
Abstract
The molecular cause of prostate cancer (PCa) is still unclear; however, its progression involves androgen, PI3K/Akt, and PTEN signaling, as cycle and apoptotic pathways. Alterations in oncogenes and tumor suppressor genes as PIK3CA, BRAF, KRAS and TP53 are not very common. Recently, somatic mutations have been discovered in relation to cancer progression mainly in genes such as PIK3CA; however, little data has been described in PCa. Nowadays genetic tools allow us to investigate multiple details about the biological heterogeneity of PCa, to better understand the mechanisms of disease progression and treatment resistance. Therefore, if the most relevant somatic mutations were included during screening, we could identify the best treatment for the right patient, bringing us closer to personalized medicine. The main objective of this article is to provide a review of the principal somatic mutations that appear to have a relevant role in hormonal cancers, like prostate cancer.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
This review covers the principal somatic mutations that may have an important role in PCa. |
Somatic mutations in androgen, PI3K/Akt, and PTEN signaling, like cycle and apoptotic pathways, can provide biomarkers useful for the prognosis and some treatment strategies in PCa. |
1 Introduction
A high percentage of PCa is considered sporadic, while a minority of cases are considered familial and hereditary PCa [1]. Sporadic PCa is caused mainly by somatic mutations, that, according to the NCI dictionary of cancer terms, are “alterations in DNA that occurs after conception and it can occur in any of the cells of the body except the germ cells (sperm and egg) and therefore are not passed on to children”. In the context of somatic mutation, the difference between hereditary and familial is that the familial PCa may be due to germline mutations (they are hereditary) and/or to somatic mutations caused by shared environmental and lifestyle factors by family members, while hereditary PCa is associated with germline mutations [2].
Advances in next-generation sequencing (NGS) will facilitate the discovery of multiple novel somatic mutations. Most of the work is focused on searching for new genetic variants that will offer relevant biomarker data, providing important prognostic and treatment response data [3]. Currently one of the principal goals in clinical medicine is the ability to offer personalized medicine and pharmacogenetic therapy to each patient. Already in tumors of metastatic colorectal cancer (CRC) patients, the status of v-Ki-ras2 Kirsten rat sarcoma viral oncogene homolog (KRAS) mutations are routinely determined, and patients with non-mutated KRAS are treated with cetuximab [a monoclonal antibody to estimated glomerular filtration rate (EGFR) receptor]. Almost all the biomarkers of treatment prediction response are genetic biomarkers; this highlights the relevance of a proper and universal standardization testing for somatic mutations [4].
Even though most cases of PCa are sporadic, by contrast, the majority of somatic mutations that appear in tumors have no impact or role in the progression of the cancer, they are known as passenger mutations [5]. For example, Fröhling et al., through high-throughput DNA sequence analysis and functional assays, identified five variants (T167A, V194M, Y364H, M737I, and G831E) of the fms related tyrosine kinase 3 (FLT3) gene identified in acute myeloid leukemia (AML) patients that do not increase kinase activity and do not contribute to leukemogenesis [6]. A minor percentage (<0.1%), known as “driver” mutations, provide selective advantages to cancer cells and have a relevant role in conferring the main characteristics of cancer development. Driver mutations occur in mutation driver genes, but these genes can also harbor passenger mutations. A total of 54 oncogenes and 71 tumor suppressor genes are considered mutation driver genes [5]. Somatic mutations could include punctual genetic changes (base substitutions, insertions, deletions, etc.) along with epigenetic alterations [7]. For example, miRNAs regulate gene expression at the post-transcriptional level and its deregulation affects critical cellular processes that contribute to the onset and progression of PCa [8].
Somatic mutations are also found in mitochondrial DNA (mtDNA), which also have a common role in many human cancers (breast, colorectal and ovarian cancer, among others) [9]. Mutations in the mtDNA appear to be a common event and are more frequent than mutations in autosomes in PCa [10]. These mutations could compromise the normal operation of the organelle, and mutations in tRNA could alter protein synthesis [11].
It is not clear how mtDNA mutations contribute to the onset and progression of cancer. Trying to clarify this point, Van Gisbergen et al. have proposed different models to explain how mutations in mitochondrial genes could cooperate in cancer. These models are not mutually exclusive; otherwise they could coexist in the same tumor [12]. However, mutations in the mitochondrial genome appear to be an early event in the carcinogenesis process being detectable before morphological alterations occur in the prostate tissue, thus, they are useful as biomarkers for diagnosing PCa in biopsy (confirmed by anatomopathological analysis) as evidenced by the use of the Prostate Core Mitomic Test™ (PCMT™). This test identifies deletions in mtDNA in the healthy tissue adjacent to tumor tissue in the biopsy. It has a sensitivity of 85%, thereby increasing tumor detection and reducing false negatives [13].
Although all cancers carry somatic mutations, most of them have been reported in CRC, especially in the BRAF, KRAS, PIK3CA and APC genes [14]. For example, somatic mutations have been related to a high incidence of metastatic disease and different treatment response has also been reported depending on the KRAS and BRAF mutation status [15, 16].
One of the main challenges of PCa is related to its high heterogeneity. This makes clinical stratification and selecting treatment strategies difficult. With the inclusion of expression patterns, molecular and genetic biomarkers in PCa, the aim is to create a specific classification profile to assess risk and treatment options [17]. In this regard, a correlation between the newly proposed grading of prognostic groups for PCa (based in Gleason grades) and genomic events has recently been established, so that the groups at risk of a biochemical recurrence are identified; those presenting with a higher frequency of polyploidy, as well as a higher frequency of point mutations in TP53, SPOP and ERG rearrangements [18]. It is very important to keep studying structural alterations such as gene fusion events, for example, the ETF gene family should be explored in different populations, to achieve better management and cancer screening programs in prostate cancer [19].
When talking about somatic mutations in PCa there are some candidate genes such as AR, TP53, KLF6, EPHB2, CHEK2, ZFHX3 (formerly known as ATBF1), NCOA2, PTEN, MYC, PIK3CA, FOXA1, KIT, and various histone-modifying genes [20]. But, there is not much information available for somatic mutations in tumors at the metastatic stage [21], in part, owing to the difficulty in accessing the target organs (bone is the primary site of metastatic PCa) and getting enough quality tissue to undertake the necessary studies [22]. Newly identified subsets of PCa have been related to mutations in the encoding SPOP gene, a class of cullin E3-ubiquitin ligase (Cullin E3) [20].
The major problem at this point is the high intra-tumoral heterogeneity and multifocality in primary tumors. That is why NGS needs to be used with hundreds tumors in order to create a better understanding of the evolution of prostate cancer [23]. One approach to this issue, which produced surprising results, was reported by Gundem et al. in April 2015 [24], which shines unprecedented light on the prostate cancer evolution process in different patients using NGS with sub-clones and metastases from the same tumor; also, the study of Cooper et al. [25] that has demonstrated the origin of multifocal disease from clonal expansion of mutations in morphologically normal tissue and the intra-tumoral heterogeneity by the coexistence of various tumor lineages with distinct ERG fusions in a tumor sample.
In 2015, the TCGA (the Cancer Genome Atlas) presented a complete analysis of more than 300 prostate carcinomas. These samples showed patterns defined by specific gene fusions or mutations. The result of the project in prostate cancer established a molecular taxonomy using data obtained from somatic mutations, gene fusions, somatic copy-number alterations, gene expression and DNA methylation analysis. The results revealed a molecular taxonomy in which 74% of the samples are included in seven subtypes defined on distinct oncogenic driver mutations or gene fusions (21 and 53% of the samples, respectively). Mutations in genes speckle-type POZ protein (SPOP) (11%), forkhead box A1 (FOXA1) (3%) and isocitrate dehydrogenase 1 (IDH1) (1%) and gene fusions in ERG (46%), ETV1 (8%), ETV4 (4%) and Friend leukemia virus integration 1 (FLI1) (1%). The results of this large analysis show molecular heterogeneity among primary prostate cancers and potentially actionable gene fusions and mutations [26].
