
Abstract
The intense drought affecting olive production in Northern Chile underscores the need to research non-traditional irrigation strategies to obtain the best crop performance. Accordingly, this study aimed to obtain preliminary data to guide future research on this topic. Different water replenishment levels on crop evapotranspiration (ETc; 13.5, 27.0, 40.5, and 54%) were established in a young orchard, cv. Arbequina, from the end of fruit drop (EFD) to full bloom in the next season. We evaluated the influence of plant water status (Ψstem) and crop load, considered as function of fruit number divided by trunk cross-sectional area, on reproductive and productive variables using multiple linear regressions. Our results show that crop load and Ψstem measured from EFD to harvest affected yield components. Nevertheless, Ψstem had the strongest influence on fruit size, pulp development, oil accumulation, and yield. Oil content and yield were reduced by 54% and 50% for each MPa, respectively, from Ψstem EFD-H − 1.8 MPa, an effect that intensified as crop load increased. During the period of flower development (September–November), the number of flowers per inflorescence and percentage of perfect flowers were reduced when Ψstem was less than − 2.0 MPa. These preliminary results showed that bud differentiation, inflorescence and flower formation are highly sensitive to water deficit.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Chile is an emerging competitor in olive oil production. One of its main olive growing areas is the Coquimbo region, with 21% of the country’s planted area. The region has an arid climate, with a high accumulation of degree-days (1600–2100; base 10 °C), is frost-free, and has an annual rainfall of 75–100 mm, mainly concentrated in winter (Uribe et al. 2012). Under these conditions, and with irrigation doses of about 8000 m3 ha−1, yield potential in full production is between 12 and 15 t ha−1 (Tous et al. 2014). However, during the last 8 years, limited irrigation has been a major issue for olive farmers. This is caused by a consistent drop in water availability in irrigation dams and groundwater, resulting from a progressive decrease in precipitation. A reduction between 20 and 30% towards the middle of the 21st century is estimated, possibly as a response to global warming and a greater competition for water from other sectors (Santibáñez et al. 2014). Thus, the prolonged and intense drought affecting the Coquimbo region makes it impossible to satisfy fruit tree water demand. The productive potential is reduced under these conditions, making it necessary to develop irrigation management strategies under conditions of limited irrigation water availability.
Variables used to assess the efficiency of different irrigation strategies include irrigation water use efficiency (IWUE), expressed as kg of fresh fruit or kg of oil per mm of water supplied per hectare (Trentacoste et al. 2015). This information is essential in areas where water is scarce and expensive, such as the Coquimbo region, where the production-related cost/benefit ratio must be evaluated. Several studies have shown a curvilinear olive yield response to the amount of water irrigation supply (Moriana et al. 2003; d’Andria et al. 2004; Berenguer et al. 2006; Grattan et al. 2006; d’Andria 2008), where maximum oil yield was achieved when water applied was close to 60–75% of the crop evapotranspiration, and a higher IWUE was reached with low levels of applied water.
Moriana et al. (2012) reported that measurements of plant water status enable a good approximation of irrigation needs, generating water potential thresholds within which a good yield can be reached, regardless of the location. These authors indicated that water potential must be kept over − 1.2 MPa and − 1.4 MPa before and after pit hardening, respectively, to maintain olive trees in non-stress condition. Likewise, Trentacoste et al. (2015) found that, regardless of tree density and climate conditions, the non-stress threshold in cv. Frantoio was − 1.3 MPa and − 1.5 MPa before and after pit hardening, respectively. By contrast, Naor et al. (2013) described this threshold as dependent on the crop load. These authors observed that oil yield in cv. Koroneiki was not affected by levels of water stress applied during the period of oil accumulation (− 1.5 MPa to − 4 MPa) when the crop load was low. However, with high crop loads, there was greater oil yield dependence on water stress levels, associated with a high demand of assimilates. Thus, olive yield response to water status is related to intensity, duration and moment of stress (Moriana et al. 2012), and their interaction with crop load (Martín-Vertedor et al. 2011; Naor et al. 2013).
Although olive trees respond positively to a lower percentage of ETc replenishment, water stress during critical periods, such as the flowering process, affects yield (Rapoport et al. 2012; Pierantozzi et al. 2013) mainly by a reduction of crop load (Ben-Gal et al. 2011). Under Mediterranean climatic conditions, where olive trees are traditionally grown, rains in winter and early spring frequently provide sufficient water to the soil for optimal development in the differentiation process, flower development, and shoot growth. Therefore, the effect of reduced water availability during this period of great sensitivity to water stress has been of little interest (Connor and Fereres 2005; Rapoport et al. 2012; Pierantozzi et al. 2013). In addition, most studies focused on different levels of reduction in irrigation supply between specific phenological periods, and establish non-stress conditions before and after specific study periods (Gómez-del-Campo 2013a, b; Naor et al. 2013; Pierantozzi et al. 2014). Nevertheless, under severe drought conditions water availability is reduced for a longer time, with the distribution of scarce water in key moments becoming more important to ensure a minimum production and preserve the orchard life.
