
Abstract
Climate change alters ecohydrological processes on different temporal and spatial scales. The aim of this study was to estimate ecological instream flow requirements (EIFR) under climate change impacts and to ascertain optimum flow magnitude to maintain the health of river ecosystems. We used the Mann–Kendall test, the River2D Hydrodynamic model, and the frequency-weighted usable area (FWUA) model on an annual scale in the Juma River to investigate changes in EIFRs caused by climate change. Findings indicated that: (1) between 1956 and 2005, annual instream flow in the Juma River exhibited an abrupt downward trend (in 1981); (2) variation in instream flow governed the area of available fish habitat, and degrees of change for low flow threshold values and low and high FWUA threshold almost exclusively occurred in descending sequential order from winter, spring, autumn, and summer; (3) changes in hydrological regimes influenced both the quantity and quality of physical habitat area, contributing greater to quality than to the distribution of area of physical habitat for Pseudorasbora parva. Climate change, reflected in the changes in EIFR and the area of optimum physical habitat, has dramatically influenced ecological structure and function, even in the small river system investigated for this study. Findings indicated that more rational measures should be put into practice to help address climate change.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Along with other important variables, climate change alters the frequency and intensity of precipitation. This has led to changes in streamflow response, resulting in the degradation of river ecosystems (Poff 2002; IPCC 2007; Liu and Cui 2011; Barranco et al. 2013). Ecological instream flow requirements (EIFRs) (or environmental flow) have been proposed as a means to protect river ecosystems. EIFR is defined as “the quantity of water required to maintain a river ecosystem in its desired state [that is] of particular importance in areas of high natural value” (Acreman and Dunbar 2004; Zhong et al. 2006; Piniewski et al. 2014). EIFRs are determinants of channel morphology, riparian and aquatic flora and fauna, water quality, and stream load transport (Song et al. 2007). Methods used to calculate EIFRs can generally be classified into four types: hydrological, hydraulic ratings, habitat simulations, and holistic methods as well as hybrids or integrations of these methods (Tharme 2003; Huckstorf et al. 2008; Cui et al. 2010; Karimi et al. 2012; Shokoohi and Amini 2014). Methods associated with habitat simulations take into account biological factors in combining hydrological indexes and ecological processes to provide more scientifically based and practical results (Daniel and Nassir 1994; Cui et al. 2010; Tian et al. 2012). These include instream flow incremental methodology (IFIM) (Bovee 1982), the physical habitat simulation model (PHABSIM) (Milhous et al. 1989), the Computer Aided Simulation Model for Instream Flow and Riparia (CASiMiR) (Zhao et al. 2008), and the River2D Hydrodynamic model (the fish habitat component of River2D is based on PHABSIM) (Steffler and Blackburn 2002; Wang et al. 2011). Although the applicability of the methods in obtaining EIFR remains an open debate (Peñas et al. 2014), River2D, which links hydrological regimes and fish habitats, has been widely used to define EIFR. Influenced by climate change, ecological structures and functions change along with hydrological regimes (Hamilton 2002; Campos et al. 2013). In this context, it was assumed that EIFR can change along with ecological structure and function. Furthermore, changes in EIFR can reflect changes in ecological structure. Thus, the objectives of this study were: (1) to explore temporal changes in streamflow and precipitation; (2) to calculate optimum EIFR using FWUA; (3) to analyze decreases in FWUA variation resulting from climate change.
Study area and data description
Juma River Basin
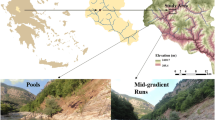
The Juma River, a tributary of the Haihe River, China, is approximately 254 km long with four drainage areas, totaling 810 km2. Originating from a spring-fed lake in Laiyuan County, the river flows into the Daqing River, a tributary of the Haihe River. Mean annual precipitation is from 530 to 660 mm, and mean air temperature is 9 °C. Impacted by climate change and anthropogenic activities (e.g., via the construction of reservoirs and the implementation of soil and water conservation projects), streamflow exhibited decreasing trends and even completely dried up in the Haihe River Basin (Yuan et al. 2005; Bao et al. 2012). Chen et al. (2010), using data obtained from the Zijingguan hydrological station, observed a reduction in streamflow resulting from decreasing trends in precipitation from 1956 to 2005. Gong et al. (2012) similarly found that decreasing trends in precipitation resulted in a reduction of streamflow in the Daqing River Basin. Moreover, the Juma Wild Animal Nature Reserve was instituted to protect the Juma River ecosystem since it is believed to play an important role in maintaining the ecological status of the river system. In this context, understanding changes in EIFR caused by climate change is critical in maintaining the integrity of this aquatic ecosystem (Fig. 1).

