
Abstract
Thymoquinone (TQ) is the active ingredient of Nigella sativa which has a therapeutic potential in cancer therapy and prevention. In this study, TQ has been shown to induce specific cytotoxicity and apoptosis and to inhibit wound healing in triple-negative breast cancer cell line. TQ also inhibited cancer growth in a mouse tumor model. Moreover, TQ and paclitaxel (Pac) combination inhibited cancer growth in cell culture and in mice. Genes involved in TQ and TQ-Pac-mediated cytotoxicity were studied using focused real-time PCR arrays. After bioinformatic analysis, genes in apoptosis, cytokine, and p53 signaling categories were found to be modulated with a high significance in TQ-treated cells (p < 10−28, p < 10−8, and p < 10−6, respectively). Important to note, TQ has been found to regulate the genes involved in the induction of apoptosis through death receptors (p = 5.5 × 10−5). Additionally, tumor suppressor genes such as p21, Brca1, and Hic1 were highly upregulated by TQ and TQ-Pac combination. Interestingly, when cells were treated with high dose TQ, several growth factors such as Vegf and Egf were upregulated and several pro-apoptotic factors such as caspases were downregulated possibly pointing out key pathways manipulated by cancer cells to resist against TQ. In cells treated with the combination of TQ and Pac, genes in apoptosis cascade (p < 10−12), p53 signaling (p = 10−5), and JAK-STAT signaling (p < 10−3) were differentially expressed. TQ has also been shown to induce protein levels of cleaved Caspase-3, Caspase-7, and Caspase-12 and PARP and to reduce phosphorylated p65 and Akt1. The in vivo therapeutic potential of TQ-Pac combination and the genetic network involved in this synergy have been shown for the first time to the best of our knowledge.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Thymoquinone (TQ) is the active ingredient of the essential oil of Nigella sativa [1]. Numerous studies demonstrated the therapeutic role of TQ in cancer therapy and prevention [2]. The therapeutic role of TQ in cancer has been shown to be mostly dependent (but not limited to) on three regulatory networks: (i) apoptosis, (ii) cell cycle, and (iii) NF-Kappa-B signaling. TQ has been shown to induce apoptosis in various cancer types through the modulation of key factors including P53; P21WAF1; caspases 3, 8, and 9; Bax; and Bcl-2 [3–7]. TQ also intervenes with cell cycle progression and inhibits G1 to S phase progression [5–8].
TNF-alpha may involve in inflammation and cell proliferation through NF-kappa-B signaling pathway [9]. TQ has been reported to inhibit NF-kappa-B activation and TNF production in various autoimmune models and cancer cell lines [9–14]. One of the mechanisms that TQ suppressed NF-kappa-B pathway is the inhibition p65 binding to DNA [7]. We recently reported that the transcription of multiple genes in the TNF-NF-kappa-B signaling axis was found to be affected during TQ-mediated apoptosis in HeLa cells [15]. TQ inhibited in vivo growth of mouse mammary tumor through NF-kappa-B [11].
TQ arrested cells in G1 phase, affected cell cycle genes (cyclin D1, cyclin E, and p27), and induced apoptosis and disturbed the structure and function of mitochondria in breast cancer cell lines [16] and in triple-negative breast cancer cells lacking functional P53 [17]. Anti-tumor effect of TQ through p38 has been found in breast cancer models [18]. TQ has been demonstrated to inhibit Akt phosphorylation and synergize with cisplatin [19]. TQ has also been found to sensitize breast cancer cells to a single dose of ionizing radiation [20] and doxorubicin in multidrug-resistant breast cancer cell line [21].
Taxol was first isolated from Taxus brevifolia in 1971 in an effort by National Cancer Institute in USA [22]. Taxol targets cancer cells through stabilization of microtubules [23]. Paclitaxel (Pac) and docetaxel are active compounds of two taxol-based commercial preparations and currently used in breast and ovarian cancer therapies [24]. There are only few studies on in vitro combination of TQ and Pac. In one study, TQ-loaded nanoparticles have been shown to sensitize cancer cells to Pac-induced cytotoxicity [14]. In another study, TQ and docetaxel combination had synergistic cytotoxicity in vitro in prostate cancer cells [25]. To the best of our knowledge, there is not a study on the combination of TQ and Pac in vivo.
Ehrlich ascite tumor is a non-differentiated tumor cell line which is derived from mouse mammary adenocarcinoma and can form ascites progressively when injected intraperitoneally in mice [26]. 4T1 is a mammary carcinoma cell line derived from a spontaneous tumor of Balb/c mice and represents a stage IV, triple-negative, highly metastatic breast cancer [27, 28].
In this study, the therapeutic effect of TQ and Pac has been demonstrated in both in vitro and in vivo breast cancer models. Importantly, detailed investigation on genetic networks involved in the action of TQ and Pac was performed using pathway focused real-time PCR panels. Genes that take part in apoptosis, cell proliferation, growth factor activity, and cytokine activity have been demonstrated to have significant importance in the therapeutic role of TQ.
Materials and methods
Animals
Eight to twelve-week female Balb/c mice obtained from the Hakan Çetinsaya Experimental and Clinic Research Center, Erciyes University, Kayseri, Turkey, were used for this study. All the animals received humane care according to the standard guidelines. Ethical approval for the study was obtained from Erciyes University Animal Researche Local Ethics Committee and the ethic regulations have been followed in accordance with international, national, and institutional guidelines.
