
Abstract
Evaluation of the immunomodulatory activity of plant compounds is an interesting and growing area of research. Teucrium ramosissimum Desf. is a native and endemic medicinal plant from the South of Tunisia traditionally used for the treatment of many diseases. The anti-inflammatory activity of apigenin-7-glucoside, genkwanin, and naringenin isolated from T. ramosissimum were assayed. The phagocytic activities of macrophage and lymphocyte proliferation were investigated in the absence and presence of mitogens (lipopolysaccharide [LPS] or lectin). Depending on the concentrations, the compounds affect macrophage functions by modulating their lysosomal enzyme activity and nitric oxide (NO) release. The tested compounds enhance significantly splenocyte proliferation, either with or without mitogen stimulation. In studies to assess any potential effects of apigenin-7-glucoside, genkwanin, and naringenin on innate immunity, the results showed that these compounds significantly enhanced the killing activity of natural killer (NK) cells and cytotoxic activity of the T lymphocyte (CTL) isolated from splenocytes. These results suggest that T. ramosissimum compounds such as apigenin-7-glucoside, genkwanin, and naringenin may be potentially useful for modulating immune cell functions in physiological and pathological conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A wide number of medicinal plants are characterized by immunomodulatory activities. Many plants used in traditional medicine are largely investigated for their potential use in the prevention against chronic disease. Modulating the immune system refers to any modification of the immune response that may involve induction, expression, amplification, or inhibition of a part or phase of the immune response.
Most clinically important medicines for treatment of inflammation-related diseases are steroidal or non-steroidal anti-inflammatory drugs. Although these drugs have potent activity, long-term administration is required for treatment of chronic disease. Furthermore, these drugs have various and severe adverse effects. Therefore, naturally occurring agents, with high effectiveness and very few side effects, are a preferable substituent for chemical therapeutics. The Teucrium genus, which belongs to the Lamiaceae family, has been instrumental in the discovery of medicinal natural products [1] and encompasses more than 300 species. About 50 of them are known in Europe and are distributed mainly in the Mediterranean basin [2]. Many species from the Teucrium genus show interesting biological properties, such as analgesic, hypoglycemic, hypolipidemic, antipyretic, anti-inflammatory, antioxidant, and antibacterial properties. However, little is known about the mechanisms responsible for their immune-regulating properties. Due to the use of the Teucrium ramosissimum genus in the food and drug industries [3] and the evidence of its biological protective activity, we were interested on how compounds extracted from T. ramosissimum can act on immune cell functions and by which mechanisms they exert their immunomodulatory effects. In the present study, our objective was to investigate the immunomodulatory, anti-inflammatory, and the antioxidant activities of compounds isolated from T. ramosissimum in splenocytes and macrophages.
Materials and methods
Plant material
The aerial part of T. ramosissimum was collected in January 2005 from the mountainous region of Gafsa in the Southeast of Tunisia. The plant was identified by Pr. Mohamed Chaieb (Department of Botany, Faculty of Sciences, University of Sfax, Tunisia) according to the Flora of Tunisia [4]. A voucher specimen (Tr-02-05) was deposited at the Herbarium of the Department of Pharmacognosy (Faculty of Pharmacy, University of Monastir, Tunisia) for future reference. The leaves were shade-dried, powdered, and stored in a tightly closed container for further use.
Preparation of plant compounds
Apigenin-7-glucoside, genkwanin, and naringenin were purified from chloroformic extract as described previously by Ben Sghaier et al. [19] from T. ramosissimum leaves. Before use, molecules were dissolved in roswell park memorial institute (RPMI)-1640 medium.
Cell preparations from mice
Specific pathogen-free male BALB/c mice (18–22 g) were obtained from the Pasteur Institute (Tunis, Tunisia). The mice were housed under standard conditions of temperature (22–28 °C), humidity (30–70 %), and light (12 h light/dark) in an accredited pathogen-free facility. All experiments were performed in accordance with the guidelines for the care and use of laboratory animals as published by the US National Institute of Health. All experiments received the explicit approval of the Ethics Animal Committee in Tunisia.
Spleen mice lymphocytes were obtained as previously reported [5]. Briefly, mice were euthanized by cervical dislocation, and each spleen was isolated aseptically and minced with a sterile forceps. Splenocytes were then isolated by centrifugation (1500 rpm, for 10 min), and any red blood cells present were lysed by resuspending the pellet in lysing buffer (144 mM NH4Cl, 1.7 mM Tris base) and placing on ice for 10 min. Cells were then washed twice with phosphate-buffered saline (PBS, pH 7.4) and then resuspended in complete RPMI medium (Gibco-BRL) containing 10 % fetal bovine serum (FBS; Gibco) and 100 mg/ml gentamycin (Gibco-BRL, Paisley, UK). Other mice were used to provide peritoneal macrophages as previously reported [5]. Peritoneal cells were obtained after intraperitoneal injection of 4 ml sterile PBS, massaging of the peritoneum, and drawing back of the fluid (≈4 ml) into the syringe. The obtained cells were washed twice and resuspended in complete RPMI-1640 medium. Cell viability was assessed using the trypan blue exclusion technique.
