
Abstract
Biosemiotics argues that “sign” and “meaning” are two essential concepts for the explanation of life. Peircean biosemiotics, founded by Tomas Sebeok from Peirce’s semiotics and Jacob von Uexkül’s studies on animal communication, today makes up the mainstream of this discipline. Marcello Barbieri has developed an alternative account of meaning in biology based on the concept of code. Barbieri rejects Peircean biosemiotics on the grounds that this discipline opens the door to nonscientific approaches to biology through its use of the concept of “interpretation.” In this article, it is noted that Barbieri does not adequately distinguish among Peirce’s semiotics, Peircean biosemiotics, and “interpretation-based” biosemiotics. Two key arguments of Barbieri are criticized: his limited view of science and his rejection of “interpretation-based” biosemiotics. My argument is based on tools taken from a different approach: Robert Rosen’s relational biology. Instead of “signs” and “meanings,” the study begins in this case from the “components” and “functions” of the organism. Rosen pursues a new definition of a law of nature, introduces the anticipatory nature of organisms, and defines the living being as a system closed to efficient cause. It is shown that Code Biosemiotics and Peircean biosemiotics can share a common field of study. Additionally, some proposals are suggested to carry out a reading of Rosen’s biology as a biosemiotic theory.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Semiotics studies the systems of signs, their production and functioning. Charles Sanders Peirce (1931–1936) argued that semiosisFootnote 1 requires a triadic relationship among a sign, its object, and its interpretant.Footnote 2
Biosemiotics is the union of biology and semiotics. It starts with the idea that signs and meanings are fundamental components of life, and its purpose is to show how semiosis allows us to explain living beings and to differentiate them from the inanimate world. As some contemporary authors claim:
Signs, not molecules, are the basic units in the study of life. (Hoffmeyer 1997, p. 940)Footnote 3
Sign science and life science are coextensive…. semiotics is biology and biology is semiotics. (Kull 2001, p. 3)
The main lines of research in biosemiotics are grouped in Peircean biosemiotics, a discipline created by Thomas Sebeok from Peirce’s semiotics and Jacob von Uexküll’s studies on animal communication, and later combined with diverse influences from general systems theory.
Marcelo Barbieri, throughout his long career, has developed a theoretical framework that also focuses on the role of signs and meanings in the explanation of biology. However, Barbieri’s biosemiotics has developed outside the mainstream, leaning on the concept of code. After a stage of collaboration with other schools, Barbieri departed from the biosemiotics field, considering that Peircean biosemiotics opens the door to nonscientific approaches through the concept of interpretation.
This article shows that Barbieri’s rejection of Peircean biosemiotics is based on: (1) a limited conception of science; and (2) an incorrect understanding of Peircean biosemiotics, which misunderstands the scope of application of Peirce’s semiotics and does not take into account the rest of the theories on which Peircean biosemiotics is based. Our goal is to show how Peircean biosemiotics and Barbieri’s Code Biology can be part of a common field.
Barbieri’s work is studied with tools taken from Robert Rosen’s relational biology. This author has been chosen for three reasons, in addition to the solidity and beautyFootnote 4 of his work. First, it provides a concept of science that allows Barbieri’s identification of science with mechanism to be overcome. Second, Rosen’s biology can be seen as a basis for Peircean biosemiotics and, therefore, allows for a better understanding of the concepts of that discipline. And finally, by relying on an author who is alien to the discussions of biosemiotic schools, the problem can be analyzed with a novel perspective.
To understand Barbieri’s arguments, it is first necessary to briefly state the basic concepts of Peirce’s semiotics and Peircean biosemiotics. This is the first section.
In the second section, Barbieri’s framework for the study of biology is laid out. His proposals of Code Biosemiotics and his criticism of interpretation-based biosemiotics (which Barbieri identifies with Peircean biosemiotics) are discussed.
The third section discusses the development of Rosen’s relational biology that leads him to the concept of anticipatory systems, and the (M, R)-system as a model of organismic organizations. Rosen characterizes living entities as systems that instantiate closure to efficient cause.
These first three sections put together for the reader the necessary elements of the theories to be analyzed. They explain that there are different approaches and objectives, but also shared concepts (that are accepted or rejected, with the same or different meaning), and imply by themselves a first comparison.
In the fourth section, two key proposals of Code Biology are criticized with arguments taken from Rosen’s relational biology. The relationship between science and mechanism is discussed, and Barbieri’s critique of the concept of interpretation (and with it his rejection of Peircean biosemiotics) is analyzed.
In section five, as an additional result of the research carried out, some lines of a study of Rosen’s biology as a biosemiotic theory compatible with Code Biology and Peircean biology are sketched.
It is concluded that: (1) Code Biology provides the adequate theoretical tools for the development of biological research; (2) it is possible to develop other scientific biosemiotics within the framework of Peircean biosemiotics; (3) Code Biosemiotics and Peircean biosemiotics, despite their different approaches, can be seen as a common framework for biology; and (4) Rosen’s biology can also be characterized as a type of biosemiotics.
Peirce’s Semiotics and Peircean Biosemiotics
Peirce’s Semiotics
Peirce’s theory of signs is central to his work on logic and his scientific studies, and constitutes a philosophical alternative to Cartesian dualism.
In Peirce’s words:
It is important to understand what I mean by semiosis. All dynamical action, or action of brute force, physical or psychical, either takes place between two subjects (whether they react equally upon each other, or one is agent and the other patient, entirely or partially) or at any rate is a resultant of such actions between pairs. But by “semiosis” I mean, on the contrary, an action, or influence, which is, or involves, a cooperation of three subjects, such as a sign, its object, and its interpretant, this tri-relative influence not being in any way resolvable into actions between pairs… and my definition confers on anything that so acts the title of a “sign.” (Peirce 1931–1936, vol. 5, p. 484)
Important ideas can be highlighted from the previous quotation. First, to develop a theory of logic and to study nature, a type of actions that cannot be characterized as ordinary physical actions must also have been considered: semiotic actions. Peirce’s semiotics is a logical and scientific study of dynamic sign action in nature. Second, a semiotic action requires a three-factor relation, a triad. Third, Peirce introduces the interpretant, the most distinctive feature of his account.
As AtkinFootnote 5 points out, an interpretant should be understood as an interpretive process in a receptive system, an effect upon someone, the understanding an agent reaches of some sign/object relation, the translation or development of the original sign. The meaning of a sign is manifest in the interpretation that the interpretant generates in sign users. The three components of the triad are closely related:
For Peirce, then, any instance of signification contains a sign-vehicle,Footnote 6 an object and interpretant. Moreover, the object determines the sign by placing constraints which any sign must meet if it is to signify the object. Consequently, the sign signifies its object only in virtue of some of its features. Additionally, the sign determines an interpretant by focusing our understanding on certain features of the signifying relation between sign and object. (Atkin 2013, Sect. 1.3)
From Zoosemiotics to Biosemiotics
Thomas Sebeok (1963, 1972) extended the communication based on signs, characteristic of human beings, to the animal world, giving rise to zoosemiotics. His work had a key antecedent in the evidence of semiosis in the animal world that Jacob von Uexküll (1928) had presented much earlier. Von Uexküll’s main proposal, the unwelt, is fundamentally a semiotic or biosemiotic concept. The unwelt is the representation of the environment inside an animal. The organism interacts with its environment in terms of how it perceives it. From this communication, the environment induces changes in the organism, and the organism modifies the environment.
Combining von Uexküll’s unwelt and Peirce’s semiotics, Sebeok founded biosemiotics as a new paradigm for all biology:
Because there can be no semiosis without interpretability—surely life’s cardinal propensity—semiosis presupposes the axiomatic identity of the semiosphere with the biosphere. (Sebeok 2001, p. 68)
Peircean Biosemiotics
As Søren BrierFootnote 7 explains:
Peircean biosemiotics is based on Peirce’s theory of mind as a basic part of reality, (in Firstness) existing in the material aspect of reality, (in Secondness) as the “inner aspect of matter” (hylozoism) manifesting itself as awareness and experience in animals, and finally as consciousness in humans. Combining this with a general systems theory of emergence, self-organization and closure/autopoiesis, it forms an explicit theory of how the inner world of an organism is constituted and, therefore, how first-person views are possible and just as real as matter. (Brier 2008, p. 40)
Following the path initiated by Sebeok, various scholars have developed their proposals within the framework of Peircean biosemiotics, especially in the Copenhagen and Tartu schools of biosemiotics, particularly Jesper Hoffmeyer, Claus Emmeche, Frederik Stjernfelt, Søren Brier, and Kalevi Kull. All of them share a common paradigm for the study of biology, although they differ in the weight that the different theories that have shaped Peircean biosemiotics have in their proposals, as shown in the following examples. First:
The theories of Heinz von Foerster and Humberto Maturana & Francisco Varela have had significant influence on the development of the Copenhagen school of biosemiotics. This school focuses mainly on a new interpretation of biology and life as having an important communicative aspect to their organization. They look at the basically biological aspect of biosemiotics especially as endosemiotics, and its significance for understanding ecological and hereditary relations in a non-reductionistic evolutionary view; it opposes views like Richard Dawkins’ theory of selfish genes. (Brier 2008, pp. 47–48)
Second, even belonging to the same school, Hoffmeyer and Brier differ when analyzing different classes of biosemiotics:
Thus Hoffmeyer sees unity on the ontological level including the consequences of an evolutionary view, as Peirce does, but qualitative differences on the epistemological level because the semiotic freedomFootnote 8 changes radically when we move into the level of symbol use in language…. Agreeing with Hoffmeyer on these points I (Brier) still think that that there are also important epistemological continuities between the zoosemiotic and the anthroposemiotics levels in the form of the understanding of knowledge coming from Peirce’s philosophy of the three categories, his theory of abduction and its connection to deduction and induction. (Brier 2008, pp. 47–48)
As a final example, relevant to our work, we highlight the importance of interpretation and learning in Hoffmeyer’s biosemiotics. For Hoffmeyer (2009), genes do not specify the phenotype. Gene expression can be explained as a process of interpretation, in which the instructions of the environment condition the genetic instructions to be executed. Semiotic interactions do not depend on a causal connection between sign and meaning but on the interpretation of an agent, which is based on communication and learning. Hoffmeyer argues that life is essentially historical, in the sense that its continuation depends on the ability to learn, to remember the strategies that have been effective, to interpret them, and to apply them when interacting with the environment. Furthermore, learning requires some type of coded representation. In other words, learning is a semiotic process.
