
Abstract
Purpose
Liver-expressed antimicrobial peptide 2 (LEAP-2) has been recently identified as the endogenous non-competitive allosteric antagonist of the growth hormone secretagogue receptor 1a (GHSR1a). In rodents, LEAP-2 blunts ghrelin-induced feeding and its plasma levels are modulated in response to nutritional status, being decreased upon fasting and increased in high-fat diet (HFD) fed mice. Clinical data support the regulation of circulating LEAP-2 by nutrient availability in humans. In this work, our primary objective was to examine the chronic effects of ghrelin and LEAP-2 administration on food intake, adiposity, and energy expenditure in young mice subjected to standard and HFD at both room temperature and at thermoneutrality. Furthermore, we aimed to assess the impact of these two hormones on aging mice.
Results
Our results indicate that LEAP-2 produces a significant decrease of body weight and adiposity, an increase in energy expenditure, and activation of the thermogenic program in white and brown adipose tissue depots. However, this effect is not maintained under HFD or under thermoneutral conditions and is only partially observed in aging mice.
Conclusion
In summary our studies describe the central effects of LEAP-2 within distinct experimental contexts, and contribute to the comprehension of LEAP-2's role in energy metabolism.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
It is well established that the ghrelin system plays a key role in energy homeostasis and the evidence are as follows. Ghrelin administration elicits a marked increase in adiposity in rodents [1] and a potent orexigenic effect in almost all species tested, including humans [2, 3]. This orexigenic effect is specifically mediated by acylated ghrelin, which acts through specific hypothalamic receptors known as GHSR1a [4,5,6]. These receptors operate via well-defined signaling pathways that have been recently elucidated [7,8,9,10]. It is noteworthy that, GHSR1a can form heterodimeric complexes with receptors such dopamine 1 receptor (D1R), dopamine 2 receptor (D2R) and melanocortin-3-receptor (MC3R), all of which are implicated in the regulation of energy homeostasis [11,12,13].
The discovery of liver-expressed antimicrobial peptide 2 (LEAP-2), as a recently identified endogenous non-competitive allosteric antagonist of the GHSR1a, stands out as the most unexpected finding in the field related to the ghrelin system [14, 15]. LEAP-2 blunts ghrelin-induced intracellular calcium mobilization in vitro and ghrelin-induced GH secretion and food intake in vivo [14]. Interestingly, in relation to energy balance, circulating levels of ghrelin and LEAP-2 behave in opposite fashion in response to feeding and fasting [16].
The discovery of LEAP-2 as a new component of the ghrelin system is considered a major breakthrough, offering potential insights into the role of the ghrelin system in energy and metabolic homeostasis and paving the way for the development of more rational therapies. Recent data have shown that treatment of obesity is feasible through a unimolecular polypharmacology, which involves simultaneously targeting multiple receptors [17,18,19]. Whether blocking GHSR1a could further enhance this approach is unknown. However, before reaching this stage, it is widely acknowledged that more preclinical data are needed to assess the influence of factors such as diet, temperature and age, all of which may variably affect the biological effects of ghrelin and LEAP-2 on energy homeostasis [11,12,13, 20].
In this study, our objective was to investigate the impact of intracerebroventricular chronic administration of ghrelin and LEAP-2 on food intake, adiposity, and energy expenditure. It has been described that ghrelin’s central effects on nutrient intake and nutrient partitioning can be separated following exposure to HFD, indicating that they act through independent neuronal pathways [21]. Therefore, we conducted these experiments in mice that were maintained on either a standard diet or a high-fat diet. While the majority of mouse experiments are typically conducted at 22–24 °C, [22], a temperature well below their thermoneutrality zone, we also conducted an experiment at 30 °C to explore potential temperature-related effects. Furthermore, recognizing the age-dependent nature of ghrelin's biological effects on energy homeostasis, we conducted a specific study focusing on older mice.
Materials and methods
Animals
C57BL6j mice were housed in air-conditioned rooms (22–24 °C) under a 12:12 h light/dark cycle. After weaning, the mice were fed with standard chow (STD) or the high fat diet (HFD) (Research Diets D12,492; 60% fat, 5.24 kcal g−1, Research Diets, New Brunswick, NJ) for 15 weeks.
For aging experiments other group was maintained until 30 months of age.
After the experiment, the mice were sacrificed, and all tissues were rapidly explanted and snap frozen on dry ice. The tissues were collected and frozen at − 80 °C until they were used.
The animal procedures were conducted according to the principles approved by the Animal Care Research Bioethics Committee from University of Santiago de Compostela (license 15010/17/002); performed in compliance with the Directive 2010/63/EU and Spanish Royal Decree 53/2013, on the protection of animals used for experimental and other scientific purposes.
Intracerebroventricular (icv) chronic treatment of ghrelin and/or LEAP-2 on food intake and body weight
Brain infusion cannulas were stereotaxically placed into the lateral ventricle as previously described [23]. A catheter tube was connected from the brain infusion cannula to an osmotic minipump flow moderator (model 1002D, Alza Corp., Palo Alto, CA). Mice were then infused with either vehicle or ghrelin (Bachem) (5 nmol/day) or LEAP-2 (LEAP-2(38–77) peptide (Biorbyt) (2 nmol/day) or ghrelin + LEAP-2 for 7 days into the lateral ventricle as described previously [24, 25]. During this time, mice were motorized in body weight and food intake. The dose of ghrelin and LEAP-2 was chosen based on our experiments or a previous study [26,27,28].
Determination of energy balance
The animals were monitored in a custom 12-cage indirect calorimetry-, food intake-, and locomotor activity-monitoring system (LabMaster; TSE Systems; Bad Homburg, Germany) as described elsewhere [23].
For thermoneutrality experiments the mice were maintained to thermoneutral environment (30 °C with relative humidity of 45–52% in cages in LabMaster; TSE Systems; Bad Homburg, Germany).
For cold exposure and thermoneutrality, mice were fasted for 1 h before they were placed in a room at 4 °C. We measured the core body temperature at various time points before and during cold exposure using a rectal probe attached to a digital thermometer (Harvard Apparatus, Holliston, MA, USA).
