
Abstract
Hfq is a global regulatory RNA-binding protein abundant in many bacteria. In terms of virulence regulation in Salmonella, Hfq binds to and stabilizes the mRNA of hilD, a master regulator of Salmonella pathogenicity island 1 (SPI-1), and Hfq activates SPI-1 gene transcription during the late exponential-growth phase. In contrast to the positive roles of Hfq in SPI-1 regulation, we observed that the absence of Hfq increased the expression of SPI-1 regulators, including HilA and InvF, during the late stationary phase. This result suggests that Hfq has pleiotropic functions, controlling SPI-1 positively or negatively depending on the growth conditions.
Similar content being viewed by others
Introduction
Salmonellae are enteric pathogens that cause gastroenteritis and enteric fever. They diverged from the genus Escherichia ~120–160 million years ago with the acquisition of a set of virulence genes, termed Salmonella pathogenicity island 1 (SPI-1) (Galan 1999). SPI-1, a 40-kb island located at centisome 63 of the genome, is composed of more than 39 genes encoding regulators, effectors, cognate chaperones, and components of a type III secretion system (SPI-1 T3SS) (Lostroh and Lee 2001). The SPI-1 T3SS plays diverse roles in multiple steps of Salmonella systemic infections, including invasion into host epithelial cells (Galan 1999), induction of proinflammatory responses (Hardt et al. 1998), stimulation of programmed cell death in macrophages (Hernandez et al. 2003), and proliferation within host cells (Que et al. 2013). Due to the multifaceted roles of SPI-1 in different locations and at different times, its expression is finely regulated by a coordinated regulatory network that integrates multiple environmental signals conclusively under the control of HilA, a direct activator of SPI-1 (Golubeva et al. 2012). HilD, another cognate activator encoded within SPI-1, is positioned at the hub of the regulatory network, linking multiple regulatory inputs to HilA; it is therefore regarded as a master regulator at the top of the SPI-1 regulatory circuit (Golubeva et al. 2012).
Hfq is one of the most abundant RNA-binding proteins in bacteria and has key roles in gene expression control (Vogel and Luisi 2011). In terms of SPI-1 regulation, Hfq was found to stabilize hilD mRNA and enhance HilD activity in late log/early stationary phase cultures at 4 h post-inoculation (p.i.) (Sittka et al. 2008; Lim et al. 2013). Accordingly, hilA promoter activity was reduced in Δhfq strains (Sittka et al. 2007; Lim et al. 2013). However, we suggest that the influence of Hfq on SPI-1 varies depending on bacterial growth phases, based on the observation in this study that the expression of SPI-1 regulators, including HilA and InvF, in Δhfq appeared to catch up with that in wild-type Salmonella in late stationary phase cultures.
Materials and methods
Bacterial strains and growth conditions
Wild-type Salmonella enterica serovar Typhimurium SL1344 was used as the parent strain in all of the strains constructed in this study. Wild-type Salmonella and its isogenic Δhfq strain harboring the hilA::lacZY fusion, respectively, were constructed in a previous study (Lim et al. 2013). Salmonella strains expressing hemagglutinin (HA)-tagged HilA or HA-tagged InvF were constructed using the phage λ Red recombination system as described elsewhere (Datsenko and Wanner 2000; Lim et al. 2013). The primers used to amplify the kanamycin cassette from pKD13 (Datsenko and Wanner 2000) were hilA-HAF (5′-ttc aaa aga tgg aaa cag gat ccc cgc ttg att aaa tta cgg tat ccg tat gat gtt cct gat tat gct agc ctc tga tgt agg ctg gag ctg ctt cg-3′) and hilA-HAR (5′-att acg atg ata aaa aaa taa tgc ata tct cct ctc tca gat tat tcc ggg gat ccg tcg acc-3′) for HilA::HA expression, and invF-HAF (5′-tcg ccg cgg aaa tta tca aat att att caa ttg gca gac aaa tat ccg tat gat gtt cct gat tat gct agc ctc tga tgt agg ctg gag ctg ctt cg-3′) and invF-HAR (5′-cgc ggc aca tgc cag cac tct ggc caa aag aat atg tgt cta ttc cgg gga tcc gtc gac c-3′) for InvF::HA expression. Transfer of the resistance allele between strains was conducted by P22HT-mediated transduction (Maloy et al. 1996).
