
Abstract
This work supports previous studies in the Great Barrier Reef lagoon that show the new nitrogen (N) load introduced by Trichodesmium is similar to or greater than that from riverine discharges. However, the current management programs aimed at improving the chronic eutrophic state of the GBR ignore the N load from Trichodesmium. These programs also ignore the evidence that Trichodesmium blooms could promote the bioavailability of heavy metals and be a source of toxins in the ciguatera food chain. Further work is urgently required to better quantify the potential impacts of Trichodesmium and develop management plans to reduce those impacts. A simple algorithm that uses MODIS imagery is developed for not only monitoring the spatial extent of Trichodesmium blooms but also for quantifying the concentration of those blooms. The algorithm is based on the readily available MODIS L2 data. A management plan that includes the harvesting of Trichodesmium is outlined.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Past farming practices in Queensland have led to serious degradation of the coastal catchments over the past century; more than 80% of the coastal catchments have been developed for agriculture and this development has led to increased loads of sediment and nutrients in the run-off entering the Great Barrier Reef (GBR) lagoon (Bell and Elmetri 1995; Kroon et al. 2012; RWQPP 2014; Reside et al. 2017). The increased nutrient and sediment loads in the run-off have led to serious degradation of water quality over the entire GBR lagoon, even in the remote Cape York Region (Bell 1992; Bell and Elmetri 1995). The principal water quality problem impacting the GBR coral reefs is related to the increased production of algae (i.e. eutrophication) that has resulted from the increased loads of nutrients/fertilizers washed into the system following high rainfall events. The degraded water quality has led to serious changes to the ecology of the GBR lagoon. There is evidence that the addition of nutrients to the GBR region has not only stimulated the growth of algae and heterotrophs (e.g. including crown of thorns (COTS) larvae, Drupella spp. larvae and protozoa such as ciliates) but has also lead to the proliferation of coral skeletal diseases and jellyfish (Bell et al. 2007, 2012; Bell et al. 2014ab). Overall it is estimated that the reduced water quality has directly and indirectly led to a 70–80% loss of hard coral within the GBR lagoon over the past century. With respect to the proliferation of COTS, there is evidence that the nutrient enrichment leads to increases in the biomass of larger size classes of phytoplankton (i.e. nano-micro-phytoplankton > 2 μm) that provide the principal food source of the COTS larvae (Brodie et al. 2005). Significant increases in such phytoplankton populations have been recorded in the GBR lagoon near to Low Isles since the 1928–1929 GBR Expedition (Bell and Elmetri 1995; Bell et al. 2014a).
Although a considerable amount of scientific evidence was available that showed that the chronic eutrophic state of GBR waters had promoted outbreaks of COTS and the massive reduction in hard coral cover, a large portion of Australian marine science community and government agencies were for many years in a state of denial of the existence of the water quality problem (Bell and Gabric 1991; Bell 1992; Bell and Elmetri 1995; Risk 1999). The denial of the existence of the problem was supported by conclusions of the multimillion dollar ENCORE study conducted in the One Tree Island Lagoon (Larkum and Steven 1994; Koop et al. 2001). A principal conclusion of ENCORE was that the addition of nutrients did not promote the growth of attached algae and hence would not promote the conversion from coral dominated communities to algal dominated communities. However, this conclusion is not supported by the fact that attached algal growth was prolific on the walls and crests of the tested micro-atolls by the end of ENCORE (Bell et al. 2007). This incorrect conclusion not only lead to a lack of action to control the nutrient loads from diffuse and point source discharges to the GBR but it also supported the proliferation of bad science and management practices overseas (Hughes et al. 1999; Szmant 2002). However it is now generally accepted by the marine science community and various government agencies that the ecology of the GBR region is suffering from the chronic effects of eutrophication brought about by the discharge of nutrients from the developed catchments (Brodie and Waterhouse 2012; Waterhouse et al. 2017). In particular it is now generally accepted that excessive algal growth hinders the recovery of corals following damage (e.g. due to storms and bleaching events) and promotes the proliferation of COTS. This recognition led to the Australian Government Reef Rescue Initiative (RRI) in 2008 (Brodie and Waterhouse 2012) which has led to the Reef Water Quality Protection Plan (RWQPP) (RWQPP 2014).
The RRI led to the provision of multi-million dollar packages for various reef restoration programs; as part of these programs significant action on the management of nutrient loads in the riverine discharges has been initiated and a large number of associated research programs are being supported. The current water quality improvement programs are based on the assumption that phytoplankton growth and in particular the growth of the nano-micro-phytoplankton that promote the growth of COTS, is nitrogen (N) limited and hence a principal aim of the program is to reduce the riverine-loads of N and particularly the loads of dissolved inorganic nitrogen (DIN = NH4++NO -3 +NO2−). However there is no program to investigate the N loads introduced by Trichodesmium spp. and other N-fixing organisms even though early work showed such inputs could be significant (Bell and Elmetri 1995; Bell et al. 1999). Available evidence suggests that the magnitude of the Trichodesmium blooms has increased significantly since the 1928–1929 GBR Expedition (Bell and Elmetri 1995; Bell et al. 1999) and a recent study by Blondeau-Patissier et al. (2018) suggests that the growth of Trichodesmium in the Southern GBR has increased significantly since 2002. It is well known that large surface accumulations of Trichodesmium occur regularly during calmer/low wind periods (Revelante and Gilmartin 1982; Jones 1992; Blondeau-Patissier et al. 2018). Such large surface accumulations can extend for 100s to 1000s of kilometres.
