
Abstract
The present study was undertaken with an aim of characterization of rabies virus (genus Lyssavirus of the family Rhabdoviridae under the order Mononegavirales) by sequencing of partial nucleoprotein (N) gene of rabies virus and phylogenetic analysis to know the genotype and lineage of rabies virus present in Gujarat state of India. A total of 32 samples (18 brain samples and 14 saliva samples) were aseptically collected from live and dead animals (viz. dog, buffalo, cow, goat, donkey and hyena) for rabies virus detection. Out of 32 samples, 24 samples were found positive by Reverse Transcriptase Polymerase Chain Reactions and from these 24 positive samples, 20 samples were selected for sequencing having good concentration of gene product. ClustalW alignment of nucleotide sequences and amino acid sequences of field rabies isolates revealed 95.20–100 and 97.95–100% similarity among themselves, respectively. Multiple sequence alignment of field rabies isolates and reference vaccine strains [Pasteur strain and Challenge Virus Strain (CVS)] indicated single nucleotide mutations at total 91 positions and amino acid mutations at total 17 different positions. Phylogenetic analysis of N gene sequences using our 20 field rabies isolates and 21 other reported isolates in Genbank resulted in 3 phylogenetic clusters. All the field rabies isolates showed same genetic lineage among themselves and with other earlier reported Indian rabies isolates placing them in Arctic like lineage of Genotype 1 Rabies virus. However, they were at genetic distance with reference Pasteur and CVS strains, which grouped in different phylogenetic cluster.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rabies is historically one of the most significant zoonotic diseases, because of nearly 100% case fatality rate and ubiquitous global distribution [3]. Rabies is present in many parts of the world, particularly in countries of Asia, Africa and Latin America since antiquity except Antarctica where the disease is enzootic despite the availability of proven prevention and control tools but more than 95% of human deaths occur in Asia and Africa [21, 26]. India has the highest incidence of rabies [14, 15] with an estimated 20,000 human death and 17.4 million exposures to animal bite every year, which corresponds to 1.7% incidence rate of animal bites [25].
Rabies virus is the prototype member of the genus Lyssavirus of the family Rhabdoviridae under the order Mononegavirales [26]. It is a single stranded, negative-sense RNA virus, Lyssavirus (genotype 1) with a genome size of approximately 12 kb [4, 7, 11]. Viral genome encodes total five structural genes which are separated by four non‐coding intergenic sequences from 3′ terminus to 5′ terminus in the order of N–P–M–G–L, which encode respectively the nucleoprotein (N), phosphoprotein (P), matrix protein (M), glycoprotein (G) and large subunit of transcriptase (L) with two untranslated regions (UTRs) at the ends of the genome [27]. All these structures are crucial for the replication and encapsidation of rabies virus RNA [6, 24].
Study of canine rabies dynamics in the field would help to understand the genetic variants in circulation and their evolutionary relationships thereby helping to devise effective control measures. The genetic typing technique, which employs Reverse Transcriptase Polymerase Chain Reaction (RT-PCR), provides an opportunity to elucidate the epidemiologic and evolutionary relationships between rabies virus (RABV) and rabies related viruses (RABVs) [10]. There are very few reports available on molecular epidemiology of rabies virus of Indian origin. Till date, the N and G genes of rabies virus have most often been targeted for genomic and evolutionary studies [16]. N gene in the whole genome of rabies virus is highly conserved and can be efficiently expressed, which is the reason why N gene is frequently used as a judging index for genotyping and population variation of rabies viruses [27]. Selection of N gene for the sequencing and phylogenetic analysis is also because of following reasons: first, the amplification of the N gene using PCR as a simple diagnostic technique can be expanded to allow a precise typing method based on nucleotide sequences [19]. Second, the N gene sequences of representative isolates of all six genotypes of Lyssavirus are available [4]. Third, from comparative point of view, the important contribution of capsid proteins to varied structural and immunological functions and the purported role of other viral nucleoprotein like Influenza nucleoprotein as a determinant of host range [20] make the evolutionary analysis and genetic variability of rabies N protein interesting but full genome sequencing of N gene of rabies virus would provide more robust data that is the reason why partial N gene has selected for sequencing. The N gene, although highly conserved allows viral strains to be accurately differentiated by analyzing genetic differences that are present within the gene [9].