Splenocytes proliferation assay
Assays of lymphocyte proliferation were performed by using the 3-(4,5 dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) method outlined by Mosmann [6]. Splenocyte suspension is a mixture of cells containing T lymphocytes, B lymphocytes, and monocytes among others; after 24 h incubation, only lymphocytes proliferate. The splenocyte suspension in RPMI-1640 (5 × 106 cells/ml; 100 μl aliquot/well) was pre-incubated in 96-well plate for 24 h before the addition of mitogens (lectin [from Phytolacca Americana (PWM)] or lipopolysaccharide [LPS, rough strain, Escherichia coli EH100, Sigma, Hamburg, Germany]) at 5 mg/ml alone or in combination with increasing concentrations of apigenin-7-glucoside, genkwanin, or naringenin freshly solubilized in RPMI. The treated cells were then incubated at 37 °C in a humidified 5 % CO2 atmosphere for a further 48 h. Thereafter, the plates were centrifuged at 1500 rpm for 10 min, and the pellet in each well was resuspended in 50 μl of an MTT (5 mg/ml RPMI) solution and incubated for 4 h at 37 °C. After that, the plate was centrifuged, the MTT solution was removed from each well, and 100 μl dimethyl sulfoxide (DMSO; 98 %) added to the wells. After incubation at 37 °C for 15 min, the formation of formazan was evaluated by following its absorbance at 570 nm in a microplate reader (Thermo Scientific, Vantaa, Finland). The percentage of proliferation (relative to that of cells receiving neither lectin nor LPS) was ultimately calculated using the following equation: Proliferation (%) = 100 × (OD sample − OD control)/OD control [7].
Assay of natural killer (NK) cell activity
Natural killer (NK) cell activity was measured as previously described [8], with minor modification. Briefly, spleens prepared as described above were used as the source of effector cells; isolated splenocytes were seeded into 96-well microtiter plates at 5 × 106 cells/ml. The cells were then stimulated at 37 °C by different concentrations of apigenin-7-glucoside, genkwanin, or naringenin for 24 h. To eliminate the direct effects of the extract on target cells, spleens were washed once with RPMI-1640, then target K562 cells (5 × 104 cells/ml; yielding a 100:1 expected effector-target ratio) were added to each well in 100-μl aliquots. The plates were then incubated for 4 h at 37 °C in 5 % CO2 atmosphere. An aliquot (40 μl) of MTT solution (5 mg/ml) was then added to each well, and the plate was incubated a further 2 h. After that, the plate was centrifuged again, the MTT in each well was removed, and 100 μl of DMSO (98 %) was added. After incubation at 37 °C for 15 min, absorbance of formed formazan in each well was measured at 570 nm in a microplate reader (Thermo Scientific, Vantaa, Finland). Three kinds of control measurements were performed: target cell control, blank control, and effector cell control. NK cell activity was calculated as follows: NK activity (%) = 100 × (ODT − (ODS − ODE))/ODT, where ODT = optical density value of target cell control, ODS = optical density value of test samples, and ODE = optical density value of effector cell control.
Assays of cytotoxic T lymphocyte (CTL) activity
T lymphocyte (CTL) assay or cell-mediated cytotoxicity was performed using the MTT assay. Splenocyte suspension in RPMI-1640 medium (5 × 106 cells/ml; 100 μl aliquot/well) was pre-incubated in a 96-well plate for 24 h; then pre-treated with different concentrations of apigenin-7-glucoside, genkwanin, or naringenin for a supplementary 24 h; and then target B16-F10 melanoma cells (5 × 104 cells/ml; yielding a 100:1 expected effector-target ratio) were added to each well in 100-μl aliquots. The plates were then incubated for 24 h at 37 ° C in 5 % CO2 atmosphere. Spleens were used as the source of effector cells called CTLs.
CTL cell activity was calculated as follows: CTL activity (%) = 100 × (ODT − (ODS − ODE))/ODT, where ODT = optical density value of target cell control, ODS = optical density value of test samples, and ODE = optical density value of effector cell control.
