
Abstract
The parallel expansion of anthropogenic environments and wild boar Sus scrofa populations has favoured the intrusion of the species into many European metropolitan areas, including Rome. In this study, we used Maxent to analyse the seasonal spatial ecology of the wild boar in Marcigliana natural reserve, a rural area located in the northeastern suburbs of Rome. The wild boar avoided urban settlements year-round. In the growing season, it had a higher probability of occurring with an increasing presence of turkey oak Quercus cerrioides woods, but seemed to be partially adapted to the presence of arable lands, which had a marked negative effect only when very abundant. These results lead us to hypothesise that the species in this part of the year adopts a spatial strategy which optimises the trade-off among the need for thermal cover and food resources. In autumn and winter, the species avoided meadows and pastures. The analysis of wild boar spatial ecology in metropolitan areas is essential to provide important information contributing to the development of effective plans for managing peri-urban populations and mitigating conflicts with humans.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The wild boar (Sus scrofa) is among the most widely distributed large mammals in the world. The natural range of the species extends from Western Europe and the Mediterranean basin to Eastern Russia, Japan and Southeastern Asia, while the introduced one includes America, Africa and Oceania (Barrios-García and Ballari 2012). In Europe, the species has recently recolonised the UK, Sweden, Finland, Estonia and Norway, and populations are growing across the entire continent (Massei et al. 2015). Different reasons may explain this trend in wild boar numbers: a very high prolificacy, a great adaptability to a wide variety of environmental conditions, the lack of large predators in most of its range, reforestation processes following the abandonment of rural areas and massive releases of individuals for hunting purposes (Apollonio et al. 2010; Massei et al. 2015; Morelle et al. 2015).
Previously confined to natural areas with low human presence, in recent decades, the wild boar has colonised urban and peri-urban environments, establishing a permanent presence in many European cities (Cahill et al. 2012; Podgórski et al. 2013; Stillfried et al. 2017a). The intrusion of the wild boar in these areas is favoured by an easy access to abundant food sources, a low hunting pressure and an expanding urbanisation into the countryside (Licoppe et al. 2013), with rivers and roads acting as main movement corridors (Stillfried et al. 2017a; Castillo-Contreras et al. 2018). This phenomenon has inevitably exacerbated boar-human conflicts, specifically collisions with vehicles, disturbance or threat to residents, spread of diseases, damage to gardens and public parks and ransacking of rubbish bins and containers (Licoppe et al. 2013; Zuberogoitia et al. 2014; Fernández-Aguilar et al. 2018).
Over the last decade, the wild boar has become quite common also within the municipality of Rome (central Italy), which is characterised by the presence of densely built-up areas interspersed with several green areas, representing relicts of a traditional landscape composed of a mosaic of agricultural, grazing and woodland patches. Currently, the wild boar has been observed predominantly in peri-urban areas, farmlands and rural forests (Pierucci et al. 2014, 2015; Monterosso et al. 2016), but individuals have been seen even close to urban settlements and in densely populated areas (Primi et al. 2016), thus worsening boar-human conflicts.
Despite the amplified interest in wild boar research, there remains a significant knowledge gap regarding its basic biology and ecology; such data are essential for the development of effective, long-term management strategies (Beasley et al. 2018). With respect to urban populations, wildlife managers should consider the urban ecosystem as a whole, including both densely built-up areas and their surroundings. It has been demonstrated, in fact, that wild boars invading urban areas are mostly rural disperders, with cities serving as attractive sinks (Stillfried et al. 2017a; Castillo-Contreras et al. 2018). Therefore, to define appropriate management solutions able to limit boar-human conflicts and the invasion of urban settlements, it is overriding to understand the spatial ecology of the wild boar even in potential source areas (i.e. rural areas) (Pepin et al. 2017; Stillfried et al. 2017a; Beasley et al. 2018), targeting efforts also on seasonal habitat analyses; actually, the wild boar is able to cope with fluctuating conditions by shifting its spatial distribution to follow seasonal changes in cover and food availability (Keuling et al. 2009; Thurfjell et al. 2009; Morelle and Lejeune 2015).