There are very few reports available on molecular epidemiology of rabies virus of Indian origin and because of that the present study was performed to characterize rabies virus by sequencing of partial N gene of rabies virus and phylogenetic analysis with earlier reported sequences of Indian and foreign origin to know the genotype and lineage of rabies virus present in Gujarat state.
Materials and methods
Rabies viruses
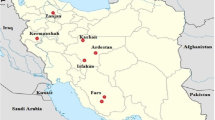
A total of 32 samples (18 brain samples and 14 saliva samples) were aseptically collected from live and dead animals (viz. dog, buffalo, cow, goat, donkey and hyena) for rabies virus detection from different geographical locations of Gujarat state. Out of 32 samples, 24 samples were found positive by RT-PCR and from these 24 positive samples, 20 samples were selected for partial N gene sequencing having good concentration of gene product of rabies virus. The details of collected samples are listed in Table 1.
RT-PCR
RT-PCR product was prepared by QIAGEN® OneStep RT-PCR Kit (Catlog No. 210210, QIAGEN, Germany). The primer set used for one step RT-PCR was JW12(F) ATGTAACACCTCTACAATG and JW6(R) CAATTAGCACACATTTTGTG targeting 605 bp size amplicon of the nucleoprotein gene of rabies virus [1]. RT-PCR was carried out in a final reaction volume of 50 μl using 200 μl capacity thin walled PCR tubes comprising of 10 μl Qiagen one-step RT-PCR Buffer (5×), 2 μl of dNTP mix (10 mMol), 3.0 μl of each primer (10 pmol) (Eurofins Genomics India Pvt Ltd), 2 μl of Qiagen one-step RT-PCR enzyme mix, 10 μl of RNA template (30 ng/μl) and 20 μl of RNAse free water (Fermentas). The RT-PCR reactions were performed in thermocycler (Biorad PCR) with following cycles; reverse transcription 50 °C for 30 min, initial denaturation of 95 °C for 15 min followed by 35 cycles of denaturation, annealing and extension at 94 °C for 30 s, 50 °C for 30 s and 72 °C for 60 s, respectively, and the final extension was carried out at 72 °C for 10 min.
Nucleotide sequencing
The purification of 50 μl RT-PCR product was carried out by QIAquick PCR Purification Kit (Cat. No. 28106, QIAGEN Co., Valencia, CA). Purified DNA was quantified by spectrophotometric analysis using NanoDrop 1000 Spectrophotometer V3.7 (Thermo Fisher Scientific, USA). Sequencing was performed following the instructions supplied along with Big Dye® Terminator V3.1 Cycle Sequencing Kit (Applied Biosystems). The reaction was carried out in a final reaction volume of 20 μl using 200 μl capacity thin wall PCR tubes comprising of Ready reaction premix (2 μl), Big Dye sequencing buffer (5×) (4 μl), Exon specific primer-forward or reverse each (2.5 pmol/μl) (2 μl), Template DNA—PCR product (60 ng) and Deionized water to make up to 20 μl volume. The cycle sequencing reactions were performed in thermocycler (Biorad, USA) with following cycles; initial denaturation of 96 °C for 2 min followed by 29 cycles of denaturation, annealing and extension at 96 °C for 12 s, 50 °C for 15 s and 68 °C for 4 min, respectively.
Purification was carried out from the final reaction volume of cycle sequencing (20 μl), using ZR DNA Sequencing Clean-up Kit, (Cat. No. D4051, Zymo Research, USA) according to manufacturer instructions. The filtrate obtained was directly loaded into MicroAmp Optical 96-Well Reaction Plate (Applied Biosystems) and analyzed using ABI 3500 Genetic analyzer. The obtained forward and reverse sequences of N gene were aligned with each other using SeqScape v 2.5 sequence analysis software. The sequence similarity search with the DNA public Database available at Genbank, maintained by NCBI, was carried out with the help of Basic Local Alignment Search Tools-BLAST. Multiple sequence analysis was carried out by clustalW within obtained sequences and with two reference sequences (Pasteur and CVS strain). The neighbor-joining tree (NJ tree) was drawn by using the MEGA version 6.0 program [23], with confidence levels assessed by the use of 1000 bootstrap replications. A total of 21 strains/isolates from India and foreign countries with their gene accession numbers were used for sequence analysis and phylogenetic study.
