
Abstract
The objective of this study was to evaluate the composition, diversity, and structure of tidal “Várzea” and “Igapó” forests in eastern Amazonia, Amapá, Brazil. All live tree individuals with diameter at breast height (dbh) ≥10 cm were registered. A total of 130 plots measuring 10 × 100 m were inventoried, distributed among 13 ha in each of the two forest typologies. A total of 10,575 trees were reported, belonging to 343 species, 172 genera, and 49 families. For all 26 ha sampled, mean tree density was 406 ± 61.27 trees ha−1 and mean basal area was 27.2 ± 11.13 m2 ha−1. Fabaceae, Arecaceae, Malvaceae, Meliaceae, and Rubiaceae were the most important families in tidal “Várzea”, together accounting for 74.76 % of the family importance value index (FIVI %). In “Igapó”, the most important families were Lecythidaceae, Euphorbiaceae, Malvaceae, and Arecaceae, which together accounted for 57.05 % of the family important value index (FIVI %). Smaller diameter trees measuring between 10 and 30 cm dbh dominated the landscape, accounting for 75.52 % of all individuals sampled. In general, 80 % (8285) individuals were under 24 m in height, while only 1.32 % of trees (140) reached heights above 34 m. There was evidence for statistically significant mean differences among tidal “Várzea” and “Igapó” with regard to the number of individuals, species, diversity, and tree height. However, no mean differences were detected for equitability, dbh, dominance, and basal area. Compositional patterns showed low similarity between the evaluated areas, indicating the existence of phytogeographic pattern based on species distribution.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
“Várzea” and “Igapó” forests are considered distinct floodplain habitats. The former covers an area of approximately 180,000 km2 and is characterized by low soil fertility and low productivity (Melack and Hess 2010). The latter covers approximately 300,000 km2 distributed along channel margins that drain the Andes and Andean foothills. Rivers of “Várzea” floodplains thus carry large suspended loads of nutrient-rich sediments (Melack and Hess 2010).
Many Amazonian wetland forests are affected by periodic inundation, resulting in cycles of terrestrial and aquatic phases. These flood regimes strongly influence vegetation composition and dynamics, thus providing a unique opportunity to further our understanding of ecological structure and function within periodically flooded systems (Junk et al. 1989). As a consequence of the often extreme environmental conditions, wetland organisms have developed a number of morpho-physiological coping strategies (Parolin et al. 2010).
Relatively few studies have compared “Várzea” and “Igapó”, e.g., Haugaasen and Peres (2006), Almeida and Jardim (2011), Ferreira and Parolin (2011), and Ferreira et al. (2013). Comparative studies in central and western Amazonia support the idea that “Várzea” is more species-rich (Haugaasen and Peres 2006; Wittmann et al. 2006). Indeed, “Várzea” is considered the most species-rich wetland tree community in the world, with well-over 1000 tree species (Wittmann et al. 2010).
The objective of this study was to compare the forest structure, composition, and diversity of tidal “Várzea” and “Igapó” in two watersheds of eastern Amazonia. The following hypotheses were tested: (1) the diversity and phytosociological patterns differ significantly between tidal “Várzea” and “Igapó” forests, and (2) the diversity and phytosociological patterns do not differ significantly between the flooded forests of this study in relation to other periodically flooded Amazon.
Materials and methods
Study area
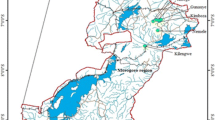
We studied two alluvial forests in different watersheds of eastern Amazonia, located in southern Amapá, northern Brazil. The first watershed represents tidal “Várzea” and is drained by the Mazagão river, located within the municipality of Mazagão between 00°14′06″ and 00°15′54″ latitude and 51°20′06″ and 51°23′42″ longitude. The second watershed represents “Igapó” and is drained by the Jari river, located in the municipality of Laranjal do Jari between 00°30′00″ and 01°06′00″ latitude and 52°18′00″ and 52°38′00″ longitude (Fig. 1). In both watersheds, the climate is tropical with a mean annual temperature of 27.5 °C and mean rainfall of 2158 mm (Vásquez and Rabelo 1999).