This review is focused in the important effect of somatic mutations mainly in PCa, the effects of which remain undiscovered. The main aim is to try to classify somatic mutations in relation to changes in tumor cell activities to identify and prioritize those that generate functional changes and enhance tumor cell proliferation.
2 Methods
We performed a methodical literature examination from December 2012 to August 2016. Published research using somatic mutations in PCa and other cancers were selected by using databases (Pubmed, Scopus and Science direct). For inclusion in the searching process we also included related terms containing PCa, somatic mutations and TP53, KRAS, PIK3CA, APC, PTEN, EGFR, KIT, AR, SPOP, ETS and FOXA1 genes.
First, we performed a global search comprising somatic mutation in main genes such as TP53, KRAS, PIK3CA, APC, PTEN, EGFR, KIT, AR, SPOP, ETS and FOXA1 as well as other somatic mutations in cancer in general. Second, our search was restricted to the principal genes (TP53, KRAS, PIK3CA, APC, PTEN, EGFR, KIT, AR, SPOP, ETS and FOXA1) and PCa. Only papers published in English are included in the review.
3 Somatic Mutations
3.1 TP53 (Tumor Protein p53)
The TP53 gene is located on chromosome 17p13.1 codifying a 393 amino acid protein. The p53 tumor suppressor protein is a crucial factor for the preservation of a stable nuclear genome as well as for suppression of cancer. This gene is implicated in the activation of transcription factors in the presence of numerous types of cell stress situations (such as DNA damage, hypoxia, spindle damage, etc.) and exerts multiple, antiproliferative functions [27]. Genetic variations of TP53 contribute to human cancers in different ways; loss of TP53 gene function (>98%) or somatic mutations, Table 1. Most of the TP53 mutations (75%) are missense substitutions, and a minor proportion of frameshift indels (9%), nonsense (7%), and silent (5%) mutations [28].
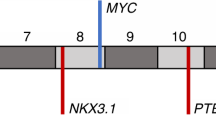
One of the principal targets for deactivating the cancer process is focused on the p53 protein, mainly due to its antiproliferative role in answer to stress situations [29]. It is related to metastasis, pathogenesis and progression in many cancers such as bladder, colorectal, and breast. Furthermore, there are also data about p53’s relevant role in the regulation of mtDNA [9]. As can be seen in Fig. 1, TP53 is included in the COSMIC database as one of the genes with the highest somatic mutation rate in PCa (138 above 1238 analyzed samples) [30]. Somatic mutations in the TP53 and AR genes, and other somatic point mutations in genes such as MTOR, BRCA2, ARHGEF12 and CHD5, can contribute to lethal PCa [21] associated with invasive metastasis [31].

Percentage of mutations in PCa and representation of the main somatic mutations. The table includes the percentage of samples with presence of mutations in the genes included in COSMIC “catalogue of somatic mutations in cancer”. A sample is examined through one or more genes for mutations. The selection parameters are “tissue type” prostate, “histology selection” carcinoma and “sub-histology selection” adenocarcinoma. CDS coding DNA sequence, Delet deletion, Inser insertion, Fram frameshift, n number of samples, Miss missense, Nons nonsense, Subst substitution. *The sum of number of mutated samples (n) do not match the number shown in the side tables because the same sample name can exist as separate entries. Date of COSMIC extraction: 09-12-2016
In PCa, abnormal p53 expression was related to increased risk of disease, as well as the development of distant metastasis [32], a high percentage of TP53 alterations were found in metastatic castration-resistant prostate cancer (mCRPC) samples compared to samples of primary tumors PCa [33]. P53 expression can be modified by miR-124 in a positive feedback loop with AR; miR-124 downregulates AR, which in turn downregulates miR-125b and this, upregulates p53 expression promoting apoptosis inhibition, but miR-124 is often repressed in PCa [34].
3.2 KRAS (v-Ki-ras2 Kirsten Rat Sarcoma Viral Oncogene Homolog)
KRAS gene on chromosome 12p12 .1 consists of six exons and spreads over 35 kb of genomic DNA. This gene encodes a protein that is a main component of Ras/MAPK signaling pathway. This protein’s main function is to interact with GTPase activating proteins (GAPs) and sending indications to nuclei.
In the Ras family, which comprises KRAS, NRAS and HRAS, mutations have been identified which encode for oncogenic proteins that promote tumorigenesis. There is little by way of pharmacological inhibition for these proteins [35].
Most of the mutations in KRAS affect codons 12, 13 and 61. Indeed they appear in many types of tumors, such as pancreatic cancers (with a mutation rate over of 90%) [36], where there is a major region of mutation in codon 12 [37]. Analysis of tissues revealed that some metastatic PCas harbored aberrations at the KRAS locus [38]. However, discrepancies in the frequency of mutation in relation to PCa have been reported across a range of geographical regions, races, and patient cohorts in KRAS [39]. For example, Wang et al. described a fusion protein whose expression exhibits transforming activity in some cells lines suggesting that this aberration may drive metastatic progression in a rare subset of PCa [38]. The protein Ras is important for local PCa growth, but it is not essential for tumor progression as it has a secondary role during the invasion and metastases processes [40]. However, it has been proven that activation of KRAS synergizes with overexpression of androgen receptor (AR) or the Akt signaling pathway and appears to significantly contribute to the progression of advanced PCa [41]. Recently it has been shown that approximately 40% of primary tumors and 90% of the metastatic tumors in PCa are altered in Ras signaling pathways [42]. KRAS expression is downregulated by miR-143 inhibiting Kras pathway, but this miRNA is frequently downregulated in PCa [8].
3.3 PIK3CA (The Phosphatidylinositol 3-Kinase Catalytic Subunit)
PIK3CA gene is on chromosome 3q26.32 and it is formed by 21 exons. One of the most common mutated oncogenes in breast cancer is the subunit p110 alpha of P13 kinase [43]. The main role of this kinase is to phosphorylate PTDs (protein transduction domains), and activate signaling cascades involved in cellular signaling in response to various growth factors and in other important cascades such as EGF, INS, IGF1, VEGFA and PDGF [44]. Most common mutations are theorized to activate the enzyme by two mechanisms: (i) release of the autoinhibition by the nSH2 domain of p85, and (ii) increase the interaction of the protein with the membrane that results in a greater accessibility of the enzyme to its substrate, a membrane component [45]. These somatic mutations are frequent in many different human diseases, many of them cancers; 43% of endometrial cancer, 4% of ovarian cancer and 2% of colon cancer [46].
However, it is not as common as in other tumors such as prostate, brain or pancreas [47–51]. In recent studies it was shown that aberrant activation of PI3K/Akt/mTOR has been implicated not only in the survival and metastasis of PCa cells but also in the development of drug resistance [52]. This is one of the signaling pathways frequently altered in patients with castration-resistant prostate cancer (CRPC), owing to amplifications of PIK3CA that give rise to a gene overexpression [33].
3.4 APC (Adenomatous Polyposis Coli)
APC is a tumor suppressor gene localized on chromosome 5q21, which encodes a protein with a relevant role in regulating proliferation and apoptosis. This protein regulates expression of β-catenin, both components of the Wnt pathway, and has several cellular functions such as adherence or stabilization in the cytoskeleton.