Results
RT-PCR
Out of total 32 brain and saliva samples, 16 brain and eight samples were found positive by amplifying the expected 605 bp size amplicon on gel documentation system. Negative samples had not showed any band (Fig. 1).

RT-PCR results using brain and saliva samples for rabies diagnosis
Nucleotide sequences
All the 20 selected isolates from different animal species yielded nucleotide sequences of desired nucleotide length ranging from 584 to 606 bp and submitted to Genbank of NCBI database. Genebank accessions numbers were KU867249, KU882952 to KU882961 and KU958724 to KU958732. Deduced Amino acid sequences of partial N gene of rabies virus were obtained using ExPAsy proteomics tools.
Sequence analysis and phylogenetic study
ClustalW alignment of all the 20 nucleotide sequences revealed 95.20–100% similarity among themselves. Alignment of Amino acid sequences using clustalW revealed 97.95–100% similarity among themselves. Multiple sequence alignment was performed with the sequences of two reference vaccine strains viz. Pasteur and CVS strains (Accession no. NC_001542 and AF406696). The multiple nucleotide sequence alignment of Pasteur strain and CVS strain with field rabies viruses exhibited 89.10–90.75 and 88.07–90.84% identity, respectively. Multiple Amino acid sequence alignment showed 94.38–95.91 and 94.89–96.42% identity with Pasteur and CVS strains.
In multiple sequence alignment of 20 nucleotide sequences of field rabies viruses with reference vaccine strains and between themselves revealed variations at total 91 positions. However, significant variations were observed at respective positions in different field isolates and reference strains were as per following. Position: 92, 337: At these positions, the reference strain CVS showed nucleotide ‘C’ (Cytosine), while all the field isolates and Pasteur strain had nucleotide ‘T’ (Thymine). Position: 112, 199, 380, 418, 433, 478, 508, 517: Pasteur strain and CVS strain showed nucleotide ‘C’ at these positions, whereas nucleotide ‘T’ was present in all the field isolates. Position: 121, 154, 166, 235, 256, 460, 532, 535: Pasteur strain and CVS strain showed nucleotide ‘T’ at these positions, whereas nucleotide ‘C’ was present in all the field isolates. Position: 134: At this position, both the reference strains had nucleotide ‘T’, whereas all the field viruses showed nucleotide ‘A’ (Adenine). Position: 142: Only the Pasteur reference strain had nucleotide ‘T’, whereas CVS reference strain and all field viruses showed nucleotide ‘C’. Position: 148, 163, 208, 295, 352, 577: Pasteur strain and CVS strain showed nucleotide ‘A’, but all the field viruses showed nucleotide ‘G’ (Guanine). Position: 151, 198, 238, 262, 333, 551: Here, Pasteur strain and CVS strain presented nucleotide ‘G’, whereas all the field viruses had nucleotide ‘A’. Position: 187: Only the CVS reference strain showed nucleotide ‘G’, while all the field isolates and Pasteur reference strain had nucleotide ‘A’. Position: 191: At this position, Pasteur reference strain showed nucleotide ‘T’, whereas all the field isolates and CVS strain had nucleotide ‘G’. Position: 194: Pasteur strain and CVS strain showed nucleotide ‘A’, but all the field viruses were different from that showing nucleotide ‘T’. Position: 241, 486: Here, only Pasteur strain had nucleotide ‘G’, while all the field isolates and CVS reference strain showed nucleotide ‘A’. Position: 244, 299: At these positions, both the reference strains had nucleotide ‘G’, whereas the field viruses had nucleotide ‘T’. Position: 391: At this position, Pasteur strain and CVS strain had nucleotide ‘C’, while all the field isolates showed nucleotide ‘A’.