Hormonal and biochemical assays
The levels of various compounds in plasma were measured by ELISAs. T4 (DRG, Germany), intra-assay and inter-assay variation coefficients was < 5% and < 8%, respectively. The assay sensitivity limit was 8 nmol/L, IGF1 (R&D Systems, USA) intra-assay and inter-assay variation coefficients was < 6% and < 10%, respectively. The assay sensitivity limit was 8.4 pg/mL, Leptin (Merk-Millipore) intra-assay and inter-assay variation coefficients was < 2% and < 5%, respectively. The assay sensitivity limit was 0.05 ng/mL and LEAP-2 (Human LEAP-2 [38–77] ELISA kit, Phoenix Pharmaceuticals, Inc) as described previously [29, 30], intra-assay and inter- assay variation coefficients was < 10% and < 15%, respectively. The assay sensitivity limit was 0.15 ng/ml. Triglycerides and cholesterol plasma levels were quantified by a homogeneous enzymatic colorimetric assay (Spinreact, S.A., Spain).
Haematoxylin–eosin staining and immunohistochemistry
Sections of 3 μm thick were made on a microtome and stained by the standard haematoxylin–eosin alcoholic procedure according to the manufacturer’s instructions (BioOptica, Italy). Immunohistochemical analysis was performed with an anti-UCP1 antibody (1:1000 dilution) (cat. #ab10983; Abcam, USA) as described [30].
Western blot analysis
Western blots were performed as previously described [25, 31] with antibodies against PPARγ (sc-7273) (Santa Cruz Biotechnology, USA); β3-AdR (ab694506), FGF21 (ab171941), PRDM16 (ab106410) and UCP1 (ab10983), FAS (ab128870), ACC (ab45174) (Abcam, USA); vinculin (clone hVIN-1) (Sigma-Aldrich cat# V9131); GAPDH (CB1001) (Millipore, USA). The protein levels were normalized to GAPDH or vinculin for each sample.
Quantitative RT-PCR analysis
Real-time PCR (TaqMan; Applied Biosystems, USA) was performed with specific primers and probes as previously described [32, 33]. Relative values were expressed in relation to hypoxanthine–guanine phosphoribosyl-transferase (Hprt, Mm03024075_m1) levels. We used the following commercially available and pre-validated TaqMan primer/probe sets (Applied Biosystems) for leap2 (Mm 00461982_g1).
Statistical analysis
Values were plotted as the mean ± SEM for each genotype. To test whether the data follows a Gaussian distribution, a normality test was performed (the Kolmogorov–Smirnov test for n between 5 and 7 or the Shapiro–Wilk test for n > 7). For normal distributions, a parametric test was conducted; for two‐group comparisons, the unpaired t‐test was carried out. In the statistical analysis data with a p‐value less than 0.05 were considered statistically significant. For a multiple comparison test, a one‐way ANOVA followed by Bonferroni´s post hoc multiple comparison test was performed. Data analysis was performed in GraphPad Prism Software Version 9.0 (GraphPad Software San Diego, CA, USA).
Results
Effect of chronic central infusion of LEAP-2 in STD diet on body weight, food intake adiposity and energy expenditure
As anticipated, ghrelin administration led to a notable increase in body weight compared to the vehicle-treated group. In contrast, mice treated with LEAP-2 exhibited a reduced weight gain (Fig. 1a). The co-administration of LEAP-2 with ghrelin attenuated the ghrelin-induced increase in body weight an effect that was independent of food intake (Fig. 1a, b). Food intake in LEAP-2 treated animals remained unaltered relative to controls, whereas it increased in ghrelin-treated animals. Circulating leptin levels, a surrogate marker of adiposity, remained unchanged in LEAP-2 treated animals compared to controls, while it suppressed the ghrelin-induced increase in leptin levels (Fig. 1c). Additionally, LEAP-2 administration led to a reduction in circulating triglyceride levels, while cholesterol levels remained unaffected (Fig. 1d), but co-administration of LEAP-2 with ghrelin failed to reverse the ghrelin-induced increase in cholesterol (Fig. 1d).

Central chronic LEAP-2 on a STD diet decreased body weight and increase energy expenditure. a Body weight change in mice with administration of LEAP-2, ghrelin, ghrelin and LEAP-2 or vehicle during 7 days. Data is expressed as mean ± SEM *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001 vehicle vs ghrelin; #p < 0.05, LEAP-2 vs vehicle; $p < 0.05, $$p < 0.01 LEAP-2 vs ghrelin + LEAP-2; &p < 0.05 ghrelin vs ghrelin + LEAP-2. b Accumulated food intake normalized to body weight during all treatment. c Circulating levels of leptin in all groups after 7 days of treatment. d Circulating cholesterol and triglycerides levels after 7 days of treatment. e Semiquantification of immunoblot analysis of de novo lipogenesis proteins FAS and ACC in WAT and representative immunoblot after 7 days treatment. f Energy expenditure corrected by body weight and locomotor activity during 48 h in control and LEAP-2 treated mice. Data is expressed as mean ± SEM *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001
In the white adipose tissue (WAT) LEAP-2 decrease fatty acid synthase (FAS), and acetyl CoA carboxylase 1 (ACC1) compared with control animals with statistically differences in ACC and clear tendency in FAS. However, LEAP-2 failed to reverse the ghrelin action in the expression of these enzymes when the LEAP-2 and the ghrelin are co-administered (Fig. 1e).
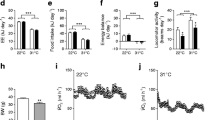
Because LEAP-2 administration reduced the body weight compared with control mice and to uncover the underlying mechanism responsible for the reduced body weight and adiposity, we assessed energy expenditure through indirect calorimetry in both control and LEAP-2 treated mice. This analysis revealed a significant increase in energy expenditure in mice treated with LEAP-2, evident in both light and dark phases, and this effect was not linked to changes in locomotor activity (Fig. 1f).
Chronic central infusion of LEAP-2 in STD diet-fed mice increases thermogenic markers in BAT and browning in scWAT.
We delved into the molecular mechanisms underlying the observed increase in energy expenditure. In line with the lower body weight and heightened energy expenditure in LEAP-2-treated mice, histomorphological analysis of brown adipose tissue (BAT) revealed smaller lipid droplets (Fig. 2a). Immunohistochemistry showed elevated uncoupling protein 1 (UCP1) levels in mice treated with LEAP- 2 (Fig. 2b). This result was corroborated by Western blot (WB) analysis, indicating increased protein levels of various thermogenic biomarkers, including UCP1, peroxisome proliferator-activated receptor gamma (PPARγ), and PR domain zinc finger protein 16 (PRDM16), indicative of heightened thermogenic activity (Fig. 2c).

