
Abstract
Although Inonotus linteus complex is placed in the genus Inonotus, its perennial basidiocarps with a dimitic hyphal system, at least in the trama, distinguish the complex from other species in the genus, which have an annual habit and a monomitic hyphal system. The species number of Inonotus linteus complex has been increased in China and tropical America in recent publications. However, whether species in this complex belong to Inonotus has not specifically been addressed. To explore the phylogenetic placement of species of Inonotus linteus complex, we performed a comprehensive study using morphological and phylogenetic data based on global samples. nLSU (53 with 13 new) and ITS (70 with 18 new) datasets were used to produce the phylogenetic results. Taking into consideration the phylogenies inferred from the nLSU and ITS datasets, Inonotus is a polyphyletic genus comprising at least three clades. Clade A, the core Inonotus clade, contains the generic type, Inonotus hispidus, while Clades B and C comprise species from the Inonotus linteus complex. Morphological and phylogenetic evidence indicates that Clades B and C are new genera, and Sanghuangporus and Tropicoporus are introduced in this study. Ten species are transferred to Sanghuangporus and seven to Tropicoporus. Tropicoporus excentrodendri and T. guanacastensis spp. nov. are described, and their distinctive characters are discussed. Keys to the two new genera and the Inonotus sensu stricto, and to species of each new genus are provided.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Inonotus P. Karst. (Hymenochaetaceae), was introduced by Karsten (1879) and Inonotus hispidus (Bull.) P. Karst. was later selected as its type species (Donk 1960). Inonotus sensu lato is traditionally characterized by annual basidiocarps and a monomitic hyphal system and includes more than 100 species (Ryvarden 2005). These morphological characteristics can distinguish Inonotus from Phellinus Quél., another large genus within Hymenochaetaceae that often produces perennial basidiocarps with a dimitic hyphal system (Larsen and Cobb-Poulle 1990). Molecular studies have indicated that the delimitation between the two genera is weak, and several species of Phellinus, viz. Phellinus baumii Pilát, P. linteus (Berk. & M.A. Curtis) Teng, P. tropicalis M.J. Larsen & Lombard, P. vaninii Ljub. and P. weirianus (Bres.) Gilb. were transferred to Inonotus (Wagner and Fischer 2002). Later Dai (2010), Wu et al. (2012) and Vlasák et al. (2013), respectively, combined Phellinus lonicericola Parmasto, P. lonicerinus (Bondartsev) Bondartsev & Singer and P. dependens (Murrill) Ryvarden to Inonotus. These are members of the Inonotus linteus (Berk. & M.A. Curtis) Teixeira complex.
Previous phylogenetic analyses (Wagner and Fischer 2002; Dai 2010) clearly indicated that Inonotus sensu lato had polyphyletic origins, and four more narrowly defined genera, namely Inocutis Fiasson & Niemelä, Inonotopsis Parmasto, Mensularia Lázaro Ibiza and Onnia P. Karst., were segregated from Inonotus. The Inonotus linteus complex was considered to belong to the reduced Inonotus (Wagner and Fischer 2002; Dai 2010). However, this opinion is questionable, because no comprehensive phylogeny study has specifically focused on the reduced Inonotus. Taxa in the Inonotus linteus complex also differ distinctively from other species of Inonotus. Several new taxa from East Asia and tropical America belonging to the Inonotus linteus complex have recently been introduced (Wu et al. 2012; Tian et al. 2013; Vlasák et al. 2013), which brings the number of species in this complex to 15 (Table 1). In our previous phylogenetic analyses, taxa of the Inonotus linteus complex formed two distinctly separate lineages (Tian et al. 2013; Vlasák et al. 2013). However, the phylogenetic relationships between the Inonotus linteus complex, especially those African members, as well as other species of Inonotus are poorly investigated.
The aim of this study is to investigate the diversity and taxonomy of the Inonotus linteus complex using a worldwide set of collections and to elucidate the phylogenetic position of the Inonotus linteus complex.
Materials and methods
Morphological studies
The studied specimens are deposited at the herbaria of the Institute Applied of Ecology, Chinese Academy of Sciences (IFP), the Institute of Microbiology, Beijing Forestry University (BJFC), the Botanical Museum, University of Oslo (O), the National Museum of Natural Science, Taichung, Taiwan (TNM), the Mycothèque de l’Universitè catholique de Louvain (MUCL), the Farlow Reference Library and Herbarium of Cryptogamic Botany, Harvard University (FH), and the private herbarium of J. Vlasák (JV). Microscopic procedures follow Dai (2010). Sections were prepared in Cotton Blue (CB), Melzer’s reagent (IKI) and 5 % potassium hydroxide (KOH), and studied using a Nikon Eclipse 80i microscope at magnifications of up to 1000×. The measurements and drawings were made from sections stained with CB. When presenting the size range of basidiospores, 5 % of the measurements were excluded from each end of the range and are given in parentheses. Drawings were made with the aid of a drawing tube. Special colour terms follow Petersen (1996). The abbreviations are used as follows: L = mean basidiospore length (arithmetical average of all basidiospores), W = mean basidiospore width (arithmetical average of all basidiospores), Q = variation in the L/W ratios between the specimens studied, n = the number of basidiospores measured from given number of specimens, IKI– = neither amyloid nor dextrinoid, CB+ = cyanophilous and CB– = acyanophilous.