Approximately 90% of CRC present APC-inactivating mutations in somatic cells [53, 54]. Recent studies have detected various mutation types in CRC, for example missense, frameshift, deletions/insertions and stop/gain variants [55, 56]. Therefore, it is assumed that samples of CRC proliferation and tumor progression are affected by gene regulation or protein modifications [55]. Furthermore, mutations in the APC gene correlate with DNA breakage in late-replicating, low percentage GC and untranscribed regions of the genome [57]. In sporadic CRC, the loss of function of the APC protein is an early event already occurring during the formation of adenoma and cancerous lesions [58, 59]. It has also been identified in other cancer such as malignant mesothelioma where the mRNA of APC, Wnt4, Fzd3, sFRP4 and Axin2 was downregulated in relation to primary mesothelial cells [60].
As can be seen in Table 1, APC mainly plays a role in CRC and PCa. Wnt/β-catenin signaling is generally inactive in normal differentiated prostate cells but is crucial for regulating prostate development. Active Wnt/β-catenin signaling is associated with human PCa, and its progression. Approximately 20% of advanced prostate tumors and 85% of skeletal bone metastases have an elevated nuclear β-catenin expression [61]. The absence of APC expression may also be due to epigenetic regulation. APC is hyper-methylated and silenced in the tissue of poorly differentiated PCa, where their low level of expression is inversely correlated with the expression level of DNMT1, which is responsible of gene methylation [62].
3.5 PTEN (Phosphatase and Tensin Homolog)
PTEN is a tumor suppressor gene on chromosome 10q23.3 encoding a 403 amino acid. This protein is localized in cytoplasm and the nucleus, regulating cell cycle progression by maintenance of the G2/S cell cycle checkpoint, and prevents genomic instability by increasing the double strand break repair activity [63]. There are a wide range of studies focused on the relationship of PCa and mutations in PTEN [63], so it is interesting to view this gene when we study phenomena related to PCa. A relevant role in initiation, development and progression of PCa has been proven in recent studies [61, 64]. As can be seen in Fig. 1, there are data in the COSMIC database on the PTEN gene which indicate that it is one of the genes with the highest somatic mutation rate (133 above 1698 analyzed samples) [30]. PTEN inhibits the spread and migration of cells by regulating kinase adhesion and p53 protein activity [65]. As a consequence of PTEN’s regulation of the PI3K-AKT-mTOR pathway, the transcription of the GLUT1 gene is also affected. PTEN dysfunction increases the production of GLUT1, increasing the glucose intake and lactate production, which is called the Warburg effect. This effect is well known in cancerous metabolism as a great generator of energy to allow the cells to proliferate and spread easily [63]. In PCa, the PI3K/Akt pathway is one of the most commonly altered signaling pathways. PTEN loss and upregulation of the Akt pathway have begun to emerge as potentially important aberrations in PCa biology. Abnormalities of this pathway have been shown to induce proliferation in PCa [66]. MicroRNAs can down regulate the PTEN expression, like the miR-22 and miR-106b-25 cluster, which are overexpressed in human PCa. But there is a PTEN pseudogene (PTENP1) which can act as a target for PTEN’s miRNAs, inhibiting their action over PTEN’s regulation [63].
There are many studies indicating the role of miRNAs in the progression of PCa. miR-125b is found to have altered expression patterns of PTEN in PCa cells and tissues [64]. Likewise, miR-153 can suppress PTEN expression in PCa cells to promote cellular proliferation and it is upregulated in PCa [67].
3.6 EGFR (Epidermal Growth Factor Receptor)
The epidermal growth factor receptor (EGFR) has a relevant role in many tumorigenic processes, such as cell proliferation, survival, invasion and metastasis. EGFR is a member of the tyrosine kinase receptors ErbB family located on chromosome 7p12 and is responsive to extracellular ligands such as EGF and TGF-α [68].
Most of the regulatory effects on tumorigenesis (cell proliferation, differentiation, survival and migration) are controlled by the activation of the Ras-Raf-MEK- ERK1/2, STAT-3, STAT-5; and the PI3K/PTEN-Akt- mTOR cascades [65]. Activation of the EGFR oncoprotein is one of the principal events in human cancers. There are many alterations related to EGFR activation, like substitutions, indels (in kinase domain in cases of thyroid and lung cancers), high-copy amplifications (shown in epithelial, lung, gastrointestinal cancers, head, neck cancers and glioblastoma); or even overexpression rates of EGFR protein in others cancers (colorectal) [69]. Many studies suggest multiple roles for EGFR in developing prostate, mature prostate, and in androgen-responsive or in dependent malignant PCa, for example cellular invasion and bone metastasis. A number of results suggest that some relevant processes are mediated by EGFR activation in PCa including overexpression of mutation of the receptor, or its ligands; heterodimerization with other members of the ErbB receptor family, and transactivation by other receptors [70].
3.7 KIT (V-Kit Hardy-Zuckerman 4 Feline Sarcoma Viral Oncogene Homolog)
Normal cell migration and development is controlled by the KIT gene. This proto-oncogene is located on chromosome 4q12, and consists of 21 exons [71]. Most frequent KIT mutations are clustered at exons 11 and 17, but others in KIT exons 2, 8, and 9 or KIT-TK1 (exons 13 and 14).
Generally, KIT is inactive when its ligand is not present, but when KITLG/SCF binding occurs, there is a dimerization and activation on tyrosine residues of the kinase in an inactive conformation. When there are mutations, KIT activates the Ras/MEK/MAPK pathway and they produce cell proliferation, survival, differentiation, adhesion, and motility alterations [72].
c-Kit is a transmembrane receptor that is activated by the binding to its ligand (KL). Previous studies by Simak et al. [73] have shown that c-Kit is expressed in cultured prostate stromal cells. The same group showed that KL protein was expressed in normal prostate stromal cells as well as approximately 40% of PCa. Thus, there is abundant KL present in PCa from stromal cells or from cancer cells that can activate c-Kit [74]. There are previous recent studies reporting somatic mutations in KIT and PCa, but their presence in other cancers, such as leukemia, may be a clue to follow through the study of different kind of neoplasias as PCa [75].