Central chronic LEAP-2 on a STD diet increase thermogenic markers in BAT and browning in scWAT. a Representative Hematoxylin & Eosin staining of BAT of treated mice after 7 days of treatment. b Representative UCP1 immunohistochemistry and semiquantification of BAT after 7 days treatment. c Semiquantification of immunoblot analysis of thermogenesis proteins as UCP1, β3-adrenergic R, PRDM16, PPARγ in BAT and representative immunoblot. d Representative Hematoxylin & Eosin staining and size measurements of scWAT. e UCP1 immunohistochemistry of scWAT and semiquantification of UCP1. f Quantification of immunoblot analysis of thermogenesis proteins as UCP1, β3-adrenergic R, FGF21 and PPARγ in scWAT and representative immunoblot. Data is expressed as mean ± SEM. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001
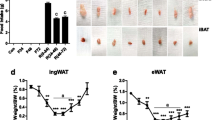
Subsequently, we investigated the subcutaneous white adipose tissue (scWAT). Ghrelin-treated animals exhibited expanded adipose tissue size, which was partially ameliorated when co-administered with LEAP-2 (Fig. 2d). This alteration mirrored characteristics commonly associated with brown adipose tissue (BAT) development, as corroborated by elevated UCP1 expression observed through immunostaining in LEAP-2-treated animals (Fig. 2e). Additionally, LEAP-2 administration induced a rise in β3-adrenergic receptor (β3AdrR) expression, but not UCP1, compared to controls. Notably, ghrelin displayed a non-statistically significant trend towards reduced β3AdrR expression. Moreover, LEAP-2 administration elicited a pronounced surge in fibroblast growth factor 21 (FGF21) expression (Fig. 2f).
In sum, LEAP-2 can influence energy balance, under normal dietary and housing conditions, through an enhancement of energy expenditure which is explained by increased BAT thermogenic activity and the browning of scWAT, a mechanism independent of thyroids hormones (Fig S1a).
Effect of chronic central infusion of LEAP-2 in Energy Homeostasis under Thermoneutral Conditions and cold exposure
We evaluated the effect of LEAP-2 treatment in a thermoneutral environment (30 °C) to eliminate the extra metabolism necessary to maintain body temperature at lower ambient temperatures.
A marginal, significant decrease in body weight gain was observed at the end of the experiment (day 7) (Fig. 3a). However, no significant differences were observed in final body weights (Fig. 3a) or food intake (Fig. 3b) in mice treated with LEAP-2 compared to controls. Of note, under thermoneutral conditions, energy expenditure and locomotor activity did not change with the treatment (Fig. 3c). These results indicate that chronic central infusion of LEAP-2 failed to exert a major impact on body weight, with significance observed only at day 7, and even then, the effects were less pronounced than those observed at 22ºC.

Effect of chronic central infusion of LEAP-2 in Energy Homeostasis under Thermoneutral Conditions and cold exposure. a Body weight change in mice with administration of LEAP-2 or vehicle during 7 days in thermoneutrality starting at day 1. b Accumulated food intake in relation of body weight during thermoneutrality. c Energy expenditure corrected by body weight and locomotor activity during 48 h in control and LEAP-2 treated mice. d Rectal temperature of mice treated with LEAP-2 or vehicle upon cold exposure (+ 4 °C) for 3 h. e Representative Hematoxylin & Eosin staining of BAT and representative UCP1 immunohistochemistry of treated mice after 3 h of 4ªC and semiquantification of UCP1 immunohistochemistry of BAT. f Representative Hematoxylin & Eosin staining of scWAT and scWAT adipocyte size of treated mice after 3 h of 4ªC. g Representative UCP1 immunohistochemistry and semiquantification of UCP1 immunohistochemistry of scWAT. Data is expressed as mean ± SEM *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001 vehicle vs LEAP-2
To gain deeper insight into the physiological relevance of LEAP-2 during cold adaptation, we conducted a cold tolerance test. At the 3-h mark of cold exposure, LEAP-2 treated mice appeared to better maintain their body temperature compared to controls (Fig. 3d). Histological analysis further revealed smaller lipid droplets in the BAT of LEAP-2 group, accompanied by a significant up-regulation of UCP1 protein as observed in immunostaining (Fig. 3e). Additionally, we conducted a histomorphological analysis of scWAT for study browning, which demonstrated a reduction in adipocyte size in LEAP-2-treated mice (Fig. 3f). Consistently, UCP1 immunostaining was increased in LEAP-2-treated animals compared to those in the control group (Fig. 3g). Altogether, these findings could suggest that LEAP-2 would improve the cold tolerance, at least partially, by increasing the browning of scWAT and the activation of BAT.
HFD fed mice are resistant to the weight reduction of central LEAP-2
As expected, our data indicated an increase in both circulating and liver mRNA expression of LEAP-2 in mice exposed to HFD for 12 weeks (Fig. S1c, d). Therefore, we decided to investigate the effect of chronic central infusion of LEAP-2 in diet-induced obese animals. In contrast to the data obtained in animals under standard diet (STD), we did not observe any difference in body weight gain (Fig. 4a), food intake (Fig. 4b) or adiposity as assessed by circulating leptin levels (Fig. 4c) between any of the treatments. Consistent with these findings, there were no changes in circulating levels of triglycerides or cholesterol among all the groups (Fig. 4d), and we also failed to detect changes in protein levels of FAS and ACC1 in WAT (Fig. 4e) or in energy expenditure and locomotor activity (Fig. 4f) in LEAP-2 treated mice.