Importance of N-loads from Trichodesmium
Calculations based on the distribution and the measured N-fixation rates of Trichodesmium spp. in the GBR lagoon showed that the N load introduced by Trichodesmium is probably similar to or even greater than that entering via riverine discharges (Bell and Elmetri 1995; Bell et al. 1999) and hence could be a major source of DIN. Also the annual water quality/ecological (phytoplankton+zooplankton) study conducted by Revelante and Gilmartin (1982) shows that increases in DIN concentrations and blooms of other phytoplankton taxa occur following Trichodesmium blooms. The importance of these early findings has not generally been recognised within the Scientific Consensus (Bartley et al. 2017; Waterhouse et al. 2017) that is driving the current water quality improvement programs. However, some recent studies do support these early findings. For example the study by McKinna (2010) based on remotely sensed satellite data and observations of surface accumulations of Trichodesmium using a ship mounted spectrophotometer concludes the N input from Trichodesmium is similar to or even greater than that introduced by the rivers. Also Furnas et al. (2011) have estimated that the average pelagic cross-shelf N load from Trichodesmium to the Wet Tropical region is of the same order as that introduced by the rivers. However Furnas et al. (2011) did not consider the N load introduced by blooms of Trichodesmium. They used a relatively low average concentration of Trichodesmium in their estimations (~ 1000 trichomes L−1) which corresponds to measured values in the less impacted region off Low Isles (Bell and Elmetri 1995). The monitoring work carried out in waters off Townsville (Revelante and Gilmartin 1982; Bell et al. 1999) suggest a much higher value should be used in the Wet Tropical region and hence the N load from the N-fixation by Trichodesmium would be far greater than that given by Furnas et al. (2011). A recent study by Messer et al. (2017) suggests that there is a need to include biological N-fixation as a significant source of N within GBR waters and Erler et al. (2020) argue that 15N/14N isotopic analyses in coral cores suggest that the N-load introduced by N-fixation has increased significantly over the past century or so. Also Blondeau-Patissier et al. (2018) note that their results derived from satellite imagery suggest that new N introduced to the GBR by Trichodesmium could be of the same order as that entering via catchment runoff.
Satellite remote sensing of Trichodesmium
It has been known for some time that surface accumulations of Trichodesmium that occur during calmer/low wind periods exhibit a high reflectance signature over the visible and near-infrared (NIR) wavelengths (e.g. see Borstad et al. 1992) which is clearly visible from space. Kuchler and Jupp (1988) demonstrated the potential use of this high reflectance phenomenon with a space shuttle photograph that captured a massive bloom in the GBR lagoon. This high reflectance phenomenon has been used by several workers to develop satellite remote-sensing algorithms to monitor Trichodesmium on a large scale. For example, Dupouy et al. (1988) presented a Nimbus-7 Coastal Zone Color Scanner (CZCS) image of a 90 000 km2 Trichodesmium bloom in the southwest Pacific Ocean near New Caledonia; they estimated that this single bloom could fix 3600 tonnes N in 10 days. Borstad et al. (1992) and Dupouy (1992) developed reflectance-based models applicable to surface accumulations of Trichodesmium recorded by the CZCS images. Also Subramaniam et al. (1999) used the high reflectance feature of Trichodesmium in conjunction with sea truth data and data from the advanced very high resolution radiometer (AVHRR) to map a 300 000 km2 Trichodesmium bloom off the Somali Coast in May 1995. They estimated that the N fixed by this bloom was 940 tonnes day−1.
A number of studies utilizing this high reflectance phenomenon have been carried out in the GBR region. McKinna et al. (2011) report results from a classification algorithm that uses high resolution data from the Moderate Resolution Imaging Spectroradiometer (MODIS). The classification algorithm was tested on a subset of MODIS images with corresponding sea-truth data. The algorithm was found to be robust in the presence of highly reflective, potential confounding effects such as shallow bathymetry and high suspended sediments. Tran (2015) used the McKinna algorithm to map the existence of dense Trichodesmium blooms following dust storm events using MODIS-derived images. His results imply that the dust storm events induce blooms of Trichodesmium over large regions of the southern GBR lagoon suggesting that the dust deposition could be a source of necessary trace elements. However the application of McKinna’s algorithm requires a significant amount of manipulation of the MODIS L0 data and the method ignores the use of negative remote sensing reflectance (Rrs) readings, which as discussed below, can be a good indicator of pixels with a high reflectance.