Map of the study area showing the location of the Juma River and its location in China
Topographic and hydrological data
Typical inputs of the River2D Hydrodynamic model include bed topography, estimates of initial roughness, discharge at the upper cross section, and water surface elevation at the downstream cross section (Steffler and Blackburn 2002). We assumed that no significant changes in river morphology and channel substrate occurred during the modeling period. Accordingly, we used the Electronic Total Station, a geological mapping tool, to measure the relative X coordinate, Y coordinate, and elevation of a 3-km-long river channel using 234 survey points, recording the substrate at each survey point. We collected discharge data from 1951 to 2005 from the Haihe River Water Conservancy Committee and obtained current discharge rates and values from the monitoring instrument used at the Zijingguan hydrological station. We determined water surface elevation using a rod (surveying pole) along the study area. Data were then loaded into the River2D Hydrodynamic model to generate an initial data file of the Juma River (shown in Fig. 2).

Modeled river bed elevation (a), roughness (b), and water surface elevation (c) in the study area. X and Y are relative coordinates (m)
Materials and methods
The Mann–Kendall test
The nonparametric Mann–Kendall (M–K) test was developed by Mann (1945) and refined by Kendall (1948). It is widely used to determine temporal and spatial trends in hydrological and meteorological time series and can reasonably identify abrupt points of change (Liu et al. 2008; Liang et al. 2011; Yang and Liu 2011). For this study, we used the M–K test with a nominal rejection rate of 5% to detect trends and years of abrupt change in annual streamflow and precipitation at the Zijingguan hydrological station. Test parameters \(a_{ij}\) and \(d_{k}\) are defined as:
where x 1, x 1,…,x n are the time series, and n is the total number of data in the time series. We calculated the expected value \(E\left( {d_{k} } \right)\) and variance \(\text{var} \left( {d_{k} } \right)\) as follows:
We tested the null hypothesis using \({\text{UF}}\left( {d_{k} } \right)\), and if the standard normal probability \(\left| { {\text{UF}}\left( {d_{k} } \right)} \right|\) > u α , the null hypothesis was rejected. \({\text{UF}}\left( {d_{k} } \right)\) constituted the UF curve:
By applying the method to the inverse series, we could obtain the \({\text{UB}}\left( {d_{k} } \right)\) series as follows:
If intersection points of UF and UB were situated between two confidence lines (α = 0.05, μ 0.05 = ±1.96), we assumed that an abrupt change took place at that point.
The River2D hydrodynamic model
The River2D Hydrodynamic model is a two-dimensional, depth-averaged finite element model specifically developed to simulate river flow and fish habitat in a given area (Steffler and Blackburn 2002). The model can simulate transient flow of subcritical–supercritical flow and wet–dry flow by applying changes in surface flow to groundwater flow equations expressed in a conservative format, referred to as Saint–Venant equations. They include three equations and represent the conservation of water mass and the two components of the momentum vector, respectively. They are defined as follows:conservation of mass:
Conservation of x-direction momentum:
Conservation of y-direction momentum:
where H is the depth of flow, and U and V are the depth-averaged velocities in x and y coordinate directions, respectively. q x (q x = HU) and q y (q y = HV) are the respective discharge intensities, which are related to velocity components; g is the gravitational acceleration; ρ is the water density; S ox and S oy are the bed slopes in x and y directions; S fx and S fy are the corresponding friction slopes; and τ ij is the component of the horizontal turbulent shear stress tensor (i, j = x, y).
The River2D Hydrodynamic model is composed of a hydrodynamic model and a fish habitat model. The fish habit module is based on the PHABSIM-weighted useable area (WUA) approach, which is used to determine the optimum flow that can provide the largest physical habitat in rivers. WUA is calculated by summing the product of cell area and its respective combined suitability index (CSI) value. CSI values are obtained from individual habitat suitability index values for depth, velocity, and channel indexes as follows:
where A i is the surface area of a cell in partition i; f(v i ), f(D i ), and f(C i ) are individual habitat suitability index (HSI) values for velocity, depth, and the channel index, respectively, in partition i (shown in Fig. 3).