Bacteria grown in Luria–Bertani (LB) broth were added to fresh LB broth at a 1:100 ratio and cultivated at 37 °C with shaking. Kanamycin was added when required at 50 µg/ml.
β-galactosidase assay
Wild-type Salmonella and an isogenic Δhfq strain harboring the chromosomal hilA::lacZY fusion were cultivated in LB broth and the activity of hilA::lacZY transcriptional fusions was analyzed by determining the β-galactosidase activity according to a standard method (Maloy et al. 1996).
Western blot analysis
Salmonella strains producing HA-tagged proteins were harvested by centrifugation at 4 and 12 h after inoculation. The cell lysates were prepared by lysis of cells using B-PER solution (Pierce, Rockford, IL) and lysates corresponding to 20 μg of protein were separated by 12 % SDS-PAGE. The proteins on the gel were transferred to nitrocellulose membranes and the membranes were incubated with anti-HA (Sigma, St. Louis, MO, USA) or anti-DnaK (Stressgen Biotechnologies Corp., Victoria, BC, Canada) antibodies and anti-mouse IgG horseradish peroxidase-linked antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA) in succession. The blots were developed using the WEST-ZOL detection system (Intron Biotechnology, Seongnam, Korea).
Results and discussion
In contrast to the known positive role of Hfq in SPI-1 regulation, we have observed that the hilA expression seemed to be increased by the absence of Hfq at 12 h p.i. in our previous study (Lim et al. 2013). To gain better insight into this issue, we first monitored hilA expression in a Δhfq strain using hilA::lacZY chromosomal fusion across various growth phases (Fig. 1a). Wild-type Salmonella enterica serovar Typhimurium SL1344 and an isogenic Δhfq strain harboring the hilA::lacZY fusion were cultivated in Luria–Bertani (LB) broth, and the activity of chromosomal hilA::lacZY transcriptional fusions was analyzed by determining the β-galactosidase activity every 2 h. As expected, hilA expression was reduced by approximately two-fold at 4 h p.i. in the Δhfq strain when compared with wild type. However, a time-course observation demonstrated comparable hilA expression levels between the two strains at 6 h p.i. and higher hilA expression in the Δhfq strain at later time points (the reduced expression of hilA after 6 h in wild type, while the increased expression of hilA in the Δhfq strain was maintained for the remainder of the experiment).

Growth phase-dependent pleiotropic effects of Hfq on hilA expression. a Wild-type Salmonella Typhimurium SL1344 and isogenic Δhfq strains harboring hilA::lacZY fusion were cultivated in LB broth at 37 °C, and the activity of hilA::lacZY was determined using a β-galactosidase assay every 2 h and represents the average result from three independent biological cultures. b Wild-type and Δhfq Salmonella strains producing HA-tagged HilA were grown in LB broth for 4 and 12 h and the levels of HilA::HA were compared by Western blot analysis using anti-HA antibodies. The amount of protein loaded in each lane was normalized to the level of intracellular DnaK
The time-dependent differential effects of Hfq on hilA expression were further verified by comparing HilA protein levels between the wild-type and Δhfq strains (Fig. 1b). Salmonella strains expressing HA-tagged HilA were cultivated in LB medium and subjected to Western blot analysis as described in Materials and Methods. In accordance with the β-galactosidase assay results, the absence of Hfq decreased HilA levels during the late log phase (4 h p.i.), whereas it increased HilA levels during the late stationary phase (12 h p.i.).
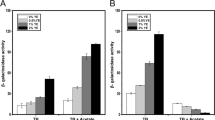
HilA can activate the expression of inv/spa and prg/org operons, which encode components of the SPI1-T3SS machinery, by binding to the invF and prgH promoters (Lostroh and Lee 2001). InvF, which is transcriptionally activated by HilA and HilD, was also differentially regulated by Hfq between the late log and late stationary phases; decreased InvF expression at 4 h p.i. and increased InvF expression at 12 h p.i. were observed in the Δhfq strain compared with wild type (Fig. 2). Taken together, these results suggest that Hfq activates the transcription of SPI-1 genes by stabilizing hilD mRNA during the late log phase but decreases their expression during the late stationary phase; however, the role of Hfq in the negative regulation at the late stationary phase remains undetermined.