The first systematic analysis of spatial and temporal occurrences of Trichodesmium in the GBR was carried out by Blondeau-Patissier et al. (2018) using satellite data from the decade-long (now non-operational) MERIS ocean colour mission. Surface accumulations of Trichodesmium were detected using the Maximum Chlorophyll Index (MCI) which is based on reflectance of NIR wavelengths. The method was tested on a set of images coincident with available field data; the field data were only qualitative in nature i.e. observations of the absence or presence of Trichodesmium. The results of Blondeau-Patissier et al. (2018) show that large surface accumulations > 10 000 km2 occur in the GBR lagoon and that larger accumulations and more frequent accumulations occur in the more eutrophic Central and Southern GBR. The results from their time series analysis show there has been an increase in bloom frequency of Trichodesmium over the 10-year period (2002–2012), suggesting that the N load resulting from the blooms has also increased over the same period.
The high reflectance signature over the Red-Edge and NIR wavelengths can interfere with satellite remote sensing algorithms because the usual methods for estimating the atmospheric correction factor assume that the water leaving radiance is zero at these longer wavelengths; this assumption can lead to erroneous and even negative values of Rrs in the visible range (Hu et al. 2010; Rousset et al. 2018). McKinna (2010) ignored negative Rrs values in development of the Trichodesmium-detection algorithm whereas Rousset et al. (2018) used the existence of negative values of Rrs (678), as determined by SeaDAS processing of the MODIS L2 data (https://seadas.gsfc.nasa.gov/downloads), as a principal feature for detecting surface accumulations of Trichodesmium. They also suggest that the absolute value of Rrs (678) could be used to determine the relative concentration of Trichodesmium .
In the present study the presence of surface accumulations of Trichodesmium is assumed, as was done by Rousset et al. (2018), in regions where the SeaDAS analysis of MODIS L2 data shows Rrs(678) < 0. In addition the developed algorithm evaluates the use of negative values of Rrs(645), Rrs(555) and Rrs(547) as a measure of the concentration of Trichodesmium in the surface accumulations. The method uses the standard SeaDAS analysis of the MODIS L2 data and hence can be used by workers not familiar with detailed analysis of remote sensing data.
Model development
Results from experimental and modelling studies (Borstad et al. 1992) suggest algorithms based on reflectance phenomena should be able to detect Trichodesmium filament concentrations > 100 000 filaments L−1 with corresponding chlorophyll a (Chl a) concentrations ~ 10 mg m−3 (Borstad 1978). Borstad et al. (1992) showed that the magnitude of the reflection signal in the Red-Edge-NIR region increased with the concentration of Trichodesmium filaments and hence one would expect that the error introduced by the normal atmospheric correction methods would also increase with the concentration of Trichodesmium filaments and hence these higher concentrations could lead to negative Rrs values at shorter wavelengths. In the present study the presence of Trichodesmium is assumed to occur, as was done by Rousset et al. (2018), in regions where Rrs(678) < 0. In addition an algorithm that includes negative values of Rrs(645), Rrs(555) and Rrs(547) is evaluated to test the possibility of measuring increased levels of reflectance and hence increased concentrations of Trichodesmium. Four levels of concentration are included in this classification algorithm with the Level 1 classification corresponding to the lowest concentration of Trichodesmium within the surface accumulation and Level 4 classification corresponding to the highest concentration in the surface accumulation. Application of this algorithm is achieved by simply adding the following Masking Criteria in the standard SeaDAS analysis of the MODIS-Aqua images: Level 1: Rrs(678) < 0 and Rrs(645) > 0 and Rrs(555) > 0 and Rrs(547) > 0; Level 2: Rrs(678) < 0 and Rrs(645) < 0 and Rrs(555) > 0 and Rrs(547) > 0; Level 3: Rrs(678) < 0 and Rrs(645) < 0 and Rrs(555) < 0 and Rrs(547) > 0; Level 4: Rrs(678) < 0 and Rrs(645) < 0 and Rrs(555) < 0 and Rrs(547) < 0.
A number of MODIS-Aqua images that have been previously identified by other workers as blooms of Trichodesmium were chosen to evaluate the above algorithm. Use of this approach is not specific for identifying blooms of Trichodesmium; its use is restricted to estimating the extent/concentration of field verified Trichodesmium blooms.
Application of Model
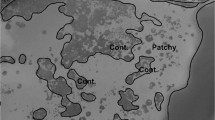
Figure 1 shows the application of Level 1 and Level 2 classifications of the MODIS-Aqua Chl a image examined by Rousset et al. (2018) in the Noumea region which they concluded was a bloom of Trichodesmium. The image shows a close resemblance to that presented by Rousset et al. (2018) but in addition the results suggest there are regions of higher (Level 2) and lower (Level 1) surface concentrations of Trichodesmium.