Velocity (a), water depth (b), and channel index (c) preference curves for Pseudorasbora parva in the Juma River, China
The River2D Hydrodynamic model has been verified by a number of comparisons with theoretical, experimental, and field results (Ghanem et al. 1995; Waddle et al. 1996; Hayes et al. 2007; Kozarek et al. 2014). It has been widely used in China to determine optimum EIFR and to evaluate ecological consequences of instream flow regulations on the basis of the available physical habitat of specific species (e.g., Jiang et al. 2010; Yang et al. 2010; Tan et al. 2011; Sun et al. 2013).
Habitat suitability curves
According to fish species distribution and diversity surveyed by Yang et al. (2013) in the Juma River, the proportion of Pseudorasbora parva (P. parva) for all species caught was 80.1%. Moreover, by way of survey data collected from twenty field sites upstream and downstream of the river, we found that the local populace most often caught P. parva for purposes of cooking. Accordingly, we selected P. parva as the target fish in this investigation on the impact of climate change on physical habitat and calculated suitability curves using velocity, water depth, and channel indexes. The channel index is a categorical variable used in habitat simulation models to describe the structural characteristics of stream channels (Bovee 1986), which we obtained according to channel substrates for P. parva. These values were interpolated from a separate channel index file to computational notes. We obtained relevant habitat preference curves by combining literature research, field investigations, and data from particular species/life stages (Parasiewicz et al. 2012; Xu et al. 2012; Zeng 2012; Wang et al. 2013) (shown in Fig. 3). The optimum range of velocity, water depth, and channel indexes were from 1.0 to 1.5 m/s, from 1.0 to 1.75 m, and from 1.0 to 6.0, respectively.
WUA modification
We calculated f(V i ), f(D i ), and f(C i ) on the basis of the frequency of fish appearing at different velocities, water depths, and channel indexes, and this was based on the hypothesis that f(V i ), f(D i ), and f(C i ) had the same influence as the suitability index did. In addition, values of f(V i ), f(D i ), and f(C i ) were all between 0 and 1. Therefore, if we added an adjusting factor to CSIs and assumed that they had the same influence on final results, such as f(V i ), f(D i ), and f(C i ) would, modifications of WUA could be considered reasonable. Thus, we conducted a “weight” by adding all CSIs calculated from different guaranteed hydrological frequencies and a value of their corresponding discharge frequencies (namely f(F i )) to obtain an optimum dynamic equilibrium between EIFR and the available habitat area. To be more specific, each WUA calculated from a different guaranteed hydrological frequency was multiplied by its corresponding frequency factor. This is referred to as the frequency-weighted usable area (FWUA) method. WUA modifications were conducted as follows:
Furthermore, Leclerc et al. (1995) divided the quality of the physical habitat of fish into different grades in accordance with CSIs: extremely low: 0 < CSI < .1; low: 0.1 < CSI < 0.4; moderate: 0.4 < CSI < 0.7; and high: 0.7 < CSI < 1.0. We adopted this standard to analyze variety in the quality of the physical habitat of fish during normal periods and periods of change during different seasons. In order to validate the accuracy of the modified WUA method (FWUA), we compared our EIFR results to those obtained using other methods, such as the range of variability approach (RVA) method (Richter et al. 1996), the monthly frequency calculation method (MFC) (Yang et al. 2003), and the Tennant method (Tennant 1976).
Results and discussion
Temporal trends in streamflow and precipitation
The M–K test showed that annual streamflow at the Zijingguan hydrological station exhibited an abrupt downward trend in 1981 (α = 0.05) (Fig. 4a). Therefore, annual streamflow at the Zijingguan hydrological station can be divided into two separate periods: a normal period (1956–1980) and a period of change (1981–2005).