In this sense, species distribution models can be a helpful planning tool, as they describe empirical correlations between ecological conditions and corresponding species distributions, allowing to identify the areas where the probability of occurrence of the target species is highest and consequently to highlight priority areas where to intervene (Franklin and Miller 2009; Gormley et al. 2011). In this study, we analysed the spatial ecology of the wild boar in Marcigliana natural reserve (hereafter, MNR), a rural area located in the northeastern suburbs of Rome. In particular, we used the maximum entropy modelling approach (Maxent) (Phillips et al. 2006) to describe the spatial distribution of the wild boar during two contrasting seasons (autumn-winter and spring-summer), to identify the main drivers of its habitat preferences and to draw seasonal maps of habitat quality, so to provide pertinent information to wildlife managers for improving management strategies.
Materials and methods
Study area
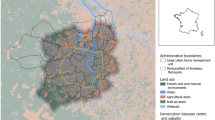
The study was carried out in MNR, a protected area which extends over 4696 ha within the municipality of Rome (central Italy), northeast of the urbanised area (Fig. 1a, b), nearby the Tiber River Valley. The climate is Mediterranean, with an average yearly temperature of 15.7 °C and an annual rainfall of 798 mm that shows a peak in late autumn and early winter and a minimum in summer. The area is dominated by arable lands (79.5%) (Fig. 1c), which include wheat (Triticum spp.), barley (Hordeum vulgare), oat (Avena sativa), clover (Trifolium spp.), alfalfa (Medicago sativa) and broad bean (Vicia faba). Permanent crops (3.4%) are mainly present as olive groves. Woodlands represent 11.3% of the total surface and are constituted chiefly by turkey oaks (Quercus cerrioides); common hornbeam (Carpinus betulus), hazel tree (Corylus avellana), narrow-leafed ash (Fraxinus angustifolia), white willow (Salix alba) and black locust (Robinia pseudoacacia) are less common. Urban settlements (3.1%), scrublands (1.5%) and pastures (1.2%) are scarcely present (Cafiero 2003; Macchiolo and Serafini Sauli 2007) (Fig. 1c). The first sighting of the wild boar dates back to 2010, when an individual was observed in the northeastern part of the MNR. Then, direct observations and tracks were not recorded until the end of 2013, when the species returned in the area from surroundings through natural ecological corridors (Amendolia et al. 2016). Since 2014, sightings have been more and more frequent, and the negative impact of the wild boar, mainly represented by damage to croplands, has drastically increased (Amendolia et al. 2016). At present, population density data are not available. Among ungulates, besides the wild boar, there is a small population (15–20 individuals) of fallow deer (Dama dama), originated following an escape of a few individuals from an enclosure. Large predators are absent: the wolf (Canis lupus) returned within the municipality of Rome in 2013 (Antonelli et al. 2018), but to this day it has not been observed in the MNR, and the presence of feral dogs is not reported (S. Amendolia, pers. comm.). Game shooting on wild boars is forbidden in the study area.

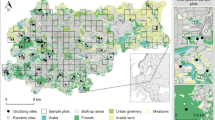
a Location of the study area (Marcigliana natural reserve, Rome, central Italy). b Location of the study area (in light grey) compared to the position of urban Rome (the area encompassed by the Great Ring Road). Within the Great Ring Road, in white: green spaces, which include historical villas, archaeological sites, meadows, grasslands, cultivated and uncultivated grounds, public gardens and parks; in dark grey: densely built-up areas. c Land use map of the study area and location of refined wild boar records (spring-summer, n = 52; autumn-winter, n = 15)
Species occurrence data
We collected 92 wild boar presence records from October 2013 to June 2016 from three different sources: (1) records of damage to croplands (n = 61), evaluated by agricultural engineers designated by the managing authority of the reserve; (2) diurnal observations reported by citizens to the rangers of the reserve (n = 17); and (3) diurnal observations made by the rangers of the reserve during patrolling activity (n = 14) (Table 1). Wild boar presence records were mapped and georeferenced with the geographic information system Quantum GIS version 2.12.3 (QGIS Development Team 2015) and divided into two groups for seasonal analyses: spring-summer (1 April–30 September, n = 72) and autumn-winter (1 October–31 March, n = 20). Finally, the presence records were processed to reduce the negative effect of sampling bias on model performance and to avoid spatial autocorrelation; following Morelle and Lejeune (2015), we applied a spatial filter, removing records within 300 m of one another and keeping the most records possible. Therefore, only 52 spring-summer and 15 autumn-winter presence points were included in the analyses (Fig. 1c shows a map of the study area with refined wild boar records).