LEAP-2 Resistance in Energy Homeostasis and Thermogenesis Program in Mice on a HFD. a Body weight change in mice on a HFD with administration of LEAP-2, ghrelin, ghrelin and LEAP-2 or vehicle during 7 days. b Accumulated food intake in relation of body weight during all treatment. c Circulating levels of leptin in all groups after 7 days of treatment. d Circulating cholesterol and triglycerides levels after 7 days of treatment. e Quantification of immunoblot analysis of de novo lipogenesis proteins FAS and ACC in WAT and representative immunoblot after 7 days treatment. f Energy expenditure corrected by body weight and locomotor activity during 48 h in control and LEAP-2 treated mice under HFD. Data is expressed as mean ± SEM #p < 0.05, ##p < 0.01, ###p ≤ 0.001 ghrelin vs ghrelin + LEAP-2; $p < 0.05, $$p < 0.01, $$$p ≤ 0. LEAP-2 vs ghrelin + LEAP-2. g Representative Hematoxylin & Eosin staining of BAT of treated mice after 7 days of treatment in mice on a HFD. h Representative UCP1 immunohistochemistry and semiquantification of UCP1 of BAT after 7 days treatment. i Quantification of immunoblot analysis of thermogenesis proteins as UCP1, β3-adrenergic R and FGF21 in BAT and representative immunoblot. j Representative Hematoxylin & Eosin staining, weight and adipocyte size measurements of scWAT. k Representative UCP1 immunohistochemistry and semiquantification of UCP1 of scWAT. Data is expressed as mean ± SEM. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001
As we expected, the histomorphological analysis revealed no major differences in BAT morphology (Fig. 4g) or UCP1 levels by immunohistochemistry (Fig. 4h) or Western blot (Fig. 4i). In relation of browning, we did not find any differences in scWAT histomorphology, adipocyte size (Fig. 4j) or UCP1 staining (Fig. 4k), indicating that under HFD, the mice exhibit resistance to the actions of LEAP-2. Overall, these findings confirm the resistance to adiposity actions for both hormones.
Effect of LEAP-2 on energy homeostasis and thermogenesis in aged mice
The relationship between ghrelin and GHS-R in aging has been reported, with GHS-R exerting opposite effects on WAT and BAT during aging. Ghsr−/− mice fed on regular chow have reduced fat mass and an improved lipid profile [34], highlighting the differential effects of ghrelin and GHS-R deficiency on body composition and energy homeostasis during aging [35]. We found increased circulating levels of LEAP-2 (Suppl. Figure 1d) and liver mRNA expression in aged animals (Suppl. Figure 1e).
Thereby, we decided to study the actions of LEAP-2 in aging mice. We chronically administered LEAP-2, ghrelin, ghrelin-LEAP-2 or vehicle intracerebroventricularly (icv) to 30-month-old mice. Compared to untreated control mice, LEAP-2 failed to produce changes in body weight (Fig. 5a), on the other hand, ghrelin administration led to a clear increase in body weight, which was blunted by co-administration of LEAP-2 (Fig. 5a). Both LEAP-2 and ghrelin failed to affect food intake (Fig. 5b). In aged mice, ghrelin treatment elevated circulating leptin levels, a marker of adiposity, while LEAP-2 administration decreased leptin levels. Co-administration of ghrelin and LEAP-2 effectively reversed the ghrelin-induced increase in leptin (Fig. 5c). This aligns with the observed reductions in circulating triglycerides and cholesterol compared to controls (Fig. 5d). Similarly, no significant effect of LEAP-2 was observed under basal or stimulated conditions on FAS or ACC (Fig. 5e).

Central chronic LEAP-2, ghrelin, ghrelin and LEAP-2 or vehicle in aging mice maintain thermogenic effect. a Body weight change during treatment (7 days) in mice with 30 months age old. b Accumulated food intake in relation of body weight during all treatment. c Circulating levels of leptin in all groups after 7 days of treatment. d Circulating cholesterol and triglycerides levels after 7 days of treatment. e Quantification of immunoblot analysis of de novo lipogenesis proteins FAS and ACC in WAT and representative immunoblot after 7 days treatment. f Energy expenditure corrected by body weight and locomotor activity during 48 h in control and LEAP-2 treated mice. g Representative Hematoxylin & Eosin staining of BAT of treated mice after 7 days of treatment in aging mice. h Representative UCP1 immunohistochemistry and semiquantification of UCP1 of BAT after 7 days treatment. i Quantification of immunoblot analysis of thermogenesis proteins as UCP1, β3-adrenergic R, FGF21, PRDM16 and PPARγ in BAT and representative immunoblot. j Representative Hematoxylin & Eosin staining, weight and adipocyte size measurements of scWAT. k Representative UCP1 immunohistochemistry and semiquantification of UCP1 of scWAT. Data is expressed as mean ± SEM. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001
Surprisingly, LEAP-2 treatment led to a slight decrease in energy expenditure (Fig. 5f), potentially contributing to the observed trend towards higher body weight compared to control mice. No changes in locomotor activity were observed (Fig. 5f). We assessed the potential effect of LEAP-2 in the BAT of aged animals. Histological analysis revealed smaller lipid droplets (Fig. 5g) and increased UCP1 immunolabelling (Fig. 5h). Protein levels of other thermogenic proteins, including PRDM16, PPARα, and FGF21 were elevated, with a clear trend for UCP1 (Fig. 5i), suggesting partial activation of the thermogenic program in LEAP-2-treated mice. Ghrelin administration resulted in an increase in adipocyte size and subcutaneous WAT weight (Fig. 5j). Administration of LEAP-2 alone or in combination with ghrelin significantly reduced subcutaneous WAT size under both basal and stimulated conditions. However, LEAP-2 treatment did not exhibit a significant effect on UCP1 levels under basal or stimulated conditions (Fig. 5k).
These findings suggest that in aged mice, central LEAP-2 administration counteracts ghrelin-induced weight gain by triggering thermogenic activity in BAT but does not induce browning of scWAT.
Discussion
This study demonstrated that the brain infusion of LEAP-2 reduces body weight in young mice and blocks ghrelin-induced weight gain in both young and old mice fed a chow diet. However, this effect is lost in diet-induced obese rodents. The effect on body weight was associated with a decrease in adiposity, as assessed by measuring leptin levels, as well as a decrease in basal triglyceride levels and ghrelin-induced cholesterol levels. Furthermore, our data indicate that the effect of LEAP-2 on body weight is associated with an increase in energy expenditure (EE). Indeed, we provide evidence that its stimulatory effect EE could be mediated by the increased activation of BAT and/or increased browning of scWAT. Taken together, this effect of LEAP-2 on EE provides further support for its role as a robust regulator of energy homeostasis. Our data showing this effect of LEAP-2 at the central level resembles that previously reported [36] and is consistent with data showing that neuronal knock down of the GHSR1a HFD-induced weight gain by increasing EE [37]. Hence, we decided to study in depth its effects taking into consideration the environmental temperature at which the animals were exposed to, their diet and their age. All these factors are considered highly relevant when assessing the role of ghrelin in relation to the different components of energy homeostasis [22]. This is necessary to reach firm conclusions regarding the homeostatic role of LEAP-2 and its potential as a therapeutic agent [22, 38].