In this context, this study aimed to develop a preliminary evaluation on irrigation management strategies for olive crop under limited irrigation water availability, by (1) exploring the influence of plant water status from the end of fruit drop to the next season’s full bloom, considering crop load on plant production variables; and (2) comparing the yield and irrigation water use efficiency for oil production using low ETc replenishment levels in a young orchard of ‘Arbequina’ olive trees.
2 Materials and methods
2.1 Site and orchard characteristics
The study was carried out from December 2012 to November 2013 in a commercial 4-year-old ‘Arbequina’ olive orchard planted in 2008, belonging to Valle Arriba S.A. and located at Tabalí, 15 km from Ovalle in the Limarí province, Coquimbo region, Chile (30°39′S, 71°25′W). The experimental plot is part of the Tabalí soil series, characterized by alluvial terraces, varying from flat to rolling lands (IREN 1964). The clayey soil (ISSS classification) contains 52% clay, 30.5% sand, and 17.5% silt, a bulk density of 1.1 Mg m−3, with gravimetric water content at field capacity of 32.8% (− 0.03 MPa) and 22% at the permanent wilting point (− 1.5 MPa). The average root depth observed was 0.65 m. The olive trees were planted in an east–west orientation on hedgerows with a 6 × 3 m spacing. Their harvest in the previous season was 3500 kg ha−1.
2.2 Irrigation treatments
Prior to this study, the orchard was fully irrigated. From the last week of December 2012 (end of fruit drop) to November 2013 (full bloom), different irrigation treatments were carried out based on crop evapotranspiration (ETc) replenishment levels of 13.5, 27.0, 40.5, and 54.0%. The ETc was calculated with Eq. (1), using reference evapotranspiration (ETo) calculated by the Penman–Monteith method (Allen et al. 1998), and a crop coefficient (Kc) of 0.7 (Girona et al. 2002), which was adjusted by a factor dependent on the coverage (Kr) (Fereres et al. 1981; 2012). The Kr was estimated as the average daily fraction of the intercepted photosynthetically active radiation (fPARi), described below, obtaining an average intercepted fraction of 0.35 for all trees. This value was used throughout the study.
Irrigation was supplied with a double-irrigation dripper line with emitters spaced 1 m apart (6 emitters per tree) delivering 4, 3, 2, and 1 l h−1 (depending on the treatment). Irrigation scheduling was the same for all treatments and determined based on orchard water availability. Thus, from mid-December (beginning of treatments) until early March, weekly irrigations were carried out. Later, irrigations were scheduled every 15 days (Fig. 2). Total irrigation from the beginning of the treatment until harvest was 74.6, 56, 37.3 and 18.65 mm for the treatments of 54, 40.5, 27 and 13.5% ETc, respectively. Finally, in October 2013 a single irrigation was applied (Fig. 2). The irrigation time was the same for all treatments.
2.3 Meteorological data
During the study, maximum and minimum temperatures, relative humidity, precipitation, solar radiation, vapor pressure deficit, and wind speed were recorded through a weather station (Campbell Sci; Utah, USA) located 1 km from the study site.
2.4 Fraction of intercepted solar radiation (fPAR i)
The fPARi was calculated (Eq. 2) by measuring the radiation not intercepted by the trees (PARni) along with the incident radiation on the orchard (PARo) on a completely sunny day at the beginning of the study. The PAR measurements were taken with a SunScan SS1 ceptometer (Delta-T Devices Ltd., Cambridge, UK) in the area assigned under the canopy of the tree in the center of each plot. Measurements were taken every 0.5 m cross-sectionally to the row (42 measurements per tree), capturing the shaded and non-shaded areas on the entire surface of the soil assigned to the tree (PARni). They were taken five times during the day: at solar noon, two, and four hours before and after solar noon. Immediately before taking the measurements under the canopy, the incident PAR (PARo) was measured in a non-shaded area without canopy interference, at a height of 1.5 m.
2.5 Plant water status
Stem water potential was measured at pre-dawn (Ψpd) and midday (Ψstem) as indicators of plant water status (Moriana et al. 2012). In both measurements, one small branch per tree was selected from the central part of the canopy under direct sun exposure. To measure Ψstem, branches were covered with bags of aluminum sheets wrapped in plastic at least 1 h before taking the measurements to allow the leaf water potential to balance with the stem water potential (Trentacoste et al. 2015). Ψpd and Ψstem were measured between 5:30–6:00 a.m. and 12:30–1:30 p.m., respectively, once a week until February 2013, then once a month. Measurements were taken according to the procedure described by Guerfel et al. (2009). A Scholander pressure chamber (PMS Model 600, USA) was used for the measurement.