Cell culture
Mouse breast cancer cell line, 4T1 (ATCC CRL-2539), was cultured in RPMI-1640 growth medium (Biochrom, Berlin, Germany) with the supplements (10 % FBS (Biochrom), 2 mM l-glutamine (Biochrom), 1 mM sodium pyruvate (Biochrom), 10 mM HEPES (Biochrom), and penicillin–streptomycin (Biochrom)) at 37 °C and 5 % CO2 under sterile conditions. Mouse embryonic fibroblasts, PMEF (PMEF-CFL-P1), was cultured in DMEM growth medium (Biochrom) with the supplements.
Cell viability assays
4T1 and mouse embryonic fibroblast (MEF) cells were incubated in the presence of certain doses of TQ (Sigma, Taufkirchen, Germany) for 48 h. Cell viability assays were performed as described by us earlier [15].
Assessment of apoptosis with Giemsa staining of 4T1 cells
4T1 breast cancer cells were grown to 60–70 % confluency in six-well plate for 24 h at a density of 3–4 × 104 cells/cm2. Then, cells were incubated at 37 °C and 5 % CO2 with certain concentrations of TQ (control, 6.25, 12.5, 25, 50, and 100 μM) for 24 h. Assessment of apoptosis with Giemsa staining was performed as described in [15].
Wound healing assay
4T1 cells were grown to 100 % confluency in 25-cm2 flasks. A scratch wound was made in all flasks. Cells were incubated at certain different concentrations (0, 6.25, 12.5, 25, 50, and 100 μM) of TQ for 48 h. Wound healing assay was performed as described in [15].
In vitro cytotoxicity assay for the combination of thymoquinone and paclitaxel
4T1 breast cancer cells were grown to 70 % confluency in six-well plate at a density of 3–4 × 104 cells/cm2. TQ (Sigma) and Pac (Bristol Myers Squibb, New York, USA) were added at certain concentrations, and cells were incubated for 24 h. Cell viability assays were performed as described in [15]. Ehrlich tumor cells were injected intraperitoneally (i.p.) into mice, and after 8–10 days, cells were collected from tumor ascites of mice and counted using trypan blue staining. Approximately, 5 × 105 cells were plated in 2-ml 24-well plates. TQ and Pac were added at certain concentrations, and cells were incubated for 24 h. Total number of viable cells was calculated by counting in hemacytometer using trypan blue staining.
In vivo mouse tumor model
Ehrlich tumor ascites (EAC) were kindly provided by Assoc. Prof. Dr. Tolga Ertekin, Erciyes University. EAC cells were obtained from tumor ascite fluids of previously injected female Balb/c mice. Each mice were injected with 5 × 106 cells in 200 μl volume, and mice were weighed daily. Tumor growth was monitored by the evaluation of the weights of mice. The doses of TQ and Pac (Bristol Myers Squibb) were administered intraperitoneally daily for TQ and weekly for Pac. TQ was dissolved in dimethyl sulfoxide (DMSO) and then diluted in ddH2O on the day of injection. Pac was in a suspension including 50 % ethanol and diluted in ddH2O on the day of injection. Control mice were injected with the same amount of DMSO and/or ethanol in ddH2O. Tumor growth was monitored by weighing the mice daily. Student’s t test was used to compare the difference between the groups.
RNA isolation and cDNA synthesis
For RNA isolation, 4T1 cells were grown to 90–100 % confluency for 48 h in 25-cm2 flasks. Then, certain concentrations of TQ were added, and cells were incubated for 24 h. As a control, only DMSO was added as vehicle. 4T1 cells were washed with PBS and trypsinized. RNA isolation was performed using RNeasy Plus Mini Kit (QIAGEN, Hilden, Germany). RNA quality and concentration were checked with NanoDrop. OD 260/280 and 260/230 values were above 1.8 for all samples. RNAs were kept frozen at −80 °C. Five hundred nanograms of RNA was used for cDNA synthesis using RT2 First Strand Kit (QIAGEN). Briefly, RNA, BufferGE2, and RNAse-free H2O were mixed and incubated at 37 °C for 5 min for genomic DNA elimination. Then, Buffer BC4 including reverse transcriptase, random hexamers, oligo-dT, and other components were added. The mix was incubated at 42 °C for 15 min and 95 °C for 5 min. cDNAs were diluted with RNAse-free H2O and kept frozen at −20 °C until the gene expression analysis.
Gene expression analysis by quantitative real-time PCR
In this study, QIAGEN RT2 Profiler PCR Arrays were utilized. Three panels used were the following: (i) Mouse Apoptosis Array, (ii) Mouse Breast Cancer Array, and (iii) Mouse Wound Healing Array. These panels have primers for 84 pathway-related genes, 5 housekeeping genes, additionally 3 primers for reverse transcription control, 3 primers for positive PCR control, and 1 primer for genomic DNA contamination control. For real-time PCR experiment, briefly, 2xRT2SYBR Green Mastermix, cDNA, and RNAse-free H2O were mixed and added into each well of 96-well plate. Roche LightCycler 480 instrument was used to run real-time PCR. The protocol for real-time PCR was the following: 1 cycle of preincubation for 10 min at 95 °C and 45 cycles of amplification for 15 s at 95 °C, for 1 min at 60 °C. Each experiment was performed twice. The expression data was analyzed online using the “Data Analysis Center” software (QIAGEN). Ct cutoff threshold was set as 35. Average of the expression values of five housekeeping reference genes were used for the normalization of target genes. Genes that were downregulated or upregulated more than twofold were considered to be modulated. The bioinformatics analysis of the gene list was performed using the functional annotation tool of the bioinformatics software, DAVID [29, 30].
