studies in History and Philosophy of Biological and Biomedical Sciences, 57, 129–136.
Marsh, G. P. (1964). Man and nature; or, physical geography as modified by human action. C. Scribner.
Martins, L. (2022). Wild by design. Harvard University Press.
Mathevet, R., & Bousquet, F. (2014). Résilience & environment. Penser le changement socio-écologiques. Libella
Mayr, E. (1982). The growth of biological thought. Belknam Press.
McDonnell, M. J., & Pickett, S. T. A. (1993). Humans as components of ecosystems: The ecology of subtle human effects and populated areas. Springer-Verlag. https://doi.org/10.1007/978-1-4612-0905-8
McIntosh, R. P. (1975). Gleason ‘Individualistic Ecologist’ 1882–1975: His contributions to ecological theory. Bulletin of the Torrey Botanical Club, 102(5), 253–273.
Mori, A. S. (2011). Ecosystem management based on natural disturbances: Hierarchical context and non-equilibrium paradigm. Journal of Applied Ecology, 48, 280–292. https://doi.org/10.2307/2484142
Morison, B. (2002). On location. Aristotle’s concept of place. Oxford University Press. https://doi.org/10.1093/0199247919.001.0001
Mumby, P., Wolff, N., Bozec, Y., Chollet, I., & Halloran, P. (2013). Operationalizing the resilience of coral reefs in an era of climate change. Conservation Letters, 7(3), 176. https://doi.org/10.1111/conl.12047
Odum E. (1953), Fundamentals of ecology. W. B. Saunders.
Palmer, M. (2016). Persistent and emerging themes in the linkage of theory to restoration practice. In M. Palmer, J. Zedler, & D. Falk (Eds), Foundations of restoration ecology. Island Press.
Palmer, M., Zedler, J., & Falk, D. (2016). Ecological theory and restoration ecology. In M. Palmer, J. Zedler, & D. Falk (Eds), Foundations of restoration Ecology. Island Press.
Pickett, S. T. A., Kolasa, J., & Jones, C. G. (1994). Ecological understanding. Academic Press.
Pickett, S. T. A., Kolasa, J., & Jones, C. G. (2007). Ecological understanding. The nature of theory and the theory of nature. Academic Press.
Pickett, S. T. A., & Ostfeld, R. S. (1995). The shifting paradigm in ecology. In R. L. Knight & S. F. Bates (Eds), A new century for natural resource management. (pp. 261–281). Island Press.
Pickett, S. T. A., & Parker, F. P. L. (1992). The new paradigm in ecology Implications for conservation biology above the species level. In P. L. Fiedler & S. K. Jain (Eds), Conservation biology. The theory and practice of natural conservation, preservation and management. (p. 102). Chapman and Hall.
Pickett, S. T. A., & White, P. S. (1985). The ecology of natural disturbance and patch dynamics. Academic Press Inc.
Pimm, S. L. (1991). The balance of nature? University of Chicago Press.
Reghezza-Zitt, M., Rufat, S., Djament-Tran, G., Le Blanc, A., & Lhomme, S. (2012). What resilience is not: Uses and abuses. Cybergeo, 621, 12. https://doi.org/10.4000/cybergeo.25554
Reid, R., & Botterill, L. C. (2013). The multiple meanings of ‘Resilience’: An overview of the literature. Australian Journal of Public Administration, 72(1), 31–40. https://doi.org/10.1111/1467-8500.12009
Ren, J., & Coffman, G. (2023). Integrating the resilience concept into ecosystem restoration. Restoration Ecology, 31, 5. https://doi.org/10.1111/rec.13907
Rhode, K. (2005). Nonequilibrium ecology. Cambridge University Press.
Rosenberg, A. (1997). Reductionism redux: Computing the embryo. Biology and Philosophy, 12, 445–470. https://doi.org/10.1023/A:1006574719901
Shugart, H. H. (2001). Succession, phenomenon of. In S. A. Levin (Ed.), Encyclopedia of biodiversity (pp. 541–552). Academic Press.