2.6 Yield and yield components
Olives were harvested on May 6, 2013. From a 2-kg sample per tree, 100 fruits were weighed to determine their average weight (wt). Total fruit number per tree was estimated from the average fruit weight and total harvest weight. Fruit yield and crop load efficiency per tree were expressed dividing the total harvest weight and fruit number, respectively, by the trunk cross-sectional area (TCSA) measured 30 cm above the soil at harvest, to account for possible differences in tree size. Oil concentration in the pulp was determined by the Soxhlet method (AOAC 2000) with a sample of 25 fruits per tree. The pulp was dried in an atmospheric pressure oven at 70 °C, until achieving a constant weight. The result was expressed in percentage of oil pulp based on dry matter. Percent water concentration was calculated as 100 × (fresh wt − dry wt)/fresh wt. Oil yield efficiency at harvest (kg oil cm−2 of TCSA) was calculated per tree according to the following equation:
where OCM is the pulp oil concentration on a dry-weight basis, DW/FW is the dry-to-fresh-weight ratio, M/F is the fresh mesocarp/fruit ratio, and P is the fruit yield efficiency in kg cm−2 TCSA per tree. Irrigation water use efficiency for oil yield (IWUEo) was estimated as the kg of oil per mm of irrigation per hectare applied from the beginning of the treatments to harvest.
2.7 Shoot growth and flower development
Shoot growth was measured monthly in three fruitless shoots per tree, from the beginning of treatments (end of December 2012) until harvest (first week of May 2013). The number of flowers per inflorescence and the percentage of perfect flowers were determined in full bloom (November 1, 2013), when 70% of the inflorescences showed at least 50% open flowers (Sanz-Cortés et al. 2002). To count the number of flowers per inflorescence, a sample of 20 inflorescences per tree (2 inflorescences × 10 branches) was selected from the middle third of each branch located at the center of each side of the hedgerow. Then, the percentage of perfect flowers present in each inflorescence was determined. A flower was considered perfect when it presented both functional androecium and gynoecium, and staminate when it only had stamens or showed an atrophied gynoecium (without development).
2.8 Experimental design and statistics
Homogeneous trees were selected in full bloom based on flowering, vegetative development (coefficient of variation for TCSA and fPARi of 13% and 21%, respectively), vigor, height (~ 2.2 m), and health (no visible nutritional or pathological problems). However, high variability in crop load was observed at harvest. Therefore, crop load was expressed as fruit number per cm2 of TCSA to normalize for tree size (Bustan et al. 2016). A randomized complete block design was used, considering the row as a block (three blocks). The experimental plot of each treatment consisted of three trees, with the central plant used as the experimental unit, and two other trees used as borders between treatments, to avoid any influence between neighboring treatments.
Simple and multiple regression models were adjusted for each variable evaluated as a function of average Ψstem between phenological periods, which were determined according to Sanz-Cortés et al. (2002), and crop load. Phenological periods were: end of fruit drop to pit hardening (EFD-PH), end of fruit drop to harvest (EFD-H), pit hardening to harvest (PH-H), harvest to budbreak (H-B), and budbreak to full bloom (B-FB). A t test was used for individual coefficients on the regression parameters estimated with a 5% level of significance. For model selection, both the Akaike (AIC) and the Bayesian (BIC) information criteria were assessed (Yang 2005). Mallows’s Cp-statistic was used as an indicator of the contribution of the regressor variables in the fit regression models (Balzarini et al. 2008). To visualize the association between regressor variables, the relationship between the response and the other regressors was removed, then a partial residual analysis was performed with scatter plots (Draper and Smith 1998). The statistical program used was InfoStat v. 2013 (Di Rienzo et al. 2013).
3 Results and discussion
3.1 Seasonal conditions
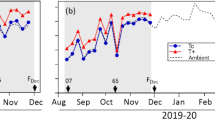
The average maximum temperature from the beginning of the study until harvest was 24.5 °C, and the average vapor pressure deficit (VPD) was 1 kPa, with a mid-summer maximum of 1.87 kPa (Fig. 1). During the spring of 2013, the average maximum temperature was 20.6 °C (Fig. 1). Over the study period, there were no temperatures below 2.8 °C (data not shown). Maximum reference evapotranspiration (ET0) in the summer was 7.6 mm day−1. There was no precipitation during the summer, whereas in winter (end of May 2013) there were two important consecutive precipitation events of 39 and 42 mm day−1 (Fig. 2), leading experimental plots to field capacity and olive trees with optimal water status (Fig. 2).