Differential regulation of InvF by Hfq between the late log and late stationary phases of growth. The expression level of InvF::HA was compared between wild-type and Δhfq Salmonella strains at 4 and 12 h post-inoculation using Western blot analysis. Equal amounts of cell lysate (20 μg protein) were loaded in each lane, and the level of InvF protein produced in each strain was normalized to the intracellular DnaK level
Hfq binds 727 Salmonella mRNAs to control their translational efficiency in combination with small noncoding RNAs (sRNAs) (Sittka et al. 2008). To gain further insights into the role of Hfq in the late stationary phase-dependent negative regulation of SPI-1 genes, we searched for regulators in the SPI-1 regulatory circuit, the expression of which might be affected by Hfq under late stationary phase conditions. CsrA, a global regulatory RNA-binding protein, binds to a region within the hilD mRNA transcript that overlaps with the Shine-Dalgarno sequence and prevents its translation, leading to accelerated turnover (Martinez et al. 2011). CsrA transcription is in turn activated by RpoS, an alternative sigma factor, responding to stressful conditions such as the starvation-induced stationary phase of growth (Mukherjee et al. 1998; Dong and Schellhorn 2010; Knudsen et al. 2013). RpoS mRNA is a representative target of Hfq, whose translation is released by the coordinated binding of Hfq and sRNAs (Lease and Woodson 2004). The translation of rpoS mRNA is self-repressed in the absence of Hfq by a stem loop within its 5′ leader; thus, Salmonella Δhfq strains suffer from the lack of RpoS under starvation-induced stationary phase growth (Brown and Elliott 1996). The depletion of RpoS caused by the absence of Hfq downregulates CsrA expression, which may relieve the repression of SPI-1 genes under late stationary phase conditions. Knudsen et al. (2013) demonstrated that the downregulation of SPI-1 genes in clpP mutant strains was abolished by the deletion of rpoS, which most likely nullified the increase in CsrA caused by the deletion of clpP.
To investigate the possible indirect regulation of SPI-1 genes by Hfq via RpoS and CsrA, we compared the transcriptional profiles of SPI-1 genes in Δhfq, ΔrpoS, and ΔcsrA strains under log and stationary phase conditions (Fig. 3). Transcriptomic data for the Δhfq, ΔrpoS, and ΔcsrA strains were retrieved from a previous study (Yoon et al. 2009) and reanalyzed. In accordance with the results presented in this study, the effects of Hfq on the expression of SPI-1 genes differed between the log and stationary phases. The transcription of SPI-1 genes in the absence of Hfq was decreased in log phase LB cultures, whereas it tended to be elevated in stationary phase LB cultures. Interestingly, the SPI-1 expression patterns in the ΔrpoS and ΔcsrA strains paralleled each other in the two growth phases, and SPI-1 gene transcription appeared increased in both strains during the stationary phase. Furthermore, the negative effects of RpoS and CsrA on SPI-1 expression were more distinct during the stationary phase, when the expression of rpoS is elevated in response to nutrient depletion. Given that the transcriptional responses of the SPI-1 region were similar among the Δhfq, ΔrpoS, and ΔcsrA strains during the stationary phase, we intend to explore the roles of RpoS and CsrA in the negative regulation of SPI-1 by Hfq during late stationary phase growth in a future study.