Abrupt changes in annual flow series revealed by the Mann–Kendall test (a), and temporal trends in annual streamflow and precipitation taken from the Zijingguan hydrological station between 1956 and 2005 (b)
As shown in Fig. 4, we detected significant decreasing trends (P < 0.05) in annual streamflow and precipitation with an average slope of −0.09 × 108 m3 a−2 and −2.20 mm a−2, respectively. During the normal period, average annual streamflow was 3.1 × 108 m3 a−1, while during the period of change average annual streamflow decreased to 0.5 × 108 m3 a−1. Annual precipitation was 564.3 and 340.4 mm a−1 during the normal period and period of change, respectively. Streamflow showed decreasing trends in precipitation (with a 0.78 Pearson product-moment correlation coefficient).
Optimum EIFR calculated by FWUA
The area of physical habitat for P. parva (Fig. 5) reached a maximum of 4816.8 m2 when instream flow was 65 m3/s. Moreover, the optimum range of flow was from 30 to 75 m3/s and the area of physical habitat for P. parva was >4000 m2 during this period. Furthermore, the area of physical habitat decreased rapidly when flow exceeded 85 m3/s, and the area of physical habitat for P. parva decreased to zero when flow reached 850 m3/s. The traditional approach typically used for areas of physical habitat omits the influence of climate change as well as changes that occur during different seasons. For example, when climate change was taking into account, the maximum high flow threshold for different guaranteed hydrological frequencies was only 66.0 m3/s.

Variation in the area of physical habitat of different flows
As shown in Table 1, the extent of maximum habitat had typically occurred during flow exceedance frequencies of 75% with the exception of the spring and winter seasons that fell within the period of change, which exhibited maximum habitat values during exceedance frequencies of 50%. Therefore, the area of optimum physical habitat can be elicited as shown in Table 2. Results showed that the area of optimum physical habitat dramatically decreased in winter and spring with autumn and summer following in descending order. The loss of the optimum area of physical habitat in winter and spring could be key factors in reducing P. parva populations.
As shown in Table 3, we calculated EIFR during the normal period from FWUA and compared it to results from the RVA, MFC, and Tennant methods. The range of EIFR calculated using FWUA was consistent with the other three methods. FWUA was highly reliable and was more conducive to the maintenance of the ecological health of rivers as well as the protection of instream biological habitats.
FWUA variation under climate change
Figure 6 shows that variations in optimum minimum and maximum flow and FWUA exhibited the same trends during both normal periods and periods of change. During the normal period (1956–1980), high flow threshold and FWUA values all occurred in the following descending order: summer, autumn, winter, and spring, while during the period of change (1981–2005), low flow threshold and FWUA values all occurred in the following descending order: summer, autumn, spring, and winter.

Variation in low and high thresholds of flow and the area of optimum physical habitat (AOPH) for the normal period (a) and the period of change (b) during different seasons
Variation in low flow threshold values and low and high FWUA threshold values during both the normal period and the period of change showed that the degree of change in winter was highest with a value change greater than 76.5% for instream flow and greater than 97.1% for FWUA. It was followed in descending order by spring, autumn, and summer. The high flow threshold by itself showed that winter also experienced the highest changes in flow. Conversely, changes in high flow threshold values in summer exceeded those in spring, reaching 46.2%. At the same time, low flow threshold values during the period of change were greater than during the normal period for summer.
Variation in the quality and distribution of FWUA
Figure 7 provides the four physical habitat grades for all seasons. For the normal period, the four grades during spring, autumn, and winter showed little proportional differences, whereas in summer, physical habitat quality was mostly in the moderate to high levels. During the period of change, moderate and high levels of physical habitat quality in spring and winter ceased altogether, and low and extremely low quality levels accounted for the majority of the habitat. In summer, high levels decreased and low levels increased proportionally. Conversely, moderate and high levels in autumn increased and low and extremely low levels decreased proportionally. Given that the quality of moderate and high physical habitats only changed slightly during the summer and autumn compared to that of winter and spring, the overall degree of change in FWUA for fish habitat during the summer and autumn was not significant when habitat area did not change during the four seasons.

Variation in the quality of fish habitat for the normal period and the period of change during different seasons
The available habitat for P. parva (Fig. 8) was mainly located in the eastern middle reaches of the Juma River during both the normal period and the period of change. Differences were mainly in the extent and degree of CSIs. This means that there was only a slight impact on the distribution of P. parva FWUA (Fig. 8), while there was a significant impact on quality of P. parva FWUA (Fig. 7).