Western blotting
4T1 cells were grown to 60–70 % confluency in 10-cm petri dishes at a density of 3–4 × 104 cells/cm2. Then, cells were incubated at 37 °C and 5 % CO2 with certain concentrations of TQ (control, 12.5, 25, and 50 μM), Pac (10 μg/ml), and Pac + TQ (10 μg/ml + 12.5 μM) for the indicated time. Cells were washed and lysed in 150 μl RIPA buffer (no. R0278, Sigma) for 30 min, and supernatants were collected after centrifugation. Twenty micrograms of protein were run using ready-to-use gels (no. 456-8123, Bio-Rad, California, USA) and blotted to PVDF membrane (no. 10413096, GE Healthcare Life Sciences, USA). Membranes were blocked with nonfat dry milk (no. 170-6404, Bio-Rad) for 1 h and probed overnight at 4 °C with the following primary antibody sets: Apoptosis antibody sampler kit (mouse preferred, no. 9930, Cell Signalling Technology, MA, USA), Caspase 7 antibody (no. 9492), Cleaved Caspase 7 (no. 9491), Phospho-Akt (no. 4060), Beta-Actin (no. 3700) and Beta-Tubulin (no. 2146), Phospho-p65 (no. 3033), p65 (sc-109, Santa Cruz Biotechnology, Dallas, USA), Bcl-2 (sc-492), Bax (sc-493), and appropriate secondary antibodies (All from Cell Signalling Technology). Membranes were treated with ECL (no. 170-5060, Bio-Rad) and visualized using ChemiDoc MP imaging system instrument (Bio-Rad).
Results
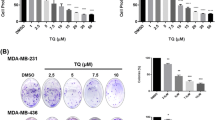
Thymoquinone induces stronger cytotoxicity in 4T1 mouse breast cancer cells compared to control fibroblast cells
Cytotoxic effect of TQ was studied using cell viability assay by the utilization of 4T1 mouse breast cancer cell line and mouse embryonic fibroblasts (MEF). 4T1 and MEF cells were incubated at different concentrations of TQ ranging from 3.125 to 100 μM by twofold increments. TQ induced effective cytotoxic effect on 4T1 cells with an IC50 value of approximately 1.3 μM (Supplementary Fig. 1B). Cytotoxic effect of TQ on MEF was milder with an IC50 value of 10.3 μM (Supplementary Fig. 1C).
TQ inhibits wound healing and migration in 4T1 cells
An in vitro cell culture wound healing assay was performed to evaluate the impact of TQ on the proliferation and migration of 4T1 breast cancer cells. After making the scratch wound, cells were grown in culture medium containing TQ in concentrations of 6.25, 12.5, 25, 50, and 100 μM together with control. Microscope images were taken at the beginning and at every 12 h (Fig. 1a). The closure of the wound gap was monitored and calculated by making multiple measurements using specific image software. TQ significantly inhibited wound healing at all doses except for 6.25 μM (Fig. 1a, b). Averages of measurements of wound healing were plotted (Fig. 1b, Supplementary Table 1).

a Scratch wound healing assay in the presence of different concentrations of TQ. a Representative images were obtained at the beginning, at 12 and 24 h. a–f Images of scratch wounds in cultures for the control, 6.25, 12.5, 25, 50, and 100 μM concentrations of TQ, respectively, at the beginning. g–l Images of scratch wounds in cultures at 12 h. m–s Images of scratch wounds in cultures at 24 h. Arrows indicate the width of the wound. b Wound width was depicted at the beginning, 12 and 24 h for the control and all concentrations of TQ in the graph. Error bars indicate standard error. Star signs indicate the significant difference between the control and TQ-treated samples (p < 0.05)
TQ induces apoptosis in 4T1 cells
Degree of apoptosis induced by TQ was evaluated in 4T1 cells using Giemsa staining. Treatment with TQ for 24 h resulted apoptosis in 4T1 cells at all doses ranging from 6.25 to 100 μM (Supplementary Fig. 2). Apoptotic cells were clearly identified by their condensed and darker nucleus and changed morphology (Supplementary Fig. 2A–E). Percentages of apoptotic cells were 11.8 ± 1.6 for the control, 25.8 ± 2.8 for 6.25 μM, 67.6 ± 3.2 for 12.5 μM, 86.6 ± 1.6 for 25 μM, 98.4 ± 0.9 for 50 μM, and 100 ± 0 for 100 μM dose of TQ in 4T1 breast cancer cells, respectively. Percentages of apoptotic cells were plotted (Supplementary Fig. 2F).
Combination of TQ and Pac strongly induces cytotoxicity in 4T1 cells and Ehrlich tumor cells in vitro
TQ and Pac both induced cytotoxicity in 4T1 cells (Fig. 2a). When TQ and Pac were combined together (TQ 6.25, 12.5, and 25 μM and Pac 10 μg/ml), they induced a stronger cytotoxicity compared to each agent alone (TQ 6.25 μM + Pac 10 μg/ml vs. Pac 10 μg/ml, p = 0.0004; TQ 12.5 μM + Pac 10 μg/ml vs. Pac 10 μg/ml, p = 0.0001; TQ 25 μM + Pac 10 μg/ml vs. TQ 25 μM, p = 0.0002) (Fig. 2a). Interestingly, when TQ was added before Pac, TQ sensitized 4T1 cells to Pac and the induced cytotoxicity was significantly higher compared to when two agents were given at the same time (TQ 12.5 μM + Pac 10 μg/ml vs. TQ 12.5 μM/Pac 10 μg/ml, p = 0.0025). The same phenomenon was observed in Ehrlich tumor cells. TQ and Pac both induced cytotoxicity in Ehrlich tumor cells (Fig. 2b). When TQ and Pac were combined together (TQ 6.25 μM and Pac 5 and 10 μg/ml), they induced a stronger cytotoxicity compared to each agent alone (TQ 6.25 μM/Pac 5 μg/ml vs. Pac 5 μg/ml, p = 0.0009; TQ 6.25 μM/Pac 10 μg/ml vs. Pac 10 μg/ml, p = 0.02) (Fig. 2b).