Application of Level 1 and Level 2 Classifications of MODIS-Aqua (Chl a) image (5 March 2015) in the South Pacific Noumea region
Figure 2 shows the application of Level 1 to Level 4 classifications of a SeaDAS RGB MODIS-Aqua Image (using default RGB-NASA MODIS L2 wavebands 667–531–443 nm with Turbidity Mask activated) of the Capricorn Channel region of the GBR that was examined for the large scale distribution of Trichodesmium by McKinna et al. (2011) and Rousset et al. (2018). The results are in general agreement with those of McKinna et al. (2011) and Rousset et al. (2018) but in addition the results suggest quite a distinctive variation from low to high concentrations from the outer to inner regions of the bloom formation. A number of black pixels in the inner region were denoted as indeterminate values by SeaDAS. It is suggested that, assuming these pixels are not associated with cloud cover, these indeterminate values are most probably due to very high reflectance values promoted by surface mats of Trichodesmium.

Application of Level 1 to Level 4 Classifications of a RGB MODIS-Aqua image (17 Oct 2007) of the Capricorn Channel region of the GBR
Figure 3 shows the application of Level 1 and Level 2 classifications of a SeaDAS RGB MODIS-Aqua image of the West Coast of Florida region that was identified by Hu et al. (2010) as probably being a Trichodesmium bloom. A number of black pixels in the inner region of this image were denoted as indeterminate values by SeaDAS. Again it is suggested that these indeterminate values are most probably due to very high reflectance values promoted by surface mats of Trichodesmium.

Application of Level 1 and Level 2 Classifications of RGB MODIS-Aqua Image (22 May 2004) off the West Coast of Florida
Figure 4 shows the application of Level 1 to Level 4 classifications of the RGB MODIS-Aqua Image (with Turbidity Mask activated) of the Central GBR region that was identified (Riebeek 2011) as probably being a significant Trichodesmium bloom. The results suggest quite a distinctive variation from low to high concentrations from the outer to inner regions of the bloom formation. The black pixels in the inner region of this very clear image probably represent surface mats of Trichodesmium.

Application of Level 1 to Level 4 Classifications of RGB MODIS-Aqua Image (9 August 2011) in the Townsville to Mackay GBR region
Figure 5 shows the application of Level 1 and Level 2 classifications of the RGB MODIS-Aqua image (with Turbidity Mask activated) in the Whitsunday/Mackay GBR region which was identified by McKinna et al. (2011) as a Trichodesmium bloom that followed a river flood plume event. This analysis shows a distribution pattern of Trichodesmium similar to that derived by McKinna et al. (2011) but in addition suggests that higher concentrations occur towards the inner bloom regions. McKinna (2010) notes that dense surface aggregations of Trichodesmium were reported during sampling of the flood plume on the previous day; a surface sample was analysed and yielded a Chl a concentration of ~ 2300 mg m−3. The results in Fig. 5 show that high suspended sediment, coloured dissolved and detrital matter associated with the flood plume are not misclassified as Trichodesmium by the present model.

Application of Level 1 and Level 2 Classifications of RGB MODIS-Aqua image (28 January 2005) in the Whitsunday/Mackay GBR region
Figure 6 shows the application of Level 1 to Level 4 classifications of a RGB MODIS-Aqua image (with Turbidity Mask activated) in the Central GBR region. This image shows that the bloom is quite extensive and again, that the nearshore waters which are generally characterised by high suspended sediment and coloured dissolved and detrital matter are not misclassified as Trichodesmium. McKinna (2010) identified extensive surface accumulations of Trichodesmium in the Southern region of this image. The area covered by this bloom is estimated to be ~ 10 000 km2.

Application of Level 1 to Level 4 Classifications of RGB MODIS-Aqua image (31 July 2008) in the Central GBR region
Estimated N-Loads from Trichodesmium
The above images show that the significant reflectance signal recorded by the satellite data is useful in estimating the aerial extent of the surface accumulations of the Trichodesmium and possibly a range of concentrations within those accumulations. However the surface accumulations can be notoriously patchy at all scales; very high concentrations at the surface and probably not very deep. In order to estimate the probable impact of such accumulations on water quality we require an estimate of the N content of the Trichodesmium, the proportion of that N content that is fixed by the Trichodesmium, some measurement of average volumetric concentration of the Trichodesmium and the depth variation of that concentration. The volumetric concentration is usually determined by microscopic counts (e.g., filaments/trichomes per litre and average cells per filament/trichome) and/or by associated Chl a measurements. Few such data are available for the GBR, far more data are available for other regions around the world. For example Devassy et al. (1978) measured bloom Chl a concentrations ~ 500 mg m−3 in the top 5 cm of the water column and Karl et al. (1992) measured an average Chl a concentration ~ 1100 mg m−3 in surface accumulations. McKinna (2010) notes that the Trichodesmium accumulations in the GBR were generally concentrated in a layer 1–2 m deep and that the measured Chl a concentrations in two grab samples were 57 and 315 mg m−3. McKinna (2010) reports that a grab sample of the surface accumulation depicted in Fig. 5 had a Chl a concentration of ~ 2300 mg m−3. If this were concentrated in the top 5 cm layer, as found by Devassy et al. (1978), it would equate to a value ~ 115 mg Chl a m−3 over a 1 m mixed layer. The results in the above images show that there can be quite a range in reflectance and hence in the Trichodesmium biomass concentration in the surface accumulations. The results from the experimental reflectance studies and modelling studies of Borstad et al. (1992) show that Trichodesmium concentrations greater than 100 000–200 000 filaments L−1 (which based on measurements of Borstad (1978) corresponds to ~ 10–20 mg Chl a m−3) should produce a significant reflectance. These results suggest that the Level 1 classification in the above images could represent Chl a concentrations of the order of ~ 10–20 mg m−3 but this would need to be verified by field data collected during Trichodesmium bloom events.