Environmental variables
As predictors of wild boar habitat quality, we used seven environmental variables: turkey oak woods, other woods, scrublands, arable lands, permanent crops, meadows and pastures and urban settlements (Fig. 1c, Table 2). These variables were extracted from the land use map of the Latium Region, which is based on the same categories of the Corine Land Cover dataset, but at a higher level of detail (scale 1:10,000; http://dati.lazio.it/catalog/it/dataset/cus-lazio-approfondimento-delle-formazioni-naturali-e-seminaturali-iv-e-v-livello-corine-land-cover). Then, the study area was divided into 1-km2 squares forming a grid, and the percentage coverage of all variables was calculated in each square. The grid resolution corresponds to the approximate home range size of the wild boar in metropolitan areas (Podgórski et al. 2013). We considered potential multicollinearity among variables using Pearson’s coefficient, retaining r = 0.7 as a threshold value (Dormann et al. 2013). None of the pairwise comparisons resulted in a higher correlation value; therefore, we used all the variables in the analyses.
Model building and evaluation
We modelled the seasonal distribution of the wild boar using the maximum entropy algorithm implemented in the software Maxent (version 3.3.3k). This method creates a prediction based on spatially defined variables by comparing occurrence localities with a sample of background pixels (Phillips et al. 2006). We ran two models (spring-summer and autumn-winter) with auto features (which apply the feature class or classes estimated to be appropriate for the particular sample size of occurrence records), a logistic output (with predicted probability values ranging from 0 to 1), 5000 background points (Merow et al. 2013; Sieber et al. 2015) and a regularisation multiplier = 3, to lower the effect of overfitting (Radosavljevic and Anderson 2014).
To evaluate the performance of the models, we used a ten-fold cross-validation procedure. Data were split into ten independent subsets, and for each subset, models were trained with nine subsets and evaluated on the tenth one. The receiver operating characteristic (ROC) curve was used to assess the accuracy of models, as measured by the area under the ROC curve (AUC). AUC provides a measure of model discrimination ability, varying from 0.50, for a model with discrimination ability no better than random, to 1.00, for a model with a perfect discrimination ability. Models with AUC > 0.70 are considered to have an acceptable predictive capacity; to aid interpretation, we considered models as having an adequate (0.70 < AUC < 0.80), good (0.80 < AUC < 0.90) or excellent (AUC > 0.90) predictive capacity (Swets 1988).
We identified the variables most capable of predicting wild boar presence using two different parameters. The relative contribution of variables was estimated by calculating the percent contribution of each variable during the model training process. Then, permutation importance was estimated as the relative loss in AUC value of a model when the values of a given variable are randomly permuted among the presence points and random background points. A higher value of permutation importance means a greater relative loss in AUC value after random permutation, and, therefore, a greater reliance on the dependent variable (Phillips 2010).
Finally, we produced two seasonal maps describing the predicted probability of the presence of the wild boar in the study area.
Results
Spring-summer model
Model performance, as indicated by AUC value, was adequate (average AUC ± standard deviation, 0.72 ± 0.06). Urban settlements, turkey oak woods and arable lands were the most important variables in explaining the spatial distribution of the wild boar in spring and summer; nevertheless, turkey oak woods produced a low decrease in AUC value when randomly permuted (Table 3). The wild boar had a higher probability of occurring with an increasing presence of turkey oak woods, whereas the probability diminished with an increasing presence of urban settlements (Fig. 2). Arable lands had a strong negative effect only when very abundant (> 90%); when present in a smaller proportion, their negative effect was slighter (Fig. 2). All the other variables were less important. High-quality habitats for the wild boar are restricted to the northwestern part of the study area, while low-quality habitats are located predominantly in the southeastern part. Central and northeastern sectors are characterised by the dominance of medium-quality habitats (Fig. 3).