Combined suitability index (CSI) of the high flow threshold for the normal period and the period of change during spring (a, e), summer (b, f), autumn (c, g), and winter (d, h)
Influenced by climate change, streamflow exhibited significant decreasing trends (P < 0.05) with a reduction in precipitation (that also showed significant decreasing trends; P < 0.05) in the Juma River Basin (Fig. 4), which is consistent with results from Chen et al. (2010), Ji et al. (2010), Xu (2011), and Gong et al. (2012). As shown in Fig. 6, climate change greatly reduced the frequency of high flow while increasing the frequency of low flow during summer. A reduction in streamflow could occur by means of associated biological changes, such as growth, breeding and the migration of fish species, the growth rate of vegetation as well as some chemical alterations in water temperature, oxygen content, and photosynthesis (Richter et al. 1996; Ruiz et al. 2013). Alterations in hydrological regimes [e.g., the reduction in frequency of high flow and increases in frequency of low flow in summer (Fig. 6)] could directly influence hydrological processes and riverine ecosystems (Gibson et al. 2005; Yang et al. 2012; Yan et al. 2013; Dindaroğlu et al. 2015). As shown in Fig. 6, low and high pulses exhibited similar decreasing trends, which also led to decreases in area of low and high optimum physical habitat between the normal period and the period of change and resulted in decreases in quantity and quality for the entire habitat area for P. parva in the Juma River (Fig. 7). As a result of the complex interactions between flow regimes and ecological systems (Ryberg et al. 2012; Karimi et al. 2012), changes in seasonal streamflow between the normal period and the period of change dramatically altered the available habitat of P. parva (see Fig. 6, Table 2). Due to the assumption that no significant changes in river morphology and channel substrate occurred, we set the channel index as the steady state during the study period, which is consistent with other studies (e.g., Steffler and Blackburn 2002; Clark et al. 2008; Lee et al. 2010). In fact, changes in hydrological regimes can alter river morphology and channel substrate and result in changes to optimum physical habitat. Considering that changes in hydrological regimes were mainly affected by climate, we used the modified WUA method to calculate optimum physical habitat area and EIFR in the Juma River Basin. Compared to the RVA, MFC, and the Tennant methods, FWUA obtained results consistent with the other methods (Table 3), which indicated that EIFR derived from FWUA is a creditable approach. Furthermore, climate change altered hydrological regimes and affected available habitat (Abbaspour et al. 2012), even for the small river area investigated in this study. Alterations in ecological structure and function that result from changes in hydrological regimes can strengthen and amplify their influencing factors on larger scales. More suitable water management practices should therefore be put into place to maintain natural hydrological regimes (Karimi et al. 2012; Shokoohi and Amini 2014), which could help address the impact of climate change on riverine ecosystems.
Conclusion
EIFR plays an important role in maintaining the balance of river ecosystems and promoting the harmonious and sustainable development of local economies. Using the M–K test, the River2D Hydrodynamic model, and FWUA, we investigated EIFR response to climate change. Several conclusions can be drawn from our results:
-
1.
Impacted by climate change, annual streamflow in the Juma River showed a significant decreasing trend. Specifically, annual streamflow exhibited a downward trend from an abrupt change that occurred in 1981. We used this abrupt change to partition the study period (1956–2005) into a normal period and a period of change;
-
2.
The area of optimum physical habitat for P. parva and instream flow both exhibited significant alteration to climate change. The degrees of change in area of optimum physical habitat and flow magnitude were magnified in winter and followed in descending order by spring, autumn, and summer. The quality of optimum physical habitat was severely degraded; however, the area distribution of optimum physical habitat changed little under climate change;
-
3.
Changes in EIFR and area of optimum physical habitat showed that hydrological regimes impact ecological structures and functions, even in a small river system, and therefore rational measurements of such values should be put into practice to adapt to climate change in small river systems, which could help to maintain their ecological integrity on a large basin scale.