Multiple Amino acid sequence comparison showed single Amino acid variations at total 17 positions. Among that variations at positions 34, 45, 170, 181, 193, 194, 195 and 196 were between field rabies isolates themselves Fig 2. However, significant variations were observed at respective positions in different field isolates and reference strains, Position: 26: At this position, Reference strain CVS showed H (Histidine) in place of Y (Tyrosine) which was present in all the field isolates and Pasteur reference strain. Position: 40: Pasteur strain and CVS strain, both had amino acid C (Cysteine), but all the field isolates showed amino acid S (Serine) at this position. Position: 59: All the field samples and CVS reference strain showed amino acid G (Glycine), but amino acid C (Cysteine) was presented by Pasteur reference strain. Position: 60: Pasteur strain and CVS strain were with amino acid M (Methionine) at this position, while all the field isolates showed amino acid L (Leucine). Position: 61: At this position, Pasteur strain and CVS strain had amino acid S (Serine), but all the field isolates showed amino acid N (Asparagine). Position: 95: Amino acid V (Valine) was present in both the reference strains, but amino acid L (Leucine) was present in all the field isolates. Position: 106: Field isolates differed from reference strains by showing amino acid D (Aspartic acid) in place of amino acid G (Glycine). Position: 157: All the field isolates along with CVS reference strain showed amino acid N (Asparagine) while Pasteur reference strain had amino acid S (Serine). Position: 179: Here, reference strains, Pasteur and CVS had amino acid V (Valine), while all the field isolates showed I (Isoleucine) amino acid. Position: 195: At this position, only one isolate And_Rabies_Hyena_Brain_1 showed L (Leucine) amino acid, whereas remaining all the field isolates and reference strains had A (Alanine) amino acid.

Multiple sequence alignment of deduced amino acid sequences of field rabies isolates with each other and reference strains
Phylogenetic analysis
The phylogenetic study using MEGA 6 with NJ algorithm using consensus of 1000 bootstrap replicates with 21 sequences of known RV isolates including two reference strains revealed three distinct clusters with Indian and foreign isolates (Fig. 3). Cluster I included all our field isolates along with India-Human DQ521242, India-Dog1 DQ521215, India Dog2 DQ521216, India-Dog3 DQ521218, India-Cattle AY854599, Nepal-Dog JX987742, Canada-Dog U03769, Canada-Fox L20672 and Russia-Fox U22656 isolates. Cluster II was formed with Pasteur virus NC001542, CVS strain AF406696, Namibia-Kudu JQ691441, Hungary-Red Fox U43000, Germany-Red Fox U42701, Russia-Dog AY352476, Iran-Wolf U22482, Turkey-Dog DQ837476 and Sudan-Dog FJ94702 isolates. Lastly, cluster III was formed by Sri Lanka-Water Buffalo AB041969 and Nepal-Buffalo JX987737. One isolate Indonesia-Dog EU086192, remained out grouped by not falling into the above three clusters.

Phylogenetic relationship of 20 field rabies isolates to other rabies isolates along with reference strains (0.02 means nucleotide changes or substitutions per site)
Discussion
In present study, we found that all the 20 field rabies virus isolates were found to have 89.10–90.75 and 88.07–90.84% nucleotide sequence similarity with Pasteur and CVS strains, respectively. Multiple Amino acid sequence alignment showed 94.38–95.91 and 94.89–96.42% similarity with Pasteur and CVS strains. The phylogenetic study using MEGA 6 with NJ algorithm using consensus of 1000 bootstrap replicates with 21 sequences of known RV isolates including two reference strains revealed three distinct clusters with Indian and foreign isolates.