Code Biology: A Framework for the Study of Biology
Code Biosemiotics
The starting point for Code BiosemioticsFootnote 9 is the assertion that what defines semiosis is coding, in which “a code is always a set of rules that establish a correspondence (or a mapping) between two independent worlds” (Barbieri 2003).
Barbieri’s biosemiotics is based on two key ideas: (1) cells contains an internal codemaker (Barbieri 1981, 1985), and (2) coding cannot be reduced to copying, so natural selection (based on copying) and natural conventions (based on coding) are two different mechanisms of evolution (Barbieri 1985, 2003).
The Cell and Organic Codes
Barbieri’s analysis of organic codes begins with the study of the cell model that emerges with the genetic code.Footnote 10 Barbieri considers that a cell should not be seen as a duality of genotype and phenotype but as a triad of genotype, phenotype, and ribotype: the genetic code requires the existence of internal codemakers.
In general, a semiotic system is
a system made of two independent worlds that are connected by the conventional rules of a code. A semiotic system, in conclusion, is necessarily made of at least three distinct entities: signs, meanings and code. (Barbieri 2008, p. 578)
This proposal gives priority to the codemaker. The codemaker is the agent of semiosis, while signs and meanings are its instruments. It is the codemaker that creates signs and meanings, which do not exist (as such) outside a codemaking process. In Barbieri’s words “a semiotic system is a set of signs, meanings and code that are all produced by the same agent, i.e., by the same codemaker” (Barbieri 2015, p. 30).
Barbieri focuses on organic codes, in which the worlds to connect are two sets of different molecules. In this case, a third type of molecular structures, which act as adapters and independently bind to molecules of both sets, is required. The adapters establish an indirect correspondence that does not respond to the physical properties of the molecules of the connected sets but to conventional rules. Only an authentic code guarantees biological specificity.Footnote 11 Adapters are organic codemakers, i.e., molecules that reveal the existence of organic codes.
These considerations can be applied not only to the genetic codeFootnote 12 but also to the other codes and codemakers that can be identified in living beings. Barbieri’s works (2015, Sect. 3) analyze various organic codes at the cellular and supracellular levels and include a large number of references to works on the identification of this type of code (2015, Introduction). Examples of organic codes are splicing codes, signal transduction codes, cytoskeleton codes, and compartment codes. In the fourth section there is a subsection entitled “Barbieri’s Concept of Mechanism,” in which Barbieri’s account of signal transduction codes is summarized.
Information, Sign, and Meaning
Barbieri (2008) considers the classification of signs into natural signs and conventional signs. In conventional signs, there is no physical relationship between signs and meanings; each relationship is defined by an arbitrary rule (convention). Barbieri analyzes nucleotides as signs and concludes that they behave as natural signs in the copying process, but they are also conventional signs in the coding process. In the first case, organic information comes into play, while the second gives rise to organic meaning. Organic information and organic meaning are not, therefore, intrinsic properties of the molecules that carry them but the results of natural processes. Faced with the cohabitation of two different paradigms of biology, the “chemical paradigm” (the idea that life is chemistry) and the “information paradigm” (the idea that life is chemistry + information), Barbieri poses that: “(T)he copying of the genes and the coding of proteins are equally fundamental processes, and this leads to a third theoretical framework that is referred to as the ‘code paradigm’ (the idea that life is chemistry + information + codes)” (Barbieri 2015, p. XV).
Barbieri generalizes these considerations for all organic codes and provides the following definitions. The sequence used (or produced) by a copymaker during a copying process is organic information. The sequence used by a codemaker during a coding process is an organic sign. The sequence produced by a codemaker during a coding process is an organic meaning. An organic code is a set of rules of correspondence between signs and meanings.
For Barbieri (2015), recognizing the existence of a code, whether mental (or neural) or organic, entails affirming the existence of a meaning. A code establishes a correspondence between two entities and, with it, converts one into a sign and another into its meaning. If the code is neural, the meaning is a mental entity; if the code links two types of molecules, the sign and meaning are organic entities. It is possible, therefore, to affirm the existence of organic signs and meanings if the existence of organic codes is demonstrated. Barbieri proposes as a biological research goal the identification of organic codes at all levels as an integral part of life.
Unification and Rupture
Barbieri (2009) reports that in 2004, several schools of biosemiotics agreed to place what united all of them (the introduction of meaning in biology) ahead of their differences and to consider Sebeok’s (2001) claim that life and semiosis are coextensive as a foundational principle of biosemiotics. This principle and the assertion that signs, meanings, and codes are natural entities were agreed upon as necessary and sufficient conditions for the establishment of biosemiotics as a natural science and as the basis of minimal unity among different schools.
However, at the end of 2012, Barbieri left the common field of biosemiotics and, together with 11 colleagues, founded the International Society of Code Biology (ISCB). Barbieri and his colleagues said they were rebelling against what they considered the application of Peirce’s semiotics to all living beings and the growing presence of humanities studies in biosemiotics. These researchers considered it necessary to preserve the scientific approach, and abandoned the biosemiotic community to constitute a different theoretical framework for biology: Code Biology. In the constitution of the ISCB, Code Biology is defined as “the study of all codes of life with the standard methods of science” (Barbieri 2014, p. 247).
Codes Versus Interpretation
Barbieri (2009) compares the following definitions of code-based and interpretation-based semiotics:
The necessary and sufficient condition for something to be a semiosis is that A provides a conventional association between B and C, where A is a set of adaptors and B and C are the objects of two independent worlds. (Barbieri 2008, p. 236)
The necessary and sufficient condition for something to be a semiosis is that A interprets B as representing C, where A is the interpretant, B is an object and C is the meaning that A assigns to B. (Posner et al. 1997, p. 4)
Peircean semiotics argues that all living beings, from a cellular level, maintain a behavior that depends on their context and that they can do so because they are able to interpret the world. Conversely, Barbieri argues that a cell can respond to the changing environment without performing any interpretation. A combination of several organic codes is sufficient to show context-dependent behavior. At the cellular level, he advocates for a unique semiosis based on organic codes, which can play three different and complementary roles: (1) manufacturing (production of new objects, such as proteins); (2) signaling (association of objects from independent worlds, as in signal transduction); and (3) regulation (as in the control of transcription of genes). Biosemiotics based on interpretation leads, according to Barbieri, to the abandonment of scientific objectivity and the conversion of biology into a division of the humanities.
Theoretical Frameworks for the Study of Biology
Barbieri (2015) analyzes the Modern Synthesis and systems biology as main theoretical frameworks for the study of biology. The Modern Synthesis focuses on the population aspects of the problem of life, and its main mechanism (its only mechanism of evolution) is natural selection. Systems biology prioritizes the study of the individual organism as a system that is self-manufactured, and its central process is autopoiesis (self-production).
As a critique of the Modern Synthesis, Barbieri believes that evolution requires a second mechanism of evolution, natural conventions, based on codes. Further, faced with systems theory, he asserts that the organism is not explained by autopoiesis but by codepoiesis, which is the ability to acquire codes and maintain them. For Barbieri, “the cell is a codepoietic system, i.e., a system that is capable of creating and conserving its own codes” (2015, p. 176). There are two distinct versions of Code Biology. In the restricted version, only organic codes are studied. In the general version, Code Biology examines all codes of life: organic, neural, and language.Footnote 13 Accordingly, Barbieri proposes Code Biology as a third and very ambitious framework for biology.
The stated intention of the International Society of Code Biology is the study of all codes of life using the standard methods of science. Barbieri argues that this intention is already applied rigorously in the study of organic codes, but he admits that the study of neural and language codes remains a more speculative field that requires both scientists from various disciplines and linguists and philosophers (Barbieri 2015).
Science and Mechanisms
According to Barbieri, to understand something is equivalent to explaining it with a model that is a mechanism, which has the logic of a machine. Further, scientific knowledge is obtained by constructing mechanistic models of what we observe in nature.
Barbieri considers the study of organic codes to be a strictly scientific discipline since it proposes a model from which to perform experiments: a mapping between two sets of independent molecules interconnected by a third set of molecules that act as adapters. The discovery of adapters reveals the presence of organic codes. The semiosis of Barbieri is explained by the mechanisms by which adapters associate signs and meanings. In addition, coding “is a generative mechanism, a mechanism of evolution” (Barbieri 2015, p. 180).