Molecular study
About 1 mm3 of the dried specimens collected from China and culture WD 1838 from the Forestry and Forest Products Research Institute, Ibaraki, Japan (FFPRI) were directly employed to PCR amplification using Phire® Plant Direct PCR Kit (Finnzymes Oy, Finland) according to the manufacturer’s instruction. The PCR procedure included these steps: initial denaturation at 98 °C for 5 min, followed by 39 cycles denaturation at 98 °C for 5 s, annealing at 48 °C for 5 s for nLSU region/59 °C for 5 s for ITS region and extension at 72 °C for 5 s, and a final extension of 72 °C for 10 min. The primer pairs LR0R and LR7 (Vilgalys and Hester 1990), and ITS5 and ITS4 (White et al. 1990) were, respectively, used in the amplifications of nLSU and ITS regions, and also to sequence them in Beijing Genomics Institute, China. In addition, primers LR3R and LR3 (Vilgalys and Hester 1990) were sometimes used to sequence the nLSU region as well.
DNA was isolated from 0.02 to 0.2 g of the dried basidiocarps of non Chinese specimens using CTAB/NaCl extraction, followed by repeated extractions with chloroform and isopropanol precipitation. Crude DNA was purified using Wizard Clean Up kit PROMEGA and was diluted as a template for subsequent amplifications. nLSU sequences were amplified with primers LR0R and LR7, while ITS sequences used primers ITS5 and ITS4. Annealing temperature is 55 °C for both regions. Amplified DNA sequences were sequenced with the same primers in the Genomics Laboratory of Biology Centre, Academy of Sciences of the Czech Republic, České Budějovice, on ABI 3730xl DNA analyzer, using BigDye Terminator 3.1 kit. All newly generated sequences were deposited at GenBank (http://www.ncbi.nlm.nih.gov/; Table 2)
Phylogenetic analysis
The new sequences generated for this study, and additional nLSU and ITS sequences (Figs. 1 and 2) retrieved from GenBank were assembled for phylogenetic analysis. The nLSU dataset was first used to explore the phylogeny of Inonotus sensu lato. Besides Inonotus species, other representative species of Inocutis, Inonotopsis, Mensularia, Fomitiporella Murrill, Fulvifomes Murrill and Phylloporia Murrill were included in the dataset. ITS dataset, including more representatives of the Inonotus linteus complex, were used to further elucidate the phylogenetic relationship between Inonotus linteus complex and other species of Inonotus. Phellinus laevigatus (P. Karst.) Bourdot & Galzin and P. populicola Niemelä were selected as outgroup taxa for both datasets (Wagner and Fischer 2002). The two datasets were aligned using MAFFT 7 (Katoh et al. 2002) with Q-INS-i option (Katoh and Toh 2008). The resulting alignments are deposited at TreeBASE (http://purl.org/phylo/treebase; submission ID S16557).

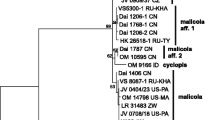
Phylogeny of Inonotus sensu lato inferred from nLSU sequences. Topology is from maximum likelihood analysis, while bootstrap values from maximum likelihood analysis and Bayesian inference (simultaneously not below 50 % and 0.8, respectively). The newly generated sequences for this study are in boldface. The species names in brackets were names before this study

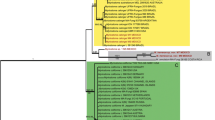
Phylogeny of Inonotus sensu lato inferred from ITS sequences. Topology is from maximum likelihood analysis, while bootstrap values from maximum likelihood analysis and Bayesian inference (simultaneously not below 50 % and 0.8, respectively). The newly generated sequences for this study are in boldface. The species names in brackets were names before this study
When performing phylogenetic analyses of the alignments, all characters equally weighted and gaps were treated as missing. The maximum likelihood (ML) trees were constructed using raxmlGUI 1.2 (Stamatakis 2006; Silvestro and Michalak 2012) with GTR + I + G model and auto FC option (Pattengale et al. 2010) in bootstrap (BS) replicates. Bayesian inference (BI) was conducted using MrBayes 3.2 (Ronquist and Huelsenbeck 2003) following the best-fit evolutionary model estimated by jModelTest 2.1.4 (Guindon and Gascuel 2003; Darriba et al. 2012). Two independent runs were employed and each run with four chains started from random trees. Trees were sampled every 1000th generation. The first 25 % of sampled trees was set as burn-in, while the other trees were used to produce a 50 % majority consensus tree and for calculating Bayesian posterior probabilities (BPPs). Chain convergence was determined with Tracer 1.5 (http://tree.bio.ed.ac.uk/software/tracer/). The two phylogenetic methods resulted in similar topologies for each dataset. Therefore, only the ML tree is presented, and BS and BPP values simultaneously not below 50 % and 0.8, respectively, were indicated at the nodes.
Results
Molecular phylogeny
A total of 53 nLSU and 70 ITS sequences, including 13 nLSU and 18 ITS sequences newly generated in this study, were used in the phylogenetic analyses. The alignment, resulting from the nLSU dataset, contained 909 characters. Its best-fit evolutionary model for BI was estimated as GTR + I + G. All chains in BI converged after 10 000 000 generations, where the effective sample sizes (ESSs) of all parameters were greater than 2000 and the potential scale reduction factors (PSRFs) were equal to 1.000. The BS search for ML analysis stopped after 250 replicates.
The nLSU-based phylogeny (Fig. 1) recovered Inonotus as monophyletic with moderate support in ML and full support in BI. Within the genus Inonotus, I. andersonii (Ellis & Everh.) Nikol., I. chihshanyenus T.T. Chang & W.N. Chou, I. compositus Han C. Wang, I. cuticularis (Bull.) P. Karst., I. glomeratus (Peck) Murrill, I. hispidus (the generic type), I. nidus-pici Pilát, I. obliquus (Ach. ex Pers.) Pilát, I. pachyphloeus (Pat.) T. Wagner & M. Fisch., I. pegleri Ryvarden, I. plorans (Pat.) Bondartsev & Singer, I. pruinosus Bondartsev, I. quercustris M. Blackw. & Gilb., I. rodwayi D.A. Reid and I. ulmicola Corfixen formed a clade that we term Clade A here. Species of Inonotus linteus complex clustered in two other clades. The Clade B comprised Inonotus cubensis, I. linteus, I. tropicalis and one unidentified collection, while the Clade C comprised I. alpinus, I. baumii, I. lonicericola, I. sanghuang, I. vaninii, I. weigelae, I. weirianus, I. zonatus, Phellinus microcystideus Har. & Pat., and one unidentified collection.