For modelling purposes McKinna (2010) assumed the Trichodesmium was concentrated in the top 2 m layer and that the concentration was characterised by a Chl a value of 100 mg m−3. In the present work a more conservative approach is taken in order to estimate the N load associated with the recorded surface accumulations, namely a 1 m mixed layer with an average Chl a concentration of 20 mg m−3 is assumed to occur over the Level 1 to Level 4 regions in Fig. 6. In addition we assume a N:Chl a (w/w) ratio of 71:1 (Mague et al. 1977) and that the N content of the Trichodesmium has been supplied predominately by N-fixation. Using this approach we estimate that the N content and hence the new-N load introduced by the bloom in Fig. 6 is ~ 14 000 tonnes which is far greater than the estimated annual DIN load from riverine discharges entering the Central GBR lagoon (~ 4200 tonnes, Kroon et al. 2012). This new-N load is of the same order as the estimated N-fixation load based on 15N/14N analysis of coral cores (Erler et al. 2020) and the available data on the N-fixation rates and distribution of Trichodesmium in the GBR lagoon (Bell et al. 1999; McKinna 2010). The actual annual N load could be much higher than this estimated value because much of the N fixed by Trichodesmium is released as DIN to the water column during the exponential growth phase and during the more senescent surface accumulation phase (Mulholland and Bernhardt 2005). Also more than one bloom of Trichodesmium can occur annually (Revelante and Gilmartin 1982). It is essential that a more accurate estimate of the N load be determined; this will require the conducting of various field studies prior to and during bloom events.
Importance of Phosphorus and Trace Elements
Various studies have shown that the N-fixation rates and growth rates of Trichodesmium and other diazotrophs common to the GBR would be increased by relatively small increases in the background concentrations of reactive inorganic phosphorus (P-PO4), dissolved organic phosphorus (DOP) and trace elements such as iron (Fe) (Bell et al. 2005; Mulholland and Bernhardt 2005; Fu et al. 2005). The phosphorus and trace elements can be supplied by various sources including riverine discharges, waste-water discharges and dust deposition (Shaw et al. 2008; Tran 2015). Also on decay, the DOP utilized by Trichodesmium could be released as biologically available phosphorus (BAP) which could promote the growth of other phytoplankton which cannot use the available background DOP. Studies are required to evaluate if DOP in the GBR waters supports the growth of Trichodesmium and if the BAP released by Trichodesmium promotes the growth of other phytoplankton.
Discussion
The simple satellite algorithm developed in the present work should be useful for not only monitoring the spatial extent of Trichodesmium blooms but also for quantifying the concentration of those blooms provided enough coincident field data is obtained to help ‘calibrate’ the satellite imagery. The current research and monitoring plans need to be updated to more accurately assess the potential impacts of Trichodesmium and management plans need to be developed to reduce those impacts.
Shortcomings of Current Research and Monitoring Programs
The evidence that Trichodesmium blooms are probably providing a significant source of bioavailable nutrients to the GBR lagoon has been ignored by the current Scientific Consensus in establishing the RWQPP. Also the importance of loads of phosphorus and trace components that would promote the growth of Trichodesmium has not been recognised. Current management programs also ignore the evidence that Trichodesmium blooms could promote the bioavailability of heavy metals (Jones 1992) and could be a potential source of toxins in the ciguatera food chain (Kerbrat et al. 2010) and hence could be having a significant negative impact on the fisheries industry. A number of critical questions regarding the impacts of water quality on GBR ecosystems are posed in the RWQPP but no integrated water quality/ecological (phytoplankton + zooplankton) monitoring program is proposed for answering these questions. Such a program similar to that undertaken by Revelante and Gilmartin (1982) and Ikeda et al. (1980) would be useful for answering many of the posed questions. For example as shown by other workers (Revelante and Gilmartin 1982; Bell 1992; Jones 1992; Bell et al. 1999; Bell et al. 2014a and b) data from such an integrated water quality/ecological monitoring program would provide valuable information on:
-
the relative importance of various sources of nutrients e.g. riverine discharges, wind-resuspension of sediments, dust storms, upwellings and growth of Trichodesmium;
-
the links between the growth of Trichodesmium and the production of bioavailable toxins;
-
the identification of probable links between eutrophication and the proliferation of corallivores (e.g. COTS and Drupella spp.), jelly-fish, coral skeletal diseases and coral bleaching.