Relational Biology of Robert Rosen
Relational biology began with Nicolas Rashevsky (1954) and his attempt to develop a “mathematical” biology. Initially, Rashevsky tried to model each of the organismic functions and formalize the whole organism from its components. However, he soon became convinced that this strategy was not viable and, therefore, proposed that it is necessary to focus on the search for a principle that governs the organization, rather than on the phenomena themselves.
Continuing the line of work initiated by his professor, Rosen considered that the study of life must be based on the notion of function and on the relationships among the components of the organism. He proposed a theoretical framework for the study of biology that escapes the conception of science that has remained valid since Newton. Rosen argued that anticipatory behavior, which depends on the existence of predictive models, is the general rule of biological systems. He defined a type of system, the (M, R)-system, which incorporates the fundamental functions of living beings.
Laws, Models and Mechanisms
Laws of Nature and Models
Rosen (1991) explains that the concept of law of nature is based on the assertion that there exists, and can be established, a relation between two different modes of implication: the causality of natural systems and the inferential implication of formal systems. This relation is established through a model.
Consequently, a natural law would be the congruence between two systems of implications: one corresponding to a natural system and the other to a formal system that is a model of the natural system. (See Fig. 1 for Rosen’s representation of it.)

Modified from Rosen (1991, p. 60, Fig. 3H.2)
Natural law and model.
As Rosen described it, formal system F is a model of a natural system N as long as we obtain the same result in the following two cases:
-
1.
Moving from one phenomenon to another in N; and
-
2.
Encoding (E), applying F, and decoding (D).
In this case, F is a model of N, and N is a realization of F.
The model defines an encoding of the qualities of N into the formal objects of F. The objects of F are vehicles of signs that represent the qualities of N with which they are associated. The formal relations among the objects of F (the inferences that occur among them) correspond, once F is decoded into N, to causal relations among the qualities of the system N. Inferences in F allow us to make predictions about N.
A natural system N can have two or more formal models. One might wonder what types of models are possible, what the class of all formal models of N is, and whether there is a model that is the largest.
Models and Analogy
Consider two systems, N1 and N2, which can be encoded into the same formal system F, as shown in Fig. 2.

Modified from Rosen (2012, p. 75, Fig. 2.2)
Two natural systems encoded into the same formal system.
N1 and N2 share a common model and can be considered alternative realizations of formal system F. N1 and N2 are analogous systems. As Rosen (2012) suggests, E2 − 1E1 can be seen as the encoding of N1 into N2, and E1 − 1E2 as the encoding of N2 into N1. Thus, a relation of analogy establishes a modeling relation between two natural systems. N1 contains a model of N2, and N2 contains a model of N1.
These outcomes are shown in Fig. 3.

Analogy between two natural systems
Once an analogy has been established between two natural systems, we can learn about one of them by studying the other. As is explained in the “Anticipatory Systems” section below, the concept of analogy is the basis of the definition of an anticipatory system.
Models and Metaphors
Figure 4 shows two natural systems that encode into two different formal systems. Consider the case in which formal systems F1 and F2 satisfy some mathematical relation, although there is no objective function between sets of encoded propositions that preserve the structure. In this case, an analogy between N1 and N2 cannot be established (a system is not a model of the other), but the existence of a certain relationship between the two natural systems can be affirmed. We say that N1 is a metaphor of N2 (and N2 is a metaphor of N1), indicating that their models share some property. This notion of metaphor also allows for learning about a natural system from the study of the other. Rosen acts metaphorically in his analysis of the concepts of fitness, adaptation, natural selection, and evolution, as well as in the concept of learning (see the “Anticipatory Systems” section).

Modified from Rosen (2012, p. 76, Fig. 2.3)
Two natural systems that encode into two different formal systems.
Model and Simulation
The concept of simulation provides a method for comparing formalisms that is different from modeling.
Consider Fig. 5. Let F1 and F2 be two formalisms such that F2 is a model of F1. An inference in F1 equates to an encoding of F1 into F2 plus an inference in F2 plus a decoding of F2 into F1. Note that the inferential structure of F1 is not implicated by F2.

Modified from Rosen (1991, p. 53, Fig. F.2)
Modeling between two formalisms.
Let’s now analyze a simulation. Consider that, in Fig. 5, F1 is a map to be simulated, and F2 is the simulator. To perform the simulation, the map F1 is incorporated into F2. The left part of the graph (F1, E, and D) disappears inside the right part: F1 becomes the effect of F2. In this case, F1 is simulated by F2, congruence between two inferential structures is not established, and we cannot learn anything about the simulated map from its simulation.
According to Rosen, a mapping is simulable (or computable) if it is definable by an algorithm, i.e., if it is evaluable by a mathematical (Turing) machine.
Mechanisms
Rosen (1991) argues that an organism is different from a mechanism. He claims that a natural system N is a mechanism if and only if all its models are simulable, and his analysis concludes that, if a natural system N is a mechanism, then:
-
1.
N has a maximal model that is the largest of all its models and includes everything that can be known about N, according to the laws of nature;
-
2.
There is a finite set of minimal models of N;
-
3.
The maximal model is equal to the direct sum of the minimal models, allowing us to define states for the maximal model in terms of the minimal models;
-
4.
In the category of all models of N, analytic and synthetic modeling coincide (the direct product is equal to the direct sumFootnote 14); and
-
5.
Every property of N is fractionable.Footnote 15
Rosen’s exposition concludes that, unlike a mechanism, an organism cannot be decomposed as a direct sum, i.e., it is not fractionable into components.
Rosen (1991) studies mechanisms from a relational perspective (see the “(M, R)-Systems” section for Rosen’s definitions of relational model and efficient cause) and analyzes the implications among their different parts. For each component f, the answer to the question “why f?” (that is, the efficient cause of f) must be found in another component of the system or in the environment. Rosen’s analysis concludes that there can be no closed path of efficient causation in a mechanism. Conversely, an organism is a system in which all its components are implied by other components of the system. Organisms manifest maximal implication.
According to these arguments, mechanisms are systems in which syntactics and semantics coincide. Conversely, an organism, like a natural language, possesses semantic modes of implication that do not appear in any formal piece that we can extract and study syntactically. The concept of mechanism is a (restrictive) means of expressing a law of nature. Mechanisms are specializations. A mechanism is a purely syntactic construct; it is a simple (not complex) system. Unlike mechanisms, an organism is a complex system, in which complexity should not be seen as a property of a particular coding but as the capacity to define different codings of a natural system.Footnote 16
That a system is not a mechanism does not mean that it cannot have mechanistic models. However, in an organism, the limit of its mechanistic models is not a mechanism; an organism is a system congruent with an impredicative model.Footnote 17 A living system must have non-computable models.
According to Rosen, physics is the science of mechanisms and a special science, while biology is a more general science that studies complex systems that should consider more types of models, some of which are non-simulable.
Anticipatory Systems
Definition of Anticipatory Systems
Tentatively, Rosen defines an anticipatory system as
…a system containing a predictive model of itself and/or of its environment, which allows it to change state at an instant in accord with the model’s predictions pertaining to a later instant. (Rosen 2012, p. 313)
Later, Rosen (2012, Sect. 6.1) specifies the concept by requiring five conditions that a system must fulfill to be an anticipatory system, leading to the following formal definition:
An anticipatory system S2 is one which contains a model of a system S1 with which it interacts. This model is a predictive model; its present states provide information about future states of S1. Further, the present state of the model causes a change of state in other subsystems of S2; these subsystems are (a) involved in the interaction of S2 with S1, and (b) they do not affect (i.e., are unlinked to) the model of S1. In general, we can regard the change of state in S2 arising from the model as an adaptation, or pre-adaptation, of S2 relative to its interaction with S1. (2012, pp. 317–318)
Since S1 and S2 are two natural systems, we can see that the basis of the concept of an anticipatory system is the property of an analogy between two natural systems.
Adaptive Behavior and Anticipatory Systems
Rosen (2012, Sect. 6.5) explains metaphorically the concepts of fitness, adaptation, natural selection, and evolution. He begins by studying a simple organism that can move on a two-dimensional surface and develops its metaphor, idealizing the type of behavior found. From this metaphor, he extracts the essential aspects that characterize any system in which the concepts of fitness, adaptation, and selection can be defined. Following this line, Rosen states that adaptive behavior implies anticipation and that “a behaviour or phenotype which is adaptive necessarily is of an anticipatory character” (Rosen 2012, p. 345). Moreover, “(t)he retrospective or reactive mode through selection generates adaptation, becomes converted in the adapted organism to a prediction about how present behaviour will affect future behaviour” (2012, p. 346).
Finally, Rosen shows that the organism analyzed meets the conditions required in the rigorous definition of an anticipatory system.
In conclusion, selection and adaptation in fact generate specific predictive models, in such a way that the behavior of an organism at an instant of time bears a definite relation to an internal prediction about a later instant. Additionally, although without developing his statement, Rosen claims that “a general theory of macroevolution can readily be built on the framework we have introduced, incorporating all of the traditional biological features of the Darwinian picture” (2012, p. 350).Footnote 18
Learning and Anticipatory Systems
Rosen (2012, Sect. 6.6) analyzes the relation between evolution and learning. Both concepts turn out to be inseparable, and show how learning is the basis of anticipatory ability.