The ITS dataset resulted in an alignment of 1091 characters. The best-fit evolutionary model for BI was HKY + I + G. After 1 000 000 generations, the ESSs of all parameters were greater than 2000 and the PSRFs were close to 1.000, indicating all chains converged. The BS search for ML analysis stopped after 150 replicates.
The phylogeny inferred from the ITS dataset (Fig. 2), accommodating more representatives of the Inonotus linteus complex than the nLSU dataset, clearly indicated that Inonotus was a polyphyletic genus. The ITS dataset also showed that species of Inonotus formed at least three clades. The Clade A, receiving low support (64 % in ML, 0.61 in BI), comprised I. andersonii, I. compositus, I. henanensis Juan Li & Y.C. Dai, I. hispidus, I. latemarginatus Y.C. Dai, I. micantissimus (Rick) Rajchenb., I. obliquus, I. pachyphloeus, I. patouillardii (Rick) Imazeki, I. quercustris and I. rickii (Pat.) D.A. Reid. Species of Inonotus linteus complex clustered outside the Clade A; Clade B comprised I. dependens, I. pseudolinteus, I. sideroxylicola, Xanthochrous rudis Pat. and three unidentified lineages, besides I. cubensis, I. linteus and I. tropicalis also indicated in the nLSU-based phylogeny. Clade C additionally comprised I. lonicerinus and Phellinus microcystideus, and two unidentified collections compared with that in the nLSU-based phylogeny. In addition, Inonotus glomeratus and I. tricolor (Bres.) Y.C. Dai clustered outside the three clades.
Based on the phylogenetic evidence and morphology, we introduced two new genera to accommodate species of the Inonotus linteus complex. In addition, two new species are described and 17 new combinations are proposed for these two new genera.
Taxonomy
Key to Inonotus sensu stricto , Sanghuangporus and Tropicoporus
-
1. Hyphal system monomitic in both context/subiculum and trama·························································································································································Inonotus sensu stricto
-
1. Hyphal system monomitic to dimitic in context/subiculum, dimitic in trama·······················2
-
2. Basidiocarps perennial, effused-reflexed to pileate; basidiospores when ellipsoid mostly < 3.5 μm wide, when subglobose > 3.5 μm wide; in boreal, temperate, subtropical to tropical zones············································································································Sanghuangporus
-
2. Basidiocarps annual to perennial, resupinate, effused-reflexed to pileate; basidiospores when ellipsoid mostly > 3.5 μm wide, when subglobose mostly < 3.5 μm wide; exclusively in tropical zone···············································································Tropicoporus
Sanghuangporus Sheng H. Wu, L.W. Zhou & Y.C. Dai, gen. nov.
MycoBank no.: MB 812143
Type species. Sanghuangporus sanghuang (Sheng H. Wu, T. Hatt. & Y.C. Dai) Sheng H. Wu, L.W. Zhou & Y.C. Dai (combined below).
Etymology. Sanghuangporus (Lat.): refers to the species of this genus called as ‘sanghuang’ in Chinese herbal markets.
Basidiocarps perennial, effused-reflexed to pileate. Pileal surface brown, dark greyish to black, hispid, tomentose, velutinate to glabrous with a radially cracked crust. Pore surface yellow to brown. Context homogeneous to duplex with a black line. Hyphal system monomitic to dimitic in context, dimitic in trama; generative hyphae simple septate; tissues darkening in KOH, hyphae unchanged in KOH. Hymenial setae present, hyphoid setae absent. Basidiospores ellipsoid, broadly ellipsoid to subglobose, yellowish, slightly thick- to thick-walled, smooth, IKI–, CB– or slightly CB+. On angiosperm wood, causing a white rot. In boreal, temperate, subtropical to tropical zones.
Sanghuangporus alpinus (Y.C. Dai & X.M. Tian) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812144
Basionym. Inonotus alpinus Y.C. Dai & X.M. Tian, in Tian et al., Fungal Diversity 58: 162. 2013.
Sanghuangporus baumii (Pilát) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812145
Basionym. Phellinus baumii Pilát, Bull. trimest. Soc. mycol. Fr. 48(1): 25. 1932.
Synonym. Inonotus baumii (Pilát) T. Wagner & M. Fisch., Mycologia 94(6): 1009. 2002.
Sanghuangporus lonicericola (Parmasto) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812146
Basionym. Phellinus lonicericola Parmasto, in Parmasto & Parmasto, Folia cryptog. Estonica 38: 59. 2001.
Synonym. Inonotus lonicericola (Parmasto) Y.C. Dai, Fungal Diversity 45: 276. 2010.
Sanghuangporus lonicerinus (Bondartsev) Sheng H. Wu, L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812147
Basionym. Fomes lonicerinus Bondartsev, Acta Inst. Bot. Acad. Sci. USSR Plant. Crypt., Ser. II: no. 500. 1935.
Synonym. Inonotus lonicerinus (Bondartsev) Sheng H. Wu, Y.C. Dai & T. Hatt., in Wu et al., Botanical Studies 53(1): 140. 2012.