Proposed Action Plan
I recommend that an integrated water quality/ecological monitoring program be established at three cross-shelf locations (e.g. Northern Transect off Port Douglas; Central Transect off Townsville; Southern Transect off Gladstone). In addition:
-
Satellite imagery should be used to provide information on the frequency and spatial extent of the Trichodesmium blooms and various insitu monitoring methods should be employed to help quantify the loads of nutrients and toxins associated with such blooms.
-
The sources of the principal nutrient/trace components that promote the growth of Trichodesmium need to be identified and management programs should be implemented to reduce the loads of such components.
-
The potential benefits of harvesting Trichodesmium from the concentrated surface accumulations, as has been proposed for cyanobacterial blooms in the Baltic Sea (Gröndahl 2009), should be investigated. The harvested algae could have a significant monetary value (e.g. as a source of bio-fertilizers, biofuels and/or fine-chemicals) that could more than offset the cost of harvesting. This proposed “Cleaning the Fishpond” program has the potential to have immediate positive effects on the environment by providing a significant reduction in both the nutrient loads and the resultant promotion of algal blooms. Also, if it is demonstrated that Trichodesmium blooms are promoting the bioavailability of heavy metals and/or toxins in the ciguatera food chain, then harvesting of the Trichodesmium would provide valuable assistance to the fisheries industry.
References
Bartley, R., D. Waters, R. Turner, F. Kroon, S. Wilkinson, A. Garzon-Garcia, P. Kuhnert, S. Lewis, et al. 2017. Scientific Consensus Statement 2017: A synthesis of the science of land-based water quality impacts on the Great Barrier Reef, Chapter 2: Sources of sediment, nutrients, pesticides and other pollutants to the Great Barrier Reef. State of Queensland, 2017.
Bell, P.R.F. 1992. Eutrophication and coral reefs-some examples in the Great Barrier Reef lagoon. Water Research 26: 553–568.
Bell, P.R.F., and I. Elmetri. 1995. Ecological Indicators of Large Scale Eutrophication in the Great Barrier Reef (GBR) Lagoon. Ambio 24: 208–215.
Bell, P.R.F., and A.J. Gabric. 1991. Must GBR pollution become chronic before management reacts? Search. 22: 117–119.
Bell, P.R.F., I. Elmetri, and P. Uwins. 1999. Nitrogen fixation of Trichodesmium spp. in the Great Barrier Reef Lagoon-importance to the overall nitrogen budget. Marine Ecology Progress Series 186: 119–126.
Bell, P.R.F., P.J.R. Uwins, I. Elmetri, J.A. Phillips, F. Fu, and A.J.E. Yago. 2005. Laboratory culture studies of Trichodesmium isolated from the Great Barrier Reef Lagoon, Australia. Hydrobiologia 532: 9–21.
Bell, P.R.F., B.E. Lapointe, and I. Elmetri. 2007. Reevaluation of ENCORE: support for the eutrophication threshold model for coral reefs. Ambio 36: 418–424.
Bell, P.R.F., I. Elmetri, and B.E. Lapointe. 2012. Synoptic Scale Monitoring supports the coral reef eutrophication threshold model. 12th International coral reef symposium. Cairns, Australia. http://www.icrs2012.com/eposters/P227.pdf
Bell, P.R.F., I. Elmetri, and B.E. Lapointe. 2014a. Evidence of large-scale chronic eutrophication in the Great Barrier Reef: Quantification of Chlorophyll a thresholds for sustaining Coral Reef Communities. Ambio 43: 361–376.
Bell, P.R.F., I. Elmetri, and B.E. Lapointe. 2014b. Response to “selective evidence of eutrophication in the Great Barrier Reef” by Furnas et al. Ambio 43: 379–380.
Blondeau-Patissier, D., V.E. Brando, C. Lønborg, S.M. Leahy, and A.G. Dekker. 2018. Phenology of Trichodesmium spp blooms in the Great Barrier Reef lagoon, Australia, from the ESA-MERIS 10-year mission. PLoS ONE 13: e0208010. https://doi.org/10.1371/journal.pone.0208010.
Borstad, G.A. 1978. Some aspects of the occurrence and biology of Trichodesmium (cyanophyta) in the Western tropical Atlantic near Barbados, West Indles. Ph. D. Thesis. Marine Science Centre McGill University, Montreal, Quebec, Canada.