The apparently disparate phenomena of evolution and of learning are in fact linked to each other, in the sense that a metaphor for the one is, at the same time, a metaphor for the other. In fact, we can translate an evolutionary metaphor into a learning metaphor by means of a specific mapping process in which observables of the former are simply re-interpreted, or translated, into observables of the latter. From this it will immediately follow, from the arguments of the preceding chapter, that learning processes generate predictive models. (Rosen 2012, p. 352)
[Learning] can be regarded either as a part of adaptation, or as a metaphor for it. (Rosen 2012, p. 357)
(M, R)-Systems
The Relational Model
The formal systems proposed by Rosen to model natural systems are relational systems. Consider a separable part of a system. The difference in the behavior of the complete system, versus the case in which a part has been separated, defines the function of the separate part. A component is a part of the system that can be assigned a function and is the basic unit of the organization of the system. It can be represented as a mapping between two sets, f: A → B, where
-
A → B represents the flow from A to B; and
-
f corresponds to the efficient causeFootnote 19 exerted on that flow, which is the efficient cause of B (of each element of B).
See Fig. 6. We can say that the mapping

A component f
-
encodes component f; and
-
is an implication that can be expanded as
 for every a Є A, being f(a) = b Є B. (Hollow-headed arrows represent the flow, and solid-headed arrows symbolize the effect of the component.)
for every a Є A, being f(a) = b Є B. (Hollow-headed arrows represent the flow, and solid-headed arrows symbolize the effect of the component.)
 for every a Є A, being f(a) = b Є B. (Hollow-headed arrows represent the flow, and solid-headed arrows symbolize the effect of the component.)
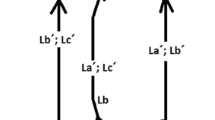
for every a Є A, being f(a) = b Є B. (Hollow-headed arrows represent the flow, and solid-headed arrows symbolize the effect of the component.)The models of natural systems are abstract block diagrams (ABDs). A combination of mappings constitutes an ABD of the relational system to which it corresponds and the organization it describes. However, mappings can also be elements of sets; for example, f belongs to the set of all mappings that can be set between A and B. Therefore, augmented abstract block diagrams (AABDs) can be defined, which could include mappings that are the results of other mappings, increasing the organizational level of the system described. At the limit, all elements of the diagram could be implied by others.
Closure to Efficient Cause
As Mossio (2013) notes, the concept of closure, in general terms, designates a feature of biological systems by virtue of which their constitutive components and operations depend on each other for their production and maintenance and collectively contribute to determining the conditions under which the system itself can exist.
Among the closure proposals, mention should be made of operational closure (Varela 1979), the closure of catalytic functions (Kauffman 2000), and the closure of constraints (Moreno and Mossio 2015). These three closure models respond to the problems of metabolism and self-maintenance of an organizationally closed but thermodynamically open system.
In Rosen’s account, a material system is an organism if and only if it is closed to efficient cause (Rosen 1991).
(M, R)-Systems
Rosen proposed the (M, R)-system first as a cell metaphor (2012)Footnote 20 and then as a general model for an organism (1991).
To change from a self-organized system to a living being, closure to efficient cause must resolve not only the problem of metabolism but also the self-repair and organizational invariance of the system.
According to Rosen, an organism is a closure among the following three classes of functions.
-
Metabolism f: A → B
-
Repair Φ: B → f
-
Replication β: f → Φ
The relationship among the three classes of functions, as shown in Fig. 7, gives rise to what Rosen called the metabolism-repair system or (M, R)-system.Footnote 21

Modified from Rosen (1991, p. 251, Fig. 10C.6)
An (M, R)-system.
Anticipation in (M, R)-Systems
As Pattee (2007) points out, Life Itself (hereafter LI; Rosen 1991) represents a qualitative change compared to Rosen’s previous work, specifically versus Anticipatory Systems (hereafter AS; Rosen 2012). In both works, the modeling relation is the same, but its application is different:
In LI Rosen no longer characterized life by its internal predictive models that allow adaptive evolution. Instead he develops a timeless relational concept of organisms based on natural and inferential “entailments” derived from Aristotle’s causal categories …. (T)he central issue in LI is no longer the evolving predictive model in the organism, but the limitations of formal models in our brain. (Pattee 2007, p. 2274)
However, in both AS and LI, the concept of anticipation is key to the definition of an organism. (M, R)-systems—which in AS are a metaphor for the cell, while in LI they are a model—respond to this requirement. According to Rosen:
Anticipatory behaviour is in fact damned as “acausal”, because causality is construed precisely as allowing only the past to affect the present. I initially softened this by interposing a “predictive model” as a transducer between now and later. But nevertheless, the presumed telic or finalistic aspects of anticipation seemed to violate the one-way causal flow on which “objective science” itself is presumed to rest. And I noticed that my own (M, R)-systems have an inherent anticipatory aspect, built into their very organisation. (Rosen 2006, p. 11)
Rosen argues as follows. In mathematics, the analogue to anticipation is impredicativity. An (M, R)-system is a mathematical construction that inherently manifests an impredicative loop.Footnote 22 In conclusion, an (M, R)-system is a system that includes anticipatory behavior.
With (M, R)-systems, Rosen reaffirms the anticipatory character as an intrinsic property of organisms. Without needing to return to the technical definition of anticipatory systems, the concept of anticipation can be applied to (M, R)-systems: the model of an (M, R)-system allows the system to change its state in an instant, according to its predictions about a later instant.
Relational Biology Versus Code Biology
Having analyzed the theoretical frameworks proposed by Barbieri and Rosen, I will now discuss some key proposals of the former in light of the latter.
Science and Mechanisms
The Meanings of “Mechanism”
Mechanisms have received renewed attention in the philosophy of biology during the last 25 years. Among the new mechanists William Bechtel, Carl Craver, Lindley Darden, Stuart Glennan, and Peter Machamer stand out. It is beyond the scope of this work to analyze the points in common and the differences between the proposals of these scholars.
To compare the role that Rosen and Barbieri give to the mechanisms in science and, more specifically, in biology, it is convenient to clarify what meaning each author gives to the concept of mechanism. My argument will lean on the analysis of Nicholson (2012). According to this author, in biology the term “mechanism” is used with different meanings,Footnote 23 which can be defined as follows:
- (a)
Mechanicism the philosophical thesis that conceives living organisms as machines that can be completely explained in terms of the structure and interactions of their component parts.
- (b)
Machine mechanism the internal workings of a machine-like structure.
- (c)
Causal mechanism a step-by-step explanation of the mode of operation of a causal process that gives rise to a phenomenon of interest. (Nicholson 2012, p. 153)
The causal mechanism sense of “mechanism” has become predominant today in biology. Causal mechanisms enable the identification of causal relations. To inquire about the causal mechanism of a phenomenon is to inquire about the causes that explain how it is brought about.
Traditionally, mechanicism and machine mechanisms share an ontological status.Footnote 24 Conversely, Nicholson argues that causal mechanisms “are actually better understood as heuristic models which target specific causal relations and thereby facilitate the explanation of the particular phenomena scientists investigate” (2012, p. 154). And,
This view of causal mechanisms significantly departs from the ontic conception of them as autonomous systems akin to machine mechanisms (defended by Glennan, Bechtel, and at times by Craver…), given that the parts of a causal mechanism do not even need to be structurally demarcated. All that matters is that they are causally relevant to the production of the explanandum phenomenon. (Nicholson 2012, p. 160)
Contemporary philosophers of science refer to mechanistic explanations, even when they appeal to causal mechanism. However, “Mechanistic explanations are ones in which wholes are accounted for in terms of the structure and interactions of their parts” (Nicholson 2012, p. 154). Nicholson refers to mechanismic explanations for the explanations given in terms of causal mechanisms.
Barbieri’s Concept of Mechanisms
Barbieri (2015) provides a definition that partially conforms to the machine mechanism concept. He talks about machine-like models and gives descriptions of fully functional working systems. To propose his mechanisms based on codes, he starts from the examples of mechanistic models of mechanicism and machine mechanism: the clock-machine, steam-engine-machine, and the computer-machine.
However, Barbieri’s mechanisms should better be understood as causal mechanisms. In his analysis of the functioning of codes (Barbieri 2008, 2015, Chap. 5), he explains step-by step the causal processes that give rise to the phenomena studied.
Consider, for example, Barbieri’s explanation of the signal transduction codes. The transfer of information from environment to genes takes place in two distinct steps: one in which the external signals (first messengers) are transformed into internal signals (second messengers), and a second path from second messengers to genes, which is known as signal integration. Barbieri, quoting Alberts et al. (2007), highlights that there are hundreds of first messengers (hormones, growth factors, neurotransmitters, etc.), whereas the known second messengers are only of four types (cyclic AMP or GMP, calcium ions, inositol trisphosphate, and diacylglycerol). First and second messengers belong to two very different worlds, and there is no necessary connection between first and second messengers because it has been proved that the same first messengers can activate different types of second messengers, and that different first messengers can act on the same type of second messengers. The explanation proposed by Barbieri is that signal transduction is based on organic codes, which must be identified finding the molecules that play the role of adaptors. In Barbieri’s words:
The transduction system consists of at least three types of molecules: a receptor for the first messengers, an amplifier for the second messengers, and a mediator in between (Berridge 1985). The system performs two independent recognition processes, one for the first and the other for the second messenger, and the two steps are connected by the bridge of the mediator. The connection, however, could be implemented in countless different ways since any first messenger can be coupled with any second messenger, and this makes it imperative to have a code in order to guarantee biological specificity.