Induction of cytotoxicity by the combinations of TQ and paclitaxel (Pac) in vitro a in 4T1 breast cancer cells and b in Ehrlich breast tumor cells. Doses of TQ (μM) and Pac (μg/ml) were indicated. Error bars indicate standard deviation. TQ + Pac indicates that, first, TQ was added, and then Pac was added 4 h later. TQ/Pac indicates that TQ and Pac were added at the same time. Single-star sign indicates the significant difference from the control. Double-star sign indicates the significant difference between compared groups
TQ significantly inhibits the growth of Ehrlich tumor in vivo
Ehrlich tumor cells form ascites in 8–12 days when injected into mice intraperitoneally. TQ significantly inhibited the growth of i.p. injected Ehrlich tumor in mice when given five times a week at a dose of 3.2 mg/kg (p = 0.012) (Fig. 3a). A lower dose of TQ (0.64 mg/kg) inhibited the growth of tumor to some degree, although it was not statistically significant (p = 0.14) (Fig. 3a).

The effect of TQ, Pac, and their combination on the growth of Ehrlich tumor in vivo. a The effect of low dose (0.64 mg/kg) and high dose (3.2 mg/kg) TQ on the growth of Ehrlich tumor in mice. b The effect of the combination of low dose TQ (0.64 mg/kg) and Pac (1.25 mg/kg) on the growth of Ehrlich tumor in mice. c The effect of the combination of high dose TQ (2.4 mg/kg) and Pac (1.25 mg/kg) on the growth of Ehrlich tumor in mice. The significant differences (p < 0.05) between individual groups were indicated by using a star sign. Error bars indicate standard error. The number of mice in each group (n) was shown. Student’s t test was used to compare the difference between the groups
Combination of TQ and Pac inhibits the growth of Ehrlich tumor in vivo
A moderate (suboptimal) dose of Pac (1.25 mg/kg) inhibited the growth of Ehrlich tumor to some degree at earlier stages; however, the inhibition was not significant at later stages. A lower dose of TQ (0.64 mg/kg) did not have an inhibitory effect on tumor growth. The combination of Pac and the lower dose TQ significantly inhibited the tumor growth compared to both TQ alone and Pac alone groups (p = 0.003 and p = 0.007, respectively) (Fig. 3b). A higher dose of TQ (2.4 mg/kg) inhibited the tumor growth until later stages (p < 0.001) (Fig. 3c). The combination of Pac and the higher dose TQ significantly inhibited the tumor growth compared to both TQ alone and Pac alone groups (p < 0.001 in both) (Fig. 3c).
TQ modulates extrinsic and intrinsic apoptosis genes, p53 signaling genes, and growth factor genes as revealed by focused real-time PCR panels
4T1 cancer cells were untreated (control) or treated with TQ (12.5, 25, and 50 μM). Profiling of the expression of the genes modulated by low dose (12.5 μM) and high dose TQ (50 μM) was performed using three focused real-time PCR panels containing 224 genes. These three panels were apoptosis, breast cancer, and wound healing panels. More than 70 genes were found to be either downregulated or upregulated by high dose TQ. These genes were analyzed and categorized into functional groups. Genes in the regulation of programmed cell death, immune response, regulation of cell proliferation, and p53 signaling pathway categories were found to be enriched significantly. Details of categories, modulated genes, and p values were given in Table 1. A representative list of genes that were strongly modulated by low dose and high dose TQ, and their fold changes were given in Table 2. Interestingly, tumor suppressor genes such as Cdkn1a and Hic1 have been found to constitute a pivotal portion of the highly upregulated genes (Table 2).
Combination of TQ and Pac differentially modulates Fasl- and Trail-mediated apoptosis, p53 signaling, and JAK-STAT signaling genes as revealed by focused real-time PCR panels
4T1 cancer cells were untreated (control) or treated with TQ (12.5 μM), Pac (10 μg/ml), and their combination (TQ, 12.5 μM + Pac, 10 μg/ml). Genes that were differentially expressed in the combination of TQ (12.5 μM) and Pac (10 μg/ml) compared to Pac and TQ alone were investigated by real-time PCR methodology. These doses of TQ and Pac were selected because their combination resulted in the most synergistic cytotoxicity in 4T1 cells (Fig. 2a).
Profiling of the expression of the genes modulated by the combination of TQ and Pac was also performed using three focused real-time PCR panels. The genes that were modulated in the combination compared to Pac alone samples were selected and analyzed. Sixty-four genes were found to be either downregulated or upregulated in the combination of TQ and Pac. These genes were analyzed and categorized into functional groups. Genes in apoptosis, p53 signaling, cytokine-cytokine receptor interaction, and JAK-STAT signaling were found to be enriched specifically in the combination TQ and Pac. Especially, genes involved in Fasl- and Trail-mediated extrinsic apoptosis such as Fasl, Fas, Trail, Tnfrsf11b, Dff45, Casp7, and Bid were affected. Additionally, genes hypermethylated in breast cancer such as Hic1 (~sevenfolds), Gstp1 (~fourfolds), and Brca1 (~twofolds) were upregulated in TQ-Pac combination compared to Pac alone treatment. Details of categories, modulated genes, and p values were given in Table 3.