Response curves of the most important variables explaining the distribution of the wild boar in Marcigliana natural reserve in spring-summer. Values are averaged over ten replicates. Light grey margins show ± one SD calculated over ten replicates

Probability of the presence of the wild boar in Marcigliana natural reserve in spring-summer
Autumn-winter model
Model performance, as indicated by AUC value, was adequate (average AUC ± standard deviation, 0.78 ± 0.14). Meadows and pastures and urban settlements were the most important variables in explaining the spatial distribution of the wild boar in autumn and winter (Table 4), and they both had a negative effect on its probability of presence (Fig. 4). All the other variables were less important. High-quality habitats for the wild boar are restricted to some patches in the northern part of the study area, while central and southern sectors are dominated by medium- and low-quality habitats (Fig. 5).

Response curves of the most important variables explaining the distribution of the wild boar in Marcigliana natural reserve in autumn-winter. Values are averaged over ten replicates. Light grey margins show ± one SD calculated over ten replicates

Probability of the presence of the wild boar in Marcigliana natural reserve in autumn-winter
Discussion
The AUC values obtained for our habitat quality models are in accordance with the results from Bosch et al. (2014) and Morelle and Lejeune (2015) that used Maxent to predict wild boar distribution in Spain and Belgium, respectively. Our results support the idea that modelling the distribution of generalist species (as is the wild boar) results in models with lower performance (Franklin et al. 2009; Grenouillet et al. 2011). For specialist species with a small ecological niche, changes in the distribution are easier to detect because their requirements lead to movements towards habitats offering the necessary resources; in contrast, for generalist species, relating habitat changes to their requirements is more hazardous due to their ecological plasticity.
Maxent models emphasised the existence of seasonal differences in the habitats selected by the wild boar, with only urban settlements avoided year-round. Indeed, wild boars living in metropolitan areas exploit densely built-up areas only as corridors to move among patchy natural areas (Podgórski et al. 2013; Stillfried et al. 2017b), when access to natural resources is limited or when anthropogenic resources (e.g. garbage, vegetables in gardens, dry cat food) are more abundant than the natural ones (Hafeez et al. 2011; Stillfried et al. 2017c; Castillo-Contreras et al. 2018; Toger et al. 2018).
The spring-summer model stressed the importance of turkey oak woods and arable lands as predictors of wild boar distribution. Deciduous forests are commonly selected by the wild boar (Gerard et al. 1991; Merli and Meriggi 2006; Thurfjell et al. 2009; Rodrigues et al. 2016) because they can offer protection from predators and human disturbance, food resources and thermal cover. Predation risk, together with experience with humans and their recreational activities, commonly has a significant impact on ungulate behaviour (Mysterud and Østbye 1999; Stankowich 2008). Animals increase their vigilance to reduce the risk of predation and human disturbance, select habitats with good cover to decrease their visibility or shift their home ranges towards protected areas (Theuerkauf and Rouys 2008; Tadesse and Kotler 2012; Merli et al. 2017). In our study case, however, we do not link the exploitation of woodlands to the need for shelter. Indeed, in MNR, large predators are absent, and human disturbance is limited, since hunting is forbidden; forestry activities, although present, are strictly supervised and tourism is scarce, being represented essentially by few hikers who walk along the “Via Francigena” that crosses the area (S. Amendolia, pers. obs.). In view of this, we hypothesise that turkey oak woods are important for the wild boar as feeding areas and thermal refuges. In Italy, likewise elsewhere in southern Europe, hard mast (i.e. acorns, chestnuts and beech nuts) is one of the most important sources of food for the wild boar (Fournier-Chambrillon et al. 1995; Massei et al. 1996; Herrero et al. 2005). In particular, acorns produced by Quercus spp. are highly energetic and easy to digest, being essential to maintain a good physical condition and to keep a high reproductive rate (Massei et al. 1996; Frauendorf et al. 2016). Large ungulates, when the ambient temperature is too low or too high, commonly thermoregulate by seeking refuge under continuous cover associated with woodlands (Fernández-Llario 2004; van Beest et al. 2012; Cuevas et al. 2013; Marchand et al. 2015). In our study area, which is characterised by a Mediterranean climate, thermal cover is particularly important in the summer, when solar radiation is high, when temperatures are high and when water is scarce. In addition, wild boars do not have functional sweat glands; thus, they need to thermoregulate in the summer by resting in shady and cool places.