In signal transduction, in short, we find all the three characteristics of the codes: (1) a correspondence between two independent worlds, (2) a system of adaptors that give meanings to molecular structures, and (3) a collective set of rules that guarantee biological specificity. (Barbieri 2008, p. 588)
Barbieri’s explanation of signal transduction codes is comparable to that given by Nicholson (2012) of the causal mechanism for the membrane trafficking of the delta-opioid receptor (DOR) induced by pain stimulation. What Nicholson affirms of that causal mechanism can also be applied to Barbieri’s description of signal transduction codes:
It is a step-by-step explanation of the mode of operation of the signal transduction pathway …. The causal mechanism is abstracted both temporally and spatially… [O]nly the features that are causally relevant … (i.e., the explanandum phenomenon) are featured… [T]he organismic context (in this case, the cell) is almost completely abstracted away and yet it is heavily presupposed, as it provides the enabling conditions that are ultimately necessary. (Nicholson 2012, pp. 160–161)
Barbieri’s mechanisms should be seen as heuristic models with causal relations that facilitate the explanation of the phenomena. The explanation is not based on the general and autonomous organization of a machine but on the parts of a causal mechanism that are causally relevant to the production of the explanandum phenomenon.
Barbieri does not attribute to mechanisms an ontological status, and makes clear his rejection of the typical characteristics of mechanicism: “Mechanism is not reductionism…is not determinism…is not physicalism…” (Barbieri 2015, p. 16).
Barbieri argues that the scientific method is based on the definition of models, and identifies mechanisms with models, which leads him to identify mechanism with the scientific method: “Mechanism, in other words, is ‘scientific modelling’” (Barbieri 2015, p. 16). Finally, Barbieri admits the limitations that models have in explaining reality.
Rosen’s Concept of Mechanism
We now analyze the concept of mechanism in Rosen’s work according to Nicholson’s taxonomy. First, Rosen’s concept of mechanism (discussed above) is intrinsically linked to the definition of law of nature inherited from Newton and prevailing in contemporary science. Therefore, his definition of mechanism should be considered as a formalization of the machine mechanism concept, although deprived of any ontological status.
Rosen rejects mechanistic biology. As we have seen, he argues that a mechanism is a restrictive way of expressing a law of nature that is not sufficient to account for biology. He does not reject the use of mechanisms as an explanation of some organismic biological processes, but holds that there may be processes in an organism that cannot be explained as mechanisms; and, in any case, a complete description of an organism cannot be a description of a mechanism. An organism is a complex system that can have mechanistic (and nonmechanistic) models, but the limit of which is not a mechanism.
On the other hand, Rosen shares with present-day biologists the use of the term “mechanism” in the study of biological processes. For example, Rosen (2012) includes expressions like “sensory mechanism,” “effector mechanism,” “morphogenetic mechanisms,” “mechanism for replication,” “anticipatory mechanisms,” “selection mechanism,” or “integrating mechanisms.” However, Rosen bases his explanations of the corresponding processes on the concept of metaphor. Consider, for instance, his accounts of morphogenesis and natural selection.
On morphogenesis, he holds that:
The biologist knows that there are three basic kinds of processes underlying all specific morphogenetic or developmental phenomena. These are: (1) Differentiation…; (2) Morphogenetic movement…; and (3) Differential birth and death.… It will be observed that these are exactly the three processes manifested metaphorically in the Ising model,Footnote 25 arising from the different interpretations of state transitions of individual lattice elements. Thus, the Ising model provides us with metaphors for all of these basic morphogenetic mechanisms. (Rosen 2012, p. 180; emphasis and footnote added)
Regarding natural selection, Rosen’s mathematical study of the movement of an idealized organism on a two-dimensional surface, such as the one mentioned in the “Anticipatory Systems” section above, leads to a definition of a selection mechanism. If natural selection actually took place in that two-dimensional surface, it could be argued that Rosen describes a machine mechanism. However, the metaphorical approach to the selection mechanism in the real world makes the argument and results comparable to a causal mechanism.
It is important to highlight that Rosen’s metaphorical explanations share with the mechanismic explanations, based on causal mechanism, that both are idealized representations, abstractions of the processes being studied.
Some Criticisms of Rosen’s Account of Mechanism
Before comparing the conceptions of science of Rosen and Barbieri, it is convenient to discuss a couple of criticisms that are posed to Rosen’s arguments.
First, it can be questioned if the new mechanists’ account of mechanism invalidates Rosen’s arguments. For example, Bechtel (2007) analyzes Rosen’s work from his mechanistic perspective and concludes that the rupture with mechanistic science can be avoided by considering self-organization, autonomy, and closure to efficient cause as a help “to fill out the picture of what mechanisms are capable of doing when they are organized appropriately” (p. 273).
According to Bechtel, “[a] mechanism is a structure performing a function in virtue of its components parts, component operations, and their organization.… The orchestrated functioning of the mechanism is responsible for one or more phenomena” (Bechtel 2006, p. 26). Moreover, his explanation of biological phenomena of an organism (such as digestion, cell division, and protein synthesis) must take into account not only the organization of the components and operations of the mechanism, but the entire organism, that is, “the particular modes of organization that are required in biological systems” (Bechtel 2007, p. 269).
To save mechanistic science, Bechtel adds system organization, typical of systems biology, to causal mechanism. This addition strengthens mechanistic science (and systems biology). However, it does not justify the claim that an organism is a mechanism.
Furthermore, according to Nicholson (2012), system organization is not present in mechanismic explanations (including Bechtel’s explanation of biological processes):
[M]echanismic explanations specify only those features of the underlying causal networks that biologists deem most relevant for manipulating and controlling the phenomena whilst at the same time presupposing a great deal of the organismic context that makes them possible. (Nicholson 2012, p. 159)
With his extended definition of mechanism, Bechtel seeks to avoid the problem of laws in biology:
Until the recent rise of mechanist accounts, most philosophical accounts of explanation viewed universal laws as the key element in an explanation.… This has seemed particularly problematic in the context of biology. (Bechtel 2007, footnote 2)
Bechtel, like Barbieri, identifies science with mechanism. In contrast, Rosen argues the need for (new) laws for biology that bring us closer to understanding what life is, notwithstanding accepting other tools (models, analogies, metaphors, simulations, even mechanisms) to understand the biological processes.
Finally, Bechtel’s mechanistic science, unlike Rosen’s biology, does not adequately distinguish between model and nature. According to Nicholson (2012, p. 158):
The fact that the overwhelming majority of mechanismic philosophers speak of them as “real systems in nature” (Bechtel 2006, p. 33) I attribute to an inadvertent transposition of the ontic status of machine mechanisms… onto the notion of causal mechanism.
Consider now a second criticism of Rosen’s proposals. Rosen’s claim that biological systems are not mechanisms is directly related to another claim, i.e., that organisms have non-simulable models. To be more precise, what Rosen states can be decomposed into two statements: (R1) an organism must have non-simulable models; and (R2) an (M, R)-system is a general model for an organism. Some authors have tried to refute Rosen’s statements, by means of both formal demonstrations and practical examples.
For instance, an analysis in terms of λ-calculus (Mossio et al. 2009) concludes that a system closed to efficient causation can certainly have computable models. However, Cárdenas et al. (2010) offer a detailed reply to Mossio’s analysis and claim that:
The conclusion of Mossio et al. (2009) that (M, R)-systems can have computable models is based on an analysis of the fundamental equations of (M, R)-systems in terms of the theory of computer programming, specifically in terms of λ-calculus. Their analysis omits an essential part of the argument, however, and arrives in consequence at a result that we contest. (Cardenas et al. 2010, p. 85)
Furthermore, the examination of the work of other authors leads Cárdenas et al. (2010, p. 90) to a more general conclusion: “Efforts to mathematically disprove Rosen’s contention that an organism cannot have simulable models have not resolved the question.”
In addition, Cornish-Bowden and his group have proposed a biochemical model of an (M, R)-system, and they have simulated it in a computer. They account for a simulation of a model of an (M, R)-system (i.e., a simulation of a model of a model of an organism) that, as they explain:
In terms of this distinction between models and simulations, the results that we shall discuss in the remainder of this paper are clearly not computer models of organisms, but they can still be models of (M, R)-systems, which themselves incorporate some (but not all) of our understanding of the way an organism is organized. (Cornish-Bowden et al. 2013, p. 387)
And, moreover:
In attempting here to relate the biochemical model to Rosen’s mathematical formalism we have oversimplified some points in the hope of remaining intelligible in chemical terms. It is hardly possible at the present state of understanding to resolve all the problems and arrive at a mathematically rigorous analysis, but we can note some points that will need to be clarified in the future. (Cardenas et al. 2010, p. 85)
Consequently, Rosen’s statements R1 and R2 cannot be considered formally refuted. On the other hand, Cornish-Bowden’s model does not exclude the need for an additional mathematical analysis of Rosen’s model.