Locations of the investigated plots distributed among two watersheds representing “Igapó” and “Várzea” forests in eastern Amazonia, Brazil
The studied “Igapó” forest lies predominantly within the Guiana Shield, a geological formation of pre-Cambrian, crystalline bedrock underlying northern Amazonia. Soils in this geological region are predominantly sandy to clayey latosols (Vásquez and Rabelo 1999). Water supply to the local watershed is derived from local rainfall, without contribution from tidal flow. The studied tidal “Várzea” occurs within an estuarine complex, deriving its water supply from Andean runoff, innumerous tributaries, and local rainfall on the watershed. In addition, two daily tidal flows of predominantly freshwater affect the estuarine “Várzea”, thus giving rise to the name tidal “Várzea”. Soils are generally haplic gleysols with very clayey to silty texture (Queiroz and Machado 2008).
Floristic inventory and data collection
Vegetation was sampled in 26 1-ha plots (100 m × 100 m), divided into 13 ha for each habitat (“Várzea” and “Igapó”). Each plot comprised 10, 1000 m2 (10 × 100 m) subplots. Only live individuals with trunk diameters at breast height (dbh) ≥10 cm were considered. Each branch was reported in cases where individuals had multiple branches ≥10 cm dbh. All plots were georeferenced with a GPS. All individuals were tagged with aluminum plaques and identified in the field with the aid of parataxonomists. APG III (2009) was adopted for taxonomic classification. Voucher specimens of individuals that could not be identified in the field were collected for subsequent comparison at the Herbário Amapaense (HAMAB—IEPA). After determination to the lowest possible taxonomic rank, the vouchers were incorporated into the herbarium: numbers 18,185 to 18,201.
Data analysis
In order to elucidate the compositional differences between tidal “Várzea” and “Igapó”, for each community we ranked species and families from the most to the least important based on the summation of relative abundance, relative frequency, and relative basal area of each taxon. Importance values for each taxon are reported as a percentage of the total. Trends in compositional pattern were also assessed through a cluster analysis of plots by an average linkage method (UPGMA). The dissimilarity matrix used for clustering was based on pair-wise comparisons of plot composition using the Bray–Curtis similarity metric for abundance data.
To quantify vegetation structure of “Várzea” and “Igapó” habitat, for each plot we calculated stem density, mean basal area, and mean tree height. Also, to assess differences in diversity among habitats, plot-level diversity was estimated as Fisher’s α coefficient and species richness (Fisher et al. 1943). Additionally, species accumulation curves were compared for “Várzea” and “Igapó”. Mean differences in the plot-level structure and diversity measurements among the two habitats were assessed using t tests (Zar 2010).
To understand how structure and diversity metrics might vary within tidal “Várzea” and “Igapó”, we also assessed the mean differences among upstream, intermediate, and downstream sites within each habitat. For this, we used an analysis of variance and Tukey’s test for multiple comparisons. To check whether the assumptions of normally distributed data were fulfilled for use of parametric analyses, we used the Shapiro–Wilk test. We used PAST v.2.04 software for all analyses (Hammer et al. 2001).
Results
Vegetation composition, structure, and diversity
In total, we recorded 10,575 trees (dbh ≥ 10 cm) distributed among 343 species, 172 genera, and 49 families (Fig. 2). For “Várzea”, we recorded 5461 trees distributed among 98 species, 80 genera, and 29 families. For “Igapó”, we recorded 5114 trees distributed among 285 species, 150 genera, and 45 families.

Total number of individuals sampled in terms of species, genera, and families in the studied “Várzea” and “Igapó” forests located in eastern Amazonia, Brazil
In “Várzea”, the most important plant families were Fabaceae, Arecaceae, Malvaceae, Meliaceae, and Rubiaceae, which together accounted for 74.76 % of the family importance value index (FIVI). In “Igapó”, the most important families were Fabaceae, Lecythidaceae, Euphorbiaceae, Malvaceae, and Arecaceae, together accounting for 57.05 % of the family importance value index (FIVI). A total of 25 families (~51 %) are common to both “Várzea” and “Igapó”, but Fabaceae stands out as the most important family in both habitats (Table 1). Several rare families contributed only one individual to “Várzea” and “Igapó” each, including Amaryllidaceae, Araliaceae, Bixaceae, Convolvulaceae, Hypericaceae, Olacaceae, Salicaceae, and Violaceae.