3.8 AR (Androgen Receptor)
AR gene is on chromosome Xq12 and it is composed of 919 amino acids. Androgens are required for prostate differentiation and growth, playing an important role in carcinogenesis. Androgens act via the AR that performs a relevant role in steroid hormone production and forms part of one of the most important activated signaling pathways in PCa [76]. Antiandrogen therapy is one of the most effective treatments for PCa [77]. Moreover, AR is implicated in normal prostate growth and differentiation as well as in treatment response and metastases [78]. PCa has many genetic alterations involved in its survival, some of them related to PI3K and AR pathways [79]. Any alteration of AR activation in PCa AR-independent could be related to somatic or germline mutations. According to the latest update published in 2012 by Androgen Receptor Gene Mutations Database, 159 different mutations in the AR have been identified, most of them somatic mutations. The most common site of mutations is the ligand-binding domain (LBD), where around half of the mutations have been found; followed by exon 1, wherein 30% of the mutations of AR occur [80]. Although, there is not much detailed information about mutations in AR gene, mainly due to its large size (around 90 kb), they are present in coding (rs192696, rs1926927) and noncoding sequences; and they are often missense mutations. AR mutations appear to be a late event in the PCa, since metastatic tumors have high percentages of AR alterations in relation to primary tumors [26]. In the case of somatic mutations, there are some previously described, such as the R726L mutation, although it is a germline mutation, it has also been identified in several sporadic cases of prostate cancer in Finland [81]. Other cases also report point mutations, G2T and C214A in 5′-UTR (noncoding) region and an AR-Q798E mutation in PCa patients [82]. Recent studies have discovered the AR P340L mutation in PCa that seems to reduce the transcriptional response to ART-27 (AR N-terminal coactivator associated with AR-mediated growth inhibition) [83]. One of the most commonly identified somatic mutations is T877A, located in the LBD, producing a receptor capable of promiscuous binding and activation by a variety of steroid hormones and ligands including estrogens, progestins, glucocorticoids, and several anti-androgens [84, 85]. Other AR somatic mutations, such as L702H, also allow activation by glucocorticoids, which are usually administered by the antiandrogen, abiraterone acetate, reducing its effectiveness [86]; or the F876L mutation, that seems to be able to void the antagonistic activity of the androgen enzalutamide [87]. Many of the recent studies are focused on truncated forms of the androgen receptor, such as ARV-7 and its relationship with responses of the anti-androgen treatments [88]. This splice variant AR is resistant to degradation mediated by SPOP [89] and even though it lacks the LBD, in the absence of androgens, it can activate AR target genes [90]. One of the possible implications of somatic mutations in the AR gene could be to reduce or not produce AR activity at low hormone levels [91]; however, there is no clear current data on the function of these somatic mutations in cancer.
miRNAs play an important role in regulation of AR expression and AR pathway. Among them, miR-34a, mR-34c, miR-205 and miR-124, suppress AR expression by direct targeting of AR [92]. Other miRNAs indirectly regulate AR activity, such as let-7, that inhibits AR expression by targeting Myc [93] or miR-21 that can be directly regulated by AR, promoting the proliferation and tumor growth [94].
4 Other Somatic Mutations
4.1 SPOP (Speckle-Type POZ Protein)
This gene is located at chromosome 17q21.33 [95] encoding a substrate for the Cullin-based E3 ubiquitin ligase [96]. The SPOP protein consists of a BTB domain for binding Cul3 [97]. Around 15% of the somatic mutations of PCa tumors are related to SPOP gene, but in CRC or gastric cancers, these are rare events [98]. In PCa, SPOP mutations affect the substrate-binding region, altering substrate interactions [99]. SPOP mutations alter the DNA double strand break repair mechanism, contributing to genomic instability and increased genomic rearrangements in PCa [100]. According to Kim et al. [97] the mutation p.Phe133Leu is relevant for PCa development. Recently SPOP was related to AR interactions, with SPOP mutations avoiding AR degradation and developing a relevant role in PCa progression and carcinogenesis [98]. In addition, SPOP mutations prevent the degradation of ERG oncoprotein [101], which is involved in cellular processes such as cell migration and invasion [102].
4.2 FOXA1 (Forkhead Box A1)
FOXA1 belong to forkhead class of DNA-binding proteins. FoxA proteins allow access by other transcription factors to condensed chromatin [103]. FOXA1 proteins have direct interactions with AR and are involved in transcriptional activity of androgen-regulated genes. Moreover, increased expression of FOXA1 is related to PCa tumorigenesis and progression. FOXA1 is one of the major genes mutated in primary prostate tumors. In fact, in a recent study, primary tumors have been classified into seven molecular subtypes based on oncogenic profile drivers, patients with FOX1A being one of them. The molecular subtype of primary tumors with FOXA1 mutations also showed SPOP mutations but not mutations in any of the other genes that defined subtypes, elevated AR-mediated transcription and uniform epigenetic profiles [26].
4.3 ETS Gene Fusions
There are several alterations like gene fusions with erythroblast transformation-specific family of transcriptions factor (ETS), which regulate AR and affect PCa. The most common of these include the TMPRSS2 gene, an androgen-regulated gene transmembrane protease serine 2 and ERG oncogene, resulting in different TMPRSS2-ERG gene fusions [104]; rearrangements that occurs in approximately 50% of prostate tumors [105]. The fusion of both genes causes overexpression of ERG under hormonal stimulation due to androgen-responsive promoter elements of TMPRSS2 [102]. Furthermore, this fusion gene can express a truncated protein ERG, that is resistant, in most cases, to degradation by SPOP [101]. TMPRSS2-ERG gene fusions and SPOP mutation, are both mutually exclusive, so they could represent different molecular subtypes [106]. While TMPRSS2 was the most frequent fusion partner in all ETS fusions, it has identified fusions with other previously described androgen-regulated 50 partner genes, including SLC45A3 and NDRG1 [26].
5 Conclusions
We have focused our analysis on the BRAF, PIK3CA, TP53, KIT, AR, EGFR, KIT, APC, KRAS, PTEN, SPOP and FOXA1 genes, which were described in other hormonal-dependent cancers which have a high similarity to PCa. Details of somatic mutations could be vital for the customization of medical decisions, practices, and drug administration. For example, recent publications have identified multiple concordant somatic mutations in cfDNA and primary tumor samples and in cfDNA and metastatic tumor samples from one patient. Using somatic mutations as genetic biomarkers in esophageal squamous cell carcinoma, has shown the possibility of diagnosing tumor recurrence with greater accuracy than using standard tumor markers or imaging methods [107]. Data has also proved that phenotypic changes, such as a partial or complete epithelial to mesenchymal transition (EMT), which play important roles in survival and proliferation, and development of resistance to therapeutic treatments in PCa, are thought to to arise due to somatic mutations in the genome [108].
Somatic genetic alterations can cause differences in histopathology, gene expression, gene amplifications and deletions. The interaction between germline genetic variations [insertions and deletions, single-nucleotide polymorphisms (SNPs), copy number variants, mini- and microsatellites] and somatic alterations can influence the clinical outcome of cancer. Somatic point mutations in PCa may be rare relative to other tumor types such as glioblastoma, lung cancer and melanoma [55] and this could be one of the main reasons for the abundance of data in these cancers. Clinical relevant alterations in CRPC (castration-resistant prostate cancer) include defects in DNA damage repair (at either the somatic or germline level) in up to 20% of patients (with implications for PARP1 inhibitor therapy), PI3K/PTEN/Akt pathway activation, WNT signaling pathway alterations, cell cycle gene alterations, and less common but potentially targetable alterations involving RAF and FGFR2. Somatic aberrations involving DNA defect repair genes [such as deletions/mutations involving BRCA1/2, ataxia telangiectasia mutated (ATM), CHEK2, Fanconi (FANC) genes] are present in up to 20% of CRPC tumors. Somatic loss of the BRCA2 gene, for instance, occurs in approximately 3% of localized prostate cancer cases (TCGA) and 13% of CRPC cases (SU2C-PCF) [109].
The important role of somatic mutations has been proven in cancers such as colorectal, non-small cell lung (NSCLC) and breast cancer. In the case of NSCLC, some studies have confirmed EGFR mutations as predictive biomarkers of treatment response to tyrosine kinase inhibitors, gefitinib and erlotinib [110]. For that reason, the screening in EGFR mutations is performed before offering the drugs to the patients. Similar results have been obtained in KRAS mutations in patients of CRC. It is proven that KRAS mutations confer resistance to treatment with EGFR antibodies and only patients with wild-type KRAS tumors obtain benefit from these agents [4].