Assessment of lysosomal enzyme activity
Lysosomal enzyme activity (reflected by acid phosphatase [AP] activity) in macrophages was measured as previously described [9], with some modification. Briefly, macrophage suspensions (100 μl aliquot of 6 × 106 cells/ml) were seeded into flat-bottom 96-well plates; treated with different concentrations of apigenin-7-glucoside, genkwanin, or naringenin; and incubated at 37 °C in a 5 % CO2 humidified atmosphere for 48 h. The medium was then removed and 20 μl of 0.1 % Triton X 100 (Sigma, St. Louis, MO), 100 μl of 100 mM p-nitrophenyl phosphate solution (Sigma), and 50 μl of citrate buffer (pH 5.0, 0.1 M) were added to each well. The plate was then incubated for 30 min at 37 °C before adding 150 μl of borate buffer (pH 9.8, 0.2 M) to each well and the absorbance then measured at 405 nm. The percentage of lysosomal enzyme activity in treated cells relative to that in control cells was calculated as previously reported [7]: Lysosomal enzyme activity (%) = 100 × (OD sample − OD control)/OD control.
Nitrite assay
Nitrite levels were measured using the diazotization method based on the Griess reaction, which is an indirect assay for nitric oxide (NO) production as previously described by Green et al. [10]. In brief, cells were incubated for 48 h in the presence of increasing concentrations of apigenin-7-glucoside, genkwanin, or naringenin. Nitrite was then measured by adding 100 μl Griess reagent (1 % sulfanilamide and 0.1 % naphthylenediamine in 5 % phosphoric acid) to 100 μl of harvested culture supernatant. The optical density at 570 nm (OD 570) was then measured in a microplate reader (Thermo Scientific, Vantaa, Finland). NO concentrations were calculated by comparison with the OD 570 of a standard solution of sodium nitrite diluted in culture medium and placed in parallel wells in the assay plates.
Cellular antioxidant activity (CAA) assay
A cellular antioxidant activity (CAA) assay, developed by Wolfe and Liu [11], was employed to measure the effects of the antioxidant potentials of apigenin-7-glucoside, genkwanin, and naringenin. Briefly, splenocytes were seeded at a density of 5 × 105 cells (in 100 μl PBS), in a 96-well microplate. Triplicate wells were then treated for 1 h with 10 μl of each tested molecule (concentrations ranging from 5 to 40 μM, for both apigenin-7-glucoside and genkwanin, and from 40 to 80 μM for the naringenin) along with 5 μl of a 25 μM solution of 2′,7′-dichlorofluorescein (DCFH). After this incubation, a 100-μl aliquot of 1.2 mM 2, 2’-azobis (2-amidino-propane) ABAP (Sigma Aldrich, Steinheim, Germany) in PBS was applied to the cells. The ABAP is an exogenous source of peroxyl radicals used to oxidize DCFH-DA to the fluorescent product dichlorofluorescein (DCF). Accordingly, cells treated with natural compounds that have any antioxidant activity should have lower fluorescence compared to untreated cells. The fluorescence in each well was read every 5 min for a total of 1 h in a fluorescence microplate reader (Biotek, Winooski, USA), using 538-nm emission and at 485-nm excitation filters. Each plate included triplicate control and blank wells: control wells contained cells treated with DCFH-DA and ABAP and blank wells contained cells with DCFH-DA and PBS without ABAP. Fluorescence values for the blank samples and initial fluorescence values were subtracted from the sample values. The area under the fluorescence vs. time curve was integrated at each time point to calculate the CAA units using the following equation:
Where ∫SA is the integrated area under the sample fluorescence vs. time curve and ∫CA is the integrated area from the control curve.
Statistical analysis
All data were expressed as mean (±SD) and compared using Student’s t test. Statistical significance was assigned at P values <0.05.
Results
Splenocyte proliferation
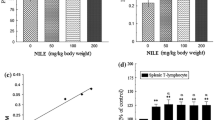
To assess whether apigenin-7-glucoside, genkwanin, and naringenin from T. ramosissimum can modulate cell proliferation, BALB/c splenocytes were cultured with tested molecules alone or with LPS (B lymphocyte mitogen) or lectin (T lymphocyte mitogen). In the absence of mitogen, apigenin-7-glucoside, genkwanin, and naringenin were able to induce splenocyte proliferation at tested concentrations in a dose-dependent manner (Fig. 1a). A higher rate of cell proliferation occurred when both tested molecules and lectin or LPS were incubated with a cell culture, in a dose-dependant manner (Fig. 1b, c).