Borstad, G.A., J.F.R. Gower, and E.J. Carpenter. 1992. Development of algorithms for remote sensing of Trichodesmium spp. blooms. In Marine pelagic cyanobacteria: Trichodesmium spp. and other Diazotrophs, ed. E.J. Carpenter, D.G. Capone, and J.G. Reuter, 193–210. Dordrecht: Kluwer Academic Publishers.
Brodie, J., K. Fabricius, G. Death, and K. Okaji. 2005. Are increased nutrient inputs responsible for more outbreaks of crown-of-thorns starfish? An appraisal of the evidence. Marine Pollution Bulletin 51: 266–278.
Brodie, J., and J. Waterhouse. 2012. A critical review of environmental management of the ‘not so Great’ Barrier Reef. Estuarine Coastal and Shelf Science 104–105: 1–22.
Devassy, V.P., P.M.A. Bhattathiri, and S.Z. Qasim. 1978. Trichodesmium spp. Phenomenon. Indian Journal of Marine Science 7: 168–186.
Dupouy, C. 1992. Discoloured waters in the Melanesian Archipelago (New Caledonia and Vanuatu). The value of the Nimbus-7 Coastal Zone Colour Scanner observations. In Marine pelagic cyanobacteria: Trichodesmium spp. and other Diazotrophs, ed. E.J. Carpenter, D.G. Capone, and J.G. Reuter, 177–191. Dordrecht: Kluwer Academic Publishers.
Dupouy, C., M. Petit, and Y. Dandonneau. 1988. Satellite detected cyanobacteria bloom in the southwestern tropical Pacific: Implications for oceanic nitrogen fixations. International Journal of Remote Sensing 9: 389–396.
Erler, D.V., H.T. Farid, T.D. Glaze, N.L. Carlson-Perret, and J.M. Lough. 2020. Coral skeletons reveal the history of nitrogen cycling in the coastal Great Barrier Reef. Nature Communications 11: 1500. https://doi.org/10.1038/s41467-020-15278-w.
Fu, F., Y. Zhang, P.R.F. Bell, and D. Hutchins. 2005. Phosphate uptake and growth kinetics of Trichodesmium spp. (Cyanobacteria) isolates from the North Atlantic Ocean and the Great Barrier Reef, Australia. Journal of Phycology 41: 62–73.
Furnas, M., D. Alongi, D. McKinnon, L. Trott, and M. Skuza. 2011. Regional-scale nitrogen and phosphorus budgets for the northern (141oS) and central (171oS) Great Barrier Reef shelf ecosystem. Continental Shelf Research 31: 1967–1990.
Gröndahl, F. 2009. Removal of surface blooms of the Cyanobacteria Nodularia spumigena: A pilot project conducted in the Baltic Sea. Ambio 38: 79–84.
Hu, C., J. Cannizzaro, K.L. Carder, F.E. Muller-Karger, and R. Hardy. 2010. Remote detection of Trichodesmium blooms in optically complex coastal waters: Examples with MODIS full-spectral data. Remote Sensing of Environment 114: 2048–2058.
Hughes, T.P., A.M. Szmant, R. Steneck, R. Carpenter, and S. Miller. 1999. Algal blooms on coral reefs: What are the causes? Limnology and Oceanography 44: 1583–1586.
Ikeda, T., M. Gilmartin, N. Revelante, A.W. Mitchell, J.H. Carleton, P. Dixon, S.M. Hutchinson, E. Hing Fay, et al. 1980. Biological, chemical and physical observations in inshore waters of the Great Barrier Reef, North Queensland. Aust. Inst. of Mar. Sci. Tech. Bull. Oceanography Series No. 1. AIMS-OS-80-1. Aust. Inst. Mar. Sci., Cape Ferguson.
Jones, G.B. 1992. Effect of Trichodesmium blooms on water quality in the Great Barrier Reef Lagoon. In Marine pelagic cyanobacteria: Trichodesmium spp. and other Diazotrophs, ed. E.J. Carpenter, D.G. Capone, and J.G. Reuter, 273–285. Dordrecht: Kluwer Academic Publishers.
Karl, D.M., R. Letelier, D.V. Hebel, D.F. Bird, and C.D. Winn. 1992. Trichodesmium blooms and new nitrogen in the North Pacific gyre. In Marine pelagic cyanobacteria: Trichodesmium spp. and other Diazotrophs, ed. E.J. Carpenter, D.G. Capone, and J.G. Reuter, 218–237. Dordrecht: Kluwer Academic Publishers.
Kerbrat, A., H. Darius, S. Pauillac, M. Chinain, and D. Laurent. 2010. Detection of ciguatoxin-like and paralysing toxins in Trichodesmium spp. from New Caledonia lagoon. Marine Pollution Bulletin 61: 360–366.
Koop, K., D. Booth, A. Broadbent, J. Brodie, D. Bucher, D. Capone, J. Coll, W. Dennison, et al. 2001. ENCORE: The effect of nutrient enrichment on coral reefs. Synthesis of results and conclusions. Marine Pollution Bulletin 42: 91–120.