Apoptosis induced by TQ and TQ-Pac combination is characterized by increased protein levels of full length and cleaved Caspase-3, Caspase-7, and Caspase-12
Since the apoptosis was the most significantly affected network in our gene expression studies, we further characterized the protein levels of full length and cleaved caspases in 4T1 cells treated with TQ and TQ + Pac at different time points. At the same time, we characterized levels of Bax, Bcl-2, phosphorylated p65, and Akt1. We have found that TQ causes a strong increase of full length and cleaved Caspase-3 at 6 and 10 h, respectively (Fig. 4a, b). At 14 h, we observed the increase of cleaved Caspase-7 and cleaved PARP and a moderate increase of cleaved Caspase-12 especially at high dose TQ (50 μM) (Fig. 4c). Interestingly, we could not detect cleaved initiator caspases (Caspase-8 and Caspase-9) at 2, 3, 6, and 10 h. We detected a weak signal for cleaved Caspase-8 at 14 h; however, the signal was also present in the control (Fig. 4c). In our transcription profiling, we have shown that mRNA levels of Caspase-7 and Caspase-12 were reduced by TQ at 24 h. At 6 and 24 h, we have shown that TQ also reduces the protein levels of full length Caspase-7 and Caspase-12 (Fig. 4a, d). We have also characterized the activity of survival factors Akt1 and p65 in 4T1 cells and found that TQ inhibits the phosphorylation of Akt1 and p65 (Fig. 4a, d).

Western blot analysis. TQ 12.5, TQ 25, TQ 50, Pac 10, Pac + TQ represent the concentrations of TQ 12.5, 25, and 50 μM; Pac 10 μg/ml, and Pac 10 μg/ml + TQ 12.5 μM used in 4T1 cells, respectively. a Protein levels of indicated factors in treated 4T1 cells at 6 h, b at 10 h, c at 14 h, and d at 24 h. Cl cleaved, P phosphorylated. Beta-actin and Beta-tubulin were used as loading controls
Discussion
In this study, the anti-tumor effect of TQ has been shown both in vitro and in vivo in mouse breast cancer model. Treatment of triple-negative breast cancer cells with TQ resulted in induction of cytotoxicity and apoptosis and inhibition of wound healing. Next, we have demonstrated that TQ and Pac shows cooperative anti-tumor effect when combined in vitro (Fig. 2) and in vivo (Fig. 3). These data suggest a synergistic cooperation of these two therapeutic agents and suggest that TQ sensitizes tumor cells to Pac.
In order to find out the molecular mechanisms through which TQ exerts its anti-tumor impact, gene expression profile of TQ-treated 4T1 cells was investigated. Focused real-time PCR arrays were used. TQ has been found to modulate the expression of genes involved in apoptosis through death receptors and mitochondrial pathway, immune response, regulation of cell proliferation, and p53 signaling pathway (Table 1).
Another important point to note is the presence of interesting patterns in the gene expression profile induced by TQ. In the expression profile, we observe both the direct impact of TQ and also the reaction of cancer cells to survive. For instance, several anti-apoptotic factors such as Bcl2a1a, Birc3, and Birc5 were found to be downregulated by high dose TQ; however, several pro-apoptotic factors such as Casp4 and Casp12 were also found to be downregulated probably as a reaction of cancer cells. Similarly, growth factors including Vegfa, Egf, and Fgf2 were upregulated when cancer cells were treated with high dose TQ. This is expected since the expression profile has been studied using the cells surviving after 24 h treatment of high dose TQ. Therefore, this might be a result of the response of cancer cells to resist to the apoptosis induced by TQ or a result of genetic feedback mechanism. These genes may give strong clues about the pathways that TQ intervenes with. As we already know that TQ induces apoptosis and activates the caspase cascade [3–7], the downregulation of caspases with high dose TQ suggests that cancer cells downregulate caspase expression to escape apoptosis. In other words, the downregulation of caspases might point out a resistance mechanism of cancer cells against TQ-induced apoptosis. Therefore, a comprehensive analysis of upregulated and downregulated genes together seems to be a more useful approach in order to find out key cascades that TQ hits.
Gene expression profile of 4T1 cells treated with the combination of TQ and Pac was analyzed to reveal the potential molecular actors responsible for their cooperative anti-tumor activity. Genes involved in apoptosis, growth factor activity, and cell cycle were found to be modulated in TQ-Pac combination compared to each agent alone. These findings suggest that TQ increase the sensitivity and decrease the resistance of 4T1 cells to Pac by tuning the activity of these factors. In our study, we found that the expression of cytokine genes were modulated in TQ-Pac combination. In accordance with our findings, TQ has been reported to modulate the levels of several cytokines and chemokines such as IL-6 and MCP-1 [31, 32]. TQ has been reported to inhibit NF-kappa-B and IL-8 expression which is overexpressed in hepatocellular carcinoma [33]. TQ also has been reported to suppress STAT3 phosphorylation and Cxcl12-induced migration in multiple myeloma cells [34–36]. TQ has also been shown to inhibit proinflammatory mediators including TNF-alpha and IL-1beta in pancreatic cancer cells [13]. Therefore, it seems that the modulation of inflammatory mediators may be an important aspect of the anti-tumor impact of TQ (Table 3).
TQ has been found to radiosensitize breast cancer cells through the modulation of the levels of integrin αV, MMP9, and MMP2 [37]. Similarly, we have also found out TQ-modulated expression of many wound healing-related genes including Ctsk, Mmp9, and Serpine1. It has been shown that TQ inhibits the phosphorylation of EGFR through JAK2 and Src [38]. Interestingly, in our study, TQ was found to affect levels of several growth factors including Egf, Fgf2, Pdfga, and Vegfa (Table 1). At the same time, JAK-STAT signaling was found to be affected in TQ-Pac combination (Table 3).