In our study, all the 20 nucleotide sequence alignment using clustalW revealed 95.20–100% similarity and amino acid sequence alignment using clustalW revealed 97.95–100% similarity, among themselves. The multiple nucleotide sequence alignment of 20 field rabies viruses with Pasteur strain and CVS strain exhibited variations at total 91 positions indicating 89.10–90.75 and 88.07–90.84% nucleotide sequence similarity, respectively whereas multiple amino acid sequence alignment showed variations at total 17 positions indicating 94.38–95.91 and 94.89–96.42% identity with Pasteur and CVS strains (Fig. 2). Chiou et al. [5] found 97–99% nucleotide identity of the N gene of rabies virus within three isolates from Taiwan Ferret Badgers. Jamil et al. [8] revealed 98–100% nucleotide identity between N gene sequences of rabies virus from seven brain samples. Reddy et al. [18] found 88.8–99.7% homology of nucleoprotein gene between 30 Indian rabies isolates and the standard Pasteur virus strain. Nagrajan et al. [16] found that South Indian RV isolates showed >95% sequence homology and analysis of the deduced N terminus amino acid sequences of nucleoprotein revealed that South Indian RV isolates differed from North Indian RV isolates at amino acid 134 (aa134), but all the South Indian RV isolates exhibited 100% amino acid homology. Marston et al. [12] found Aspartic Acid at position 106 (Asp106) which was unique in Africa 1a group members, in contrast Glu106 is unique to Sudan, which supports the concept of a single distinct virus strain circulating in Sudan. Susetya et al. [22] compared 34 rabies isolates with 20 sequences from other regions or three vaccine strains and showed nucleotide homologies of 88.4–90.2, 86.1–88.4, 85.9–87.4 and 86.2–87.4% with sequences from China, Thailand, India and Shri Lanka, respectively. The present study findings endorsed the finding of Chiou et al. [5], Jamil et al. [8] and Nagrajan et al. [16]. Deduced amino acid sequence similarity was 97.95–100% amongst the field isolates which was almost similar to observations of Nagrajan et al. [16].
In present study, phylogenetic analysis of field rabies isolates revealed that the field rabies isolates and the reference Pasteur and CVS strains grouped into different phylogenic clusters indicating genetic distance. Our field rabies isolates were also at genetic distance with the other known isolates from different countries, which grouped into cluster II and cluster III. It was noteworthy that all our field isolates were of high genetic similarity among themselves as well as with other Indian isolates of human, dog and cattle origin as all of them were found in the same phylogenetic cluster I. The genetic distance between cluster I and cluster II was 0.03, which was 0.10 between cluster I and cluster III. Pasteur strain was at 0.10 genetic distance with our field isolates and at 0.08 genetic distance with whole cluster I. CVS strain was at 0.11 genetic distance with our field isolates and at 0.09 genetic distance with whole cluster I.
Chiou et al. [5] revealed that the strain of RABV of TWFBs (RABV-TWFB) belongs to Lyssavirus genotype 1 by phylogenetic analysis. Mehta et al. [13] found that phylogenetic analysis of 20 rabies suspected samples of humans from Maharashtra, India revealed genetic relatedness to RABV strains reported from Indo-Arctic lineages and India-South and Japan. Pant et al. [17] found the presence of a surprising wide genetic diversity of RABV circulating in Nepal. Phylogenetic analysis of 24 isolates revealed three different co-existing phylogenetic groups: an Indian subcontinent clade and two different Arctic-like sub-clades within the Arctic-related clade, namely Arctic-like (AL)-1, lineage a (AL-1a), and AL-3. Jamil et al. [8] found that the rabies viruses in Bangladesh belong to Arctic/Arctic-like group 2 (AAL2) also known as Arctic-like-1, in close association with the strain from Bhutan by phylogenetic analysis. Reddy et al. [18] found that 30 rabies virus isolates from different states of India were genetically closely related with a single cluster under Arctic/Arctic-like viruses. Arvindhbabu et al. [2] found that two adult cow isolates belonged to the genetic cluster 3 (GC3) which is restricted to southern India and GC3 was found to be related to Sri Lankan rabies isolates with a strong bootstrap value of 96%. Nagrajan et al. [16] showed that RV isolates in Southern India belong to genotype 1 by phylogenetic analysis of the portion of nucleoprotein gene sequence of 22 rabies virus isolates obtained from domestic animals in Southern India.
The present phylogentic analysis showed that our field rabies isolates were of high nucleotide sequence similarity with each other and with earlier reported Indian sequences by falling into one cluster that confirms that our field rabies isolates belongs to genotype 1 [16], and because of clustering with other reported Indian isolates which are of Arctic like lineage, our isolates also belong to Arctic like lineage. This endorses the findings of Mehta et al. [13], Pant et al. [17], Jamil et al. [8], Reddy et al. [18], Arvindhbabu et al. [2] and Nagrajan et al. [16].