Approximately 13 % (44) of the species occurred in both “Várzea” and “Igapó”. Among these, the most important were Pentaclethra macroloba (Willd.) Kuntze, Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg., Spondias mombin L., Virola surinamensis (Rol. ex Rottb.) Warb., and Mora paraensis (Ducke) Ducke (Table 2). The five most important species in “Igapó” and tidal “Várzea” accounted for 35.23 and 35.7 % of the cumulative importance for each habitat, respectively. The 15 most important species in “Igapó” accounted for approximately 50 % of total importance, while in “Várzea”, the 15 most important species accounted for 60 %. Species represented by a single individual accounted for about 20 % (n = 56) of species in “Igapó” and 17 % (n = 16) in tidal “Várzea”.
Diameter distributions in both tidal “Várzea” and “Igapó” showed an inverse J curve (Fig. 3). The smallest diameter class (between 10 and 30 cm) included the most individuals, accounting for 74.75 and 76.34 % of individuals in tidal “Várzea” and “Igapó”, respectively. Only 80 trees (0.76 %) had trunks greater than 90 cm dbh, with 56 (1.03 %) belonging to tidal “Várzea” and 24 (0.47 %) to “Igapó”.

Diameter distribution of dominant trees in tidal “Várzea” and “Igapó” in eastern Amazonia, Brazil
In “Igapó”, the three most important families accounted for 45 % (143.17 m2) of the total basal area, including Fabaceae, Lecythidaceae, and Euphorbiaceae. The species accounting for the largest proportion of “Igapó” basal area included P. macroloba (40.88 m2), H. brasiliensis (32.03 m2) and Attalea maripa (Aubl.) Mart. (13.39 m2). In “Várzea”, the three most important families, Fabaceae, Arecaceae, and Malvaceae, accounted for approximately 60 % (230.37 m2) of the total basal area. The species accounting for the largest proportion of tidal “Várzea” basal area included Mora paraensis, Pentaclethra macroloba, and Astrocaryum murumuru Mart. The relatively high basal areas of these species were due largely to their high abundance (Fig. 4).

Basal area of different diameter classes in tidal “Várzea” and “Igapó” in eastern Amazonia, Brazil
The height distribution curves of individuals showed different patterns for tidal “Várzea” and “Igapó” (Fig. 5). In “Várzea”, the number of individuals monotonically decreased with increasing height class. The largest proportion of individuals (53.74 %) were under 14 m, while 36.11 % were between 14 and 24 m, and 8.61 % were between 24 and 34 m in height. A total of 84 trees (1.54 %) had heights above 34 m, representing canopy—the emergents in tidal “Várzea”. These tidal “Várzea” emergents included species such as Mora paraensis (Ducke) Ducke, Manilkara huberi (Ducke) A. Chev., Calycophyllum spruceanum, Olmedia caloneura Huber., and Ceiba pentandra with heights between 35 and 43 m.

Distribution of individuals among four height classes in tidal “Várzea” and “Igapó” in eastern Amazonia, Brazil
In contrast, “Igapó” showed a monomodal height class distribution, with the largest fraction of individuals (49.51 %) in the second height class between 14 and 24 m in height. The lower height class accounted for 35.82 % of individuals, while larger height classes accounted for 13.57 and 1.09 % (56 emergent trees) of individuals. Emergents in “Igapó” include Ceiba pentandra, Hevea brasiliensis, and Goupia glabra Aubl. In general, individuals <24 m in height accounted for 80 % of all individuals in both tidal “Várzea” and “Igapó”.
For most structural variables assessed, there was more variation within tidal “Várzea” and “Igapó” habitats than between them. Mean tree density for all 26 ha was estimated to be 406.73 ± 61.27 trees ha−1. Tidal “Várzea” had 27 more trees ha−1 than “Igapó”, on average, but this difference was not statistically significant (P < 0.01) (Table 3). Mean basal area for all plots was 27.2 ± 11.13 m2 ha−1. Although tidal “Várzea” had a mean basal area 5.57 m2 ha−1 higher relative to “Igapó”, there was no evidence for consistent differences among habitats (P > 0.05). For mean tree height, “Igapó” trees were, on average, 2.26 m taller than those of tidal “Várzea” (P < 0.05).