Sanghuangporus microcystideus (Har. & Pat.) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812148
Basionym. Phellinus microcystideus Har. & Pat., Bull. Mus. Hist. Nat., Paris 15: 90. 1909.
Sanghuangporus sanghuang (Sheng H. Wu, T. Hatt. & Y.C. Dai) Sheng H. Wu, L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812149
Basionym. Inonotus sanghuang Sheng H. Wu, T. Hatt. & Y.C. Dai, in Wu et al., Botanical Studies 53(1): 141. 2012.
Sanghuangporus vaninii (Ljub.) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812150
Basionym. Phellinus vaninii Ljub., Bot. Mater. Otd. Spor. Rast. 15: 115. 1962.
Synonym. Inonotus vaninii (Ljub.) T. Wagner & M. Fisch., Mycologia 94(6): 1009. 2002.
Sanghuangporus weigelae (T. Hatt. & Sheng H. Wu) Sheng H. Wu, L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812151
Basionym. Inonotus weigelae T. Hatt. & Sheng H. Wu, in Wu et al., Botanical Studies 53(1): 143. 2012.
Synonym. Inonotus tenuicontextus L.W. Zhou & W.M. Qin, Mycol. Progr. 11(3): 793. 2012.
Sanghuangporus weirianus (Bres.) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812152
Basionym. Fomes weirianus Bres., Stud. Trent., Classe II, Sci. Nat. Econ. 7(1): 5. 1926.
Synonym. Inonotus weirianus (Bres.) T. Wagner & M. Fisch., Mycologia 94(6): 1009. 2002.
Sanghuangporus zonatus (Y.C. Dai & X.M. Tian) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812153
Basionym. Inonotus zonatus Y.C. Dai & X.M. Tian, in Tian et al., Fungal Diversity 58(1): 165. 2013.
Key to species of Sanghuangporus
-
1. Pores 4–5 per mm·······························································································S. lonicerinus
-
1. Pores > 5 per mm···················································································································2
-
2. Pores 8–10 per mm····························································································S. lonicericola
-
2. Pores 5–8 per mm···················································································································3
-
3. Context thin, < 3 mm thick····································································································4
-
3. Context thick, > 10 mm thick·································································································5
-
4. Context duplex; distributed in the warm temperate zones·····································S. weigelae
-
4. Context homogeneous; distributed in alpine zones·················································S. alpinus
-
5. Setae mostly < 20 μm long·····································································································6
-
5. Setae mostly > 20 μm long····································································································8
-
6. Basidiocarps with a sharp edge················································································S. zonatus
-
6. Basidiocarps with an obtuse edge··························································································7
-
7. Basidiospores broadly ellipsoid, mostly < 4.5 μm long, < 3.5 μm wide···················S. baumii
-
7. Basidiospores subglobose, mostly > 5 μm long, > 4 μm wide·····················S. microcystideus
-
8. Basidiospores basically subglobose; restricted to Juglans····································S. weirianus
-
8. Basidiospores basically broadly ellipsoid; restricted to Morus or Populus··························9
-
9. Basidiocarps resupinate to pileate; basidiospores 3.8–4.4 × 2.8–3.7 μm; restricted to Populus····················································································································S. vaninii
-
9. Basidiocarps distinctly pileate; basidiospores 4–4.9 × 3.1–3.9 μm; restricted to Morus····································································································································S. sanghuang
Tropicoporus L.W. Zhou, Y.C. Dai & Sheng H. Wu, gen. nov.
MycoBank no.: MB 812154
Type species. Tropicoporus excentrodendri L.W. Zhou & Y.C. Dai (described below).
Etymology. Tropicoporus (Lat.): refers to a distribution of the genus exclusively in tropics.
Basidiocarps annual to perennial, resupinate, effused-reflexed to pileate. Pileal surface brown, dark greyish to black, hispid, tomentose, velutinate to glabrous with a radially cracked crust. Pore surface yellow to brown. Context homogeneous to duplex with a black line. Hyphal system monomitic to dimitic in context/subiculum, dimitic in trama; generative hyphae simple septate; tissues darkening in KOH, hyphae unchanged or swollen in KOH. Hymenial setae present, hyphoid setae absent. Basidiospores ellipsoid, broadly ellipsoid to subglobose, yellowish, slightly thick- to thick-walled, smooth, IKI–, CB–. On angiosperm wood, causing a white rot. In tropical zone exclusively.
Tropicoporus excentrodendri L.W. Zhou & Y.C. Dai, sp. nov. Figs. 3 and 4

A basidiocarp of Tropicoporus excentrodendri (holotype). — Scale bar = 1 cm

Microscopic structures of Tropicoporus excentrodendri (drawn from the holotype). a. Basidiospores; b. Basidia and basidioles; c. Hymenial setae; d. Hyphae from trama; e. Hyphae from context
MycoBank no.: MB 812155
Basidiocarps annual, resupinate, effused-reflexed to distinctly pileate, imbricate. Pileal surface tomentum to hispid, concentrically sulcate; margin obtuse, tomentose to hispid. Pores circular to angular, 7–8 per mm; dissepiments thick, entire. Context duplex with a black line. Hyphal system dimitic in both context and trama; hyphae unchanged in KOH. Hymenial setae occasionally present, 20–25 × 5–8 μm. Basidiospores broadly ellipsoid to subglobose, pale yellowish, slightly thick-walled, IKI–, CB–, 3.4–4 × 2.9–3.6 μm.
Holotype. CHINA . Guangxi, Longzhou County, Nonggang Nature Reserve, on stump of Excentrodendron tonkinense, 22 July 2012, Yuan 6232 (IFP).
Etymology. excentrodendri (Lat.): refers to the host genus Excentrodendron.