The wild boar seems to be partially adapted to the presence of arable lands, which have a negative effect only when very abundant. In the growing season, wild boars often utilise agricultural areas (Keuling et al. 2009; Thurfjell et al. 2009), as they represent an interesting source of food in this period of the year. This result, combined with the positive effect of turkey oak woods, leads us to hypothesise that in spring and summer, the species adopts a spatial strategy which optimises the trade-off between the need for thermal cover (provided by turkey oak woods) and the need for food resources (provided by turkey oak woods and arable lands).
Our autumn-winter model showed lower importance of the arable lands for the wild boar, probably linked to a lesser attraction of croplands in this part of the year, and a negative effect of meadows and pastures. An avoidance of this habitat has been highlighted by several previous studies (Fonseca 2008; Cuevas et al. 2013; Caruso et al. 2018); certainly, this is an open habitat that provides little cover and likely does not offer abundant food resources in autumn and winter relative to other habitats (i.e. woodlands).
Environmental conditions seem to be quite good for the wild boar. Despite the clear prevalence of cultivated areas, the non-negligible extension of woodlands undoubtedly favours the species, which has established in the last years a permanent presence, as demonstrated by the increasing number of records collected during the study period. The results presented here are expected to contribute to the development of effective management plans, with the main aims of limiting the negative impact of the species (Amendolia et al. 2016) and preventing the invasion of adjacent urban settlements. Numerous methods are usable to manage boar-human conflicts (see review in Massei et al. 2011). In MNR, evaluating both landscape features and the position of the study area, the use of trapping to control the expansion of the wild boar is advisable, focusing efforts in the areas identified by the Maxent models as the best ones for the species. Trapping, widely practiced and commonly considered as a cost- and time-effective method for wild boar control, targets primarily family groups that have the highest reproductive potential. In addition, it can achieve high reductions at small spatial scales and is usable in residential areas, where other methods are difficult to use (Massei et al. 2011). To increase trapping efficacy, a temporal planning of interventions is necessary. About that, besides the importance of seasonal habitat selection analyses, at least two important aspects require further research: the evaluation of food availability throughout the year and the analysis of wild boar reproductive phenology. Indeed, when food is scarce, wild boars are relatively easy to catch and a large number of animals can be captured (Massei et al. 2011), and trapping is most efficient in the reduction of invasive boar populations if focused during the low-birthing period (Pepin et al. 2017).
These results could be transferred to other urban and suburban areas, which have environmental characteristics comparable to those of MNR and have to face analogous boar-human conflicts.