Fisher’s α ranged from 8.46 to 16.53 (mean = 11.10 ± 2.1) for tidal “Várzea” and from 9.78 to 28.32 (mean = 18.54 ± 5.2) for “Igapó”, indicating higher mean alpha diversity for “Igapó” (P < 0.001) (Table 3).
Rarefaction curves also showed that “Igapó” is almost three times more species-rich than “Várzea” in the area investigated (Fig. 6). In tidal “Várzea”, the expected rate of new species was low. In contrast, in “Igapó”, new species were accumulated throughout. There was no evidence for differences in equitability (J) between tidal “Várzea” and “Igapó” (P > 0.05).

Expected species accumulation curves by plot, based on random samples of the data, for tidal “Várzea” and “Igapó” in eastern Amazonia, Brazil
In “Igapó”, comparison of upstream, intermediate, and downstream sites showed significant differences with regard to mean trunk diameter (P < 0.05), mean tree height (P < 0.05), basal area (P < 0.01), and the number of individuals (P < 0.001). However, there was no evidence for differences in Fisher’s alpha or plot richness (P > 0.05). For all variables assessed, except for tree density, upstream sites showed the highest values, while downstream sites showed the lowest.
In tidal “Várzea”, there was no evidence of mean differences in Fisher’s alpha, plot richness, and structural variables among upstream, intermediate, and downstream site classes. However, downstream sites had, on average, between 9 and 15 % less tree stems than upstream site classes (P < 0.001) and showed higher values for mean trunk diameter, height, and basal area (Table 4).
The dendrogram clusters revealed that the major division with regard to vegetation composition corresponded to the different habitats: tidal “Várzea” and “Igapó” (Fig.7). Moreover, the dendrogram aided in identifying the differences in compositional pattern within tidal “Várzea” and “Igapó”. Specifically, in “Igapó”, dendrogram clusters are concordant with upstream, intermediate, and downstream groups, indicating compositional change along the river course.

Dendrogram resulting of an UPGMA cluster analysis based on the Bray–Curtis dissimilarity matrix of all 26 plots studied in tidal “Várzea” and “Igapó” in eastern Amazonia, Brazil
Upstream sites in “Igapó” had more variable composition than both intermediate and downstream groups. In contrast, plot-wise comparisons in tidal “Várzea” all had similarities above 0.5, indicating less compositional variation among plots relative to “Igapó”. Unlike tidal “Várzea”, upstream, intermediate, and downstream sites did not form clusters as in “Igapó”.
It is worth mentioning that despite strong compositional differences, several species were widely distributed among most plots regardless of plot position along the river course or among dendrogram clusters. Among these, the most important are Pentaclethra macroloba, Gustavia augusta, Mora paraensis, Astrocaryum murumuru, Matisia paraensis, Carapa guianensis and Swartzia cardiosperma.
Discussion
The five most important plant families in each habitat accounted for about 82 and 60 % of trees in tidal “Várzea” and “Igapó”, respectively. They also accounted for the largest proportion of species richness, including 50 and 35 % of species reported for tidal “Várzea” and “Igapó”, respectively. Our study confirms the idea that few plant families and species dominate tropical forests (Wittmann et al. 2002, 2004; Wittmann and Parolin 2005; Parolin et al. 2010; Ferreira et al. 2010; Parolin et al. 2010). In the floodplains studied here, the patterns of dominance are even more severe, likely contributing to the over-dominant status reported for some families and species (Ter Steege et al. 2013).
Among the most important families occurring in both tidal “Várzea” and “Igapó” communities are Fabaceae, Arecaceae, Lecythidaceae, and Rubiaceae. Dominance of the Fabaceae was due to high species richness, abundance, and mean basal area. The family’s most important representatives include Pentaclethra macroloba in “Igapó” and Mora paraensis in tidal “Várzea”. These species have been reported in other floristic studies of eastern Amazonian tidal “Várzea” (Almeida et al. 2004) and “Igapó” (Ferreira et al. 2013), indicating a wider pattern of dominance beyond the samples of this study. The ranking of other families with regard to importance varies between tidal “Várzea” and “Igapó”.