a Seasonal daily maximum air temperature and vapor pressure deficit (VPD). b Seasonal daily reference ET0 from end of fruit drop to full bloom of next season in Ovalle, 2012–2013. Bloom occurs at mid-November; oil accumulation starts at the end of January, and harvest occurs in mid-May

Irrigation amount (a) and midday stem water potential (b) in the irrigation levels (% of reference ETc) from end of fruit drop to full bloom of next season. Diamond symbols indicate daily irrigation. PH pit hardening, H harvest, B budbreak, FB full bloom. Each data point represents a mean (n = 3)
3.2 Plant water status
Pre-dawn leaf water potential (Ψpd) for different phenological stages is detailed in Table 1. The Ψpd at the beginning of the study was similar among irrigation treatments, varying between − 0.62 and − 0.85 MPa. Pre-dawn values during the first eight weeks after bloom should be over − 2.0 MPa and − 3.0 MPa according to Costagli et al. (2003) and Rapoport et al. (2004), respectively, for optimal early fruit growth without significant reduction in cell number or cell size. In this study, average midday Ψstem among treatments was − 2.11 MPa (Fig. 2), consequently olive trees were already stressed according to the ranges proposed previously (Moriana et al. 2012; Trentacoste et al. 2015). Before pit hardening, similar pre-dawn and midday water potential among treatments was largely due to the high capacity of the olive tree to extract water, even under water stress conditions (Sofo et al. 2007). During pit hardening, both soil and plant water status were significantly different among treatments (Table 1 and Fig. 2). The 13.5% ETc treatment showed the lowest Ψpd (− 1.52 MPa), whereas the other irrigation treatments increased pre-dawn leaf water potential when irrigation contribution increased (Table 1). At harvest, the trees showed increased stress levels, which were greater in treatments with 40.5, 27, and 13.5% ETc replenishment (average Ψstem − 3.8 MPa) compared to the 54% ETc (Ψstem − 3.0 MPa). It should be noted that the two lowest irrigation rates were similar in terms of Ψstem over the study period; therefore, plant water status was similar between the 27% ETc and 13% ETc treatments.
Winter rains improved soil water status, rehydrating the olive trees (Fig. 2). However, at the onset of budbreak the Ψpd was between − 1.9 and − 2.22 MPa (Table 1), and stem water potential was lower than − 2.0 MPa, with non-significant differences among treatments. Finally, during the period of flower development until bloom, Ψstem varied around − 2.25 MPa at the highest replenishment irrigation rate, whereas the other treatments reached an average Ψstem of − 3.1 MPa.
3.3 Oil yield and yield component responses
As previously reported, crop load has a strong influence on water tree requirement (Bustan et al. 2016), shoot growth (Dag et al. 2010), yield and yield components, i.e., oil content and fruit size (Trentacoste et al. 2010). In this context, and given the variability of the crop load on the evaluated trees (variation coefficient of 33.1% Table 3), multiple linear regressions were adjusted, accounting for the crop load and water status of olive trees (Table 2). A slightly better fit (5%) in the adjusted models was found, considering the water potential of EFD-H vs. PH-H period (data not shown). Although there was a better fit considering Ψstem EFD-H, i.e., when different irrigation regimens began, a greater contribution of water status to the yield and its components was observed after pit hardening. It has been widely reported that the least responsive period to water stress in olive is the EFD-PH period (Goldhamer 1999; Alegre et al. 2002; Lavee et al. 2007; Gómez-del-Campo 2013a, b). Indeed, Gómez-del-Campo (2013a, b) found in olive hedgerows of cv. Arbequina that water can be saved in this period until a Ψstem threshold of -2.9 MPa without any yield penalty. In the present study, Ψstem EFD-PH ranged from − 1.7 to − 2.5 MPa (Table 3), which was higher than the Ψstem threshold indicated by Gómez-del-Campo (2013a, b). Although stress intensity is relevant, stress duration also plays an important role (Moriana et al. 2012); therefore, it is likely that this slightly better fit found throughout the period is due to stress duration.
The multiple linear regression analyses showed that plant water status, measured as Ψstem EFD-H, was the variable that most negatively affected fruit size, pulp development (pulp/fruit ratio), oil accumulation (oil concentration and content in the pulp), and yield (higher Mallows’s Cp value, Table 2). In turn, linear regression coefficients (slope) related to water potential showed a significantly higher impact on oil content and oil yield (kg of oil cm−2 TCSA), which decreased from Ψstem EFD-H − 1.8 MP by 54% and 50% for each MPa, respectively (Tables 2, 3). The partial residuals of all multiple regressions showed that yield and its components were correlated linearly to crop load and water potential in the ranges of both variables (Fig. 3); therefore, it was not necessary to fit more complex models (e.g. polynomials). Although crop load (range from 89 to 305 fruits cm−2 of TCSA, Table 3) also influenced the reduction in productive traits due to deficit irrigation, its contribution was smaller (lower Mallows’s Cp value, Table 2). This may be because (1) severe water stress affects mesocarp weight, in both cell size and number, whereas crop load mainly affects mesocarp weight through cell number (Gucci et al. 2009; Lodolini et al. 2011); and (2) photoassimilate synthesis and partitioning were more limited by water stress rather than by competition among sinks. In this scenario, there is a greater dependency on irrigation to achieve higher yields, despite olive trees being young and with high levels of crop load. This agrees with Martín-Vertedor et al. (2011) and Naor et al. (2013), who indicate that the impact of water stress on oil yield and quality increases with crop load.