Personalized screening may potentially confer additional benefits. It can detect cancer in younger subjects at high risk. Prostate and breast cancer detected in younger subjects tends to behave more aggressively. If a high polygenic risk is associated with disease aggressiveness, then potentially additional life years would be gained by early detection of cancer in younger subjects [111]. Nowadays, in respect to PCa the main strategies in personalized medicine are focused on a gene-based approach to prostate cancer prevention; specifically, in susceptibility alleles in genes BRCA1, CHEK2, NBS1 and HOXB13, that can improve the performance of the PSA test in a population-based setting [112]. So, if we add the most relevant somatic mutations in the screening offered to the personalized medicine, we could improve the genetic information, creating a more specific genetic profile in relation to the cancer and achieving the aim of the right treatment for the right patient at the right time [113].
In conclusion, some somatic mutations can interact with tumor suppressor gene mutations or other cancer factors and they are considered as risk factors. In PCa, although it is one of the most prevalent cancers worldwide, and it has a complex genetic landscape, we hypothesis that somatic mutations could also confer risk in its progression and development. A recently published study suggested that BRCA2-disrupted tumors represent a unique and clinically relevant molecular subtype of aggressive PCa, highlighting both the promise and utility of this mutation signature as a prognostic and treatment-selection biomarker [114]. However, at the moment, data are scarce in this cancer compared to others. One of the main reasons could be the heterogeneity of this cancer, manifested histologically as multifocal or unifocal PCa, and the difficulty in detecting these kinds of mutations in this complex tissue.
References
Kral M, Rosinska V, Student V, Grepl M, Hrabec M, Bouchal J. Genetic determinants of prostate cancer: a review. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2011;155(1):3–9.
Ostrander EA, Markianos K, Stanford JL. Finding prostate cancer susceptibility genes. Annu Rev Genomics Hum Genet. 2004;5:151–75.
Paolillo C, Londin E, Fortina P. Next generation sequencing in cancer: opportunities and challenges for precision cancer medicine. Scand J Clin Lab Invest Suppl. 2016;245:S84–91.
Aung KL, Board RE, Ellison G, Donald E, Ward T, Clack G, et al. Current status and future potential of somatic mutation testing from circulating free DNA in patients with solid tumours. Hugo J. 2010;4(1–4):11–21.
Vogelstein B, Papadopoulos N, Velculescu VE, Zhou S, Diaz LA, Kinzler KW. Cancer genome landscapes. Science. 2013;339(6127):1546–58.
Fröhling S, Scholl C, Levine RL, Loriaux M, Boggon TJ, Bernard OA, et al. Identification of driver and passenger mutations of FLT3 by high-throughput DNA sequence analysis and functional assessment of candidate alleles. Cancer Cell. 2007;12(6):501–13.
Pleasance ED, Cheetham RK, Stephens PJ, McBride DJ, Humphray SJ, Greenman CD, et al. A comprehensive catalogue of somatic mutations from a human cancer genome. Nature. 2010;463(7278):191–6.
Jackson BL, Grabowska A, Ratan HL. MicroRNA in prostate cancer: functional importance and potential as circulating biomarkers. BMC Cancer. 2014;14:930.
Lai CH, Huang SF, Liao CT, Chen IH, Wang HM, Hsieh LL. Clinical significance in oral cavity squamous cell carcinoma of pathogenic somatic mitochondrial mutations. PLoS One. 2013;8(6):e65578.
Lindberg J, Mills IG, Klevebring D, Liu W, Neiman M, Xu J, et al. The mitochondrial and autosomal mutation landscapes of prostate cancer. Eur Urol. 2013;63(4):702–8.
Kloss-Brandstätter A, Schäfer G, Erhart G, Hüttenhofer A, Coassin S, Seifarth C, et al. Somatic mutations throughout the entire mitochondrial genome are associated with elevated PSA levels in prostate cancer patients. Am J Hum Genet. 2010;87(6):802–12.
van Gisbergen MW, Voets AM, Starmans MH, de Coo IF, Yadak R, Hoffmann RF, et al. How do changes in the mtDNA and mitochondrial dysfunction influence cancer and cancer therapy? Challenges, opportunities and models. Mutat Res Rev Mutat Res. 2015;764:16–30.
INC MLS. The Prostate Core Mitomic Test™. 2016 [cited 2016 15 March]; Available from: http://mdnalifesciences.com/prostate-core-mitomic-test/.
Shull AY, Clendenning ML, Ghoshal-Gupta S, Farrell CL, Vangapandu HV, Dudas L, et al. Somatic mutations, allele loss, and DNA methylation of the Cub and Sushi Multiple Domains 1 (CSMD1) gene reveals association with early age of diagnosis in colorectal cancer patients. PLoS One. 2013;8(3):e58731.
Chaiyapan W, Duangpakdee P, Boonpipattanapong T, Kanngern S, Sangkhathat S. Somatic mutations of K-ras and BRAF in Thai colorectal cancer and their prognostic value. Asian Pac J Cancer Prev. 2013;14(1):329–32.
Wood LD, Parsons DW, Jones S, Lin J, Sjöblom T, Leary RJ, et al. The genomic landscapes of human breast and colorectal cancers. Science. 2007;318(5853):1108–13.
Barbieri CE, Tomlins SA. The prostate cancer genome: perspectives and potential. Urol Oncol. 2014;32(1):53.e15–22.
Rubin MA, Girelli G, Demichelis F. Genomic correlates to the newly proposed grading prognostic groups for prostate cancer. Eur Urol. 2016;69(4):557–60.
Miyagi Y, Sasaki T, Fujinami K, Sano J, Senga Y, Miura T, et al. ETS family-associated gene fusions in Japanese prostate cancer: analysis of 194 radical prostatectomy samples. Mod Pathol. 2010;23(11):1492–8.
Tapia-Laliena MA, Korzeniewski N, Hohenfellner M, Duensing S. High-risk prostate cancer: a disease of genomic instability. Urol Oncol. 2014;32(8):1101–7.
Robbins CM, Tembe WA, Baker A, Sinari S, Moses TY, Beckstrom-Sternberg S, et al. Copy number and targeted mutational analysis reveals novel somatic events in metastatic prostate tumors. Genome Res. 2011;21(1):47–55.
Mehra R, Kumar-Sinha C, Shankar S, Lonigro RJ, Jing X, Philips NE, et al. Characterization of bone metastases from rapid autopsies of prostate cancer patients. Clin Cancer Res. 2011;17(12):3924–32.
Mitchell T, Neal DE. The genomic evolution of human prostate cancer. Br J Cancer. 2015;113(2):193–8.
Gundem G, Van Loo P, Kremeyer B, Alexandrov LB, Tubio JM, Papaemmanuil E, et al. The evolutionary history of lethal metastatic prostate cancer. Nature. 2015;520(7547):353–7.
Cooper CS, Eeles R, Wedge DC, Van Loo P, Gundem G, Alexandrov LB, et al. Analysis of the genetic phylogeny of multifocal prostate cancer identifies multiple independent clonal expansions in neoplastic and morphologically normal prostate tissue. Nat Genet. 2015;47(4):367–72.
Network CGAR. The molecular taxonomy of primary prostate cancer. Cell. 2015;163(4):1011–25.
Petitjean A, Achatz MI, Borresen-Dale AL, Hainaut P, Olivier M. TP53 mutations in human cancers: functional selection and impact on cancer prognosis and outcomes. Oncogene. 2007;26(15):2157–65.
Richardson RB. p53 mutations associated with aging-related rise in cancer incidence rates. Cell Cycle. 2013;12(15):2468–78.
Olivier M, Hollstein M, Hainaut P. TP53 mutations in human cancers: origins, consequences, and clinical use. Cold Spring Harb Perspect Biol. 2010;2(1):a001008.