We found that tidal “Várzea” and “Igapó” are similar with regard to several of the measured structural variables. For example, over 75 % of individuals had dbh between 10 and 30 cm, and the diameter class distribution observed in both habitats is similar, matching those found in other studies of “Várzea” and “Igapó” (Wittmann et al. 2002, 2004, 2013; Almeida et al. 2004; Wittmann and Parolin 2005; Haugaasen and Peres 2006; Santos and Jardim 2006; Carim et al. 2008; Ferreira et al. 2010; Almeida and Jardim 2011).
The inverted J-shaped distribution curve for dbh is characteristic for tropical forests (Haugaasen and Peres 2006; Carim et al. 2008, 2015) and indicates a positive balance between recruitment and mortality. We did not find evidence that the studied forests differed in terms of basal area and mean trunk diameter. Mean tree height was greater in “Igapó” than in tidal “Várzea”; however, in both habitats, the heights of majority of the individuals were less than 24 m. In tidal “Várzea”, the largest proportion of individuals were binned into the shortest height class, an understandable finding considering the limiting conditions of environmental instability, where specialized morpho-anatomical attributes are required for establishment (Wittmann et al. 2002, 2004; Wittmann and Parolin 2005; Parolin et al. 2010).
Tidal “Várzea” and “Igapó” showed similar physiognomic structure, distinguished only by a slight increase of mean height of trees of “Igapó”. Many studies point to ecological tradeoffs for species occurring in habitats with contrasting soil nutrient resources—one manifestation of these tradeoffs is the size of individuals, with smaller individuals expected in sites with lower nutritional resources (Fine et al. 2006; Targhetta et al. 2015). Contrary to these studies, we show that, despite large differences in soil fertility, the tidal “Várzea” and “Igapó” investigated here have similar physiognomic structure and comprise trees of similar size.
Carim et al. (2015) analyzed 14 ha of terra firme forest adjacent to the “Igapó” forest of this study, reporting similar findings with regard to composition, diversity, and structure. For example, Pentaclethra macroloba is among the 15 most important species in “Igapó” (this study) and that of Carim et al. (2015) terra firme study. Carim et al. (2008) studied 5 ha of tidal “Várzea” in a different location of the Amazonian estuary, finding practically the same results for vegetation structure, composition, and diversity as the tidal “Várzea” considered in the current study. Gama et al. (2005) compared the distribution of species in “Várzea” and “terra firme” in the State of Pará, reporting that 85.6 % of species were exclusive to terra firme, 5.2 % were exclusive to “Várzea”, and 9.2 % occurred in both terra firme and “Várzea”. These general findings have been corroborated by a number of authors (Almeida et al. 2004; Santos and Jardim 2006; Queiroz and Machado 2008; Carim et al. 2014).
Numerous studies point out the differences between “Várzea” and “Igapó” in the Amazon region. Some authors report greater richness and diversity in “Igapó” (Ferreira et al. 2010). On the other hand, the majority of comparative studies find that “Várzea” is richer in species, especially in central and western Amazonia (Haugaasen and Peres 2006; Wittmann et al. 2006). In fact, “Várzea” is widely considered the richest floodplain flora in the world, contributing more than 1000 tree species (Wittmann et al. 2010). We found a large difference with regard to species richness among habitats, with “Igapó” approximately three times richer than tidal “Várzea”. This finding stands as a striking contrast with most comparative studies (e.g., ter Steege et al. 2000; Wittmann and Junk 2003; Ferreira et al. 2010).
While the two systems we studied are completely different with regard to species richness and diversity, we found strong similarity in physiognomic structure. Conversely, other studies indicate minimal variation in composition, diversity, and structure within tidal “Várzea” and within “Igapó” ecosystems in the eastern Amazon. Composition, however, is likely to change at larger biogeographic scales including the central and western Amazon (Wittmann and Junk 2003). In sum, this study does not support the hypothesis that “Várzea” is more species-rich and diverse than “Igapó”.