Partial residuals for oil content as a function of a crop load (fruit/trunk cross-sectional area) and b average midday water potential from end of fruit drop (EFD) to harvest (H), corresponding to a multiple linear regression model (n = 12)
Previous studies have indicated that reductions in irrigation rates during the summer can improve fruit water concentration with a consequent higher oil extractability, without a significant effect on oil yield (Goldhamer 1999; Alegre et al. 2002). In contrast, Girona et al. (2002) and Grattan et al. (2006) observed a lower oil production with summer irrigation deficits. In our study, we found that fruit water concentration was strongly correlated to water potential (R2 = 0.76), decreasing by 12% for each average MPa reduction from pit hardening to harvest (Table 2). Furthermore, crop load had a non-significant effect, as previously reported (Trentacoste et al. 2010). Although fruit water concentration decreased as water stress increased, both oil concentration and content were affected, decreasing 21% and 0.13 g fruit−1 for each Ψstem EFD-H MPa, respectively (Table 2). The Ψstem range of olive trees varied between − 1.9 and − 3.4 MPa in PH-H, i.e. a moderate stress according to Moriana et al. (2002). The slopes of these correlations could be important for designing irrigation strategies for greater productivity.
In this context, an irrigation threshold of Ψstem EFD-H greater than − 2.0 MPa must be maintained to avoid affecting yield components, in agreement with Gómez-del-Campo (2013a, b), although this threshold also depends on the crop load (Naor et al. 2013). However, reducing fruit water concentration requires values of Ψstem PH-H less than − 2.0 MPa. Puertas et al. (2012) found that a Ψstem PH-H of − 1.65 MPa did not affect fruit water concentration, yield components or extractability, but allowed for a 16% water saving. Further research is needed to determine the moment and degree of pre-harvest stress and when water concentration could be reduced, to improve extractability without affecting oil yield.
3.4 Irrigation water use efficiency
Oil yield was correlated to irrigation rates through a polynomial quadratic curve (Fig. 4). The highest level of ETc replenishment (54% ETc), generated the greatest yield, which was practically twice that of the other treatments. A similar response was found by Lodolini et al. (2016), who reported that a positive effect on yield could only be reached when irrigation supply was over 35% ETc. For the lowest irrigation rates, no reduction in oil yield was observed. Irrigation water use efficiency (IWUEo) for oil yield had the inverse response, i.e., the lowest irrigation rate was the most efficient, showing twice the efficiency of the other irrigation levels, whereas no distinction was seen at the highest irrigation rate (Fig. 4b). On the other hand, the multiple linear regression analysis showed that IWUEo was linearly and positively correlated to the crop load and the degree of stress increased (Ψstem EFD-H), with both variables contributing similarly to IWUEo (Table 2). This would explain why the three lowest irrigation rates did not show any differences in yield among them, due to highly efficient water use of the trees under scarce availability (Trentacoste et al. 2015). In addition, these relations underscore the importance of increased irrigation rates when the crop load is greater, as IWUEo is positively associated with fruit number. However, it is necessary to assess a strategy of severe, long term water restriction. Although IWUEo increases, it may affect the olive tree reserves, and therefore, reduce the productive life of the orchard (Bustan et al. 2011).

a Oil yield and b irrigation water use efficiency for oil yield (IWUEo) as a function of olive tree evapotranspiration (n = 12)
3.5 Shoot growth and inflorescence response
In the Coquimbo region, most of the shoot growth occurs from the beginning of the season in August until January (Fichet and González 2011), which is why severe water stress is not desirable during this period (Naor et al. 2013). The soil water content in these months, from winter and spring rains (65 mm August 2012; data not shown), in addition to the increased water availability for irrigation due to snowmelt from the Andes, was sufficient for optimal shoot growth (Fig. 5). It has been shown that, depending on the stress severity, regulated deficit irrigation (RDI) during the summer could severely affect bud induction, when photosynthesis is affected (Alegre et al. 2001). However, in the present study we observed that, regardless of the treatment, more than 90% of the buds that developed in the 2012 spring–summer were floral buds in 2013 (Fig. 5). This occurred despite the fact that the Ψstem in the induction period (mid-January to the end of February) fluctuated between − 2.0 and − 3.0 MPa (Fig. 2). It is worth noting that, although shoot length was negatively affected by Ψstem EFD-H and crop load (Table 2), the maximum magnitude of this growth was only 4.2 cm (Table 3), which is why the effect of the irrigation levels on potential flower sites in the next season was minimal. This result may be because shoot growth rate in this period is low (Fichet and González 2011). Secondly, less shoot growth can be expected under Ψstem less than − 2.0 MPa as observed during this period, according to Ψstem thresholds reported in previous studies (− 1.8 MPa, Gómez-del-Campo et al. 2008; − 2.0 MPa, Moriana et al. 2012; − 1.3 MPa, Gómez-del-Campo 2013a, b).