References
Abbaspour M, Javid AH, Mirbagheri SA, Givi FA, Moghimi P (2012) Investigation of lake drying attributed to climate change. Int J Environ Sci Technol 9(2):257–266
Acreman MC, Dunbar MJ (2004) Defining environmental river flow requirements—A review. Hydrol Earth Syst Sci Discuss 8(5):861–876
Bao ZX, Zhang JY, Wang GQ, Fu GB, He RM, Yan XL, Jin JL, Liu YL, Zhang AJ (2012) Attribution for decreasing streamflow of the Haihe River basin, northern China: climate variability or human activities? J Hydrol 460:117–129
Barranco LM, Javier AR, Olivera F, Olivera A, Quintas L, Estrada F (2013) Assessment of the expected runoff change in Spain using climate simulations. J Hydrol Eng 19(7):1481–1490
Bovee KD (1982) A guide to stream habitat analysis using the instream flow incremental methodology. Fish and Wild life Service, Washington, DC
Bovee KD (1986) Development and evaluation of habitat suitability criteria for use in the instream flow incremental methodology. Washington, DC: USDI Fish and Wildlife Service. Instream flow information paper no. 21 FWS/OBS/86/7. http://www.fort.usgs.gov/Products/Publications/pub_abstract.asp?PubID=1183
Campos GEP, Moran MS, Huete A, Zhang YG, Bresloff C, Huxman TE, Eamus D, Bosch DD, Buda AR, Gunter SA, Scalley TH, Kitchen SG, McClaran MP, McNab WH, Montoya DS, Morgan JA, Peters DPC, Sadler EJ, Seyfried MS, Starks PJ (2013) Ecosystem resilience despite large-scale altered hydroclimatic conditions. Nature 494:349–352
Chen FL, Wang J, Yang G, Jia XJ (2010) Analysis of the rainfall and runoff change trend in the Zijingguan basin. J Shihezi U (Nat. Sci.) 28(1):101–105 (in Chinese with English abstract)
Clark JS, Rizzo DM, Watzin MC, Hession WC (2008) Statial distribution and geomorphic condition of fish habitat in streams: an analysis using hydraulic modelling and geostatistics. River Res Appl. 24(7):885–899
Cui BS, Hua YY, Wang CF, Liao AL, Tan XJ, Tao WD (2010) Estimation of ecological water requirements based on habitat response to water level in Huanghe River Delta, China. Chin Geogr Sci 20(4):318–329
Daniel C, Nassir EJ (1994) Comparison and regionalization of hydrologically based instream flow techniques in Atlantic Canada. Can J Civ Eng 22(2):235–246
Dindaroğlu T, Reis M, Akay AE, Tonguc F (2015) Hydroecological approach for determining the width of riparian buffer zones for providing soil conservation and water quality. Int J Environ Sci Technol 12(1):275–284
Ghanem A, Steffler PM, Hicks FE, Katopodis C (1995) Dry area treatment for two-dimensional finite element shallow flow modelling. In: Proceedings of the Canadian Hydrotechn
Gibson CA, Meyer JL, Poff NL, Hay LE, Georgakakos A (2005) Flow regime alterations under changing climate in two river basins implications for freshwater ecosystems. River Res Appl 21:849–864
Gong AX, Zhang DD, Feng P (2012) Variation trend of annual runoff coefficient of Daqinghe River Basin and study on its impact. Water Res Hydrop Eng 43(6):1–4 (in Chinese with English abstract)
Hamilton SK (2002) Hydrological controls of ecological structure and function in the Pantanal wetland (Brazil). In: McClain M (ed) The ecohydrology of South American Rivers and Wetlands, vol 6. International Association of Hydrological Sciences, Wallingford, pp 133–158
Hayes JW, Hughes NF, Kelly LH (2007) Process-based modelling of invertebrate drift transport, net energy intake and reach carrying capacity for drift-feeding salmonids. Ecol Mod 207:171–188
Huckstorf V, Lewin WC, Wolter C (2008) Environmental flow methodologies to protect fisheries resources in human-modified large lowland rivers. River Res Appl 24(5):519–527
IPCC (2007) Climate change 2007: the physical science basis. In: Solomon S et al (eds) Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
Ji ZH, Yang CX, Qiao GJ (2010) Reason analysis and calculation of surface runoff rapid decrease in the Northern branch of Daqinghe River. South North Water Transf Water Sci Tech 2(8):73–76