References
Amendolia S, Lombardini M, Pierucci P, Meriggi A (2016) Human-wild boar conflicts in the metropolitan area of Rome (Central Italy). In: Angelici FM, Rossi L (eds) . Atti del III Congresso della Fauna Problematica, Cesena, pp 161–162
Antonelli M, De Lorenzis A, Benvenuti A, Fedrigo G, Lorenzini AR, Baini S, Festa F, Donfrancesco V, Huamán M, Bertagni G, Lanzi Mazzocchini D, Donfrancesco S (2018) Assessing the first reproduction of the wolf (Canis lupus italicus) in the coastal area of Rome in the last century. Hystrix It J Mamm 29:48 (supplement)
Apollonio M, Andersen R, Putman R (2010) European ungulates and their management in the 21st century. Cambridge University Press, Cambridge
Barrios-García MN, Ballari SA (2012) Impact of wild boar (Sus scrofa) in its introduced and native range: a review. Biol Invasions 14:2283–2300
Beasley JC, Ditchkoff SS, Mayer JJ, Smith MD, VerCauteren KC (2018) Research priorities for managing invasive wild pigs in North America. J Wildl Manag 82:674–681
Bosch J, Mardones F, Pérez A, de la Torre A, Muñoz MJ (2014) A maximum entropy model for predicting wild boar distribution in Spain. Span J Agric Res 12:984–999
Cafiero G (2003) La pianificazione del sistema delle aree protette di Roma: le nove Riserve naturali di RomaNatura. Collana dell’Istituto nazionale di urbanistica, Quaderni 37 (in Italian)
Cahill S, Llimona F, Cabañeros L, Calomardo F (2012) Characteristics of wild boar (Sus scrofa) habituation to urban areas in the Collserola Natural Park (Barcelona) and comparison with other locations. Anim Biodivers Conserv 35:221–233
Caruso N, Valenzuela AEJ, Burdett CL, Luengos Vidal EM, Birochio D, Casanave EB (2018) Summer habitat use and activity patterns of wild boar Sus scrofa in rangelands of central Argentina. PLoS One 13(10):e0206513
Castillo-Contreras R, Carvalho J, Serrano E, Mentaberre G, Fernández-Aguilar X, Colom A, González-Crespo C, Lavín S, López-Olvera JR (2018) Urban wild boars prefer fragmented areas with food resources near natural corridors. Sci Total Environ 615:282–288
Cuevas MF, Ojeda RA, Jaksic FM (2013) Multi-scale patterns of habitat use by wild boar in the Monte Desert of Argentina. Basic Appl Ecol 14:320–328
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G, García Marquéz JR, Gruber B, Lafourcade B, Leitão PJ, Münkemüller T, McClean C, Osborne PE, Reineking B, Schröder B, Skidmore AK, Zurell D, Lautenbach S (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:27–46
Fernández-Aguilar X, Gottschalk M, Aragon V, Càmara J, Ardanuy C, Velarde R, Galofré-Milà N, Castillo-Contreras R, López-Olvera JR, Mentaberre G, Colom-Cadena A, Lavín S, Cabezón O (2018) Urban wild boars and risk for zoonotic Streptococcus suis, Spain. Emerg Infect Dis 24:1083–1086
Fernández-Llario P (2004) Environmental correlates of nest site selection by wild boar Sus scrofa. Acta Theriol 49:383–392
Fonseca C (2008) Winter habitat selection by wild boar Sus scrofa in southeastern Poland. Eur J Wildl Res 54:361–366
Fournier-Chambrillon C, Maillard D, Fournier P (1995) Diet of the wild boar (Sus scrofa L.) inhabiting the Montpellier garrigue. Ibex J Mount Ecol 3:174–179
Franklin J, Miller JA (2009) Mapping species distribution: spatial inference and prediction. Cambridge University Press, Cambridge
Franklin J, Wejnert KE, Hathaway SA, Rochester CJ, Fisher RN (2009) Effect of species rarity on the accuracy of species distribution models for reptiles and amphibians in southern California. Divers Distrib 15:167–177
Frauendorf M, Gethöffer F, Siebert U, Keuling O (2016) The influence of environmental and physiological factors on the litter size of wild boar (Sus scrofa) in an agriculture dominated area in Germany. Sci Total Environ 541:877–882
Gerard J-F, Cargnelutti B, Spitz F, Valet G, Sardin T (1991) Habitat use of wild boar in a French agroecosystem from late winter to early summer. Acta Theriol 36:119–129