Representative samples of flowering shoots at bloom in ‘Arbequina’ olive trees irrigated with different water levels (% ETc). Flowering shoots of 13.5% ETc and 27% ETc treatments showed 90% inflorescences per node (white circles); however, they did not develop because of water restriction from budbreak to full bloom (Ψstem < −2.5 MPa)
Additionally, according to Rapoport et al. (2012), water restriction during flower development (elongation and branching of the inflorescence axis, and formation of the individual flower) significantly reduced inflorescence structure and quality (i.e. flowers per inflorescence, number and proportion of perfect flowers, and ovary quality). Similarly, it was observed that the average Ψstem from budbreak to full bloom (Ψstem B-FB) had a strong impact on the percentage of perfect flowers (R2 = 0.80; p < 0.0001) and on flowers per inflorescence (R2 = 0.72; p < 0.0001), where flowers per inflorescence decreased linearly from 20 to 0 as Ψstem B-FB decreased from − 2.0 MPa to − 3.0 MPa (Fig. 6a, b). Also, Pierantozzi et al. (2014) observed that irrigation with 75% ETc and average Ψstem greater than − 2 MPa during flower development did not affect either flower number per inflorescence or the yield in a year with heavy flowering. In contrast, in a year with low flowering, trees that presented a water potential lower than − 3.0 MPa beared similar fruit loads to those that presented water potentials above − 1.5 MPa, although shoot growth did decrease with low Ψstem. Therefore, irrigation thresholds between − 1.3 and − 1.5 MPa (as a safety factor) can be used during the spring without affecting reproductive variables, but partially reducing shoot growth, which could be used to control vigor in super intensive orchards (Connor and Gómez-del-Campo 2013).

a Flowers and b percentage of perfect flowers per inflorescence as a function of average midday stem water potential from budbreak (B) to full bloom (FB), inflorescence development period (n = 12). *indicates a significant linear relationship at p ≤ 0.05
4 Concluding remarks
Under water stress conditions, both vegetative and productive development of ‘Arbequina’ olive trees were more affected by water status than crop load, from the end of fruit drop to harvest. Nevertheless, the effect of water status was intensified as fruit number increased. In addition, the flower development period was highly sensitive to water stress. During this period, a midday stem water potential threshold above − 2.0 MPa should be maintained, to avoid affecting the number of flowers per inflorescence and the percentage of perfect flowers. The results presented here are preliminary, and it is necessary to continue exploring the irrigation thresholds that must be applied in conditions of low water availability without compromising the productive life span of olive orchards and maintaining yield over time.
References
Alegre S, Girona J, Arbonés A, Mata M, Marsal J (2001) Estrategias de riego deficitario controlado para el riego del olivar. Especial Olivicultura III, Fruticultura Profesional 120:19–28
Alegre S, Marsal J, Mata M, Arbonés A, Girona J, Tovar MJ (2002) Regulated deficit irrigation in olive trees (Olea europaea L. cv. Arbequina) for oil production. Acta Hortic 586:259–262. https://doi.org/10.17660/ActaHortic.2002.586.49
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration (guidelines for computing crop water requirements), Irrigation and drainage no.56
AOAC (2000) Official methods of analysis of AOAC international. association of official analysis chemists international. https://doi.org/10.3109/15563657608988149
Balzarini M, Gonzalez L, Tablada M, Casanoves F, Di Rienzo J, Robledo C (2008) InfoStat. Manual de usuario. Editorial Brujas, Córdoba
Ben-Gal A, Yermiyahu U, Zipori I, Presnov E, Hanoch E, Dag A (2011) The influence of bearing cycles on olive oil production response to irrigation. Irrig Sci 29:253–263. https://doi.org/10.1007/s00271-010-0237-1
Berenguer MJ, Vossen P, Grattan S, Connell J, Polito V (2006) Tree irrigation levels for optimum chemical and sensory properties of olive oil. HortScience 41:427–432
Bustan A, Avni A, Lavee S, Zipori I, Yeselson Y, Schaffer A, Riov J, Dag A (2011) Role of carbohydrate reserves in yield production of intensively cultivated oil olive (Olea europaea L.) trees. Tree Physiol 31:519–530. https://doi.org/10.1093/treephys/tpr036
Bustan A, Dag A, Yermiyahu U, Erel R, Presnov E, Agam N, Kool D, Iwema J, Zipori I, Ben-Gal A (2016) Fruit load governs transpiration of olive trees. Tree Physiol 36:380–391. https://doi.org/10.1093/treephys/tpv138
Connor D, Fereres E (2005) The physiology of adaptation and yield expression in olive. Hortic. Rev. 34:155–229