Barbieri’s and Rosen’s Conceptions of Science
Although Barbieri’s and Rosen’s definitions of mechanism are different, both have points in common that are relevant to our argumentation. For both Barbieri and Rosen, mechanisms are only models, and science is constructed from models. Both scholars use mechanistic explanations, and also other explanations based on causal abstractions of the processes they study (mechanismic explanations and metaphorical explanations). However, while Barbieri identifies mechanism with scientific modeling, according to Rosen’s account, the scientific task should not be restricted to mechanistic models. In Rosen’s words:
My (M, R)-systems inherently manifest such an impredicative loop; one which cannot be straightened out without losing everything. They are thus not approachable via “machines” in the usual sense; they are not purely syntactic objects. They are what I call complex; they must have non-computable models. I would argue that, precisely by excluding temporally closed causal loops, and indeed by identifying this exclusion with science itself, we have lost not only life, in my sense, but most of its material basis, its physics, as well. To invoke a parallel mentioned earlier: just as the “closed system” is too impoverished, to special, to be a basis for (say) the physics of morphogenesis, exactly so is the simple system, one which can be described entirely as software to a machine, too impoverished to accommodate the living. In fact, these two situations are closely related, but it would take too long to explain that relation here. (Rosen 2006, p. 12)
Other authors (for example, Kauffman 2000), with arguments different from those of Rosen, also defend the non-simulability of living beings, the basis of our criticism of Barbieri’s limited conception of science.Footnote 26 Further studies must also consider that the claim that mechanisms are not sufficient to explain an organism is closely related to Gödel’s incompleteness theorem.
The science of life can draw not only from Newton’s sources but also from Rosen’s and others. The study of biological mechanisms and, in particular, of those associated with coding, is indisputable as a scientific method, but is not necessarily the only possible way to carry out a scientific biosemiotics.
Interpretation
Interpretation According To Barbieri
It is necessary to first present some considerations about the fact that Barbieri does not distinguish properly among three different (although related) theories and concepts: Peirce’s semiotics, Peircean biosemiotics, and interpretation.
As we have seen in the first section, Peircean biosemiotics is more than the application of Peirce’s semiotics to biology (other varied influences contribute to this field of study); and within Peircean biosemiotics, there are important differences between schools and authors, both in the application of Peirce’s semiotics, and in the weight they give to the other theories that make up Peircean biosemiotics. When Barbieri rejects biosemiotics, he really seem to refer to his reading of Hoffmeyer’s work.
On the other hand, it should be kept in mind that Barbieri’s definition of interpretation-based semiosis is taken from Posner and not directly from Peirce. Surely, putting the weight on the interpretant rather than on the interpretation would bring Code Biology closer to Peircean biosemiotics.
In any case, what Barbieri rejects is the application of the concept of interpretation in biology, and this is what we analyze in this section.
Barbieri (2015) discards interpretation-based semiosis because:
-
Interpretation is based on abduction, and abductive reasoning is extrapolation, i.e., the jump to a conclusion from limited data;
-
What is interpreted is not the world but representations of the world; and
-
The result of the interpretation depends on the environment and previous experience (interpretation requires memory).
According to Barbieri, it is not appropriate to define decoding at the cellular level as a type of interpretation. He holds that it is necessary to distinguish between two forms of semiosis: one based only on the processes of encoding and decoding; and another also based on interpretation.
He argues that semiosis based only on codes is the only type present at the cellular level. He specifies it for the process of protein synthesis, although he makes it extendible to the remaining organic codes:
-
The existence of the genetic code demonstrates the existence in the cell of organic semiosis based on a code;
-
The genetic code does not depend on interpretation, it is always the same; and
-
The genetic code has only internal meaning (sense, in Frege’s (1892) terminology, not reference).
However, even in prokaryotic cells, the application of the genetic code is not automatic. Gene expression is subject to a regulation that depends on the interaction between the regulatory proteins and the environment of the cell. According to Barbieri, it is the combination of a signal transduction code with the genetic code that allows the cell to regulate the synthesis of proteins according to the signals that arrive at the cell from its surroundings, which in turn causes the cell to acquire behavior dependent on the environment (Barbieri 2015). Moreover, this could be extended to more complex processes simply by adding more codes. What Barbieri proposes is the construction of a complex mechanism assembling elementary mechanisms that correspond to each code.
Conversely, the animal world, in addition to code-based semiosis, would also require interpretation-based semiosis, which includes two types of meanings: sense and reference.
Barbieri (2015) believes that Peircean biosemiotics implies adopting an ad hoc definition of interpretation, and argues against this approach.
-
This approach would only lead to a reformulation, in terms of Peirce’s semiotics, of what we already know about biological processes. What would be achieved would be a new description, but not new knowledge.
-
Interpretation-based biosemiotics assumes, in fact, the approach of the humanities, betting more on storytelling than on the mechanisms as instruments of study of the processes of life.
-
Only a scientific approach to biosemiotics can lead to new scientific knowledge.
-
Code Biosemiotics (Code Biology) aims to define a theoretical framework to perform experiments and obtain new scientific knowledge.
Interpretation In Rosen’s Work
Rosen’s proposals for the explanation of organisms (from the individual cell)—anticipatory systems and (M, R)-systems—include models that go beyond the mechanisms and imply a semiosis that includes and surpasses code-based semiosis.
On the one hand, an anticipatory system contains a model of itself and/or its environment. Those systems can change their state in an instant according to the predictions of the model about a later time. Consequently, it seems appropriate to state that the system interprets what its model indicates about what might occur in the future, from its state and the environment in the present.
On the other hand, as seen above, (M, R)-systems have an inherent anticipatory character constructed in their organization. This capacity of anticipation is an intrinsic property of organisms. The (M, R)-systems are theoretical tools that grasp the ability of the organism to interpret the information that resides in its organization. The behavior of organisms represents neither internal organization nor external information but interpretations of one through the other.Footnote 27
Moreover, predictive capacity depends on the environment and the previous experience (learning, memory) of the system,Footnote 28 which are qualities that Barbieri attributes to interpretation and the reason for which he rejects the use of this concept. As explained above, Rosen stated that (1) selection and adaptation generate predictive models; (2) learning can be seen as a part of adaptation or as a metaphor for it; and (3) learning processes generate predictive models.
If Barbieri rejects the concept of interpretation as the basis of cellular semiosis, it is because he believes that this concept opens the door to nonscientific formulations of biosemiotics. However, his association between interpretation and the humanities is not a mandatory consequence of the concept of interpretation but an abuse of the concept by some theorists that can be considered a pretext to abandon the rigor of science. Like Barbieri, Rosen also seeks to lay the foundation for a new scientific study of biology.
As an additional consideration, it is worth noting that when we speak of interpretation and meaning in the study of the cell, we refer to interpretation performed “by the cell” and meaning produced and used “by the cell.” Both concepts must therefore be linked to the utility of the associated processes (encoding and decoding, or interpretation) by/for the cell itself. This linkage unites the justification of the concepts of meaning and interpretation to the concept of function, which is the starting point of Rosen’s approach.
Rosen’s Biology as Biosemiotics
From the comparison of the theoretical frameworks of Barbieri and Rosen, it is possible to obtain an additional conclusion, which I point out in this section but will leave its fuller development to future study: Rosen’s biology not only provides support for biosemiotics, it can also be read as a biosemiotic theory.
Code-Based Biosemiotics
The concept of code (or mapping between sets) is present in Rosen’s proposals in various ways. First, the congruence between a natural system N and a formal system F, which constitutes the definition of a natural law, is based on the encoding of the qualities of N into the formal objects of F.
Second, the analogy between two natural systems N1 and N2 that share a common model assumes a modeling relation between both systems, in which the qualities of N1 are encoded into the qualities of N2. N1 and N2 are two independent worlds joined by coding rules (a code), such as those proposed by Barbieri, and this concept of coding between natural systems, associated with the concept of analogy, is the basis of the definition of an anticipatory system.
Third, abstract block diagrams that Rosen proposed as models of natural systems are constructed with mappings. The (M, R)-system, proposed by Rosen as a model of organisms, is nothing more than a special type of abstract block diagram, in which the mappings condense three types of functions (three types of encodings): metabolism, repair, and replication. As previously stated, Barbieri speaks of two independent worlds connected by a code (two sets of molecules bound by adapters, which are organic codemakers); and Rosen of two sets related by a mapping, which is the efficient cause that leads from each element of the first set to an element of the second. The formal similarity between both definitions requires, however, a clarification. In an organism, many processes in which elements of two different worlds are connected by a third type of elements (or processes) are carried out. Only some of these connections are arbitrary, and only arbitrary connections correspond to a code. Likewise, not all the mappings of an augmented abstract block diagram necessarily represent processes subject to arbitrary efficient causes. Only those mappings that correspond to arbitrary processes are comparable to Barbieri’s codes. Nevertheless, both Barbieri’s explanation of biological specificity and Rosen’s defense of non-simulability of organisms imply that life requires some arbitrariness. Surely, the parallelism between Barbieri and Rosen can be reinforced through the concept of interpretant. Peirce speaks of sign, object, and interpretant; and Brier (2008, p. 50), explaining the concept of “code” in Peircean biosemiotics, states:
A code is a set of process rules or habits (for instance how the ribosome works) which connect elements in one area (for instance genes) with another area (for instance a sequence of amino acids in proteins) in a specific meaning context (here the procreation, function, and survival of the cell) …. From Peircean biosemiotics one argues that codes are part of triadic sign processes where an interpretant makes the motivated connection between objects and signs (representamens). The functioning of living systems is based on self-constructed codes. (Brier 2008, p. 50)
Finally, it should be noted that Rosen distinguishes two different types of encodings: (1) the establishment of a correspondence between a natural system N and a formal system F; and (2) the construction of F from components (mappings, codes). Barbieri assumes the encoding and decoding between N and F and does not explicitly distinguish between natural system (the cell) and its formalization (the cell model): his notion of code refers directly to what happens inside a biological system.
Rosen’s biology not only meets the requirements demanded by Barbieri for Code Biosemiotics but also allows the role played by the codes for the cell itself to be distinguished from what they mean to the biologist who studies it.