The importance of housing temperature in comparative studies involving rodents and humans regarding energy homeostasis has been firmly established [39,40,41]. The concept of thermoneutrality in humans spans a range from 19 ºC to 30 ºC, indicating that individuals in developed societies are rarely exposed to temperatures below this range. In contrast, mice, which are typically housed at 22 °C, must increase their energy expenditure by 30% to maintain core body temperature [42]. Therefore, any studies investigating mechanisms related to energy homeostasis may be influenced by this phenomenon, particularly when considering that the activation of BAT and browning of WAT are regarded as potential strategies for preventing and treating obesity [43,44,45,46].
To address this issue, we conducted a study to assess the impact of LEAP-2 in mice at thermoneutrality. Our findings, demonstrating that LEAP-2 increased energy expenditure in animals at 22 ºC, suggest that it was reversing the inhibitory effect of the ghrelin system on BAT activation or browning, as indicated by indirect measurements based on the expression of UCP1. Conversely, our data reveal that the effect of LEAP-2 under thermoneutral conditions was significantly diminished, implying that the ghrelin system plays a minor role in suppressing energy expenditure at 30 ºC. This aligns with the concept that ghrelin can be considered a survival hormone [47]. Therefore, when animals are exposed to lower temperatures, such as 22 ºC, ghrelin suppresses thermogenesis in BAT and browning of scWAT to conserve energy. However, once animals reach thermoneutrality, the effect of ghrelin is substantially reduced, explaining the lack of impact of LEAP-2 under these conditions.
A strong interrelationship between the ghrelin system and exposure to a HFD is well-established. Genetic silencing of either ghrelin [48] or its receptor GHSR1a [37] prevents HFD-induced weight gain in mice exposed early in life. Additionally, the orexigenic effect of exogenously administered ghrelin is blunted in animals exposed to HFD, while ghrelin-induced adiposity is maintained [21]. On the other hand, it has been recently reported that mice exposed to HFD exhibit higher intrahepatic LEAP-2 synthesis and elevated circulating levels compared to chow diet, indicating overexpression of LEAP-2 in states of nutritional excess [49]. Similar data was obtained in humans when compared normal versus obese subjects, both children and adults [16, 29, 50]. Consistent with these findings, genetic deletion of LEAP-2 in male mice fed a standard diet exposed to acute peripheral administration of ghrelin exhibits increased food intake and increased GH secretion, but does not affect body weight and weekly food intake in male mice exposed to HFD [51] indicating a state of ghrelin resistance that could affect agonists and antagonists of the GHSR1a. Our data showing that in a similar experimental setting, icv administration of LEAP-2 failed to influence weekly food intake or energy expenditure in animals exposed to HFD, indicates that the reason of this resistance is unrelated to impairment of ghrelin or LEAP-2’s ability to cross the blood–brain barrier. Whether this alteration is related to changes in the expression of the GHSR1a or its signaling pathway at the central level remains to be established. Previous data assessing the effect of exposure to HFD for one week showed that ghrelin-induced food intake but not adiposity was impaired. Our data, obtain from animals exposed to chronic HFD for a more extended duration (15 weeks) showed that long-term exposure leads to an impairment of ghrelin and LEAP-2 effects in both food intake and adiposity.
In summary, the main effects of both ghrelin and LEAP-2 in energy homeostasis are largely lost upon chronic exposure to HFD.
A significant hallmark of aging is the association with decreased circulating levels of GH and insulin-like growth factor 1 (IGF-1) [52, 53]. However, the question of whether circulating ghrelin levels are altered during the aging process and to what extent aging leads to a state of ghrelin resistance remains a topic of debate [54]. Consequently, there is considerable interest in assessing the response to LEAP-2 in aged mice. Our data revealed that the administration of LEAP-2 alone in aged mice failed to produce any substantial effects on food intake or body weight, suggesting the presence of either reduced ghrelinergic activity or a state of ghrelin resistance. Notably, our findings indicate that the orexigenic effect of ghrelin is lost in aged animals, pointing towards a state of ghrelin resistance. Given that we administrated ghrelin directly at the central level, this resistance seems to be independent of ghrelin's ability to traverse the blood–brain barrier. Conversely, our data also demonstrated that ghrelin was capable of increasing body weight and adiposity, as evidenced by elevated leptin levels providing further support for the existence of distinct neuronal pathways governing these two effects. Intriguingly, while the effects of ghrelin on leptin were reversed when LEAP-2 was co-administered, it failed to modify FAS and ACC protein levels. It's also important to note that our study did not observe any discernible effects of ghrelin and LEAP-2 in aged animals in terms of their impact on biomarkers of BAT activity. This lack of effect could be explained by recent findings showing that aged mice produced much fewer beige adipocytes from these cells than their younger counterparts. The mechanisms involved could be related to aging-induced impairment of de novo beige adipogenesis from adipocyte stem and progenitor cells which is due to dysregulation of unknown signaling pathways since this impairment occurs in vivo but no in vitro [55]. Future investigations involving a comprehensive assessment of various components within the ghrelin system are warranted before mechanistic conclusions can be drawn.
In summary, our data continue to bolster the significance of LEAP-2 as a pivotal component within the ghrelin system. We present compelling evidence that both corroborates and extends our understanding of its interaction with ghrelin concerning various facets of energy homeostasis, encompassing food intake, energy expenditure, and adiposity in mice subjected to a standard diet [14]. Furthermore, our study introduces fresh insights into the central effects of LEAP-2 within distinct experimental contexts, each possessing high translational relevance. These contexts encompass exposure to a high-fat diet, thermoneutrality, and the aging process, thereby expanding our comprehension of LEAP-2's multifaceted role in regulating energy balance.