Gormley AM, Forsyth DM, Griffioen P, Lindeman M, Ramsey DSL, Scroggie MP, Woodford L (2011) Using presence-only and presence–absence data to estimate the current and potential distributions of established invasive species. J Appl Ecol 48:25–34
Grenouillet G, Buisson L, Casajus N, Lek S (2011) Ensemble modelling of species distribution: the effects of geographical and environmental ranges. Ecography 34:9–17
Hafeez S, Abbas M, Khan ZH, Rehman E (2011) Preliminary analysis of the diet of wild boar (Sus scrofa L., 1758) in Islamabad, Pakistan. Turk J Zool 35:115–118
Herrero J, Irizar I, Laskurain NA, García-Serrano A, García-González R (2005) Fruits and roots: wild boar foods during the cold season in the southwestern Pyrenees. Ital J Zool 72:49–52
Keuling O, Stier N, Roth M (2009) Commuting, shifting or remaining? Different spatial utilisation patterns of wild boar Sus scrofa L. in forest and field crops during summer. Mamm Biol 74:145–152
Licoppe A, Prévot C, Heymans M, Bovy C, Casaer J, Cahill S (2013) Wild boar / feral pig in (peri-) urban areas. International survey report as an introduction to the workshop: “managing wild boar in human-dominated landscapes”. IUGB Congress, Brussels, 28th August 2013
Macchiolo P, Serafini Sauli A (2007) La flora della Riserva Naturale della Marcigliana. Quaderni tecnici dei parchi del Lazio (in Italian)
Marchand P, Garel M, Bourgoin G, Dubray D, Maillard D, Loison A (2015) Sex-specific adjustments in habitat selection contribute to buffer mouflon against summer conditions. Behav Ecol 26:472–482
Massei G, Genov PV, Staines BW (1996) Diet, food availability and reproduction of wild boar in a Mediterranean coastal area. Acta Theriol 41:307–320
Massei G, Roy S, Bunting R (2011) Too many hogs? A review of methods to mitigate impact by wild boar and feral hogs. Hum Wildl Interact 5:79–99
Massei G, Kindberg J, Licoppe A, Gačić D, Šprem N, Kamler J, Baubet E, Hohmann H, Monaco A, Ozoliņš J, Cellina S, Podgórski T, Fonseca C, Markov N, Pokorny B, Rosell C, Náhlik A (2015) Wild boar populations up, numbers of hunters down? A review of trends and implications for Europe. Pest Manag Sci 71:492–500
Merli E, Meriggi A (2006) Using harvest data to predict habitat-population relationship of the wild boar Sus scrofa in northern Italy. Acta Theriol 51:383–394
Merli E, Grignolio S, Marcon A, Apollonio M (2017) Wild boar under fire: the effect of spatial behaviour, habitat use and social class on hunting mortality. J Zool 303:155–164
Merow C, Smith MJ, Silander JA Jr (2013) A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography 36:1058–1069
Monterosso G, Somaschini A, Gallo M, Babusci S, Di Primio S (2016) Piano per la programmazione di interventi di controllo numerico del cinghiale nel Parco di Veio. Technical report (in Italian)
Morelle K, Lejeune P (2015) Seasonal variations of wild boar Sus scrofa distribution in agricultural landscapes: a species distribution modelling approach. Eur J Wildl Res 61:45–56
Morelle K, Podgórski T, Prévot C, Keuling O, Lehaire F, Lejeune P (2015) Towards understanding wild boar Sus scrofa movement: a synthetic movement ecology approach. Mammal Rev 45:15–29
Mysterud A, Østbye E (1999) Cover as a habitat element for temperate ungulates: effects on habitat selection and demography. Wildl Soc Bull 27:385–394
Pepin KM, Davis AJ, VerCauteren KC (2017) Efficiency of different spatial and temporal strategies for reducing vertebrate pest populations. Ecol Model 365:106–118
Phillips S (2010) A brief tutorial on Maxent. Lessons Conserv 3:108–135
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Pierucci P, Semenzato P, Innocenti S, Giunti M (2014) Piano di gestione e controllo del cinghiale (Sus scrofa) nella Riserva Naturale Decima Malafede. Technical report (in Italian)
Pierucci P, Semenzato P, Mariucci A, Todini A (2015) Piano di gestione del cinghiale (Sus scrofa) nella Riserva Naturale dell’Insugherata. Technical report (in Italian)