Peircean Biosemiotics
In the “Relational Biology versus Code Biology” section, we have seen that Rosen’s proposals, both anticipatory systems and (M, R)-systems, conform to interpretation-based biosemiotics. The importance given by Rosen to previous experience, learning, and memory has been highlighted, and it has been seen that Rosen’s concepts of memory and interpretation are consistent with Hoffmeyer’s, as discussed in the first section.
We will only mention here, without developing them, some ideas that reinforce the argument that Rosen’s relational biology fulfills the conditions for inclusion in the field of biosemiotics. First, an analogy can be drawn between Rosen’s efficient cause and the interpretant. Rosen’s mappings conform to Peirce’s definition, quoted above, of semiosis and to the characterization of the interpretant that we have summarized from Atkin. Second, as indicated by Brier, self-organization and closure are two of the concepts on which Peircean biosemiotics is built. Both anticipatory systems and (M, R)-systems interpret models that are implemented in their own organization, and closure to efficient cause is the key concept in Rosen’s definition of life.
In conclusion, Rosen’s biology can be seen as a basis for Peircean biosemiotics, but can also be read as a Peircean biosemiotic theory.
Conclusions
Barbieri and Biosemiotics
The incorporation of a discipline from the humanities (semiotics) to the study of life requires determining what requirements should be demanded in the construction of a scientific framework for biology. Pattee indicates that biosemiotics must decide whether or not to be a science and act accordingly:
if Biosemiotics claims that symbolic control is the distinguishing characteristic of life, and if it also claims to be a science, then it must clearly define symbols and codes in empirical scientific terms that are more familiar to physicists and molecular biologists. (Pattee and Kull 2009, p. 328)
Barbieri’s answer to this concern is Code Biology. The pillars of this proposal for the study of biology can be summarized as follows: (1) meaning is an essential component explaining biology; (2) what defines semiosis in the cell is coding (not interpretation); (3) a code is a set of (conventional) rules that establish the correspondence between two independent worlds; (4) in organic codes, codemakers are molecules that are independently attached to molecules of two different types; (5) it is necessary to introduce a second mechanism of evolution (natural conventions) that arises from codification; and (6) the aim is to define a new framework for the scientific study of biology, in which science and mechanism are assimilated.
According to Barbieri, the framework of Code Biology conforms to the requirements of standard science. The concepts of organic information, organic signs, organic meaning, and organic codes are operationally defined. Organic codes are the basis for conducting experiments and obtaining new scientific knowledge. Therefore, we must conclude that Code Biology is a type of biosemiotics and has all of the elements to be considered as a genuine scientific discipline.
Barbieri from Rosen´s Perspective
In this article, some of Barbieri’s proposals have been criticized, using arguments taken from Rosen’s biology. The emphasis has been placed on two major points. First, Barbieri identifies mechanism with scientific modeling, which leads him to limit the study of biology to the identification and study of mechanisms, specifically, coding mechanisms. In contrast, although Rosen admits the use of mechanisms in scientific research, he does not limit modeling to them. According to this author, physics is the science of mechanisms, while biology also requires nonmechanistic models. Each mapping that is part of a block diagram can be assimilated to a code-signs-meanings triad (and, with it, to a coding mechanism), but the (M, R)-system (the model of an organism) is neither a mechanism nor (only) a set of mechanisms.
Second, Barbieri rejects the possibility of a scientific biosemiotics based on interpretation. However, Rosen’s anticipatory systems and (M, R)-systems respectively interpret an internal model of the system and the organization of the system itself. In Rosen’s work we can find answers to Barbieri’s criticisms of the use of interpretation in biosemiotics: in his relational biology, interpretation has a different meaning from that assumed by humanities disciplines, to the extent that it is not limited to a reformulation of what we already know and can lead to new scientific knowledge. Like Code Biology, Rosen’s biology aims to define a theoretical framework to obtain new scientific knowledge.
Barbieri rejects interpretation at the cellular level, branding it as unscientific, and that caused his rupture with Peircean biosemiotics. However, as has been argued in this article, Rosen’s approach offers us the theoretical tools to develop a different approach in which (1) mechanisms are a key tool for scientific research, but science is not synonymous with mechanisms; (2) the scientific study of biology requires defining the laws of nature with a different (broader) approach than that of the science inherited from Newton; and (3) scientific biosemiotics can be developed including the concept of interpretation, although not all uses of this concept in biology conform to what is required of science.
Rosen’s biology provides support for Peircean biosemiotics to be considered an appropriate framework for the development of the science of biology; a framework compatible with Code Biology, which should aim to provide new scientific conceptual tools that allow for the generation of new knowledge.
The point is that, as Favareau indicates:
[T]he goal of biosemiotics is to extend and to broaden modern science, while adhering strictly to its foundational epistemological and methodological commitments—it does not seek in any genuine sense of the term to “oppose” much less “supplant” the scientific enterprise, but to continue it, re-tooled for the very challenges that the enterprise itself entails, if not demands. (Favareau 2006, p. 4)
Biosemiotics in the Work of Rosen
As an additional result of the study carried out here, it has been pointed out that a biosemiotic reading of Rosen’s biology can also be conducted.
On the one hand, for Rosen coding is a core notion. It is the basis of concepts such as modeling, natural law, anticipatory systems, or (M, R)-systems. This prominent role of coding in Rosen’s definition of life is in accordance with the requirements demanded by Barbieri for Code Biosemiotics.
On the other hand, interpretation is also present in Rosen’s biology. Anticipatory systems and (M, R)-systems interpret organismic and environmental information and rely on previous experience (memory) to make predictions about their future. Furthermore, Rosen’s biology can be analyzed as a Peircean theory.
Pattee and Kull (2009) agree on the presence of semiosis in Rosen’s work. According to Pattee, relational biology depends on semiotic relations, rather than on material ones.Footnote 29 As Kull claims, Rosen practiced biosemiotics, probably without being conscious of it or, at least, without using that term.
The study of Rosen’s biology as a biosemiotic theory deserves further work. A promising starting point could be to develop the potential connections between the concepts of interpretant and efficient causation; and between the interpretation that an organism performs and the closure to efficient causation that it realizes.
Change history
22 August 2018
In the ‘Barbieri’s Concept of Mechanisms’ section on page 12 of above mentioned article.
Notes
The term “semiosis” describes the process of producing and responding to signs. Semiotics is the study of semiosis.
Peirce writes: “I define a sign as anything which is so determined by something else, called its Object, and so determines an effect upon a person, which effect I call its interpretant, that the later is thereby mediately determined by the former” (1998, p. 478).
Robert Rosen was also a physicist and mathematician, and in these disciplines the beauty of the theories is a criterion that counts.
Atkin (2013) provides an interesting overview of Peirce’s theory of signs.
Peirce used different terms for the signifying element including “sign,” “representamen,” “representation,” and “ground.” John Deely (1990) introduced the notion of “sign vehicle” to interpret and develop Peirce’s semiotics.
See Brier (2008) for a detailed study of Peircean biosemiotics.
Hoffmeyer defines semiotic freedom as “the ‘depth of meaning’ that an individual or a species is capable of communicating” (1996, p. 62).
Over four decades, Barbieri has been developing a theoretical framework for the study of biology. What this author began by calling Semantic Biology or Biosemantics (Barbieri 1985) was then changed to Code Biosemiotics and finally to Code Biology. These name changes correspond to three stages, which do not respond to changes in his proposals, but in his relations with other lines of research: a first stage of work independent of other schools that also consider semiosis to be the key to the study of biology; a second stage in which an attempt was undertaken to unify all schools under the common framework of Biosemiotics; and a third in which Barbieri and his collaborators took an independent path again, rejecting the prevailing line in Biosemiotics, considering it based on interpretation and often far from scientific work. In a letter to the editor of Biological Theory, published in 2014, Barbieri explained the reasons that led him to join the Biosemiotic movement in 2001 and to abandon it in 2012 to establish Code Biology as an independent research field.
The application of (conventionally established) rules determines which pairs of molecules are related. This relationship gives each molecule of the first set a meaning in the second set, and the expression of that meaning defines biological specificity.
In the case of the genetic code, the correspondence between codons and amino acids can only be the result of conventional rules. The genetic code had to appear at the same time as the translation apparatus. The result is a codemaker, a machine to produce proteins from a template and a code.
Barbieri classifies life into three worlds. The first world is based only on organic semiosis. Its mechanism is coding, and its codes are organic codes. The second world incorporates animal semiosis, which is characterized by two mechanisms (coding and interpretation) and two types of codes (organic and neural). The third world adds human semiosis. It is based on three mechanisms (coding, interpretation, and language) and three types of codes (organic, neural, and language).
According to Rosen (1991), a natural system N can have analytical and synthetic models. Analytical models are tied to the idea of the Cartesian (or direct) product of quotient spaces, while synthetic models are linked to the idea of the direct sum of subspaces.
Consider a property of system N, embodied in a model M, which is smaller than the maximum model, and the maximum model can be decomposed as the direct sum of two summands, with M being one of them. If this process can be performed for all of the properties of N, then we say that N can be fractionated, and its properties are fractionables.
“I call a system which is not simple ‘complex’. Complex systems cannot be exhausted by any finite number of simple (mechanical) models; they cannot be described as software to a ‘machine’” (Rosen 2006, p. 21).
A definition is said to be impredicative if it invokes the set being defined. It is a property that important mathematicians and logicians, especially Russell, wanted to leave out of their disciplines, but it has long been proved that elimination leads to having to abandon much of mathematics.