References
Tschop M, Dl S, Ml H (2000) Ghrelin induces adiposity in rodents. Nature 407:908–913. https://doi.org/10.1038/35038090
Mr D, Am W, Aj P, Je M, Patterson M, Frost G, Ma G, Small C, Bloom SR (2005) Ghrelin increases food intake in obese as well as lean subjects. Int J Obes 29:1130–1136. https://doi.org/10.1038/Sj.Ijo.0803001
Nakazato M, Murakami N, Date Y, Kojima M, Matsuo H, Kangawa K, Matsukura S (2001) A role for ghrelin in the central regulation of feeding. Nature 409:194–198. https://doi.org/10.1038/35051587
Ma B, Feighner Sd, Ss P, Kk M, Dl H, Mv S, Va W, Howard Ad, Van Der Ploeg Lh, Jv H (2000) Structure-function studies on the new growth hormone-releasing peptide, ghrelin: minimal sequence of ghrelin necessary for activation of growth hormone secretagogue receptor 1a. J Med Chem 43:4370–4376. https://doi.org/10.1021/Jm0001727
Howard Ad, Feighner Sd, Df C, Jp A, Pa L, Ci R, Hamelin M, Dl H, Oc P, Anderson J et al (1996) A receptor in pituitary and hypothalamus that functions in growth hormone release. Science 273:974–977. https://doi.org/10.1126/Science.273.5277.974
Jm Z, Je J, Ce L, Cb S, Jk E (2006) Expression of ghrelin receptor mrna in the rat and the mouse brain. J Comp Neurol 494:528–548. https://doi.org/10.1002/Cne.20823
Lopez M, Lage R, Ak S, Perez-Tilve D, Mj V, Varela L, Sangiao-Alvarellos S, Tovar S, Raghay K, Rodriguez-Cuenca S et al (2008) Hypothalamic fatty acid metabolism mediates the orexigenic action of ghrelin. Cell Metab 7:389–399. https://doi.org/10.1016/J.Cmet.2008.03.006
Muller Td, Nogueiras R, Ml A, Zb A, Anker Sd, Argente J, Rl B, Sc B, Cy B, Broglio F et al (2015) Ghrelin. Mol Metab 4:437–460. https://doi.org/10.1016/J.Molmet.2015.03.005
Romero-Pico A, Mj V, Gonzalez-Touceda D, Folgueira C, Kp S, Alvarez-Crespo M, Ma VG, Da V, Schwarzer C, Herzog H et al (2013) Hypothalamic kappa-opioid receptor modulates the orexigenic effect of ghrelin. Neuropsychopharmacology 38:1296–1307. https://doi.org/10.1038/Npp.2013.28
Da V, Martinez G, Romero A, Mj V, Boit Kd, Ig D-R, Lopez M, Vidal A, Nogueiras R, Dieguez C (2011) The central sirtuin 1/P53 pathway is essential for the orexigenic action of ghrelin. Diabetes 60:1177–1185. https://doi.org/10.2337/Db10-0802
Wellman M, Abizaid A (2015) Growth hormone secretagogue receptor dimers: a new pharmacological target. Eneuro. https://doi.org/10.1523/Eneuro.0053-14.2015
Kern A, Albarran-Zeckler R, He W, Rg S (2012) Apo-ghrelin receptor forms heteromers with drd2 in hypothalamic neurons and is essential for anorexigenic effects of drd2 agonism. Neuron 73:317–332. https://doi.org/10.1016/J.Neuron.2011.10.038
Rediger A, Cl P, Cx Yi, Tarnow P, Strotmann R, Gruters A, Krude H, Schoneberg T, Tschop Mh, Kleinau G et al (2011) Mutually opposite signal modulation by hypothalamic heterodimerization of ghrelin and melanocortin-3 receptors. J Biol Chem 286:39623–39631. https://doi.org/10.1074/Jbc.M111.287607
Ge X, Yang H, Ma B, Galon-Tilleman H, Chen P, Chen M, Js L, Wang Y, Dalmas O, Yin Y et al (2018) Leap2 is an endogenous antagonist of the ghrelin receptor. Cell Metab 27(461–469):E466. https://doi.org/10.1016/J.Cmet.2017.10.016
M’kadmi C, Cabral A, Barrile F, Giribaldi J, Cantel S, Damian M, Mary S, Denoyelle S, Dutertre S, Peraldi-Roux S, et al (2019) N-terminal liver-expressed antimicrobial peptide 2 (Leap2) region exhibits inverse agonist activity toward the ghrelin receptor. J Med Chem 62:965–973. https://doi.org/10.1021/Acs.Jmedchem.8b01644
Bk M, Puzziferri N, He Z, Ja R, Osborne-Lawrence S, Np M, Chhina N, Gaylinn B, Mo T, El T et al (2019) Leap2 changes with body mass and food intake in humans and mice. J Clin Investig 129:3909–3923. https://doi.org/10.1172/Jci125332
Rosenstock J, Frias J, Jastreboff Am DuY, Lou J, Gurbuz S, Mk T, Ml H, Haupt A, Milicevic Z et al (2023) Retatrutide, a gip, glp-1 and glucagon receptor agonist, for people with type 2 diabetes: a randomised, double-blind, placebo and active-controlled, parallel-group, phase 2 trial conducted in the usa. Lancet 402:529–544. https://doi.org/10.1016/S0140-6736(23)01053-X
Je C, Muller Td, Finan B, Dimarchi Rd, Tschop Mh, Da D (2023) Gipr/Glp-1r dual agonist therapies for diabetes and weight loss-chemistry, physiology, and clinical applications. Cell Metab 35:1519–1529. https://doi.org/10.1016/J.Cmet.2023.07.010
Muller Td, Tschop Mh (2022) Gut-hormone triple agonists: clinical safety and metabolic benefits. Lancet 400:1826–1828. https://doi.org/10.1016/S0140-6736(22)02350-9
Ferreira-Marques M, Carvalho A, Franco AC, Leal A, Botelho M, Carmo-Silva S, Aguas R, Cortes L, Lucas V, Real AC et al (2023) Ghrelin delays premature aging in hutchinson-gilford progeria syndrome. Aging Cell. https://doi.org/10.1111/Acel.13983
Perez-Tilve D, Heppner K, Kirchner H, Lockie Sh, Sc W, Dl S, Tschop M, Pfluger P (2011) Ghrelin-induced adiposity is independent of orexigenic effects. Faseb J: Off Publ Federation Am Soc Exp Biol 25:2814–2822. https://doi.org/10.1096/Fj.11-183632
Ac C, Dp B, Haman F, Richard D (2023) Brown adipose tissue-a translational perspective. Endocr Rev 44:143–192. https://doi.org/10.1210/Endrev/Bnac015