Forbes SA, Beare D, Gunasekaran P, Leung K, Bindal N, Boutselakis H, et al. COSMIC: exploring the world’s knowledge of somatic mutations in human cancer. Nucleic Acids Res. 2015;43(Database issue):D805–11.
Morris EV, Cerundolo L, Lu M, Verrill C, Fritzsche F, White MJ, et al. Nuclear iASPP may facilitate prostate cancer progression. Cell Death Dis. 2014;5:e1492.
Robles AI, Harris CC. Clinical outcomes and correlates of TP53 mutations and cancer. Cold Spring Harb Perspect Biol. 2010;2(3):a001016.
Robinson D, Van Allen EM, Wu YM, Schultz N, Lonigro RJ, Mosquera JM, et al. Integrative clinical genomics of advanced prostate cancer. Cell. 2015;161(5):1215–28.
Shi XB, Xue L, Ma AH, Tepper CG, Gandour-Edwards R, Kung HJ, et al. Tumor suppressive miR-124 targets androgen receptor and inhibits proliferation of prostate cancer cells. Oncogene. 2013;32(35):4130–8.
Huang L, Carney J, Cardona DM, Counter CM. Decreased tumorigenesis in mice with a Kras point mutation at C118. Nat Commun. 2014;5:5410.
Nodin B, Zendehrokh N, Sundström M, Jirström K. Clinicopathological correlates and prognostic significance of KRAS mutation status in a pooled prospective cohort of epithelial ovarian cancer. Diagn Pathol. 2013;8:106.
Yuan F, Shi M, Ji J, Shi H, Zhou C, Yu Y, et al. KRAS and DAXX/ATRX gene mutations are correlated with the clinicopathological features, advanced diseases, and poor prognosis in Chinese patients with pancreatic neuroendocrine tumors. Int J Biol Sci. 2014;10(9):957–65.
Wang XS, Shankar S, Dhanasekaran SM, Ateeq B, Sasaki AT, Jing X, et al. Characterization of KRAS rearrangements in metastatic prostate cancer. Cancer Discov. 2011;1(1):35–43.
Fu M, Zhang W, Shan L, Song J, Shang D, Ying J, et al. Mutation status of somatic EGFR and KRAS genes in Chinese patients with prostate cancer (PCa). Virchows Arch. 2014;464(5):575–81.
Reis ST, Timoszczuk LS, Pontes-Junior J, Viana N, Silva IA, Dip N, et al. The role of micro RNAs let7c, 100 and 218 expression and their target RAS, C-MYC, BUB1, RB, SMARCA5, LAMB3 and Ki-67 in prostate cancer. Clinics (Sao Paulo). 2013;68(5):652–7.
Cai H, Memarzadeh S, Stoyanova T, Beharry Z, Kraft AS, Witte ON. Collaboration of Kras and androgen receptor signaling stimulates EZH2 expression and tumor-propagating cells in prostate cancer. Cancer Res. 2012;72(18):4672–81.
Aytes A, Mitrofanova A, Kinkade CW, Lefebvre C, Lei M, Phelan V, et al. ETV4 promotes metastasis in response to activation of PI3-kinase and Ras signaling in a mouse model of advanced prostate cancer. Proc Natl Acad Sci. 2013;110(37):E3506–15.
Blair BG, Wu X, Zahari MS, Mohseni M, Cidado J, Wong HY, et al. A phosphoproteomic screen demonstrates differential dependence on HER3 for MAP kinase pathway activation by distinct PIK3CA mutations. Proteomics. 2015;15(2–3):318–26.
Karakas B, Bachman KE, Park BH. Mutation of the PIK3CA oncogene in human cancers. Br J Cancer. 2006;94(4):455–9.
Gabelli SB, Echeverria I, Alexander M, Duong-Ly KC, Chaves-Moreira D, Brower ET, et al. Activation of PI3Kα by physiological effectors and by oncogenic mutations: structural and dynamic effects. Biophys Rev. 2014;6(1):89–95.
Hou W, Liu J, Chen P, Wang H, Ye BC, Qiang F. Mutation analysis of key genes in RAS/RAF and PI3K/PTEN pathways in Chinese patients with hepatocellular carcinoma. Oncol Lett. 2014;8(3):1249–54.
Stemke-Hale K, Gonzalez-Angulo AM, Lluch A, Neve RM, Kuo WL, Davies M, et al. An integrative genomic and proteomic analysis of PIK3CA, PTEN, and AKT mutations in breast cancer. Cancer Res. 2008;68(15):6084–91.
Qiu W, Schönleben F, Li X, Ho DJ, Close LG, Manolidis S, et al. PIK3CA mutations in head and neck squamous cell carcinoma. Clin Cancer Res. 2006;12(5):1441–6.
Mullerad M, Hricak H, Kuroiwa K, Pucar D, Chen HN, Kattan MW, et al. Comparison of endorectal magnetic resonance imaging, guided prostate biopsy and digital rectal examination in the preoperative anatomical localization of prostate cancer. J Urol. 2005;174(6):2158–63.
Samuels Y, Wang Z, Bardelli A, Silliman N, Ptak J, Szabo S, et al. High frequency of mutations of the PIK3CA gene in human cancers. Science. 2004;304(5670):554.
Schönleben F, Qiu W, Ciau NT, Ho DJ, Li X, Allendorf JD, et al. PIK3CA mutations in intraductal papillary mucinous neoplasm/carcinoma of the pancreas. Clin Cancer Res. 2006;12(12):3851–5.
Tang KD, Ling MT. Targeting drug-resistant prostate cancer with dual PI3K/mTOR inhibition. Curr Med Chem. 2014;21(26):3048–56.
Wong HL, Peters U, Hayes RB, Huang WY, Schatzkin A, Bresalier RS, et al. Polymorphisms in the adenomatous polyposis coli (APC) gene and advanced colorectal adenoma risk. Eur J Cancer. 2010;46(13):2457–66.
Pećina-Slaus N, Majić Z, Musani V, Zeljko M, Cupić H. Report on mutation in exon 15 of the APC gene in a case of brain metastasis. J Neurooncol. 2010;97(1):143–8.
Ashktorab H, Daremipouran M, Devaney J, Varma S, Rahi H, Lee E, et al. Identification of novel mutations by exome sequencing in African American colorectal cancer patients. Cancer. 2015;121(1):34–42.
Christie M, Jorissen RN, Mouradov D, Sakthianandeswaren A, Li S, Day F, et al. Different APC genotypes in proximal and distal sporadic colorectal cancers suggest distinct WNT/β-catenin signalling thresholds for tumourigenesis. Oncogene. 2013;32(39):4675–82.
Drier Y, Lawrence MS, Carter SL, Stewart C, Gabriel SB, Lander ES, et al. Somatic rearrangements across cancer reveal classes of samples with distinct patterns of DNA breakage and rearrangement-induced hypermutability. Genome Res. 2013;23(2):228–35.
Gerecke C, Mascher C, Gottschalk U, Kleuser B, Scholtka B. Ultrasensitive detection of unknown colon cancer-initiating mutations using the example of the Adenomatous polyposis coli gene. Cancer Prev Res (Phila). 2013;6(9):898–907.
Rogler G. Chronic ulcerative colitis and colorectal cancer. Cancer Lett. 2014;345(2):235–41.
Fox SA, Richards AK, Kusumah I, Perumal V, Bolitho EM, Mutsaers SE, et al. Expression profile and function of Wnt signaling mechanisms in malignant mesothelioma cells. Biochem Biophys Res Commun. 2013;440(1):82–7.