Connor D, Gómez-del-Campo M (2013) Simulation of oil productivity and quality of N-S oriented olive hedgerow orchards in response to structure and interception of radiation. Sci Hortic 150:92–99. https://doi.org/10.1016/j.scienta.2012.09.032
Costagli G, Gucci R, Rapoport HF (2003) Growth and development of fruits of olive “Frantoio” under irrigated and rainfed conditions. J. Hortic. Sci. Biotechnol. 78:119–124
d’Andria R (2008) Olive responses to different irrigation management in the Mediterranean environment. In: IV. Jornadas de Actualizacion En Riego Y Fertirriego, Mendoza
d’Andria R, Lavini A, Morelli G, Patumi M, Terenziani S, Calandrelli D, Fragnito F, Alta VM (2004) Effect of water regimes on five pickling and double aptitude olive cultivars (Olea europaea L). J Hortic Sci Biotechnol 79:18–25. https://doi.org/10.1080/14620316.2004.11511731
Dag A, Bustan A, Avni A, Tzipori I, Lavee S, Riov J (2010) Timing of fruit removal affects concurrent vegetative growth and subsequent return bloom and yield in olive (Olea europaea L.). Sci Hortic 123:469–472. https://doi.org/10.1016/j.scienta.2009.11.014
Di Rienzo J, Casanoves F, Balzarini M, Gonzalez L, Tablada M, Robledo C (2013) Infostat - Sofware estadístico. Universidad Nacional de Córdoba, Argentina, Universidad Nacional de Córdoba, Argentina
Draper NR, Smith H (1998) Applied regression analysis, 3rd edn. Wiley, New York. https://doi.org/10.1002/9781118625590
Fereres E, Pruitt W, Beutel J, Henderson DW, Holzapfel E, Schelbach H, Uriu K (1981) Evapotranspiration and drip irrigation scheduling. In: Fereres E (ed) Drip irrigation management. Division of Agricultural Sciences, University of California, Oakland, pp 8–13
Fereres E, Goldhamer D, Sadras V (2012) Yield response to water of fruit trees and vines: guidelines. In: Steduto P, Hsiao T, Fereres E, Raes D (eds) Crop yield response to water, vol 66. Irrigation and drainage paper. FAO, Roma, pp 246–295
Fichet T, González C (2011) Comportamiento fenológico del olivo en la Región de Atacama. In: Fichet T, Razeto B, Curcovik T (eds) El Olivo: Estudio Agronómico En La Región de Atacama. Facultad de Ciencias Agronómicas, Universidad de Chile, Santiago, pp 13–38
Girona J, Luna M, Arbonés A, Mata M, Rufat J, Marsal J (2002) Young olive trees responses (Olea europaea, cv “Arbequina”) to different water supplies. Water function determination. Acta Hortic 586:277–280. https://doi.org/10.17660/ActaHortic.2002.586.53
Goldhamer D (1999) Regulated deficit irrigation for California canning olives. Acta Hortic 474:369–372. https://doi.org/10.17660/ActaHortic.1999.474.76
Gómez-del-Campo M (2013a) Summer deficit irrigation in a hedgerow olive orchard cv. Arbequina: relationship between soil and tree water status, and growth and yield components. Span J Agric Res 11:547–557. https://doi.org/10.5424/sjar/2013112-3360
Gómez-del-Campo M (2013b) Summer deficit-irrigation strategies in a hedgerow olive orchard cv. “Arbequina”: effect on fruit characteristics and yield. Irrig Sci 31:259–269. https://doi.org/10.1007/s00271-011-0299-8
Gómez-del-Campo M, Leal A, Pezuela C (2008) Relationship of stem water potential and leaf conductance to vegetative growth of young olive trees in a hedgerow orchard. Aust J Agric Res 59:270–279. https://doi.org/10.1071/AR07200
Grattan S, Berenguer MJ, Connell J, Polito V, Vossen P (2006) Olive oil production as influenced by different quantities of applied water. Agric Water Manag 85:133–140. https://doi.org/10.1016/j.agwat.2006.04.001
Gucci R, Lodolini EM, Rapoport HF (2009) Water deficit-induced changes in mesocarp cellular processes and the relationship between mesocarp and endocarp during olive fruit development. Tree Physiol 29:1575–1585. https://doi.org/10.1093/treephys/tpp086
Guerfel M, Baccouri O, Boujnah D, Chaïbi W, Zarrouk M (2009) Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci Hortic 119:257–263. https://doi.org/10.1016/j.scienta.2008.08.006
IREN (1964) Estudio de Suelos del Proyecto Aerofotogramétrico CHILE/OEA/BID. Instituto de Investigación de Recursos Naturales, Santiago
Lavee S, Hanoch E, Wodner M, Abramowitch H (2007) The effect of predetermined deficit irrigation on the performance of cv. Muhasan olives (Olea europaea L.) in the eastern coastal plain of Israel. Sci Hortic 112:156–163. https://doi.org/10.1016/j.scienta.2006.12.017
Lodolini EM, Gucci R, Rapoport HF (2011) Interaction of crop load and water status on growth of olive fruit tissues and mesocarp cells. Acta Hortic 924:89–94. https://doi.org/10.17660/ActaHortic.2011.924.10
Lodolini EM, Polverigiani S, Ali S, Mutawea M, Qutub M, Pierini F, Neri D (2016) Effect of complementary irrigation on yield components and alternate bearing of a traditional olive orchard in semi-arid conditions. Span J Agric Res 14:e1203. https://doi.org/10.5424/sjar/2016142-8834