Simberloff, D. (1980). A succession of paradigms in ecology essentialism to materialism and probabilism. Synthese, 43(1), 3–39. https://doi.org/10.1007/BF00413854
Sober, E. (1980). Evolution, population thinking, and essentialism. Philosophy of Science, 47(3), 350–383. https://doi.org/10.1086/288942
Sperry-Taylor, A. T. (2021). Reassessing equilibrium explanations: When are they causal explanations? Synthese, 198, 5577–5598. https://doi.org/10.1007/s11229-019-02423-2
Stoddard, J. L., Larsen, D. P., Hawkins, C. P., Johnson, R. K., & Norris, R. D. (2006). Setting expectations for the ecological conditions of streams: The concept of reference condition. Ecological Applications, 16(4), 1267–1276. https://doi.org/10.1890/1051-0761(2006)016[1267:SEFTEC]2.0.CO;2
Strunz, S. (2012). Is conceptual vagueness an asset? Arguments from philosophy of science applied to the concept of resilience. Ecological Economics, 76, 112–118. https://doi.org/10.1016/j.ecolecon.2012.02.012
Sunding, K., Spotswood, E., Chapple, D., Beller, E., & Gross, K. (2016). Ecological dynamics and ecological restoration. In M. Palmer, J. Zedler, & D. Falk (Eds), Foundations of restoration ecology. (pp. 27–56). Island Press.
Tansley, A. (1935). The use and abuse of vegetation concepts and terms. Ecology, 16(3), 284–307.
Tobey, R. C. (1981). Saving the prairies. The life cycle of the founding school of American plant ecology, 1895–1955. University of California Press
Tsing, A. (2015). The mushroom at the end of the world. Possibility of life in capitalist ruins. Princeton University Press.
Turner, M. G. (2010). Disturbance and landscape dynamics in a changing world. Ecology, 91(10), 2833–2849. https://doi.org/10.1890/10-0097.1
Van Meerbeek, K., Jucker, T., & Svenning, J. (2021). Unifying the concepts of stability and resilience in ecology. Journal of Ecology. https://doi.org/10.1111/1365-2745.13651
Voigt, A. (2011). The rise of system theory in ecology. In A. Schwarz & K. Jax (Eds), Ecology revisited. Reflecting on concepts advancing science. Springer. https://doi.org/10.1007/978-90-481-9744-6_15
Walker, B., Holling, C. S., Carpenter, S. R., & Kinzig, A. (2004). Resilience, adaptability and transformability in social–ecological systems. Ecology and Society, 9(2), 1.
Whittaker, R. H. (1957). Recent evolution of ecological concepts in relation to the eastern forests of North America. American Journal of Botany, 44(2), 197–206.
Wiesner, K., & Ladyman, J. (2020), Measuring complexity. In Wiesner, K., & Ladyman, J., What is a complex system? Yale University Press.
Wolff, L. (2013). Concise review: Erythroid versus myeloid lineage commitment: Regulating the master regulator. Stem Cells, 31(7), 1237–1244. https://doi.org/10.1002/stem.1379
Wolpert, L., & Lewis, L. J. (1975). Towards a theory of development. Federation Proceedings, 34, 14–20. https://doi.org/10.1007/978-1-4684-2631-1_4
Worster, D. (1994). Nature’s economy. A history of ecological ideas (2nd ed., p. 12). Cambridge University Press.
Wu, J., & Loucks, O. L. (1995). From balance of nature to hierarchical patch dynamics: A paradigm shift in ecology. The Quarterly Review of Biology, 70(4), 439–466. https://doi.org/10.1086/419172
Zellmer, S., & Gunderson, L. (2008). Why resilience may not always be a good thing: Lessons in ecosystem restoration from Glen Canyon and the Everglades. Nebraska Law Review, 87(4), 893.
Acknowledgements
I thank the audience at the Labont Seminar and WIP seminar in Torino where the ideas proposed in this article were presented and discussed for the first time. I also thank Elena Casetta and Davide Vecchi for incisive feedback and suggestions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Barbara, L. Resilience and the shift of paradigm in ecology: a new name for an old concept or a different explanatory tool?. HPLS 46, 2 (2024). https://doi.org/10.1007/s40656-023-00600-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s40656-023-00600-8