In vitro effects of tested molecules from T. ramosissimum on the proliferation response of splenocytes. Cells were incubated for 48 h with the following: a cells were incubated with increasing concentrations of molecules without mitogens, b cells were incubated with lipopolysaccharide (LPS) (5 μg/ml) in the absence and the presence of molecules, and c cells were treated with lectin (5 μg/ml) in the absence and the presence of molecules. Control included cells incubated in RPMI-1640 medium. Cell proliferation was assessed by the MTT test. Data shown are mean percentage proliferation (±SD) from three independent experiments. The statistical significance of results was evaluated by Student’s t test: *a P < 0.05, **a P < 0.01, and ***a P < 0.001 mean that value is significantly different compared with that of negative (RPMI) control, and **b P < 0.01 and ***b P < 0.001 mean that the value is significantly different compared with that of mitogen (LPS or lectin)-treated cells
NK cell activity
NK cells are a major component of the innate immune system and play an important role in tissue inflammation. They can represent a first line of defense against pathogens [12]. Measures of the cytotoxic activity of splenic NK cells against NK-sensitive tumor cells (i.e., K562 cell line) revealed that, as compared with the control cells, T. ramosissimum compounds significantly enhanced NK cell activity at the different tested concentrations, especially with 5 and 10 μM of apigenin-7-glucoside and genkwanin and 10 μM of naringenin (Fig. 2). However, this activity appeared to decrease when cells were incubated with compound concentrations above 40 μM of apigenin-7-glucoside and genkwanin and 80 μM of naringenin.

Effect of apigenin-7-glucoside, genkwanin, and naringenin at various tested concentrations, on mice natural killer (NK) cells in vitro. Splenocytes were cultured with different doses of molecules for 24 h before K562 target cells were added (at expected 100:1 E:T ratio). NK activity was then measured using an MTT assay. Values shown are mean (±SD) % cytotoxicity from six different observations. The statistical significance of results was evaluated by Student’s t test: ***P < 0.001 means that the value is significantly different compared with that of control cells
CTL activity
There is now ample evidence that cytotoxic T lymphocytes can be key effector cells involved in the immune-mediated destruction of virally infected and cancer cells [13]. This cell-contact-dependent cytotoxicity is a hallmark of both cytotoxic T cells and NK cells. We therefore tested the effect of T. ramosissimum compounds on the cytotoxic activities of T lymphocytes against B16F10 melanoma cells. Measures of CTL activity revealed that tested molecules enhanced CTL activity in a dose-dependent manner with a maximum potency of 51.35, 60.49, and 70.23 %, respectively, at the higher tested concentration of apigenin-7-glucoside, genkwanin, and naringenin (Fig. 3).

Effect of apigenin-7-glucoside, genkwanin, and naringenin at various tested concentrations, on mice cytotoxic T lymphocyte (CTL) cells in vitro. Splenocytes were cultured with different doses of molecules for 24 h before B16F10 target cells were added (at expected 100:1 E:T ratio). CTL activity was then measured using an MTT assay. Values shown are mean (±SD) % cytotoxicity from six different observations. The statistical significance of results was evaluated by Student’s t test: ***P < 0.001 means that the value is significantly different compared with that of control cells
Peritoneal macrophage lysosomal activity
The cellular lysosomal activity in mouse macrophages was assessed in response to apigenin-7-glucoside, genkwanin, and naringenin treatments. As shown in Fig. 4, all tested molecules markedly affect macrophage lysosomal enzyme activity in a dose-dependent manner. Inhibition of the activity of macrophage lysosomal function by the tested compounds was expressed by comparison to the control cells.

Stimulation of mouse peritoneal macrophage lysosomal enzyme activity by apigenin-7-glucoside, genkwanin, and naringenin. Macrophages (2 × 106 cells/well) were incubated in the presence of increasing concentrations of molecules for 48 h. Control cells were included with RPMI-1640 medium. Lysosomal enzyme activity was assessed as indicated in the “Materials and methods” section. Data shown are mean (±SD) percentages of lysosomal enzyme activity from three independent experiments. ***P < 0.001 means that the value is significantly different compared with that of control cells
Peritoneal macrophage NO production
NO production was monitored in macrophages stimulated by LPS in the presence or absence of increasing doses of apigenin-7-glucoside, genkwanin, or naringenin during the 48-h incubation at 37 °C, 5 % CO2. We used the Griess assay to measure the accumulation of nitrite, a stable oxidized product of NO, in culture media. As shown in Fig. 5, 5 μg/ml LPS caused a dramatic increase of NO production by 53.37 μM. This induction was inhibited by the tested molecule treatment in a dose-dependent manner. In fact, NO production was reduced from 53.37 μM in control cells to 22.33, 20.66, and 28.64 μM at the highest tested concentrations of apigenin-7-glucoside, genkwanin, and naringenin, respectively.