Jiang XH, Zhao WH, Zhang WG (2010) The impact of Xiaolangdi Dam operation on the habitat of Yellow River Carp (Cyprinus (Cyprinus) carpio haematopterus Temminck et Schlegel). Acta Ecol Sin 302(18):4941–4947
Karimi SS, Yasi M, Eslamian S (2012) Use of hydrological methods for assessment of environmental flow in a river reach. Int J Environ Sci Technol 9(3):549–558
Kendall MG (1948) Rank correlation methods. Hafner, New York
Kozarek JL, Hession WC, Dolloff CA, Diplas P (2014) Hydraulic complexity metrics for evaluating in-stream brook trout habitat. J Hydraul Eng 136(12):1067–1076
Leclerc M, Boudreault A, Bechara JA, Corfa G (1995) Two-dimensional hydrodynamic modeling: a neglected tool in the instream flow incremental methodology. Trans Am Fish Soc 124:645–662
Lee JH, Kil JT, Jeong S (2010) Evaluation of physical fish habitat quality enhancement designs in urban streams using a 2D hydrodynamic model. Ecol Eng 36:1251–1259
Liang LQ, Li LJ, Liu Q (2011) Precipitation variability in Northeast China from 1961 to 2008. J Hydrol 404:67–76
Liu Q, Cui BS (2011) Impacts of climate change/variability on the streamflow in the Yellow River Basin, China. Ecol Mod 222:268–274
Liu Q, Yang ZF, Cui BS (2008) Spatial and temporal variability of annual precipitation during 1961–2006 in Yellow River Basin, China. J Hydrol 361:330–338
Mann HB (1945) Non-parametric test against trend. Econometrika 13:245–259
Milhous RT, Updike MA, Schneider DM (1989) Physical habitat simulation system reference manual-version II. Biological report
Parasiewicz P, Castelli E, Rogers JN, Plunkett E (2012) Multiplex modelling of physical habitat for endangered freshwater mussels. Ecol Mod 228:66–75
Peñas FJ, Juanes JA, Álvarez-Cabria M, Álvarez C, García A, Puente A, Barquín J (2014) Integration of hydrological and habitat simulation methods to define minimum environmental flows at the basin scale. Water Environ J 28(2):252–260
Piniewski M, Laizé CLR, Acreman M, Schneider C (2014) Effect of climate change on environmental flow indicators in the Narew Basin, Poland. J Environ Qual 43(1):155–167
Poff NL (2002) Ecological response to and management of increased flooding caused by climate change. Philos Trans R Soc Lond Ser A Math Phys Eng Sci 360(1796):1497–1510
Richter BD, Baumgartner JV, Powell J, Braun D (1996) A method for assessing hydrologic alteration within ecosystems. Conservat. Boil 10(4):1163–1174
Ruiz F, Abad M, Bodergat AM, Carbonel P, Rodriguez-Lazaro J, Gonzalez-Regalado ML, Toscano A, Garcia EX, Prenda J (2013) Freshwater ostracods as environmental tracers. Int J Environ Sci Technol 10(5):1115–1128
Ryberg KR, Lin W, Asce M, Vecchia AV (2012) Impact of climate variability on runoff in the North-Central USA. J Hydrol Eng 19(1):148–158
Shokoohi A, Amini M (2014) Introducing a new method to determine rivers’ ecological water requirement in comparison with hydrological and hydraulic methods. Int J Environ Sci Technol 11(3):747–756
Song JX, Xu ZX, Liu CM, Li HE (2007) Ecological and environmental instream flow requirements for the Wei River—the largest tributary of the Yellow River. Hydrol Proc 21(8):1066–1073
Steffler P, Blackburn J (2002) Two-dimensional depth averaged model of river hydrodynamics and fish habitat: introduction to depth averaged modeling and user’s manual. University of Alberta
Sun JN, Zhang TQ, Zhu D, Fu J (2013) Simulative evaluation of fish habitat of backwater tributary of Baihetan Reservoir. Water Resour Hydropower Eng 44(10):17–22 (in Chinese with English abstract)
Tan YP, Wang YR, Li J, Liu M (2011) Simulation research on fish habitats in reducing reach of Jinping Dahewan in Yalong River. Water Resour Power 29(3):40–43 (in Chinese with English abstract)
Tennant DL (1976) Instream flow regimens for fish, wildlife, recreation, and related environmental resources. Am Fish Soc 1:6–10