One of the important findings of this study was the tremendous upregulation of several key tumor suppressor genes in TQ-treated breast cancer cells. These genes were normally hypermethylated in cancer [39–41]. Cdkn1a (~11-folds), Hic1 (~10-folds), Gata3, Gstp1, and Brca1 were the major tumor suppressor genes that were upregulated by TQ (Table 2). Moreover, Hic1 (~sevenfolds), Gstp1 (~fourfolds), and Brca1 (~twofolds) were also upregulated in TQ-Pac combination compared to Pac alone treatment.
We have shown that TQ sensitized cancer cells to Pac both in vitro and in vivo. There have been several reports on the basis of Pac resistance. Increased expression of anti-apoptotic factors such as Bcl-2 and Bcl-XL and decreased pro-apoptotic factors such Bax and Bad, have been shown to be related with Pac resistance [42]. Interestingly, we have shown that TQ induces apoptosis in breast cancer cells and strongly modulates many genes involved in apoptosis with a p value of 2.2E−29 (Table 1). Therefore, it is quite likely that TQ breaks the resistance of breast cancer cells to Pac through interplaying with key actors in apoptosis. We have also shown that genes in apoptosis cascade (especially genes related with extrinsic cascade such as Fas, Fasl, Trail, Casp7, and Bid) and EGFR-AKT axis (such as Egfr and Akt1) were differentially expressed in TQ-Pac combination (Fig. 5). This finding suggests that TQ may stimulate extrinsic apoptosis pathway and may inhibit EGFR-AKT signaling axis to make cancer cells more prone to Pac cytotoxicity. Especially, the affected cluster of genes involved in Fasl- and Trail-mediated apoptosis (Fasl, Fas, Trail, Opg (Tnfrsf11b), Dff45, Casp7, and Bid) should be noted in TQ-Pac combination. In agreement with our findings, TQ has been reported to induce Fas-mediated apoptosis through relocalization of Fas to the membrane in multiple myeloma cells and to sensitize HepG2 cells to TRAIL-induced death [33, 35]. Thus, Fasl-Trail-mediated apoptosis axis should be taken into consideration for their role in TQ-Pac synergy in further studies especially at the proteomics and posttranslational modification level. p65 is a key factor in NF-kappa-B signaling, may be induced by Trail and Fas and is involved in the inhibition of apoptosis [9]. We have also shown that TQ reduces the levels of full length p65 and phosphorylated p65 and Akt which may be related to the modulation of Fasl and Trail (Figs. 4 and 5).

Summarized graphical illustration of interactions of genes involved in apoptosis cascade (mainly extrinsic) and the genes involved in the crosstalk of EGFR-AKT axis with apoptosis. The illustration also depicts the involvement of JAK-STAT signaling. Genes that were modulated by TQ-Pac combination are marked by a star sign. Genes that were confirmed to be modulated by TQ at the protein level are marked by a circle. → stimulatory interaction, ┤ inhibitory interaction, - -> multiple stimulatory interactions. Pathway-related data from DAVID and KEGG databases were utilized to construct the graphical illustration [29, 30, 44]
It has also been reported that IL-6 and IL-8 are overexpressed in Pac-resistant cancer cell lines [42]. We have shown that IL-6 and Stat3 and other genes in JAK-STAT signaling are modulated in cells treated with the combination of TQ and Pac. This finding suggests that another path through which TQ breaks the resistance to Pac is possibly IL-6-Stat3 axis (Fig. 5). In another study, Pac resistance in breast cancer cells has been connected to the downregulation of caspase-7, caspase-9, and BIM [43]. In accordance with these findings, in our study, TQ and TQ-Pac combination affected the expression and the activity of Caspase-3, Caspase-7, and Caspase-12 in breast cancer cells (Tables 1, 2, and 3; Figs. 4 and 5). Therefore, the caspase activity might involve in the synergistic act of TQ and Pac.
In this study, the in vivo therapeutic potential of TQ-Pac combination and a portion of genes involved in TQ-Pac cooperation have been shown. At the same time, the detailed analysis of numerous genes involved in TQ-mediated apoptosis of breast cancer cells were performed. The collected data suggest that TQ may sensitize cancer cells to Pac through multiple cascades including extrinsic apoptosis, tumor suppressor genes, and p53 signaling.
References
El-Dakhakhany M. Studies on the chemical constitution of Egyptian N. sativa L. seeds. Planta Med. 1963;11:465–70.
Rahmani AH, Alzohairy MA, Khan MA, Aly SM. Therapeutic implications of black seed and its constituent thymoquinone in the prevention of cancer through inactivation and activation of molecular pathways. Evid Based Complement Alternat Med. 2014. doi:10.1155/2014/724658.
El-Mahdy MA, Zhu QZ, Wang QE, Wani G, Wani AA. Thymoquinone induces apoptosis through activation of caspase-8 and mitochondrial events in p53-null myeloblastic leukemia HL-60 cells. Int J Cancer. 2005;117(3):409–17. doi:10.1002/Ijc.21205.
Gali-Muhtasib H, Diab-Assaf M, Boltze C, Al-Hmaira J, Hartig R, Roessner A, et al. Thymoquinone extracted from black seed triggers apoptotic cell death in human colorectal cancer cells via a p53-dependent mechanism. Int J Oncol. 2004;25(4):857–66.
el Arafa SA, Zhu Q, Shah ZI, Wani G, Barakat BM, Racoma I, et al. Thymoquinone up-regulates PTEN expression and induces apoptosis in doxorubicin-resistant human breast cancer cells. Mutat Res. 2011;706(1-2):28–35. doi:10.1016/j.mrfmmm.2010.10.007.