Production of the nitrite by mouse peritoneal macrophages in response to apigenin-7-glucoside, genkwanin, and naringenin. Macrophages (2 × 106 cells/well) were incubated in RPMI-1640 medium in the absence or in the presence of increasing concentrations of molecules for 48 h. Cells treated with 5 μg/ml LPS alone were used as positive control. Data shown are mean (±SD) percentage of NO production from three independent experiments. The statistical significance of results was evaluated by Student’s t test: ***a P < 0.001 means that the value is significantly different compared with that of negative (RPMI) control and **b P < 0.01 and ***b P < 0.001 mean that the value is significantly different compared with that of mitogen (LPS)-treated cells
Cellular antioxidant activity (CAA)
A cellular antioxidant activity (CAA) assay was conducted to quantify the intracellular antioxidant property of T. ramosissimum compounds by preventing the formation of dichlorofluorescein (DCF) using ABAP-induced peroxyl radicals in splenocyte. For the CAA assay, decreasing the cellular fluorescence of tested molecules compared to the control indicates antioxidant capacity, because the level of fluorescence is proportional to the degree of oxidation [11]. The results show that apigenin-7-glucoside, genkwanin, and naringenin could be absorbed into splenocytes and exert antioxidant activity. The mean effective concentration (EC50) was calculated from the dose-response curve. Our result revealed that incubation of splenocytes with different concentrations ranging from 5 to 40 μM for apigenin-7-glucoside and genkwanin, and from 10 to 80 μM for naringenin, reduced DCF production in a dose-dependent manner (Fig. 6). As displayed in Fig. 7, the EC50 value of peroxyl radical-inducing DCFH oxidation inhibition by the genkwanin was 7.5 μM in splenocytes.

Kinetics of inhibited oxidation. The kinetics of the inhibition of oxidation of DCFH in splenocytes using a cellular antioxidant activity assay in the presence of apigenin-7-glucoside (a), genkwanin (b), and naringenin (c)

Dose-response curve for inhibition of oxidation. Dose-response curve for oxidation of DCFH to DCF in splenocytes using a cellular antioxidant activity assay in the presence of apigenin-7-glucoside (a), genkwanin (b), and naringenin (c); (±SD, n = 3)
Discussion
Medicinal plants and their products are recognized for their powerful biological activities in health and disease treatment [14, 15]. There is no question that the relationship between the immune system and human health is very intimate. Therefore, many new compounds are being isolated and a large variety of phytochemicals are extracted and purified from edible medicinal plants and tested for their anti-inflammatory properties. As others across the world, we are authentically interested in the research of these natural products which have conventionally been used for their medicinal value and their healing properties. In fact, many authors have reported, recently, that medicinal plants such as Phlomis crinita subsp. mauritanica Munby [5], Daphne gnidium [16], and Limoniastrum guyonianum [17, 18] have potent immunomodulatory potential as they display a strong activity on macrophages and lymphocytes. That is why we focused in this study on the potential immunomodulatory effects of T. ramosissimum compounds with regard to macrophage and lymphocyte responses as well as other inflammatory mediators.
A colorimetric MTT assay was used to assess splenocyte proliferation. We present compelling experimental evidence demonstrating that apigenin-7-glucoside, genkwanin, and naringenin modulate the proliferation of splenocytes in the absence of mitogen stimulation. These results suggest that the tested molecules may also contain some factors that are acting as mitogens. The presence of LPS and other mitogens in plant extracts argue in favor of this explanation. Several published data indicate that numerous secondary metabolites like LPS and polyphenols isolated from plants have been proven to possess a wide range of biological functions such as immunomodulation and antioxidant properties [5, 16, 19]. Our data clearly indicate that both LPS- and lectin-stimulated cell proliferations of splenocytes were enhanced by apigenin-7-glucoside, genkwanin, and naringenin at varying tested concentrations (Fig. 1b, c). It is known that LPS was used for T-cell-independent B-cell proliferation, whereas lectin was used for T-cell-dependent B-cell proliferation. The presence of mitogens in the system can postulate the possible activation pathway of the molecules [20]. The compounds with and without LPS or lectin produced different effects on splenocyte proliferation as measured by MTT tetrazolium assay. In the presence of LPS, all tested molecules enhanced the proliferation activity higher than with lectin and B-cell response seemed to be more sensible than T-cell response. These results suggest that T. ramosissimum compounds may contain some mitogenic substrates capable of stimulating cell proliferation.
Cellular immunity is also mediated by T cells, including natural killer cells. NK cells play a major role in defending the host from both tumor and virally infected cells. NK cells are able to recognize and destroy targets that have lost expression of the MHC class I molecules [21]. NK cell differentiation and maturation (to killing states) are each activated by interferon and other cytokines. In the acute study, it was demonstrated that apigenin-7-glucoside, genkwanin, and naringenin significantly enhanced NK cell function. This is the first study to show that in vitro NK cell activity can be affected by T. ramosissimum compounds. This finding is in good accordance with previous studies which showed that many flavonoids have been shown to influence NK cell cytotoxicity [22, 23], although the assays they used were different. A recent study performed by Lindqvist et al. [24] has shown that myricetin could potentiate the ability of NK cells to kill K562 erythroleukemia cells which is in agreement with our findings with apigenin-7-glucoside, genkwanin, and naringenin on NK cell activity.