Tharme RE (2003) A global perspective on environmental flow assessment: emerging trends in the development and application of Environmental flow methodologies for rivers. River Res Appl 19:397–441
Tian YY, Wang X, Li CH, Can YP, Yang ZF, Wang YL (2012) Estimation of ecological flow requirement in Zoige Alpine Wetland of southwest China. Environ Earth Sci 66(5):1525–1533
Waddle T, Steffler PM, Ghanem A, Katopodis C, Locke A (1996) Comparison of one and two dimensional hydrodynamic models for a small habitat stream. Ecohydr. Conf., p 10
Wang YK, Xia ZQ, Wang D (2011) Assessing the effect of separation levee project on Chinese sturgeon (Acipenser sinensis) spawning habitat suitability in Yangtze River, China. Aquat Ecol 45(2):255–266
Wang XN, Liu ZT, Yan ZG, Zhang C, He L, Meng SS (2013) Species sensitivity evaluation of Pseudorasbora parva. Environ Sci 34(6):2329–2334 (in Chinese with English abstract)
Xu LJ (2011) Analysis on dynamic feature of the rainfall-runoff relationship in the Daqing river basin under the influence of human activities. South North Water Transf Water Sci Technol 9(2):73–76 (in Chinese with English abstract)
Xu XL, Zhang CJ, Wu JF, Feng ZJ (2012) Study on the tolerance of salinity and pH in the fish, Pseudorasbora parva. Hubei Agric Sci 51(7):1423–1425 (in Chinese with English abstract)
Yan Y, Yang ZF, Liu Q (2013) Nonlinear trend in streamflow and its response to climate change under complex ecohydrological patterns in the Yellow River basin, China. Ecol Mod 252:220–227
Yang ZF, Cui BS, Liu JL, Wang XQ, Liu CM (2003) Theory, method and practice of ecological environmental water requirement. Science Press, Beijing
Yang ZF, Yu SW, Chen H, She DX (2010) Model for defining environmental flow thresholds of spring flood period using abrupt habitat change analysis. Adv Water Sci 21(4):567–574 (in Chinese with English abstract)
Yang ZF, Liu Q (2011) Response of streamflow to climate changes in the Yellow River Basin, China. J Hydrometeorol 12(5):1113–1126
Yang Z, Yan Y, Liu Q (2012) Assessment of the flow regime alterations in the lower yellow river, China. Ecol Inform 10(7):56–64
Yang K, Zhu DM, Wang WM (2013) Oxygen consumption rate and asphyxia point in Topmouth Gudgeon, Pseudorasbora parva. Fisher Sci 32(5):256–260 (in Chinese with English abstract)
Yuan F, Xie ZH, Ren LL, Huang Q (2005) Hydrological variation in Haihe River Basin due to climate change. J Hydraul Eng 36(3):274–279
Zeng Y (2012) Morphometrics analysis of the invasive Pseudorasbora parva from different habitats. J Hydroecol 33(2):115–120 (in Chinese with English abstract)
Zhao JY, Dong ZR, Sun DY (2008) State of the art in the field of River habitat assessment. Sci Technol Rev 26(17):82–88 (in Chinese with English abstract)
Zhong HP, Liu F, Geng LH, Xu CX (2006) Review of assessment methods for instream flow requirements. Adv Water Sci 17(3):430–434 (in Chinese with English abstract)
Acknowledgements
This study was supported by the National Natural Science Foundation of China (No. 51579008), the National Key Basic Research and Development Project (2016YFC0500402), the special fund of the State Key Joint Laboratory of Environment Simulation and Pollution Control (14L01ESPC), and the Open Research Fund Program of the Beijing Climate Change Response Research and Education Center (Beijing University of Civil Engineering and Architecture).
Funding was provided by the Beijing Higher Education Young Elite Teacher Project (Grant No. YETP0259).
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial responsibility: Tan Yigitcanlar.
Rights and permissions
About this article

Cite this article
Liu, Q., Yu, H., Liang, L. et al. Assessment of ecological instream flow requirements under climate change Pseudorasbora parva . Int. J. Environ. Sci. Technol. 14, 509–520 (2017). https://doi.org/10.1007/s13762-016-1166-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13762-016-1166-1