Transcriptomic analysis of SPI-1 genes in ΔcsrA, ΔrpoS, and Δhfq strains. The expression ratio, as determined by the microarray results, of a given SPI-1 gene in a mutant strain relative to the wild-type strain was computed in the LB log and stationary phases. Green and red represent a logarithmic (log2) decrease and increase in the mutant, respectively. Rows indicate individual genes within SPI-1, while columns correspond to regulatory mutants under two growth conditions
References
Brown L, Elliott T (1996) Efficient translation of the RpoS sigma factor in Salmonella typhimurium requires host factor I, an RNA-binding protein encoded by the hfq gene. J Bacteriol 178:3763–3770
Datsenko KA, Wanner BL (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA 97:6640–6645
Dong T, Schellhorn HE (2010) Role of RpoS in virulence of pathogens. Infect Immun 78:887–897
Galan JE (1999) Interaction of Salmonella with host cells through the centisome 63 type III secretion system. Curr Opin Microbiol 2:46–50
Golubeva YA, Sadik AY, Ellermeier JR, Slauch JM (2012) Integrating global regulatory input into the Salmonella pathogenicity island 1 type III secretion system. Genetics 190:79–90
Hardt WD, Chen LM, Schuebel KE, Bustelo XR, Galan JE (1998) S. typhimurium encodes an activator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells. Cell 93:815–826
Hernandez LD, Pypaert M, Flavell RA, Galan JE (2003) A Salmonella protein causes macrophage cell death by inducing autophagy. J Cell Biol 163:1123–1131
Knudsen GM, Olsen JE, Aabo S, Barrow P, Rychlik I, Thomsen LE (2013) ClpP deletion causes attenuation of Salmonella typhimurium virulence through mis-regulation of RpoS and indirect control of CsrA and the SPI genes. Microbiology 159:1497–1509
Lease RA, Woodson SA (2004) Cycling of the Sm-like protein Hfq on the DsrA small regulatory RNA. J Mol Biol 344:1211–1223
Lim S, Yoon H, Kim M, Han A, Choi J, Choi J, Ryu S (2013) Hfq and ArcA are involved in the stationary phase-dependent activation of Salmonella pathogenicity island 1 (SPI1) under shaking culture conditions. J Microbiol Biotechnol 23:1664–1672
Lostroh CP, Lee CA (2001) The Salmonella pathogenicity island-1 type III secretion system. Microbes Infect 3:1281–1291
Maloy SR, Stewart VJ, Taylor RK (1996) In Genetic analysis of pathogenic bacteria : a laboratory manual, vol pp. Cold Spring Harbor Laboratory Press, Plainview, NY
Martinez LC, Yakhnin H, Camacho MI, Georgellis D, Babitzke P, Puente JL, Bustamante VH (2011) Integration of a complex regulatory cascade involving the SirA/BarA and Csr global regulatory systems that controls expression of the Salmonella SPI-1 and SPI-2 virulence regulons through HilD. Mol Microbiol 80:1637–1656
Mukherjee A, Cui Y, Ma W, Liu Y, Ishihama A, Eisenstark A, Chatterjee AK (1998) RpoS (sigma-S) controls expression of rsmA, a global regulator of secondary metabolites, harpin, and extracellular proteins in Erwinia carotovora. J Bacteriol 180:3629–3634
Que F, Wu S, Huang R (2013) Salmonella pathogenicity island 1(SPI-1) at work. Curr Microbiol 66:582–587
Sittka A, Pfeiffer V, Tedin K, Vogel J (2007) The RNA chaperone Hfq is essential for the virulence of Salmonella typhimurium. Mol Microbiol 63:193–217
Sittka A, Lucchini S, Papenfort K, Sharma CM, Rolle K, Binnewies TT, Hinton JC, Vogel J (2008) Deep sequencing analysis of small noncoding RNA and mRNA targets of the global post-transcriptional regulator, Hfq. PLoS Genet 4:e1000163
Vogel J, Luisi BF (2011) Hfq and its constellation of RNA. Nat Rev Microbiol 9:578–589
Yoon H, McDermott JE, Porwollik S, McClelland M, Heffron F (2009) Coordinated regulation of virulence during systemic infection of Salmonella enterica serovar Typhimurium. PLoS Pathog 5:e1000306
Acknowledgments
This research was supported by the Nuclear R&D program of the Ministry of Science, ICT and Future Planning, Republic of Korea.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Lim, S., Choi, J., Kim, M. et al. Temporal regulation of Salmonella pathogenicity Island 1 (SPI-1) hilA by Hfq in Salmonella enterica serovar typhimurium. J Korean Soc Appl Biol Chem 58, 169–172 (2015). https://doi.org/10.1007/s13765-015-0034-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13765-015-0034-x