Cytotoxic T lymphocytes (CTL) play an important role in protection against viral infections, cancer, and rejection of organ transplants. CTLs constitute the major antitumor effector cell population and are recognized for their involvement in host resistance against tumor growth and dissemination [25]. In this report, we highlighted an important role of apigenin-7-glucoside, genkwanin, and naringenin in enhancing CTL activity against tumoral cells (Fig. 3). Among measures of cell-mediated immunity, the cytotoxic T lymphocyte response is recognized as perhaps the most relevant functional measure that reflects cell-mediated acquired immune defense against viral infections and cancer. The immune system plays an important role in surveillance against tumor growth. CTLs constitute the major antitumor effector population; NK cells are recognized for their involvement in host resistance against tumor growth and dissemination [26–28]. Accumulating evidence indicated that tumor cells play a crucial role in the control of immune protection, and tumor cell growth in vivo is not only influenced by CTL-tumor cell recognition but also by tumor susceptibility to cell-mediated death [29, 30]. The efficacy of antitumor CTL critically depends on functional processing and presentation of tumor antigen by the malignant cells and also on their susceptibility to CTL-induced lysis. Thus, a significant increase in ex vivo cytotoxic CTL activity was observed after treatment with different tested molecules.
In this study, we noticed that the cytotoxicity of lymphocytes occurred in a dose-dependent manner in the CTL assay and in a dose-independent manner in the NK cell assay. Furthermore, various cytokines released by NK cells and CTL in the tumor milieu can attenuate tumor angiogenesis by affecting not only tumor cells directly but also by regulating activity of vascular endothelial cells, as well as tumor-associated fibroblasts, myeloid-derived suppressor cells, and alternatively activated M2 macrophages. [31] Therefore, NK cells and CTLs may function in multiple ways to cause tumor rejection, either by directly killing tumor cells or by changing the microenvironment of the tumor in a way that is hostile to further tumor growth and development [32]. In the future, a fuller understanding of the mechanisms by which NK cells and CTLs can reject tumors in vivo may form a rational basis for significantly improving cancer immunotherapy.
One of the most important non-specific immune activities is phagocytosis performed by macrophages (as well as by other types of leukocytes). Macrophages play an important role in the defense mechanism against host infection and the killing of tumor cells. The modulation of the antitumor properties of macrophages by various biological response modifiers is an area of active interest in cancer chemotherapy and is closely associated with the immunomodulatory activity of test drugs [33]. Phagocytosis is accompanied by the release of lysosomal enzymes like acid phosphatase (AP) used for the killing and digesting of microbial pathogens. The higher the augmentation in AP activity, the greater the phagocytic stimulation and intracellular killing capacity [34] With respect to lysosomal enzyme activity, transformation of p-NPP by AP in stimulated macrophages correlates with degranulation processes [7]. A significant decrease in lysosomal phosphatase activity was induced in apigenin-7-glucoside-, genkwanin-, and naringenin-treated cells and showed an anti-inflammatory effect (Fig. 4). According to previous studies, numerous compounds can modulate the immune system due to the presence of hydroxyl groups in their structure, which affects the enzymes or electron-transferring system, especially phagocytic activity [7]. T. ramosissimum compounds contain also hydroxyl groups, so immunomodulatory activities could be attributed to hydroxyl groups present in it. Such results were in agreement with previous studies [34] which showed a significant decrease in macrophage phagocytosis when treated with flavonoids.
Macrophage phagocytosis is also accompanied by a release of free radicals and other reactive oxygen/nitrogen species (ROS/RNS, the latter including NO) involved in pathogen killing [34]. Compared with LPS, a potent inducer of macrophage NO production [35], the studies here showed that apigenin-7-glucoside, genkwanin, and naringenin could inhibit NO synthesis, by peritoneal macrophages (Fig. 5). The decreased release of NO reflects the anti-inflammatory effect of T. ramosissimum compounds. Many plants (and their products) have been shown to influence lysosomal activity and NO synthase activity in macrophages [19]. Moreover, some flavonoids also inhibit TNFa production as well as iNOS expression and NO production in LPS-activated macrophages, an effect that has been associated with the inhibition of the NF-kB pathway [36].