Valkenburg KC, Yu X, De Marzo AM, Spiering TJ, Matusik RJ, Williams BO. Activation of Wnt/β-catenin signaling in a subpopulation of murine prostate luminal epithelial cells induces high grade prostate intraepithelial neoplasia. Prostate. 2014;74(15):1506–20.
Zhang W, Jiao H, Zhang X, Zhao R, Wang F, He W, et al. Correlation between the expression of DNMT1, and GSTP1 and APC, and the methylation status of GSTP1 and APC in association with their clinical significance in prostate cancer. Mol Med Rep. 2015;12(1):141–6.
Mulholland DJ, Kobayashi N, Ruscetti M, Zhi A, Tran LM, Huang J, et al. Pten loss and RAS/MAPK activation cooperate to promote EMT and metastasis initiated from prostate cancer stem/progenitor cells. Cancer Res. 2012;72(7):1878–89.
Tian L, Fang YX, Xue JL, Chen JZ. Four microRNAs promote prostate cell proliferation with regulation of PTEN and its downstream signals in vitro. PLoS One. 2013;8(9):e75885.
Jin G, Kim MJ, Jeon HS, Choi JE, Kim DS, Lee EB, et al. PTEN mutations and relationship to EGFR, ERBB2, KRAS, and TP53 mutations in non-small cell lung cancers. Lung Cancer. 2010;69(3):279–83.
Barnett CM, Heinrich MC, Lim J, Nelson D, Beadling C, Warrick A, et al. Genetic profiling to determine risk of relapse-free survival in high-risk localized prostate cancer. Clin Cancer Res. 2014;20(5):1306–12.
Wu Z, He B, He J, Mao X. Upregulation of miR-153 promotes cell proliferation via downregulation of the PTEN tumor suppressor gene in human prostate cancer. Prostate. 2013;73(6):596–604.
Maire CL, Ligon KL. Molecular pathologic diagnosis of epidermal growth factor receptor. Neuro Oncol. 2014;16(Suppl 8):viii1–6.
Cho J, Bass AJ, Lawrence MS, Cibulskis K, Cho A, Lee SN, et al. Colon cancer-derived oncogenic EGFR G724S mutant identified by whole genome sequence analysis is dependent on asymmetric dimerization and sensitive to cetuximab. Mol Cancer. 2014;13:141.
Hour TC, Chung SD, Kang WY, Lin YC, Chuang SJ, Huang AM, et al. EGFR mediates docetaxel resistance in human castration-resistant prostate cancer through the Akt-dependent expression of ABCB1 (MDR1). Arch Toxicol. 2015;89(4):591–605.
National Center for Biotechnology Information USNLoM. ckit gene. http://www.ncbi.nlm.nih.gov/gene?term=(ckit%5bgene.
Database GTHG. kit gene. http://www.genecards.org/cgi-bin/carddisp.pl?gene=KIT&search=kit.
Simak R, Capodieci P, Cohen DW, Fair WR, Scher H, Melamed J, et al. Expression of c-kit and kit-ligand in benign and malignant prostatic tissues. Histol Histopathol. 2000;15(2):365–74.
Dakhova O, Rowley D, Ittmann M. Genes upregulated in prostate cancer reactive stroma promote prostate cancer progression in vivo. Clin Cancer Res. 2014;20(1):100–9.
Won D, Chi HS, Shim H, Jang S, Park CJ, Lee JH. The prognostic impact of c-KIT mutation in systemic mastocytosis associated with acute myeloid leukaemia patients. Leuk Res. 2013;37(8):883–8.
Yu J, Mani RS, Cao Q, Brenner CJ, Cao X, Wang X, et al. An integrated network of androgen receptor, polycomb, and TMPRSS2-ERG gene fusions in prostate cancer progression. Cancer Cell. 2010;17(5):443–54.
Segawa N, Nakamura M, Shan L, Utsunomiya H, Nakamura Y, Mori I, et al. Expression and somatic mutation on androgen receptor gene in prostate cancer. Int J Urol. 2002;9(10):545–53.
Taylor BS, Schultz N, Hieronymus H, Gopalan A, Xiao Y, Carver BS, et al. Integrative genomic profiling of human prostate cancer. Cancer Cell. 2010;18(1):11–22.
Carver BS, Chapinski C, Wongvipat J, Hieronymus H, Chen Y, Chandarlapaty S, et al. Reciprocal feedback regulation of PI3K and androgen receptor signaling in PTEN-deficient prostate cancer. Cancer Cell. 2011;19(5):575–86.
Gottlieb B, Beitel LK, Nadarajah A, Paliouras M, Trifiro M. The androgen receptor gene mutations database: 2012 update. Hum Mutat. 2012;33(5):887–94.
Mononen N, Syrjäkoski K, Matikainen M, Tammela TL, Schleutker J, Kallioniemi OP, et al. Two percent of Finnish prostate cancer patients have a germ-line mutation in the hormone-binding domain of the androgen receptor gene. Cancer Res. 2000;60(22):6479–81.
Koochekpour S. Androgen receptor signaling and mutations in prostate cancer. Asian J Androl. 2010;12(5):639–57.
Li W, Cavasotto CN, Cardozo T, Ha S, Dang T, Taneja SS, et al. Androgen receptor mutations identified in prostate cancer and androgen insensitivity syndrome display aberrant ART-27 coactivator function. Mol Endocrinol. 2005;19(9):2273–82.
Zaman N, Giannopoulos PN, Chowdhury S, Bonneil E, Thibault P, Wang E, et al. Proteomic-coupled-network analysis of T877A-androgen receptor interactomes can predict clinical prostate cancer outcomes between White (non-Hispanic) and African-American groups. PLoS One. 2014;9(11):e113190.
Steinestel J, Luedeke M, Arndt A, Schnoeller TJ, Lennerz JK, Wurm C, et al. Detecting predictive androgen receptor modifications in circulating prostate cancer cells. Oncotarget. 2015. doi:10.18632/oncotarget.3925.
Jentzmik F, Azoitei A, Zengerling F, Damjanoski I, Cronauer MV. Androgen receptor aberrations in the era of abiraterone and enzalutamide. World J Urol. 2016;34(3):297–303.
Korpal M, Korn JM, Gao X, Rakiec DP, Ruddy DA, Doshi S, et al. An F876L mutation in androgen receptor confers genetic and phenotypic resistance to MDV3100 (enzalutamide). Cancer Discov. 2013;3(9):1030–43.
Antonarakis ES, Lu C, Wang H, Luber B, Nakazawa M, Roeser JC, et al. AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. N Engl J Med. 2014;371(11):1028–38.
An J, Wang C, Deng Y, Yu L, Huang H. Destruction of full-length androgen receptor by wild-type SPOP, but not prostate-cancer-associated mutants. Cell Rep. 2014;6(4):657–69.
Hu R, Dunn TA, Wei S, Isharwal S, Veltri RW, Humphreys E, et al. Ligand-independent androgen receptor variants derived from splicing of cryptic exons signify hormone-refractory prostate cancer. Cancer Res. 2009;69(1):16–22.
Steinkamp MP, O’Mahony OA, Brogley M, Rehman H, Lapensee EW, Dhanasekaran S, et al. Treatment-dependent androgen receptor mutations in prostate cancer exploit multiple mechanisms to evade therapy. Cancer Res. 2009;69(10):4434–42.
Ceder Y. Non-coding RNAs in prostate cancer: from discovery to clinical applications. Adv Exp Med Biol. 2016;886:155–70.
Nadiminty N, Tummala R, Lou W, Zhu Y, Zhang J, Chen X, et al. MicroRNA let-7c suppresses androgen receptor expression and activity via regulation of Myc expression in prostate cancer cells. J Biol Chem. 2012;287(2):1527–37.