Basidiocarps annual, resupinate, effused-reflexed to distinctly pileate, imbricate, without odour or taste and woody hard when fresh, woody hard and light in weight when dry. Pilei dimidiate, projecting up to 2 cm, 4.5 cm wide and 0.7 cm thick at base. Pileal surface vinaceous brown to fuscous, tomentum to hispid, concentrically sulcate; margin obtuse, hispid, yellow-brown to honey-yellow. Pore surface buff to honey-yellow, glancing; sterile margin distinct, honey-yellow, up to 3 mm; pores circular to angular, 7–8 per mm; dissepiments thick, entire. Context woody hard, up to 2 mm, duplex, with a black line between upper tomentum part and lower hard context, upper tomentum yellow-brown, soft, up to 1 mm thick, lower context cinnamon-buff, woody hard, up to 1 mm thick. Tubes curry-yellow, woody hard, up to 5 mm long; white mycelial strands occasionally present in old tubes.
Hyphal system dimitic; generative hyphae simple septate; tissue darkening but otherwise unchanged in KOH. Context. Generative hyphae pale yellowish, thick-walled, occasionally branched, frequently septate, straight, 2–3.5 μm in diam.; skeletal hyphae yellowish, distinctly thick-walled with a narrow lumen, unbranched, aseptate, straight, 3–5 μm in diam., more or less regularly arranged. Hyphae in the tomentum or hispid similar to those in context, but loosely interwoven. Tubes. Generative hyphae hyaline to pale yellowish, slightly thick-walled, unbranched, frequently septate, 2.5–3.5 μm in diam.; skeletal hyphae dominant, yellowish, thick-walled with a wide to narrow lumen, unbranched, aseptate, subparallel along the tubes, 3–4.5 μm in diam.
Hymenial setae occasionally present, ventricose, dark brown, thick-walled, with a sharp or obtuse tip, 20–25 × 5–8 μm; cystidia and cystidioles absent; basidia barrel-shaped, with four sterigmata and a simple septum at the base, 10–15 × 4–7 μm; basidioles in shape similar to basidia, but slightly smaller; rhomboid crystals abundant.
Basidiospores broadly ellipsoid to subglobose, pale yellowish, slightly thick-walled, smooth, IKI–, CB–, (3.2–)3.4–4(−4.2) × (2.7–)2.9–3.6(−3.8) μm, L = 3.74 μm, W = 3.21 μm, Q = 1.16–1.17 (n = 60/2).
Additional specimens (paratypes) examined. CHINA. Guangxi, Longzhou County, Nonggang Nature Reserve, on living Excentrodendron tonkinense, 22 July 2012, Yuan 6227 (IFP); on dead branch of living Excentrodendron tonkinense, 22 July 2012, Yuan 6229 (IFP); on stump of Excentrodendron tonkinense, 22 July 2012, Yuan 6231 & 6234 (IFP).
Remarks. Tropicoporus excentrodendri is characterized by its annual, resupinate, effused-reflexed to pileate basidiocarps, and small pores and basidiospores within the genus Tropicoporus. Tropicoporus tropicalis (M.J. Larsen & Lombard) L.W. Zhou & Y.C. Dai (combined below) is the other species of Tropicoporus with annual, resupinate basidiocarps, and shares small pores (7–9 per mm) and basidiospores (3–4.5 × 2.5–4 μm) with T. excentrodendri, but its basidiocarps are exclusively resupinate and grow on other angiosperms other than Excentrodendron tonkinense in Brazil (Larsen and Cobb-Poulle 1990).
Tropicoporus excentrodendri is similar to Sanghuangporus weigelae that has effused-reflexed to pileate basidiocarps, duplex context separated by a black line, 6–8 per mm of pores, a dimitic hyphal system in both context and trama, hymenial setae, and 3.8–4.2 × 3–3.6 μm of basidiospores (Wu et al. 2012). However, Sanghuangporus weigelae has a perennial habit, obtuse pileal margin, and growth on Weigela in warm temperate zones (Wu et al. 2012; Zhou and Qin 2012).
Tropicoporus guanacastensis L.W. Zhou, Y.C. Dai & Vlasák, sp. nov. Figs. 5 and 6

A basidiocarp of Tropicoporus guanacastensis (holotype). — Scale bar = 1 cm

Microscopic structures of Tropicoporus guanacastensis (drawn from the holotype). a. Basidiospores; b. Basidia and basidioles; c. Hymenial setae; d. Hyphae from trama; e. Hyphae from context
MycoBank no.: MB 812156
Basidiocarps perennial, pileate, single. Pileal surface radially cracked, concentrically zonate and sulcate; margin sharp to obtuse. Pores circular to angular, 7–8 per mm; dissepiments thin to thick, entire. Context homogeneous. Tube layers distinctly stratified. Hyphal system monomitic in context and dimitic in trama; hyphae becoming swollen in KOH. Hymenial setae occasionally present, 12–25 × 4–8 μm. Basidiospores ellipsoid, pale yellowish, slightly thick-walled, IKI–, CB–, 4.1–4.9 × 3.3–3.9 μm.
Holotype. COSTA RICA. Guanacaste, Santa Rosa National Park, on dead standing angiosperm tree, 2 August 2014, JV 1408–25 (holotype in IFP, isotype in JV).
Etymology. guanacastensis (Lat.): refers to the province name of type locality in Costa Rica.
Basidiocarps perennial, pileate, single, without odour or taste and woody hard. Pilei dimidiate, applanate, projecting up to 7 cm, 17 cm wide and 4 cm thick at base. Pileal surface black, radially cracked, concentrically zonate and sulcate, covered by mosses with age; margin sharp to obtuse, dark grey. Pore surface cinnamon-buff to honey-yellow, slightly glancing; sterile margin distinct, honey-yellow, up to 1 mm wide; pores circular to angular, 7–8 per mm; dissepiments thin to thick, entire. Context dark brown, woody hard, up to 5 mm. Tubes dark brown, woody hard, up to 3.5 cm long, tube layers distinctly stratified, annual layer about 2 mm long.