Unifying the overall findings of the present study, Partial N gene sequences and their analysis revealed multiple Single Nucleotide Variations (SNVs) amongst the field rabies viruses from various species and the reference viruses, although few unique mutations and accordingly signatory amino acid positions could be recorded in reference as well as few field rabies viruses. However, phylogenetic analysis revealed the genetic relatedness among the field rabies viruses and a few earlier reported isolates of India, placing them in Arctic like lineage of Genotype 1 Rabies virus.
Partial N gene sequence analysis revealed 95.20–100% nucleotide sequence similarity and 97.95–100% amino acid sequence similarity within the field rabies isolates from different species and locations. Multiple sequence alignment of field rabies isolates and reference vaccine strains (Pasteur and CVS strain) indicated SNVs at total 91 positions and amino acid variations at total 17 different positions. The multiple sequence alignment revealed 89.10–90.75 and 94.38–95.91% similarity with Pasteur reference vaccine strain and 88.07–90.84 and 94.89–96.42% similarity with CVS reference vaccine strain at nucleotide and amino acid levels, respectively. Phylogenetic analysis of N gene sequences using our 20 field rabies isolates and 21 other reported isolates in Genbank including 5 Indian, 14 foreign and 2 reference strains resulted in 3 phylogenetic clusters. All the field rabies isolates showed same genetic lineage among themselves and with other earlier reported Indian rabies isolates placing them in Arctic like lineage of Genotype 1 Rabies virus. However, they were at genetic distance with reference Pasteur and CVS strains, which are grouped in different phylogenetic cluster.
References
ArvindhBabu RP, Manoharan S, Ramadass P. Diagnostic evaluation of RT-PCR-ELISA for the detection of rabies virus. VirusDisease. 2014;25(1):120–4.
AravindhBabu RP, Manoharan S, Ramadass P, Chandran NDJ. Rabies in South Indian Cows: an evidence of Sri Lankan rabies virus variant infection based on the analysis of partial nucleoprotein gene. Indian J Virol. 2011;22(2):138–41.
Blanton JD, Rupprecht CE. Travel vaccination for rabies. Expert Rev Vaccines. 2008;7:613–20.
Bourhy H, Kissi B, Tordo N. Molecular diversity of the Lyssavirus genus. J Virol. 1993;194:70–81.
Chiou H, Jeng C, Wang H, Inoue S, Chan F, Liao J, Chiou M, Pang VF. Pathology and molecular detection of rabies virus in Ferret badgers associated with a rabies outbreak in Taiwan. J Wildl Dis. 2016;52(1):57–69.
Finke S, Cox JH, Conzelmann KK. Differential transcription attenuation of rabies virus genes by intergenic regions: generation of recombinant viruses over expressing the polymerase gene. J Virol. 2000;74:7261–9.
Gould AR, Hyatt AD, Lunt R, Kattenbelt JA, Hengstberger S, Blacksell SD. Characterisation of a novel lyssavirus isolated from Pteropid bats in Australia. Virus Res. 1998;54:165–87.
Jamil KM, Ahmed K, Hossain M, Matsumoto T, Ali MA, Hossain S, Islam A, Nasiruddin M, Nishizono A. Arctic-like rabies virus, Bangladesh. Emerg Infect Dis. 2012;18(12):2021–4.
Johnson N, Lipscomb DW, Stott R, Gopal Rao G, Mansfield K, Smith J, McElhinney LM, Fooks AR. Investigation of a human case of rabies in the United Kingdom. J Clin Virol. 2002;25:351–6.
Kissi B, Tordo N, Bourhy H. Genetic polymorphism in the rabies virus nucleoprotein gene. Virology. 1995;209:526–37.
Kuzmin IV, Hughes GJ, Botvinkin AD, Orciari LA, Rupprecht CE. Phylogenetic relationships of Irkut and West Caucasian bat viruses within the Lyssavirus genus and suggested quantitative criteria based on the N gene sequence for lyssavirus genotype definition. Virus Res. 2005;111:28–43.
Marston DA, McElhinney LM, Ali YH, Intisar KS, Ho SM, Freuling C, Müller T, Fooks AR. Phylogenetic analysis of rabies viruses from Sudan provides evidence of a viral clade with a unique molecular signature. Virus Res. 2009;145:244–50.