Martín-Vertedor A, Pérez J, Prieto H, Fereres E (2011) Interactive responses to water deficits and crop load in olive (Olea europaea L., cv. Morisca). II: water use, fruit and oil yield. Agric Water Manag 98:950–958. https://doi.org/10.1016/j.agwat.2011.01.003
Moriana A, Villalobos F, Fereres E (2002) Stomatal and photosynthetic responses of olive (Olea europaea L.) leaves to water deficits. Plant, Cell Environ 25:395–405. https://doi.org/10.1046/j.0016-8025.2001.00822.x
Moriana A, Orgaz F, Pastor M, Fereres E (2003) Yield responses of a mature olive orchard to water deficits. J Am Soc Hortic Sci 128:425–431. https://doi.org/10.3389/fpls.2017.01280
Moriana A, Pérez-López D, Prieto MH, Ramírez-Santa-Pau M, Pérez-Rodriguez J (2012) Midday stem water potential as a useful tool for estimating irrigation requirements in olive trees. Agric Water Manag 112:43–54. https://doi.org/10.1016/j.agwat.2012.06.003
Naor A, Schneider D, Ben-Gal A, Zipori I, Dag A, Kerem Z, Birger R, Peres M, Gal Y (2013) The effects of crop load and irrigation rate in the oil accumulation stage on oil yield and water relations of “Koroneiki” olives. Irrig Sci 31:781–791. https://doi.org/10.1007/s00271-012-0363-z
Pierantozzi P, Torres M, Bodoira R, Maestri D (2013) Water relations, biochemical–physiological and yield responses of olive trees (Olea europaea L. cvs. Arbequina and Manzanilla) under drought stress during the pre-flowering and flowering period. Agric Water Manag 125:13–25. https://doi.org/10.1016/j.agwat.2013.04.003
Pierantozzi P, Torres M, Lavee S, Maestri D (2014) Vegetative and reproductive responses, oil yield and composition from olive trees (Olea europaea) under contrasting water availability during the dry winter-spring period in central Argentina. Ann Appl Biol 164:116–127. https://doi.org/10.1111/aab.12086
Puertas C, Trentacoste E, Morábito J, Perez J (2012) Effects of regulated deficit irrigation during stage III of fruit development on yield and oil quality of olive trees (Olea europaea L. “Arbequina”). Acta Hortic 889:303–310. https://doi.org/10.17660/ActaHortic.2011.889.36
Rapoport HF, Costagli G, Gucci R (2004) The effect of water deficit during early fruit development on olive fruit morphogenesis. J Am Soc Hortic Sci 129:121–127
Rapoport HF, Hammami S, Martins P, Pérez-Priego O, Orgaz F (2012) Influence of water deficits at different times during olive tree inflorescence and flower development. Environ Exp Bot 77:227–233. https://doi.org/10.1016/j.envexpbot.2011.11.021
Santibáñez F, Santibáñez P, Caroca C, Morales P, González P, Gajardo N, Perry P, Melillán C (2014) Atlas del Cambio Climático en las Zonas de Régimen Árido y Semiárido. Regiones de Coquimbo, Valparaíso y Metropolitana, Universidad de Chile, Santiago
Sanz-Cortés F, Martínez-Calvo J, Badenes M, Bleiholder H, Hack H, Llacer G, Meier U (2002) Phenological growth stages of olive trees (Olea europaea L.). Ann Appl Biol 140:151–157. https://doi.org/10.1111/j.1744-7348.2002.tb00167.x
Sofo A, Manfreda S, Dichio B, Fiorentino M, Xiloyannis C (2007) The olive tree: a paradigm for drought tolerance in Mediterranean climates. Hydrol Earth Syst Sci Discuss 4:2811–2835. https://doi.org/10.5194/hess-12-293-2008
Tous J, Romero A, Hermoso JF, Msallem M, Larbi A (2014) Olive orchard design and mechanization: present and future. Acta Hortic 1057:231–246. https://doi.org/10.17660/ActaHortic.2014.1057.27
Trentacoste ER, Sadras VO, Puertas CM (2010) Effect of fruit load on oil yield components and dynamics of fruit growth and oil accumulation in olive (Olea europaea L.). Eur J Agron 32:249–254. https://doi.org/10.1016/j.eja.2010.01.002
Trentacoste ER, Puertas CM, Sadras VO (2015) Effect of irrigation and tree density on vegetative growth, oil yield and water use efficiency in young olive orchard under arid conditions in Mendoza, Argentina. Irrig Sci 33:429–440. https://doi.org/10.1007/s00271-015-0479-z
Uribe J, de la Fuente A, Paneque M (2012) Atlas bio-climático de Chile. Universidad de Chile, Santiago
Yang Y (2005) Can the strengths of AIC and BIC be shared? A conflict between model indentification and regression estimation. Biometrika 92:937–950. https://doi.org/10.1093/biomet/92.4.937
Acknowledgements
We thank Ana María Espinoza and Sandra Benavente for revising the English language of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Beyá-Marshall, V., Herrera, J., Fichet, T. et al. The effect of water status on productive and flowering variables in young ‘Arbequina’ olive trees under limited irrigation water availability in a semiarid region of Chile. Hortic. Environ. Biotechnol. 59, 815–826 (2018). https://doi.org/10.1007/s13580-018-0088-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13580-018-0088-x