It is well known that oxidative stress induced by the elevation of free radicals or other ROS levels implicated can elicit direct or indirect damage to the body. The generation and subsequent involvement of free radicals contribute to a large number of diseases including inflammatory diseases [37], and it is widely believed that the use of natural antioxidant compounds in such diseases should be reasonably proposed. In this context, we have undertaken to measure the antioxidant activity of apigenin-7-glucoside, genkwanin, and naringenin in splenocyte cells by CAA assay. It appears that T. ramosissimum compounds prevent ABAP-induced DCFH oxidation in splenocytes cells in a dose-dependent manner (Fig. 6). In fact, apigenin-7-glucoside, genkwanin, and naringenin contain numerous double bonds and hydroxyl groups that can donate electrons through resonance to stabilize the free radicals. Furthermore, Estany et al. [38] demonstrated that flavones are the most efficient antioxidant in protecting endometrial cells against oxidative stress.
Conclusion
Based on the findings reported here, we conclude that apigenin-7-glucoside, genkwanin, and naringenin have antioxidant properties as well as immunomodulating effects upon splenocytes, NK and CTL cells, and macrophages. The findings of this study support the view that medicinal plants are promising sources of potential antioxidants and anti-inflammatory agents that may be effective for therapy of human diseases. In spite of the encouraging results, further studies are underway to elucidate the precise mechanism of T. ramosissimum compounds’ effect for the modulation of cellular antioxidant defenses as well as the signaling pathways involved in the modulation of both macrophage and lymphocyte responses.
References
Bello R, Barrachina M, Martínez-Cuesta M, Esplugues J, Yufera EP. Evaluation of the acute toxicity, analgesic and CNS activities of different species of Teucrium genus. Phytother Res. 1995;9:277–80.
Abdollahi M, Karimpour H, Monsef-Esfehani HR. Antinociceptive effects of Teucrium polium L total extract and essential oil in mouse writhing test. Pharmacol Res: Off J Italian Pharmacol Soc. 2003;48:31–5.
Menichini F, Conforti F, Rigano D, Formisano C, Piozzi F, Senatore F. Phytochemical composition, anti-inflammatory and antitumour activities of four Teucrium essential oils from Greece. Food Chem. 2009;115:679–86.
Pottier-Alapetite G. Flore de la Tunisie, Angiospermes-Dicotyledones Gamopetales. Première partie. MES & MA, 1979;764--777.
Limem I, Harizi H, Ghedira K, Chekir-Ghedira L. Leaf extracts from Phlomis crinita Cav. subs. mauritanica Munby affect immune cell functions in vitro. Immunopharmacol Immunotoxicol. 2011;33:309–14.
Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods. 1983;65:55–63.
Manosroi A, Saraphanchotiwitthaya A, Manosroi J. Immunomodulatory activities of Clausena excavata Burm. f. wood extracts. J Ethnopharmacol. 2003;89:155–60.
Sarangi I, Ghosh D, Bhutia SK, Mallick SK, Maiti TK. Anti-tumor and immunomodulating effects of Pleurotus ostreatus mycelia-derived proteoglycans. Int Immunopharmacol. 2006;6:1287–97.
Manosroi A, Saraphanchotiwitthaya A, Manosroi J. In vitro immunomodulatory effect of Pouteria cambodiana (Pierre ex Dubard) Baehni extract. J Ethnopharmacol. 2005;101:90–4.
Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal Biochem. 1982;126:131–8.
Wolfe KL, Liu RH. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J Agric Food Chem. 2007;55:8896–907.
Wang D, Ma Y, Wang J, Liu X, Fang M. Natural killer cells in innate defense against infective pathogens. J Clin Cell Immunol S. 2013;13:2.
Kagi D, Ledermann B, Burki K, Hengartner H, Zinkernagel RM. Cd8+ T cell-mediated protection against an intracellular bacterium by perforin-dependent cytotoxicity. Eur J Immunol. 1994;24:3068–72.
Dubick MA. Historical perspectives on the use of herbal preparations to promote health. J Nutr. 1986;116:1348–54.
Tapsell LC, Hemphill I, Cobiac L, Patch CS, Sullivan DR, Fenech M, et al. Health benefits of herbs and spices: the past, the present, the future. Med J Aust. 2006;185:S4–24.
Harizi H, Chaabane F, Ghedira K, Chekir-Ghedira L. Inhibition of proinflammatory macrophage responses and lymphocyte proliferation in vitro by ethyl acetate leaf extract from Daphne gnidium. Cell Immunol. 2011;267:94–101.
Krifa M, Bouhlel I, Ghedira-Chekir L, Ghedira K. Immunomodulatory and cellular anti-oxidant activities of an aqueous extract of Limoniastrum guyonianum gall. J Ethnopharmacol. 2013;146:243–9.
Krifa M, Mustapha N, Ghedira Z, Ghedira K, Pizzi A, Chekir-Ghedira L. Limoniastrum guyonianum methanol extract induces immunomodulatory and anti-inflammatory effects by activating cellular anti-oxidant activity. Drug Chem Toxicol. 2015;38:84–91.