Imbernon M, Beiroa D, Mj V, Da M, Veyrat-Durebex C, Porteiro B, Diaz-Arteaga A, Senra A, Busquets S, Da V et al (2013) Central melanin-concentrating hormone influences liver and adipose metabolism via specific hypothalamic nuclei and efferent autonomic/jnk1 pathways. Gastroenterology 144(636–649):E636. https://doi.org/10.1053/J.Gastro.2012.10.051
Folgueira C, Beiroa D, Porteiro B, Duquenne M, Puighermanal E, Mf F, Barja-Fernandez S, Gallego R, Hernandez-Bautista R, Castelao C et al (2019) Hypothalamic dopamine signaling regulates brown fat thermogenesis. Nat Metab 1:811–829. https://doi.org/10.1038/S42255-019-0099-7
Imbernon M, Sanchez-Rebordelo E, Romero-Pico A, Kallo I, Mj C, Porteiro B, Al-Massadi O, Contreras C, Ferno J, Senra A et al (2016) Hypothalamic kappa opioid receptor mediates both diet-induced and melanin concentrating hormone-induced liver damage through inflammation and endoplasmic reticulum stress. Hepatology 64:1086–1104. https://doi.org/10.1002/Hep.28716
Zb A, Dm E, Beiler R, Cs C, Gi S, Tl H (2010) Uncoupling protein-2 decreases the lipogenic actions of ghrelin. Endocrinology 151:2078–2086. https://doi.org/10.1210/En.2009-0850
Lugilde J, Casado S, Beiroa D, Cunarro J, Garcia-Lavandeira M, Alvarez CV, Nogueiras R, Dieguez C, Tovar S (2022) Leap-2 counteracts ghrelin-induced food intake in a nutrient, growth hormone and age independent manner. Cells. https://doi.org/10.3390/Cells11030324
Pt P, Tr C, Km H, Strassburg S, Kruthaupt T, Chaudhary N, Halem H, Culler Md, Datta R, Burget L et al (2011) Ghrelin, peptide Yy and their hypothalamic targets differentially regulate spontaneous physical activity. Physiol Behav 105:52–61. https://doi.org/10.1016/J.Physbeh.2011.04.025
Barja-Fernandez S, Lugilde J, Castelao C, Vazquez-Cobela R, Lm S, Dieguez C, Leis R, Tovar S (2021) Circulating leap-2 is associated with puberty in girls. Int J Obes 45:502–514. https://doi.org/10.1038/S41366-020-00703-3
Cunarro J, Buque X, Casado S, Lugilde J, Vidal A, Mora A, Sabio G, Nogueiras R, Aspichueta P, Dieguez C et al (2019) P107 Deficiency increases energy expenditure by inducing brown-fat thermogenesis and browning of white adipose tissue. Mol Nutr Food Res 63:E1801096. https://doi.org/10.1002/Mnfr.201801096
Tovar S, Paeger L, Hess S, Da M, Ac H, Hs B, Hampel B, Pj A, Evers N, Buning H et al (2013) K(Atp)-channel-dependent regulation of catecholaminergic neurons controls bat sympathetic nerve activity and energy Homeostasis. Cell Metab 18:445–455. https://doi.org/10.1016/J.Cmet.2013.08.006
Perez-Sieira S, Lopez M, Nogueiras R, Tovar S (2014) Regulation of Nr4a By nutritional status, gender, postnatal development and hormonal deficiency. Sci Rep 4:4264. https://doi.org/10.1038/Srep04264
Persson-Augner D, Yw L, Tovar S, Dieguez C, Meister B (2014) Delta-Like 1 homologue (Dlk1) protein in neurons of the arcuate nucleus that control weight homeostasis and effect of fasting on hypothalamic Dlk1 Mrna. Neuroendocrinology 100:209–220. https://doi.org/10.1159/000369069
Lin L, Saha PK, Ma X, Henshaw IO, Shao L, Chang BH, Buras ED, Tong Q, Chan L, Mcguinness OP et al (2011) Ablation of ghrelin receptor reduces adiposity and improves insulin sensitivity during aging by regulating fat metabolism in white and brown adipose tissues. Aging Cell 10:996–1010. https://doi.org/10.1111/J.1474-9726.2011.00740.X
Ma X, Lin L, Qin G, Lu X, Fiorotto M, Dixit Vd, Sun Y (2011) Ablations of ghrelin and ghrelin receptor exhibit differential metabolic phenotypes and thermogenic capacity during aging. PLoS ONE 6:E16391. https://doi.org/10.1371/Journal.Pone.0016391
Lee Jh, Lin L, Xu P, Saito K, Wei Q, Ag M, Oy B, Pradhan G, Zheng H, Xu Y et al (2016) Neuronal deletion of ghrelin receptor almost completely prevents diet-induced obesity. Diabetes 65:2169–2178. https://doi.org/10.2337/Db15-1587
Jm Z, Nakano Y, Coppari R, Balthasar N, Jn M, Ce L, Je J, Ae D, Ar W, White Rd et al (2005) Mice lacking ghrelin receptors resist the development of diet-induced obesity. J Clin Investig 115:3564–3572. https://doi.org/10.1172/Jci26002
Mj B, Enerback S (2018) Targeting thermogenesis in brown fat and muscle to treat obesity and metabolic disease. Nat Rev Endocrinol 14:77–87. https://doi.org/10.1038/Nrendo.2017.132
Aw F, Cannon B, Nedergaard J (2018) Optimal housing temperatures for mice to mimic the thermal environment of humans: an experimental study. Mol Metab 7:161–170. https://doi.org/10.1016/J.Molmet.2017.10.009
Abreu-Vieira G, Xiao C, Gavrilova O, Ml R (2015) Integration of body temperature into the analysis of energy expenditure in the mouse. Mol Metab 4:461–470. https://doi.org/10.1016/J.Molmet.2015.03.001
Speakman JR, Keijer J (2014) Not so nuanced: reply to the comments of gaskill and garner on 'not so hot: optimal housing temperatures for mice to mimic the environment of humans’. Mol Metab 3:337. https://doi.org/10.1016/J.Molmet.2013.05.007
Ml R (2018) Of mice and men—environmental temperature, body temperature, and treatment of obesity. FEBS Lett 592:2098–2107. https://doi.org/10.1002/1873-3468.13070
Nedergaard J, Von Essen G, Cannon B (2023) Brown adipose tissue: can it keep us slim? A discussion of the evidence for and against the existence of diet-induced thermogenesis in mice and men. Philos Trans R Soc Lond B Biol Sci 378:20220220. https://doi.org/10.1098/Rstb.2022.0220