In conclusion, this study is a preliminary attempt to compare “Várzea” and “Igapó” in eastern Amazonia. The comparison is based on a large, quantitative dataset that substantially increases our understanding of the plant diversity and structure of these environments, contributing essential data needed to make recommendations for the classification and management of Amazonian wetlands.
References
Almeida AF, Jardim MAG (2011) Florística e estrutura da comunidade arbórea de uma floresta de Várzea na Ilha de Sororoca, Ananindeua, Pará, Brasil. Sci For 39:191–198
Almeida SS, Amaral DD, Silva AS (2004) Análise florística e estrutura de florestas de Várzea no estuário amazônico. Acta Amazônica 34:513–524. doi:10.1590/S0044-59672004000400005
Angiosperm Phylogeny Group–APG (2009) An update of the Angiosperm phylogeny group classification for the orders and families of flowering plants: APG III. Bot J Linn Soc 161:105–121. doi:10.1111/j.1095-8339.2009.00996.x
Carim MJV, Jardim MAG, Medeiros TDS (2008) Composição florística e estrutura de floresta de Várzea no município de Mazagão, Estado do Amapá, Brasil. Sci For 36:191–201
Carim MJV, Abdon LM, Guimarães JRS, Tostes LCL (2014) Análise estrutural de açaizais nativos (Euterpe Oleracea Mart.) em floresta de Várzea, Amapá, Brasil. Biota Amazônia 4:45–51. doi:10.18561/2179-5746/biotaamazonia.v4n4p45-51
Carim MJV, Guimarães JRS, Tostes LCL, Takiyama LR, Wittmann F (2015) Composition, structure and floristic diversity in dense rain forest in the Eastern Amazon, Amapá, Brazil. Acta Sci 37:419–426. doi:10.4025/actascibiolsci.v37i4.27536
Ferreira LV, Parolin P (2011) Effects of flooding duration on plant demography in a black-water floodplain forest in central Amazonia. Pesqui Bot 62:323–332
Ferreira LV, Almeida SS, Parolin P (2010) Amazonian white-and black-water floodplain forests in Brazil: large differences on a small scale. Ecotropica 16:31–41
Ferreira LV, Parolin P, Cunha DA, Chaves PP, Leal D (2013) Variação da riqueza e composição de espécies da comunidade de plantas entre as florestas de Igapós e Várzeas da Estação Científica Ferreira Penna-Caxiuanã na Amazônia Oriental. Pesqui Bot 64:175–195
Fine LV, Miller ZJ, Mesones I, Irazuzta S, Appel HM, Stevens MHH, Sääksjärvi I, Schultz JC, Coley PD (2006) The growth-defense trade-off and habitat specialization by plants in Amazonian forests. Ecology 87:S150–S162. doi:10.1890/0012-9658(2006)87
Fisher RA, Corbet AS, Willians CB (1943) The relation between the number of species and the number of individuals in a random sample of an animal population. J Anim Ecol 1:42–58. doi:10.2307/1411
Gama JR, Souza AL, Martins SV, Souza DR (2005) Comparação entre florestas de Várzea e de terra firme do Estado do Pará. Rev Árvore 29:607–616. doi:10.1590/S0100-67622005000400013
Hammer O, Harper DAT, Ryan PD (2001) Palaetolongical statistics software package for education and data analysis. Palaetolongica Electron 4:1–9
Haugaasen T, Peres CA (2006) Floristic, edaphic and structural characteristics of flooded and unflooded forests in the lower Rio Purús region of central Amazonia, Brazil. Acta Amaz 36:25–36. doi:10.1590/S0044-59672006000100005
Junk WL, Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain systems. Canadian Special Publication of Fisheries and Aquatic Sciences. In: Dodge DP (ed) Proceedings of the International Large River Symposium. Can Spec Publ Fish Aquat Sci. 106:110–127
Melack JM, Hess LL (2010) Remote sensing of the distribution and extent of wetlands in the Amazon basin. Amaz Floodplain For 210:43–59. doi:10.1007/978-90-481-8725-6_3