Ben Sghaier M, Krifa M, Mensi R, Bhouri W, Ghedira K, Chekir-Ghedira L. In vitro and in vivo immunomodulatory and anti-ulcerogenic activities of Teucrium ramosissimum extracts. J Immunotoxicol. 2011;8:288–97.
Nakamura A, Nagai K, Suzuki S, Ando K, Tamura G. A novel method of screening for immunomodulating substances, establishment of an assay system and its application to culture broths of microorganisms. J Antibiot. 1986;39:1148–54.
Ljunggren HG, Karre K. In search of the ‘missing self’: MHC molecules and NK cell recognition. Immunol Today. 1990;11:237–44.
Ismail N, Abdullah H, Seidel V, Rotondo D. Human natural killer (NK) cell activation by luteolin from Brucea javanica leaves. J Cancer Res Exp Oncol. 2012.
Krifa M, El Mekdad H, Bentouati N, Pizzi A, Ghedira K, Hammami M, et al. Immunomodulatory and anticancer effects of Pituranthos tortuosus essential oil. Tumour Biol: J Int Soc Oncodevelopmental Biol Med. 2015.
Lindqvist C, Bobrowska-Hagerstrand M, Mrowczynska L, Engblom C, Hagerstrand H. Potentiation of natural killer cell activity with myricetin. Anticancer Res. 2014;34:3975–9.
Whiteside TL, Herberman RB. The role of natural killer cells in immune surveillance of cancer. Curr Opin Immunol. 1995;7:704–10.
Sheeja K, Kuttan G. Activation of cytotoxic T lymphocyte responses and attenuation of tumor growth in vivo by Andrographis paniculata extract and andrographolide. Immunopharmacol Immunotoxicol. 2007;29:81–93.
Guo TL, Chi RP, Hernandez DM, Auttachoat W, Zheng JF. Decreased 7,12-dimethylbenz[a]anthracene-induced carcinogenesis coincides with the induction of antitumor immunities in adult female B6C3F1 mice pretreated with genistein. Carcinogenesis. 2007;28:2560–6.
Shi X, Ruan D, Wang Y, Ma L, Li M. [Anti-tumor activity of safflower polysaccharide (SPS) and effect on cytotoxicity of CTL cells, NK cells of T739 lung cancer in mice]. Zhongguo Zhong yao za zhi = Zhongguo zhongyao zazhi = China J Chin Mater Med. 2010;35:215–8.
Trapani JA, Smyth MJ. Functional significance of the perforin/granzyme cell death pathway. Nat Rev Immunol. 2002;2:735–47.
Siveen KS, Kuttan G. Effect of Aerva lanata on cell-mediated immune responses and cytotoxic T-lymphocyte generation in normal and tumor-bearing mice. J Immunotoxicol. 2012;9:25–33.
Herberman RB. Assessment of cellular immune response to cancer of the breast. Ann Clin Lab Sci. 1979;9:467–73.
Warren HS, Smyth MJ. NK cells and apoptosis. Immunol Cell Biol. 1999;77:64–75.
Schepetkin IA, Quinn MT. Botanical polysaccharides: macrophage immunomodulation and therapeutic potential. Int Immunopharmacol. 2006;6:317–33.
Jakhar R, Paul S, Chauhan AK, Kang SC. Morin hydrate augments phagocytosis mechanism and inhibits LPS induced autophagic signaling in murine macrophage. Int Immunopharmacol. 2014;22:356–65.
MacMicking J, Xie QW, Nathan C. Nitric oxide and macrophage function. Annu Rev Immunol. 1997;15:323–50.
Tamir S, Tannenbaum SR. The role of nitric oxide (NO.) in the carcinogenic process. Biochim Biophys Acta. 1996;1288:F31–36.
Sagnia B, Fedeli D, Casetti R, Montesano C, Falcioni G, Colizzi V. Antioxidant and anti-inflammatory activities of extracts from Cassia alata, Eleusine indica, Eremomastax speciosa, Carica papaya and Polyscias fulva medicinal plants collected in Cameroon. PLoS One. 2014;9, e103999.
Estany S, Palacio JR, Barnadas R, Sabes M, Iborra A, Martinez P. Antioxidant activity of N-acetylcysteine, flavonoids and alpha-tocopherol on endometrial cells in culture. J Reprod Immunol. 2007;75:1–10.
Acknowledgments
We acknowledge the financial support from the “Ministry of Higher Education, Scientific Research and Technology, Tunisia.”
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
None
Rights and permissions
About this article

Cite this article
Nasr-Bouzaiene, N., Sassi, A., Bedoui, A. et al. Immunomodulatory and cellular antioxidant activities of pure compounds from Teucrium ramosissimum Desf.. Tumor Biol. 37, 7703–7712 (2016). https://doi.org/10.1007/s13277-015-4635-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13277-015-4635-0