Banerjee S, Kaseb AO, Wang ZW, Kong DJ, Mohammad M, Padhye S, et al. Antitumor activity of gemcitabine and oxaliplatin is augmented by thymoquinone in pancreatic cancer. Cancer Res. 2009;69(13):5575–83. doi:10.1158/0008-5472.Can-08-4235.
Sethi G, Ahn KS, Aggarwal BB. Targeting nuclear factor-kappa B activation pathway by thymoquinone: role in suppression of antiapoptotic gene products and enhancement of apoptosis. Mol Cancer Res. 2008;6(6):1059–70. doi:10.1158/1541-7786.MCR-07-2088.
Gali-Muhtasib HU, Abou Kheir WG, Kheir LA, Darwiche N, Crooks PA. Molecular pathway for thymoquinone-induced cell-cycle arrest and apoptosis in neoplastic keratinocytes. Anti-Cancer Drugs. 2004;15(4):389–99. doi:10.1097/01.cad.0000125054.43188.56.
Guicciardi ME, Gores GJ. Life and death by death receptors. Faseb J. 2009;23(6):1625–37. doi:10.1096/Fj.08.
Mohamed A, Afridi DM, Garani O, Tucci M. Thymoquinone inhibits the activation of NF-kappaB in the brain and spinal cord of experimental autoimmune encephalomyelitis. Biomed Sci Instrum. 2005;41:388–93.
Connelly L, Barham W, Onishko HM, Sherrill T, Chodosh LA, Blackwell TS, et al. Inhibition of NF-kappa B activity in mammary epithelium increases tumor latency and decreases tumor burden. Oncogene. 2011;30(12):1402–12. doi:10.1038/onc.2010.521.
Tekeoglu I, Dogan A, Ediz L, Budancamanak M, Demirel A. Effects of thymoquinone (volatile oil of black cumin) on rheumatoid arthritis in rat models. Phytother Res. 2007;21(9):895–7. doi:10.1002/ptr.2143.
Chehl N, Chipitsyna G, Gong Q, Yeo CJ, Arafat HA. Anti-inflammatory effects of the Nigella sativa seed extract, thymoquinone, in pancreatic cancer cells. HPB (Oxford). 2009;11(5):373–81. doi:10.1111/j.1477-2574.2009.00059.x.
Ravindran J, Nair HB, Sung B, Prasad S, Tekmal RR, Aggarwal BB. Thymoquinone poly(lactide-co-glycolide) nanoparticles exhibit enhanced anti-proliferative, anti-inflammatory, and chemosensitization potential. Biochem Pharmacol. 2010;79(11):1640–7. doi:10.1016/j.bcp.2010.01.023.
Sakalar C, Yuruk M, Kaya T, Aytekin M, Kuk S, Canatan H. Pronounced transcriptional regulation of apoptotic and TNF-NF-kappa-B signaling genes during the course of thymoquinone mediated apoptosis in HeLa cells. Mol Cell Biochem. 2013;383(1-2):243–51. doi:10.1007/s11010-013-1772-x.
Rajput S, Kumar BNP, Dey KK, Pal I, Parekh A, Mandal M. Molecular targeting of Akt by thymoquinone promotes G(1) arrest through translation inhibition of cyclin D1 and induces apoptosis in breast cancer cells. Life Sci. 2013;93(21):783–90. doi:10.1016/j.lfs.2013.09.009.
Sutton KM, Greenshields AL, Hoskin DW, Thymoquinone A. Bioactive component of black caraway seeds, causes G1 phase cell cycle arrest and apoptosis in triple-negative breast cancer cells with mutant p53. Nutr Cancer Int J. 2014;66(3):408–18. doi:10.1080/01635581.2013.878739.
Woo CC, Hsu A, Kumar AP, Sethi G, Tan KHB. Thymoquinone inhibits tumor growth and induces apoptosis in a breast cancer xenograft mouse model: the role of p38 MAPK and ROS. PLoS One. 2013;8(10):e75356. doi:10.1371/journal.pone.0075356.
Attoub S, Sperandio O, Raza H, Arafat K, Al-Salam S, Al Sultan MA, et al. Thymoquinone as an anticancer agent: evidence from inhibition of cancer cells viability and invasion in vitro and tumor growth in vivo. Fund Clin Pharmacol. 2013;27(5):557–69. doi:10.1111/j.1472-8206.2012.01056.x.
Velho-Pereira R, Kumar A, Pandey BN, Jagtap AG, Mishra KP. Radiosensitization in human breast carcinoma cells by thymoquinone: role of cell cycle and apoptosis. Cell Biol Int. 2011;35(10):1025–9. doi:10.1042/Cbi20100701.
Effenberger-Neidnicht K, Schobert R. Combinatorial effects of thymoquinone on the anti-cancer activity of doxorubicin. Cancer Chemother Pharmacol. 2011;67(4):867–74. doi:10.1007/s00280-010-1386-x.
Wani MC, Taylor HL, Wall ME, Coggon P, McPhail AT. Plant antitumor agents. VI. The isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J Am Chem Soc. 1971;93(9):2325–7.
Schiff PB, Fant J, Horwitz SB. Promotion of microtubule assembly in vitro by taxol. Nature. 1979;277(5698):665–7.
Erdemoğlu N, Bilge S. The antitumor effects of the taxane class compounds. Ankara Ecz Fak Derg. 2000;29(1):77–90.
Dirican A, Atmaca H, Bozkurt E, Erten C, Karaca B, Uslu R. Novel combination of docetaxel and thymoquinone induces synergistic cytotoxicity and apoptosis in DU-145 human prostate cancer cells by modulating PI3K-AKT pathway. Clin Transl Oncol. 2015;17(2):145–51. doi:10.1007/s12094-014-1206-6.