Parolin P, Wittmann F, Schongart J (2010) Tree phenology in Amazonian floodplain forests. Amaz Floodplain For 210:105–126. doi:10.1007/978-90-481-8725-6_5
Queiroz JAL, Machado AS (2008) Fitossociologia em floresta de Várzea do estuário amazônico no estado do Amapá. Pesqui Florest Bras 57:5–20
Santos GC, Jardim MAG (2006) Florística e estrutura do estrato arbóreo de uma floresta de Várzea no município de Santa Bárbara do Pará, Brasil. Acta Amaz 36:437–446. doi:10.1590/S0044-59672006000400006
Targhetta N, Kesselmeier J, Wittmann F (2015) Effects of the hydroedaphic gradient on tree species composition and aboveground wood biomass of oligotrophic forest ecosystems in the central Amazon basin. Folia Geobot 50:185–205. doi:10.1007/s12224-015-9225-9
ter Steege H, Sabatier S, Castellanos H, Van Andel T, Duivenvoorden J, Oliveira AA, Ek RC, Lilwah R, Maas PJM, Mori SA (2000) An analysis of Amazonian floristic composition, including those of the Guiana Shield. J Trop Ecol 16:801–828. doi:10.1017/S0266467400001735
ter Steege H et al (2013) Hyperdominance in the Amazonian Tree Flora. Science 342:6156. doi:10.1126/science.1243092
Vásquez MP, Rabelo FG (1999) Sustainable management of na Amazonian Forest for timber production: a myth or reality? Plec News Views 12:20–28. doi:10.3362/9781780441092.017
Wittmann F, Junk WJ (2003) Sapling communities in Amazonian white-water forests. J Biogeogr 30:1533–1544. doi:10.1046/j.1365-2699.2003.00966.x
Wittmann F, Parolin P (2005) Aboveground roots in Amazonian floodplain trees. Biotropica 37:609–619. doi:10.1111/j.1744-7429.2005.00078.x
Wittmann F, Anhuf D, Junk WJ (2002) Tree species distribution and community structure of Central Amazonian Várzea forests by remote sensing techniques. J Trop Ecol 18:805–820. doi:10.1017/S0266467402002523
Wittmann F, Junk WJ, Piedade MTF (2004) The Várzea forests in Amazonia: flooding and the highly dynamic geomorphology interact with natural forest succession. For Ecol Manag 196:199–212. doi:10.1016/j.foreco.2004.02.060
Wittmann F, Schöngart J, Montero JC, Motzer T, Junk WJ, Piedade MTF, Queiroz H, Worbes M (2006) Tree species composition and diversity gradients in white-water forests across the Amazon Basin. J Biogeogr 33:1334–1347. doi:10.1111/j.1365-2699.2006.01495.x
Wittmann F, Schöngart J, Junk WJ (2010) Phytogeography, species diversity, community structure and dynamics of Central Amazonian floodplain forests. Amaz Floodplain For 210:61–102. doi:10.1007/978-90-481-8725-6_4
Wittmann F, Householder E, Piedade MTF, Assis RL, Schöngart J, Parolin P, Jun WJ (2013) Habitat specifity, endemism and the neotropical distribution of Amazonian white-water floodplain trees. Ecography 36:690–707. doi:10.1111/j.1600-0587.2012.07723.x
Zar JH (2010) Biostatistical analysis, 5th edn. Prentice-Hall, Upsaddler Road
Acknowledgements
We thank the Institute of Scientific and Technological Research of Amapá (IEPA) for logistical support and the Fundação de Amparo à Pesquisa do Estado do Amapá for financial support. We thank Embrapa Amapá for macronutrient analyses, the National Research Institute of the Amazon (INPA), and the Botany post-graduate course at INPA for providing facilities and the opportunity to conduct the research. MJVC was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
de Jesus Veiga Carim, M., Wittmann, F.K., Piedade, M.T.F. et al. Composition, diversity, and structure of tidal “Várzea” and “Igapó” floodplain forests in eastern Amazonia, Brazil. Braz. J. Bot 40, 115–124 (2017). https://doi.org/10.1007/s40415-016-0315-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40415-016-0315-6