Hyphal system monomitic in context and dimitic in trama; generative hyphae simple septate; tissue darkening in KOH, hyphae becoming swollen in KOH. Context. Generative hyphae brown, slightly thick-walled with a wide lumen, unbranched, frequently septate, 3.5–6.5 μm in diam., regularly arranged. Tubes. Generative hyphae hyaline to yellowish, thin- to slightly thick-walled, unbranched, frequently septate, 2–4 μm in diam.; skeletal hyphae dominant, brown, thick-walled with a narrow lumen to subsolid, unbranched, aseptate, parallel along the tubes, 3–4.5 μm in diam.
Hymenial setae occasionally present, mostly from skeletal hyphae, subulate to ventricose, dark brown, thick-walled, with a sharp tip, 12–25 × 4–8 μm; cystidia and cystidioles absent; basidia barrel-shaped, with four sterigmata and a simple septum at the base, 7–9 × 4–5 μm; basidioles in shape similar to basidia, but slightly smaller.
Basidiospores ellipsoid, pale yellowish, slightly thick-walled, smooth, IKI–, CB–, 4.1–4.9(−5) × (3.1–)3.3–3.9(−4) μm, L = 4.47 μm, W = 3.64 μm, Q = 1.22–1.23 (n = 60/2).
Additional specimens (paratypes) examined. COSTA RICA. Guanacaste, Santa Rosa National Park, on angiosperm wood, 12 June 1997, I. Lindblad 3101, (O 19228); Guanacaste, Estacion Experimental Forestal Horizontes, on dead standing angiosperm tree, 31 July 2014, JV 1407-103 J (JV, IFP).
Remarks. Tropicoporus guanacastensis resembles T. cubensis (Y.C. Dai, Decock & L.W. Zhou) L.W. Zhou & Y.C. Dai (combined below) by similar hyphal structure with skeletal hyphae becoming swollen in KOH and basidiospores in shape and size. Both species have origins from Central America; the former is from Costa Rica and the latter was described from Cuba (Tian et al. 2013). Tropicoporus cubensis differed from T. guanacastensis mainly in its indistinct tube layers, larger pores (5–6 per mm) and longer hymenial setae (27–43 × 5–10 μm; Tian et al. 2013).
Inonotus costaricensis Ryvarden was another species recently described from Costa Rica (Ryvarden 2002). This species is a typical member of Inonotus, producing annual basidiocarps and a monomitic hyphal system (Ryvarden 2002), and thus distinguish from Tropicoporus guanacastensis.
Tropicoporus cubensis (Y.C. Dai, Decock & L.W. Zhou) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812157
Basionym. Inonotus cubensis Y.C. Dai, Decock & L.W. Zhou, in Tian et al., Fungal Diversity 58(1): 163. 2013.
Tropicoporus dependens (Murrill) L.W. Zhou, Y.C. Dai & Vlasák, comb. nov.
MycoBank no.: MB 812158
Basionym. Pyropolyporus dependens Murrill, N. Amer. Fl. (New York) 9(2): 106. 1908.
Synonym. Inonotus dependens (Murrill) Vlasák & Y.C. Dai, in Vlasák et al., Phytotaxa 124(1): 27. 2013.
Tropicoporus linteus (Berk. & M.A. Curtis) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812159
Basionym. Polyporus linteus Berk. & M.A. Curtis, Proc. Amer. Acad. Arts & Sci. 4: 122. 1860 (1858).
Synonym. Inonotus linteus (Berk. & M.A. Curtis) Teixeira, Revista Brasileira de Botânicas 15: 126. 1992.
Tropicoporus pseudolinteus (Vlasák & Y.C. Dai) L.W. Zhou, Y.C. Dai & Vlasák, comb. nov.
MycoBank no.: MB 812160
Basionym. Inonotus pseudolinteus Vlasák & Y.C. Dai, in Vlasák et al., Phytotaxa 124(1): 30. 2013.
Tropicoporus rudis (Pat.) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812161
Basionym. Xanthochrous rudis Pat., Bull. Soc. mycol. Fr. 23: 83. 1907.
Tropicoporus sideroxylicola (Vlasák & Y.C. Dai) L.W. Zhou, Y.C. Dai & Vlasák, comb. nov.
MycoBank no.: MB 812162
Basionym. Inonotus sideroxylicola Vlasák & Y.C. Dai, in Vlasák et al., Phytotaxa 124(1): 32. 2013.
Tropicoporus tropicalis (M.J. Larsen & Lombard) L.W. Zhou & Y.C. Dai, comb. nov.
MycoBank no.: MB 812163
Basionym. Phellinus tropicalis M.J. Larsen & Lombard, Mycologia 80(1): 73. 1988.
Synonym. Poria rickii Bres., Annls mycol. 18(1/3): 37 (1920); Inonotus tropicalis (M.J. Larsen & Lombard) T. Wagner & M. Fisch., Mycologia 94(6): 1009. 2002.