Mehta S, Charan P, Dahake R, Mukherjee S, Chowdhary A. Molecular characterization of nucleoprotein gene of rabies virus from Maharashtra, India. J Postgrad Med. 2016;62(2):105–8.
Meng S, Xu G, Wu X, Lei Y, Yan J, Nadin-Davis SA, Liu H, Wu J, Wang D, Dong G, Yang X, Rupprecht CE. Transmission dynamics of rabies in China over the last 40 years: 1969–2009. J Clin Virol. 2010;49:47–52.
Ming P, Yan J, Rayner S, Meng S, Xu G, Tang Q, Wu J, Luo J, Yang X. A history estimate and evolutionary analysis of rabies virus variants in China. J Gen Virol. 2010;91:759–64.
Nagarajan T, Nagendrakumar SB, Mohanasubramanian B, Rajalakshmi S, Hanumantha NR, Ramya R, Thiagarajan D, Srinivasan VA. Phylogenetic analysis of nucleoprotein gene of dog rabies virus isolates from Southern India. Infect Genet Evol. 2009;9:976–82.
Pant GR, Lavenir R, Wong FYK, Certoma A, Larrous F, Bhatta DR, Bourhy H, Stevens V, Dacheux L. Recent emergence and spread of an arctic-related phylogenetic lineage of rabies virus in Nepal. PLoS Negl Trop Dis. 2013;7(11):e2560.
Reddy GBM, Singh R, Singh RP, Singh KP, Gupta PK, Desai A, Shankar SK, Ramakrishnan MA, Verma R. Molecular characterization of Indian rabies virus isolates by partial sequencing of nucleoprotein (N) and phosphoprotein (P) genes. Virus Genes. 2011;43:13–7.
Sacramento D, Bourhy H, Tordo N. PCR technique as an alternative method for diagnosis and molecular epidemiology of rabies virus. Mol Cell Probes. 1991;6:229–40.
Scholtissek C, Burger H, Kistner O, Shortridge KF. The nucleoprotein as a possible major factor in determining host specificity of influenza H3N2 viruses. Virology. 1985;147:287–94.
Sudarshan MK, Madhusudana SN, Mahendra BJ, Rao NS, Ashwath Narayana DH, Abdul Rahman S, Meslin F, Lobo X, Ravikumar D, Gangaboraiah K. Assessing the burden of human rabies in India: results of a national multicentre epidemiological survey. Int J Infect Dis. 2007;11:29–35.
Susetya H, Sugiyama M, Inagaki A, Ito N, Mudiarto G, Minamoto N. Molecular epidemiology of rabies in Indonesia. Virus Res. 2008;135:144–9.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013. doi:10.1093/molbev/mst197.
Tordo N, Poch O, Ermine A, Keith G, Rougeon F. Completion of the rabies virus genome sequence determination: highly conserved domains along the L (polymerase) proteins of unsegmented negative-strand RNA viruses. Virology. 1988;165:565–76.
Wilde H, Briggs DJ, Meslin FX, Hemachudha T, Sitprija V. Rabies update for travel medicine advisors. Clin Infect Dis. 2003;37:96–100.
Wunner WH, Calisher CH, Dietzgen RG, Jackson RG, Kitajima AO, Lafon MF, Leong JC, Nichol ST, Peters D, Smith JS, Walker PJ. Rhabdoviridae. In: Classification and nomenclature of viruses. Sixth report of the international committee on taxonomy of viruses, in press. New York: Springer; 1995.
Wunner WH, Larson JK, Dietzschold B, Smith CL. The molecular biology of rabies viruses. Rev Infect Dis. 1998;10:771–84.
Author information
Authors and Affiliations
Corresponding author
Additional information
Part of M.V.Sc. thesis submitted by first author to AAU, Anand.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Vagheshwari, D.H., Bhanderi, B.B., Mathakiya, R.A. et al. Sequencing and sequence analysis of partial nucleoprotein (N) gene and phylogenetic analysis of rabies virus field isolates from Gujarat state, India. VirusDis. 28, 320–327 (2017). https://doi.org/10.1007/s13337-017-0387-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13337-017-0387-3