Aj W, Lopez M, Vidal-Puig A (2011) Using brown adipose tissue to treat obesity—the central issue. Trends Mol Med 17:405–411. https://doi.org/10.1016/J.Molmed.2011.04.001
Bartelt A, Heeren J (2014) Adipose tissue browning and metabolic health. Nat Rev Endocrinol 10:24–36. https://doi.org/10.1038/Nrendo.2013.204
Quarta C, Mazza R, Obici S, Pasquali R, Pagotto U (2011) Energy balance regulation by endocannabinoids at central and peripheral levels. Trends Mol Med 17:518–526. https://doi.org/10.1016/J.Molmed.2011.05.002
Bk M, Jm Z (2017) Ghrelin as a survival hormone. Trends Endocrinol Metab: Tem 28:843–854. https://doi.org/10.1016/J.Tem.2017.10.001
Ke W, Jp DR, Murray Jd, Garcia K, Iida K, Mo T, Mw S (2005) Absence of ghrelin protects against early-onset obesity. J Clin Investig 115:3573–3578. https://doi.org/10.1172/Jci26003
Ezquerro S, Tuero C, Becerril S, Valenti V, Moncada R, Mf L, Catalan V, Gomez-Ambrosi J, Mocha F, Silva C et al (2023) Antagonic effect of ghrelin and leap-2 on hepatic stellate cell activation and liver fibrosis in obesity-associated nonalcoholic fatty liver disease. Euro J Endocrinol/Euro Fed Endocr Soc 188:564–577. https://doi.org/10.1093/Ejendo/Lvad071
Ma X, Xue X, Zhang J, Liang S, Xu C, Wang Y, Zhu J (2021) Liver expressed antimicrobial peptide 2 is associated with steatosis in mice and humans. Exp Clin Endocrinol Diabetes 129:601–610. https://doi.org/10.1055/A-1210-2357
Shankar K, Np M, Singh O, Bk M, Osborne-Lawrence S, Varshney S, Gupta D, Sb O, Takemi S, Cp R et al (2021) Leap2 deletion in mice enhances ghrelin’s actions as an orexigen and growth hormone secretagogue. Mol Metab 53:101327. https://doi.org/10.1016/J.Molmet.2021.101327
Lanfranco F, Gianotti L, Maccario M (2003) Endocrine and metabolic alterations in obstructive sleep apnea syndrome. J Endocrinol Invest 26:491–492. https://doi.org/10.1007/Bf03345208
Sun Y, Jm G, Rg S (2007) Ghrelin and growth hormone secretagogue receptor expression in mice during aging. Endocrinology 148:1323–1329. https://doi.org/10.1210/En.2006-0782
Amitani M, Amitani H, Cheng KC, Kairupan TS, Sameshima N, Shimoshikiryo I, Mizuma K, Rokot NT, Nerome Y, Owaki T et al (2017) The role of ghrelin and ghrelin signaling in aging. Int J Mol Sci. https://doi.org/10.3390/Ijms18071511
Holman CD, Sakers AP, Calhoun RP, Cheng L, Fein EC, Jacobs C, Tsai L, Rosen ED, Seale P (2023) Aging Impairs Cold-Induced Beige Adipogenesis And Adipocyte Metabolic Reprogramming. Biorxiv. https://doi.org/10.1101/2023.03.20.533514
Acknowledgements
This work has been supported by grants from Ministerio de Economía y Competitividad (ST: PID2020-116741RB-I00; CD: PID2020-116628GB-I00) Xunta de Galicia (RN:HR18-00100), Centro de Investigación Biomédica en Red (CIBER) de Fisiopatología de la Obesidad y Nutrición (CIBERobn). CIBERobn is an initiative of the Instituto de Salud Carlos III (ISCIII) of Spain which is supported by FEDER funds.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Contributions
SC, MVM, TOD, CQV performed the in vivo experiments, analytical methods, collected and analyzed the data and draft the final figures. SC, MVM made substantial contributions to the acquisition and analysis of data. RN, CD and ST drafting the work and interpreted and revising it critically for important intellectual content. CD and ST made substantial contributions to the conception and design of the work, coordinated, secure the funding and led the project. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors have nothing to disclosure.
Ethical approval
The animal procedures were conducted according to the principles approved by the Animal Care Research Bioethics Committee from University of Santiago de Compostela (license 15010/17/002); performed in compliance with the Directive 2010/63/EU and Spanish Royal Decree 53/2013, on the protection of animals used for experimental and other scientific purposes.
Informed consent
For this type of study, no informed consent is required.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
40618_2024_2307_MOESM1_ESM.pdf
Supplementary file1 Supplementary figure 1: Variations of circulating and hepatic levels of LEAP-2 with diet and aged. a Circulating levels of T4 and IGF-1 in mice with administration of LEAP-2, ghrelin, ghrelin and LEAP-2 or vehicle during 7 days. b Circulating levels of LEAP-2 in mice under HFD during 20 weeks. c Liver leap2 mRNA expression of mice under HFD during 20 weeks. d Circulating levels of LEAP-2 in mice with 30 months of age. e Liver leap2 mRNA expression of in mice with 30 months of age. Data is expressed as mean ± SEM *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001. (PDF 64 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Casado, S., Varela-Miguéns, M., de Oliveira Diz, T. et al. The effects of ghrelin and LEAP-2 in energy homeostasis are modulated by thermoneutrality, high-fat diet and aging. J Endocrinol Invest 47, 2061–2074 (2024). https://doi.org/10.1007/s40618-024-02307-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40618-024-02307-4