Unlike Barbieri, Rosen argues that only one mechanism of evolution is necessary: natural selection.
Taking Aristotelian causes as a reference, we can consider that A is the material cause of B (each element of A is the material cause of an element of B), whereas f is the efficient cause of the occurrence of the flow, i.e., the efficient cause of B (of each element of B).
The first edition of Anticipatory Systems dates from 1985.
“β/B” has been used instead, where “B” appears in Rosen’s original diagram, to avoid the usual error of identifying β with B, or with an element b of set B. According to Rosen (1991), β is the inverse of an operator constructed from b ∈B.
See Rosen’s quotation in the next section.
Nicholson shows the problems generated by not separating the three meanings, even among the new mechanists, quoting an example (Craver and Darder 2005, p. 234) in which “(T)he concept is used in different senses, sometimes even in the same passage (Nicholson 2012, p. 154).”
As Nicholson (2012, p. 154) explains, “The new mechanism discourse is not committed to a mechanistic worldview, nor does it prescribe a mechanistic approach in biology.”
The Ising model is usually considered a mathematical model of ferromagnetism, although Rosen points out: “[T]he Ising model is a metaphor for phase transitions, and not a ‘model’ in our sense” (Rosen 2012, p. 179; italics in original).
It has been argued that Rosen’s assertions on non-simulability of (M, R)-systems have not been refuted. In any case, although the non-simulability of the (M, R)-systems was refuted, as long as the non-simulability of the organisms is maintained, the claim that an organism is not a mechanism, and, therefore, the science of biology must be broader than the science of mechanisms remains valid. This would be sufficient to support our criticism of Barbieri’s limited vision of science, and to defend the compatibility of Barbieri’s biosemiotics with Peircean biosemiotics.
Note that this last sentence, which here applies to Rosen, is literally included in Brier’s explanation of Hoffmeyer (Brier 2008, p. 49).
Pattee’s biosemiotics also requires memory. Biological constraints can only occur in individuals with memory maintained by natural selection. Only individuals with memory-based control can learn from the environment and evolve (Pattee and Kull 2009).
“(H)is emphasis on ‘relational biology’ depended on semiotic rather than material relations. Rosen and I were friends for many years beginning with our studies of hierarchy theory in the 1960s. To us, hierarchies, like measurement, are also dependent on semiotic distinctions because hierarchical levels are recognized by the necessity of different descriptions” (Pattee and Kull 2009, p. 327).
References
Alberts B, Johnson A, Lewis J et al (2007) Molecular biology of the cell, 5th edn. Garland, New York
Atkin A (2013) Peirce’s theory of signs. In: Zalta EN (ed) The Stanford encyclopedia of philosophy (summer 2013 edition). https://plato.stanford.edu/archives/sum2013/entries/peirce-semiotics/
Barbieri M (1981) The ribotype theory on the origin of life. J Theor Biol 91:545–601
Barbieri M (1985) The semantic theory of evolution. Harwood Academic Publishers, London
Barbieri M (2003) The organic codes: an introduction to semantic biology. Cambridge University Press, Cambridge
Barbieri M (2008) Biosemiotics: a new understanding of life. Naturwissenschaften 95:577–599
Barbieri M (2009) A short history of biosemiotics. Biosemiotics 2:221–245
Barbieri M (2014) From biosemiotics to Code Biology. Biol Theory 9(2):239–249
Barbieri M (2015) Code Biology: a new science of life. Springer, Cham
Bechtel W (2006) Discovering cell mechanisms: the creation of modern cell biology. Cambridge University Press, Cambridge
Bechtel W (2007) Biological mechanisms: organized to maintain autonomy. In: Boogerd F, Bruggerman F, Hofmeyr JH, Westerhoff HV (eds) Systems biology: philosophical foundations. Elsevier, Amsterdam, pp 269–302
Berridge M (1985) The molecular basis of communication within the cell. Sci Am 253:142–152
Brier S (2008) The paradigm of Peircean biosemiotics. Signs 2:20–81
Cárdenas ML, Letelier JC, Gutierrez C et al (2010) Closure to efficient causation, computability and artificial life. J Theor Biol 263(1):79–92
Cornish-Bowden A, Piedrafita G, Morán F et al (2013) Simulating a model of metabolic closure. Biol Theory 8(4):383–390
Deely J (1990) Basis of semiotics. Indiana University Press, Bloomington
Emmeche C, Kull K, Stjernfelt F (2002) Reading Hoffmeyer, rethinking biology. Tartu Semiotics Library Vol. 3. Tartu University Press, Tartu
Favareau D (2006) The evolutionary history of biosemiotics. In: Barbieri M (ed) Introduction to biosemiotics: the new biological synthesis. Springer, Berlin, pp 1–67
Frege G (1892) Über Sinn und Bedeutung. Zeitschrift für Philosophie und philosophische Kritik 100:25–50. English translation in Geach P, Black M (eds) (1952) Translations from the philosophical writings of Gottlob Frege. Blackwell, Oxford
Hoffmeyer J (1996) Signs of meaning in the universe. Indiana University Press, Bloomington
Hoffmeyer J (1997) The swarming body. In: Rauch I Carr GF (eds) Semiotics around the world: synthesis in diversity. Proceedings of the 5th congress of the International Association for Semiotic Studies. Berkeley 1994. Mouton de Gruyter, Berlin, pp 937–940
Hoffmeyer J (2009) Epilogue: biology is immature biosemiotics. In: Deely J, Shrocci L (eds.) Proceedings of the 33rd annual meeting of the Semiotic Society of America. Legas Publishing, Ottawa, pp 927–942
Kauffman S (2000) Investigations. Oxford University Press, New York
Kull K (2001) Jakob von Uexküll: a paradigm for biology and semiotics. Semiotica 134(1/4). Mouton de Gruyter, Berlin
Moreno A, Mossio M (2015) Biological autonomy: a philosophical and theoretical enquiry. Springer, Dordrecht
Mossio M (2013) Closure, causal. In: Dubitzky W, Wolkenhauer O, Cho K-H, Yokota H (eds) Encyclopedia of systems biology. Springer, New York, pp 415–418
Mossio M, Longo G, Stewart J (2009) A computable expression of closure to efficient causation. J Theor Biol 257(3):489–498
Nicholson D (2012) The concept of mechanism in biology. Stud Hist Philos Biol Biomed Sci 43(1):152–163
Pattee HH (1968) The physical basis of coding and reliability in biological evolution. In: Waddington CH (ed) Towards a theoretical biology, vol 1. Edinburgh University Press, Edinburgh, pp 67–93
Pattee HH (1972) Laws and constraints, symbols and languages. In: Waddington CH (ed) Towards a theoretical biology, vol 4. Edinburgh University Press, Edinburgh, pp 248–258
Pattee HH (2007) Laws, constraints and the modeling relation—history and interpretations. Chem Biodivers 4:2272–2295 2007)
Pattee HH, Kull K (2009) A biosemiotic conversation: between physics and semiotics. Sign Syst Stud 37(1/2):311–331
Peirce CS (1931–1936) The collected papers, vol. 1–6. In: Hartshorne C, Weiss P (eds) Harvard University Press, Cambridge
Peirce CS (1998) The essential Peirce, vol. 2. Peirce Edition Project. Indiana University Press, Bloomington
Posner R, Robering K, Sebeok TA (1997) Semiotik/semiotics: a handbook on the sign-theoretical foundations of nature and culture, vol 1. Walter de Gruyter, Berlin
Rashevsky N (1954) Topology and life: in search of general mathematical principles in biology and sociology. Bull Math Biophys 16:317–348
Rosen R (1991) Life itself. A comprehensive enquiry into the nature, origin and fabrication of life. Columbia University Press, New York
Rosen R (2006) Autobiographical reminiscences of Robert Rosen. Axiomathes 16(1–2):1–23. https://doi.org/10.1007/s10516-006-0001-6
Rosen R (2012) Anticipatory systems. Philosophical, mathematical, and methodological foundations, 2nd edn. Springer, New York. [1st edn, 1985, Pergamon Press, Ontario]
Sebeok TA (1963) Communication among social bees; porpoises and sonar; man and dolphin. Language 39:448–466
Sebeok TA (1972) Perspectives in zoosemiotics. Mouton, The Hague
Sebeok TA (2001) Biosemiotics: its roots, proliferation, and prospects. In: Kull K (ed) Jakob von Uexküll: a paradigm for biology and semiotics. Semiotica 134(1/4):61–78
Uexküll J von (1928) Theoretische biologie, 2nd edn. Julius Springer, Berlin
Varela FJ (1979) Principles of biological autonomy. Elsevier, New York
Acknowledgments
I thank Cristian Saborido and two anonymous reviewers for their valuable comments on previous versions of this article. Deborah Klosky deserves special thanks for her editorial assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
The original version of this article was corrected: In the ‘Barbieri’s Concept of Mechanisms’ section on page 12 of above mentioned article the text erroneously reads Rosen, quoting Alberts et al. (2007), highlights…’. The correct text should read ‘Barbieri, quoting Alberts et al. (2007), highlights…’.
Rights and permissions
About this article

Cite this article
Vega, F. A Critique of Barbieri’s Code Biology Through Rosen’s Relational Biology: Reconciling Barbieri’s Biosemiotics with Peircean Biosemiotics. Biol Theory 13, 261–279 (2018). https://doi.org/10.1007/s13752-018-0302-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13752-018-0302-1