Ribas J, Ni X, Haffner M, Wentzel EA, Salmasi AH, Chowdhury WH, et al. miR-21: an androgen receptor-regulated microRNA that promotes hormone-dependent and hormone-independent prostate cancer growth. Cancer Res. 2009;69(18):7165–9.
Li C, Ao J, Fu J, Lee DF, Xu J, Lonard D, et al. Tumor-suppressor role for the SPOP ubiquitin ligase in signal-dependent proteolysis of the oncogenic co-activator SRC-3/AIB1. Oncogene. 2011;30(42):4350–64.
Barbieri CE, Baca SC, Lawrence MS, Demichelis F, Blattner M, Theurillat JP, et al. Exome sequencing identifies recurrent SPOP, FOXA1 and MED12 mutations in prostate cancer. Nat Genet. 2012;44(6):685–9.
Kim MS, Je EM, Oh JE, Yoo NJ, Lee SH. Mutational and expressional analyses of SPOP, a candidate tumor suppressor gene, in prostate, gastric and colorectal cancers. APMIS. 2013;121(7):626–33.
Zuhlke KA, Johnson AM, Tomlins SA, Palanisamy N, Carpten JD, Lange EM, et al. Identification of a novel germline SPOP mutation in a family with hereditary prostate cancer. Prostate. 2014;74(9):983–90.
Theurillat JP, Udeshi ND, Errington WJ, Svinkina T, Baca SC, Pop M, et al. Prostate cancer. Ubiquitylome analysis identifies dysregulation of effector substrates in SPOP-mutant prostate cancer. Science. 2014;346(6205):85–9.
Boysen G, Barbieri CE, Prandi D, Blattner M, Chae SS, Dahija A, et al. SPOP mutation leads to genomic instability in prostate cancer. Elife. 2015;4.
An J, Ren S, Murphy SJ, Dalangood S, Chang C, Pang X, et al. Truncated ERG oncoproteins from TMPRSS2-ERG fusions are resistant to SPOP-mediated proteasome degradation. Mol Cell. 2015;59(6):904–16.
Adamo P, Ladomery MR. The oncogene ERG: a key factor in prostate cancer. Oncogene. 2016;35(4):403–14.
Robinson JL, Holmes KA, Carroll JS. FOXA1 mutations in hormone-dependent cancers. Front Oncol. 2013;3:20.
Barbieri CE, Bangma CH, Bjartell A, Catto JW, Culig Z, Grönberg H, et al. The mutational landscape of prostate cancer. Eur Urol. 2013;64(4):567–76.
St John J, Powell K, Conley-Lacomb MK, Chinni SR. TMPRSS2-ERG fusion gene expression in prostate tumor cells and its clinical and biological significance in prostate cancer progression. J Cancer Sci Ther. 2012;4(4):94–101.
García-Flores M, Casanova-Salas I, Rubio-Briones J, Calatrava A, Domínguez-Escrig J, Rubio L, et al. Clinico-pathological significance of the molecular alterations of the SPOP gene in prostate cancer. Eur J Cancer. 2014;50(17):2994–3002.
Ueda M, Iguchi T, Masuda T, Nakahara Y, Hirata H, Uchi R, et al. Somatic mutations in plasma cell-free DNA are diagnostic markers for esophageal squamous cell carcinoma recurrence. Oncotarget. 2016. doi:10.18632/oncotarget.11409.
Mooney SM, Jolly MK, Levine H, Kulkarni P. Phenotypic plasticity in prostate cancer: role of intrinsically disordered proteins. Asian J Androl. 2016;18(5):704–10.
Beltran H, Antonarakis ES, Morris MJ, Attard G. emerging molecular biomarkers in advanced prostate cancer: translation to the clinic. Am Soc Clin Oncol Educ Book. 2016;35:131–41.
Wang S, Song Y, Yan F, Liu D. Mechanisms of resistance to third-generation EGFR tyrosine kinase inhibitors. Front Med. 2016. doi:10.1007/s11684-016-0488-1.
Pashayan N, Duffy SW, Chowdhury S, Dent T, Burton H, Neal DE, et al. Polygenic susceptibility to prostate and breast cancer: implications for personalised screening. Br J Cancer. 2011;104(10):1656–63.
Cybulski C, Wokołorczyk D, Kluźniak W, Kashyap A, Gołąb A, Słojewski M, et al. A personalised approach to prostate cancer screening based on genotyping of risk founder alleles. Br J Cancer. 2013;108(12):2601–9.
Jackson SE, Chester JD. Personalised cancer medicine. Int J Cancer. 2015;137(2):262–6.
Decker B, Karyadi DM, Davis BW, Karlins E, Tillmans LS, Stanford JL, et al. Biallelic BRCA2 mutations shape the somatic mutational landscape of aggressive prostate tumors. Am J Hum Genet. 2016;98(5):818–29.
Wojnarowicz PM, Oros KK, Quinn MC, Arcand SL, Gambaro K, Madore J, et al. The genomic landscape of TP53 and p53 annotated high grade ovarian serous carcinomas from a defined founder population associated with patient outcome. PLoS One. 2012;7(9):e45484.
Shanmugam V, Ramanathan RK, Lavender NA, Sinari S, Chadha M, Liang WS, et al. Whole genome sequencing reveals potential targets for therapy in patients with refractory KRAS mutated metastatic colorectal cancer. BMC Med Genomics. 2014;7:36.
Nakayama S, Sng N, Carretero J, Welner R, Hayashi Y, Yamamoto M, et al. β-catenin contributes to lung tumor development induced by EGFR mutations. Cancer Res. 2014;74(20):5891–902.
Sun Z, Wang L, Eckloff BW, Deng B, Wang Y, Wampfler JA, et al. Conserved recurrent gene mutations correlate with pathway deregulation and clinical outcomes of lung adenocarcinoma in never-smokers. BMC Med Genomics. 2014;7:32.
Muñoz-Moreno L, Arenas MI, Carmena MJ, Schally AV, Prieto JC, Bajo AM. Growth hormone-releasing hormone antagonists abolish the transactivation of human epidermal growth factor receptors in advanced prostate cancer models. Invest New Drugs. 2014;32(5):871–82.
Rapisuwon S, Parks K, Al-Refaie W, Atkins MB. Novel somatic KIT exon 8 mutation with dramatic response to imatinib in a patient with mucosal melanoma: a case report. Melanoma Res. 2014;24(5):509–11.
Gavert N, Shvab A, Sheffer M, Ben-Shmuel A, Haase G, Bakos E, et al. c-Kit is suppressed in human colon cancer tissue and contributes to L1-mediated metastasis. Cancer Res. 2013;73(18):5754–63.
Han X, Zhao J, Ji Y, Xu X, Lou W. Expression of CK19 and KIT in resectable pancreatic neuroendocrine tumors. Tumour Biol. 2013;34(5):2881–9.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
M. J. Alvarez-Cubero; L. J. Martinez-Gonzalez; I. Robles-Fernandez; J. Martinez-Herrera MS; G. Garcia-Rodriguez; M. Pascual-Geler MD; J. M. Cozar and J. A. Lorente, no have nothing to disclose. The authors have no conflict of interest.
Rights and permissions
About this article

Cite this article
Alvarez-Cubero, M.J., Martinez-Gonzalez, L.J., Robles-Fernandez, I. et al. Somatic Mutations in Prostate Cancer: Closer to Personalized Medicine. Mol Diagn Ther 21, 167–178 (2017). https://doi.org/10.1007/s40291-016-0248-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40291-016-0248-6