Podgórski T, Baś G, Jędrzejewska B, Sönnichsen L, Śnieżko S, Jędrzejewski W, Okarma H (2013) Spatiotemporal behavioral plasticity of wild boar (Sus scrofa) under contrasting conditions of human pressure: primeval forest and metropolitan area. J Mammal 94:109–119
Primi R, Viola P, Serrani F, Balzarani M, Tiberi C, Rossi CM, Amici A (2016) Update on wild boar (Sus scrofa) distribution in the Metropolitan City of Rome. In: Angelici FM, Rossi L (eds) Atti del III Congresso della Fauna Problematica. Cesena (Italy), p 171
QGIS Development Team (2015) QGIS geographic information system. Open Source Geospatial Foundation Project. http://www.qgis.org/
Radosavljevic A, Anderson RP (2014) Making better Maxent models of species distributions: complexity, overfitting and evaluation. J Biogeogr 41:629–643
Rodrigues P, Herrero J, García-Serrano A, Prada C, Giménez-Anaya A, Ayala R, Fernández-Arberas O, Fonseca C (2016) Habitat use by wild boar Sus scrofa in Moncayo Nature Park, Spain. Pirineos 171:e023
Sieber A, Uvarov NV, Baskin LM, Radeloff VC, Bateman BL, Pankov AB, Kuemmerle T (2015) Post-Soviet land-use change effects on large mammals’ habitat in European Russia. Biol Conserv 191:567–576
Stankowich T (2008) Ungulate flight responses to human disturbance: a review and meta-analysis. Biol Conserv 141:2159–2173
Stillfried M, Fickel J, Börner K, Wittstatt U, Heddergott M, Ortmann S, Kramer-Schadt S, Frantz AC (2017a) Do cities represent sources, sinks or isolated islands for urban wild boar population structure? J Appl Ecol 54:272–281
Stillfried M, Gras P, Börner K, Göritz F, Painer J, Röllig K, Wenzler M, Hofer H, Ortmann S, Kramer-Schadt S (2017b) Secrets of success in a landscape of fear: urban wild boar adjust risk perception and tolerate disturbance. Front Ecol Evol 5(157):1–12
Stillfried M, Gras P, Busch M, Börner K, Kramer-Schadt S, Ortmann S (2017c) Wild inside: urban wild boar select natural, not anthropogenic food resources. PLoS One 12(4):e0175127
Swets JA (1988) Measuring the accuracy of diagnostic systems. Science 240:1285–1293
Tadesse SA, Kotler BP (2012) Impact of tourism on Nubian Ibex (Capra nubiana) revealed through assessment of behavioral indicators. Behav Ecol 23:1257–1262
Theuerkauf J, Rouys S (2008) Habitat selection by ungulates in relation to predation risk by wolves and humans in the Białowieża Forest, Poland. For Ecol Manag 256:1325–1332
Thurfjell H, Ball JP, Åhlén P-A, Kornacher P, Dettki H, Sjöberg K (2009) Habitat use and spatial patterns of wild boar Sus scrofa (L.): agricultural fields and edges. Eur J Wildl Res 55:517–523
Toger M, Benenson I, Wang Y, Czamanski D, Malkinson D (2018) Pigs in space: an agent-based model of wild boar (Sus scrofa) movement into cities. Landsc Urban Plan 173:70–80
van Beest FM, Van Moorter B, Milner JM (2012) Temperature-mediated habitat use and selection by a heat-sensitive northern ungulate. Anim Behav 84:723–735
Zuberogoitia I, del Real J, Torres JJ, Rodríguez L, Alonso M, Zabala J (2014) Ungulate vehicle collisions in a peri-urban environment: consequences of transportation infrastructures planned assuming the absence of ungulates. PLoS One 9(9):e107713
Acknowledgements
We are grateful to the rangers of Marcigliana natural reserve for their help in data collection and to Fabrizio Foschi for his assistance in layers digitization. We also would like to thank two anonymous reviewers for their helpful comments on the first version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by: Teresa Abaigar Ancín
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Amendolia, S., Lombardini, M., Pierucci, P. et al. Seasonal spatial ecology of the wild boar in a peri-urban area. Mamm Res 64, 387–396 (2019). https://doi.org/10.1007/s13364-019-00422-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-019-00422-9