Key to species of Tropicoporus
-
1. Basidiocarps annual, resupinate, effused-reflexed to distinctly pileate·································2
-
1. Basidiocarps perennial, pileate·······························································································3
-
2. Basidiocarps resupinate, effused-reflexed to distinctly pileate; on Excentrodendron tonkinense; in China···················································································T. excentrodendri
-
2. Basidiocarps exclusively resupinate; on other angiosperms; in Brazil·················T. tropicalis
-
3. The binding alike hyphae (frequently branched and winding) present in trama····················4
-
3. The binding alike hyphae (frequently branched and winding) absent in trama·····················5
-
4. Pores 5–7 per mm; basidiospores < 5 μm long, < 4 μm wide····························T. dependens
-
4. Pores 2–3 per mm; basidiospores > 6 μm long, > 4 μm wide······················T. sideroxylicola
-
5. Pores 4–5 per mm···························································································T. pseudolinteus
-
5. Pores > 5 per mm··················································································································6
-
6. Basidiospores mostly < 4.5 μm wide; in tropical America·····················································7
-
6. Basidiospores mostly > 4.5 μm wide; in tropical Africa·············································T. rudis
-
7. Pores 7–8 per mm, tube layers distinctly stratified; hymenial setae < 25 μm long ·····································································································································T. guanacastensis
-
7. Pores 5–7 per mm, tube layers indistinct; hymenial setae mostly > 25 μm long···················8
-
8. Dissepiments distinctly thick (≈ diameter of pores); basidiospores < 5 μm long, < 4 μm wide······················································································································T. cubensis
-
8. Dissepiments distinctly thin to slightly thick (<1/4 diameter of pores); basidiospores mostly > 5 μm long, mostly > 4 μm wide·······························································T. linteus
Other specimens examined.— Sanghuangporus microcystideus . CONGO. Oubangui, Grande Forêt (Holotype in FH). ETHIOPIA. Oromia Region, West Shewa, Menagesha National Forest, 11 July 2009, AM-08 (IFP); 26 March 2009, AM-19 (IFP). TANZANIA. Arusha Region, Arusha National Park, Mount Meru, on trunk of Olea africana, 18 February 1976, Harjula (O 915609).
— Tropicoporus rudis. NIGER . Maijirgui, 17 May 1906 (Holotype in FH). RWANDA . Eastern Province, Nyagatare District, Akagera National Park, Lake Hiema, 14 August 1976, J. Rammeloo (O 915614). TANZANIA. South Highl Province, Iringa District, Ruaha National Park, 21 May 1968, D.N. Pegler (O 915617).
Discussion
The traditional concept of Inonotus incorporated the poroid Hymenochaetaceae species with an annual habit and a monomitic hyphal system (Fiasson and Niemelä 1984). Using an nLSU-based phylogeny, Wagner and Fischer (2002) also treated some species with a perennial habit and a dimitic hyphal system, and several species bearing intermediate characters (an annual habit and a dimitic hyphal system, or a perennial habit and a monomitic hyphal system) in Inonotus. This opinion was accepted by Dai (2010), but Jeong et al. (2005) did not accept species without an annual habit and a monomitic hyphal system in Inonotus. Jeong et al. (2005) further considered taxa of Inonotus linteus complex to be a sister group of Inonotus. In this study, further samples of Inonotus, especially those from Inonotus linteus complex, were phylogenetically analyzed. Taking the phylogenies inferred from nLSU and ITS datasets, Inonotus sensu Wagner and Fischer (2002) and Dai (2010) was treated in a wide sense. Inonotus andersonii, I. chihshanyenus, I. compositus, I. cuticularis, I. glomeratus, I. henanensis, I. hispidus, I. latemarginatus, I. micantissimus, I. nidus-pici, I. obliquus, I. pachyphloeus, I. patouillardii, I. pegleri, I. plorans, I. pruinosus, I. quercustris, I. rickii, I. rodwayi, I. tricolor and I. ulmicola (21 species) were considered to be members of Inonotus sensu stricto in this study. Species of Inonotus linteus complex clustered in two clades, deviating from Inonotus sensu stricto, and are respectively treated as two new genera, Sanghuangporus and Tropicoporus.
Species of Inonotus sensu stricto
Among the above-mentioned 21 species of Inonotus sensu stricto, Inonotus glomeratus and I. tricolor clustered outside the Clade A in the phylogeny inferred from ITS dataset (Fig. 2). However, the two species have key characters of Inonotus sensu stricto, such as a monomitic hyphal system, presence of hyphoid and hymenial setae, and coloured basidiospores (Ryvarden 2005; Dai 2010). The discrepancy between ITS-based phylogeny and morphology might be caused by the low resolution of ITS region at the generic level. The nLSU-based phylogeny (Fig. 1) did confirm I. glomeratus as the members of Inonotus sensu stricto. Unfortunately, the nLSU region of I. tricolor was failed to obtain in this study. Further efforts to sequence the nLSU region from specimens of I. tricolor as well as of I. henanensis and I. latemarginatus might provide more insights into the delimitation of Inonotus sensu stricto, which is beyond the scope of this study.
All the 21 species of Inonotus sensu stricto produce annual basidiocarps with a monomitic hyphal system and coloured basidiospores, except that I. pachyphloeus and I. tricolor have a perennial habit and I. henanensis has hyaline basidiospores (Ryvarden 2005; Dai 2010; Dai et al. 2011). Inonotus andersonii, I. chihshanyenus, I. henanensis, I. micantissimus, I. nidus-pici, I. obliquus, I. pegleri, I. pruinosus and I. ulmicola produce resupinate basidiocarps, and I. glomeratus bears resupinate to effused-reflexed basidiocarps, while the other 11 species have pileate basidiocarps (Ryvarden 2005; Dai 2010; Dai et al. 2011). Inonotus hispidus, the generic type, and I. plorans has neither hyphoid nor hymenial setae, while the other 19 species has either one or both types of setae (Ryvarden 2005; Dai 2010; Dai et al. 2011).
Sanghuangporus and Tropicoporus differ from species of Inonotus sensu stricto mainly in having a dimitic hyphal system in trama. The two newly proposed genera are fairly similar in morphology, and it is hard to clearly separate them according to simple character differences (see the key for the features combined to separate them).