Kroon, F.J., P.M. Kuhnert, B.L. Henderson, S.N. Wilkinson, A. Kinsey-Henderson, B. Abbott, J.E. Brodie, and R.D.R. Turner. 2012. River loads of suspended solids, nitrogen, phosphorus and herbicides delivered to the Great Barrier Reef lagoon. Marine Pollution Bulletin 65: 167–181.
Kuchler, D., and D.L.B. Jupp. 1988. Shuttle photograph captures massive phytoplankton bloom in the Great Barrier Reef. International Journal of Remote Sensing 9: 1229–1301.
Larkum, A.W.D., and A.D.L. Steven. 1994. ENCORE: The effect of nutrient enrichment on coral reefs. 1. Experimental design and research programme. Marine Pollution Bulletin 29: 112–120.
Mague, T.H., F.C. Mague, and O. Holm-Hansen. 1977. Physiology and chemical composition of nitrogen-fixing phytoplankton in the Centarl Pacific Ocean. Marine Biology 41: 213–227.
McKinna, L.I.W. 2010. Optical detection and quantification of Trichodesmium spp. within Great Barrier Reef. PhD thesis. Townsville Australia: James Cook University.
McKinna, L.I.W., M.J. Furnas, and P.V. Ridd. 2011. A simple binary classification algorithm for detecting Trichodesmium spp. within the Great Barrier Reef using MODIS imagery. Limnology and Oceanography: Methods 9: 50–66.
Messer, L.F., M.V. Brown, M.J. Furnas, R.L. Carney, A.D. McKinnon, and J.R. Seymour. 2017. Diversity and activity of diazotrophs in Great Barrier Reef surface waters. Frontiers in Microbiology 8: 967. https://doi.org/10.3389/fmicb.2017.00967.
Mulholland, M.R., and P.W. Bernhardt. 2005. The effect of growth rate, phosphorus concentration, and temperature on N2 fixation, carbon fixation, and nitrogen release in continuous cultures of Trichodesmium IMS101. Limnology and Oceanography 50: 839–849.
Riebeek, H. 2011. https://earthobservatory.nasa.gov/images/51757/phytoplankton-bloom-in-the-great-barrier-reef.
Reside, A.E., J. Beher, A.J. Cosgrove, M.C. Evans, L. Seabrook, J.L. Silcock, A.S. Wenger, and M. Maron. 2017. Ecological consequences of land clearing and policy reform in Queensland. Pacific Conservation Biology 23: 219–230. https://doi.org/10.1071/PC17001.
Revelante, N., and M. Gilmartin. 1982. Dynamics of phytoplankton in the Great Barrier Reef lagoon. Journal of Plankton Research 4: 47–76.
Risk, M.J. 1999. Paradise lost: How marine science failed the world’s coral reefs. Marine and Freshwater Research 50: 831–837.
Rousset, G., F. De Boissieu, C.E. Menkes, J. Lefèvre, R. Frouin, M. Rodier, V. Ridou, S. Laran, et al. 2018. Remote sensing of Trichodesmium spp. mats in the western tropical South Pacific. Biogeosciences 15: 5203–5219. https://doi.org/10.5194/bg-15-5203-2018.
RWQPP. 2014. Australian Government 2014. Reef Water Quality Protection Plan 2013-Priortisation Project Report, Canberra ISBN 978-1-7600307-3-5.
Shaw, E.C., A.J. Gabric, and G.H. McTainsh. 2008. Impacts of aeolian dust deposition on phytoplankton dynamics in Queensland coastal waters. Marine and Freshwater Research 59: 951–962.
Subramaniam, A., E.J. Carpenter, and P.G. Falkowski. 1999. Bio-optical properties of the marine diazotrophic cyanobacteria Trichodesmium spp. II. A reflectance model for remote sensing. Limnology and Oceanography 44: 618–627.
Szmant, A. 2002. Nutrient enrichment on coral reefs: is it a major cause of coral reef decline? Estuaries 25: 743–766.
Tran, D.V. 2015. Factors Influencing the spatiotemporal variability in satellite-derived Chlorophyll on the Queensland continental shelf. PhD Thesis. Brisbane: Griffith University. http://hdl.handle.net/10072/366428.
Waterhouse, J., B. Schaffelke, R. Bartley, R. Eberhard, J. Brodie, M. Star, P. Thorburn, J. Rolfe et al. 2017. 2017 Scientific Consensus Statement: A synthesis of the science of land-based water quality impacts on the Great Barrier Reef, Chapter 5: Overview of key findings, management implications and knowledge gaps. State of Queensland, 2017. https://www.reefplan.qld.gov.au/science-and-research/the-scientific-consensus-statement.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Bell, P.R.F. Analysis of satellite imagery using a simple algorithm supports evidence that Trichodesmium supplies a significant new nitrogen load to the GBR lagoon. Ambio 50, 1200–1210 (2021). https://doi.org/10.1007/s13280-020-01460-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13280-020-01460-3