Lettre R, Paweletz N, Werner D, Granzow C. Sublines of the Ehrlich-Lettre mouse ascites tumour. A new tool for experimental cell research. Naturwissenschaften. 1972;59(2):59–63.
Dexter DL, Kowalski HM, Blazar BA, Fligiel Z, Vogel R, Heppner GH. Heterogeneity of tumor cells from a single mouse mammary tumor. Cancer Res. 1978;38(10):3174–81.
Heppner GH, Miller FR, Shekhar PM. Nontransgenic models of breast cancer. Breast Cancer Res. 2000;2(5):331–4.
Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44–57. doi:10.1038/nprot.2008.211.
Huang DW, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009;37(1):1–13. doi:10.1093/Nar/Gkn923.
Bai T, Yang Y, Wu YL, Jiang S, Lee JJ, Lian LH, et al. Thymoquinone alleviates thioacetamide-induced hepatic fibrosis and inflammation by activating LKB1-AMPK signaling pathway in mice. Int Immunopharmacol. 2014;19(2):351–7. doi:10.1016/j.intimp.2014.02.006.
Ghazwani M, Zhang YF, Gao X, Fan J, Li J, Li S. Anti-fibrotic effect of thymoquinone on hepatic stellate cells. Phytomedicine. 2014;21(3):254–60. doi:10.1016/j.phymed.2013.09.014.
Ashour AE, Abd-Allah AR, Korashy HM, Attia SM, Alzahrani AZ, Saquib Q, et al. Thymoquinone suppression of the human hepatocellular carcinoma cell growth involves inhibition of IL-8 expression, elevated levels of TRAIL receptors, oxidative stress and apoptosis. Mol Cell Biochem. 2014;389(1-2):85–98. doi:10.1007/s11010-013-1930-1.
Badr G, Mohany M, Abu-Tarboush F. Thymoquinone decreases F-actin polymerization and the proliferation of human multiple myeloma cells by suppressing STAT3 phosphorylation and Bcl2/Bcl-(XL) expression. Lipids Health Dis. 2011. doi:10.1186/1476-511x-10-236.
Badr G, Lefevre EA, Mohany M. Thymoquinone inhibits the CXCL12-induced chemotaxis of multiple myeloma cells and increases their susceptibility to Fas-mediated apoptosis. PLoS One. 2011;6(9):e23741. doi:10.1371/journal.pone.0023741.
Li F, Rajendran P, Sethi G. Thymoquinone. Br J Pharmacol. 2010;161(3):541–54. doi:10.1111/j.1476-5381.2010.00874.x.
Rajput S, Kumar BNP, Banik P, Parida S, Mandal M. Thymoquinone restores radiation-induced TGF-beta expression and abrogates EMT in chemoradiotherapy of breast cancer cells. J Cell Physiol. 2015;230(3):620–9. doi:10.1002/Jcp.24780.
Kundu J, Choi BY, Jeong CH, Kundu JK, Chun KS. Thymoquinone induces apoptosis in human colon cancer HCT116 cells through inactivation of STAT3 by blocking JAK2- and Src-mediated phosphorylation of EGF receptor tyrosine kinase. Oncol Rep. 2014;32(2):821–8. doi:10.3892/Or.2014.3223.
Chou J, Provot S, Werb Z. GATA3 in development and cancer differentiation: cells GATA have it! J Cell Physiol. 2010;222(1):42–9. doi:10.1002/Jcp.21943.
Radpour R, Kohler C, Haghighi MM, Fan AXC, Holzgreve W, Zhong XY. Methylation profiles of 22 candidate genes in breast cancer using high-throughput MALDI-TOF mass array. Oncogene. 2009;28(33):2969–78. doi:10.1038/Onc.2009.149.
Fujii H, Biel MA, Zhou W, Weitzman SA, Baylin SB, Gabrielson E. Methylation of the HIC-1 candidate tumor suppressor gene in human breast cancer. Oncogene. 1998;16(16):2159–64. doi:10.1038/sj.onc.1201976.
Yusuf RZ, Duan Z, Lamendola DE, Penson RT, Seiden MV. Paclitaxel resistance: molecular mechanisms and pharmacologic manipulation. Curr Cancer Drug Targets. 2003;3(1):1–19.
Ajabnoor GMA, Crook T, Coley HM. Paclitaxel resistance is associated with switch from apoptotic to autophagic cell death in MCF-7 breast cancer cells. Cell Death Dis. 2012;3:e260. doi:10.1038/cddis.2011.139.
Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000;28(1):27–30.
Acknowledgments
This work was supported by The Scientific and Technological Research Council of Turkey (TUBİTAK), Grant Number: 113S322 (C. SAKALAR) and Erciyes University Scientific Research Fund (EU-BAP), Grant Number: TOA-2014-4877 (C. SAKALAR). Authors would like to thank to Prof. Dr. Nedime Serakinci, Near East University, Cyprus and Prof. Dr. Feridoun Karimi-Busheri University of Alberta, Canada for critically reading the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
None
Ethics approval
Ethical approval for the study was obtained from Erciyes University Animal Researche Local Ethics Committee and the ethic regulations have been followed in accordance with the international, national, and institutional guidelines.
Rights and permissions
About this article

Cite this article
Şakalar, Ç., İzgi, K., İskender, B. et al. The combination of thymoquinone and paclitaxel shows anti-tumor activity through the interplay with apoptosis network in triple-negative breast cancer. Tumor Biol. 37, 4467–4477 (2016). https://doi.org/10.1007/s13277-015-4307-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13277-015-4307-0