Sanghuangporus species
Nine species of Sanghuangporus are combined from Inonotus linteus complex. This group has recently been phylogenetically studied (Wu et al. 2012; Tian et al. 2013). Therefore, only one representative for each of the nine well-known species was sampled here for parsimony. Some of these species are medicinal fungi and have been subjected to pharmaceutical studies worldwide, but especially in East Asia (Dai et al. 2009, 2010; De Silva et al. 2012a, b, 2013). Previous studies in China, Japan and Korea have often dealt with all species as the important medicinal fungal species ‘Sanghuang’ (Jeon et al. 2011; Sun et al. 2014), and for several decades Phellinus linteus (or Inonotus linteus) was commonly used as the scientific binomial for species of this group. Recently, the species diversity of this group was clarified, and several new species and new combinations were added to the Inonotus linteus complex (Wu et al. 2012; Tian et al. 2013). Inonotus sanghuang (combined to Sanghuangporus above) was recognized as the genuine ‘Sanghuang’ species (Wu et al. 2012).
The nLSU-based phylogeny (Fig. 1) showed the two Ethiopian specimens, AM-08 and AM-19, clustering within Sanghuangporus. The African specimen O 915609 collected on Olea from Tanzania also nested within Sanghuangporus in ITS-based phylogeny and formed a sister relationship with the other nine species of Sanghuangporus (Fig. 2). O 915609 was morphologically identified as Phellinus microcystideus by Dai and Xu (1998) through comparison with the holotype from Congo. In this study, besides the reexamination of holotype of P. microcystideus, we also morphologically checked AM-08 and AM-19, and confirmed that the two specimens represent P. microcystideus. Therefore, we combine P. microcystideus to Sanghuangporus. It is noteworthy that P. microcystideus was treated as a synonym of P. linteus (Ryvarden 1983) and Harjula (1980) reported that this species is used as a herb to cure headaches in Tanzania.
Tropicoporus species
Nine species are accepted in Tropicoporus in this study, including two new species and seven new combinations. Most are tropical American species, with the exception of T. excentrodendri from Guangxi, tropical China and T. rudis from tropical Africa.
Tropicoporus excentrodendri was only found on aged trees of Excentrodendron tonkinense, which is an endangered species in the IUCN red list of threatened species (http://www.iucnredlist.org/); mature trees are very scarce. It means Tropicoporus excentrodendri can be also a threatened species, if it has a strict relationship with its endangered host tree.
Another Chinese specimen within the Tropicoporus clade is Yuan 5898 from Guangxi. This specimen morphologically resembles T. tropicalis by its annual and exclusively resupinate basidiocarps, which can be distinguished from all other species of Tropicoporus. Specimen Yuan 5898 was closely related to four representatives of T. tropicalis in the ITS-based phylogeny (Fig. 2), and formed a basal lineage. The locality of Yuan 5898 (China) has a long geographic distance from type locality of Tropicoporus tropicalis (Brazil), and Yuan 5898 also has a relatively long branch in ITS-based phylogeny (Fig. 2), therefore we treat Yuan 5898 as Tropicoporus cf. tropicalis.
Tian et al. (2013) described Inonotus cubensis (combined to Tropicoporus above) based on the holotype and two paratypes from Cuba. However, one of the paratypes (MUCL 43130) is actually from Costa Rica. MUCL 43130 and JV 1408–25 also from Costa Rica formed a separated clade from T. cubensis in nLSU-based phylogeny (Fig. 1), and clustered together with another two Costa Rican specimens (JV 1407-103 J and O 19228) as a distinct clade in the ITS-based phylogeny (Fig. 2). These two clades are well-supported and clearly isolated from Tropicoporus cubensis and other species of Tropicoporus. The four Costa Rican specimens represent Tropicoporus guanacastensis, which is introduced in this paper.
Xanthochrous rudis was described from Maijirgui, Niger, West Africa (Patouillard 1907), and Ryvarden (1983) treated it as a synonym of Phellinus linteus. Two successfully sequenced specimens from Rwanda (O 915614) and Tanzania (O 915617), both in East Africa, fit well with the morphology of X. rudis based on the current holotype study, and were considered to be X. rudis. Xanthochrous rudis formed a distinct clade from Tropicoporus linteus within Tropicoporus (Fig. 2). Therefore, X. rudis is transferred to Tropicoporus based on morphology and phylogeny.
References
Dai YC (2010) Hymenochaetaceae (Basidiomycota) in China. Fungal Divers 45:131–343. doi:10.1007/s13225-010-0066-9
Dai YC, Cui BK, Yuan HS, He SH, Wei YL, Qin WM, Zhou LW, Li HJ (2011) Wood-inhabiting fungi in southern China. 4. Polypores from Hainan Province. Ann Bot Fenn 48:219–231
Dai YC, Xu MQ (1998) Studies on the medicinal polypore, Phellinus baumii, and its kin, P. linteus. Mycotaxon 67:191–200
Dai YC, Yang ZL, Cui BK, Yu CJ, Zhou LW (2009) Species diversity and utilization of medicinal mushrooms and fungi in China (Review). Int J Med Mushrooms 11:287–302
Dai YC, Zhou LW, Cui BK, Chen YQ, Decock C (2010) Current advances in Phellinus sensu lato: medicinal species, functions, metabolites and mechanisms. Appl Microbiol Biotechnol 87:1587–1593. doi:10.1007/s00253-010-2711-3
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: more models, new heuristics and parallel computing. Nat Methods 9(8):772. doi:10.1038/nmeth.2109
De Silva DD, Rapior S, Fons F, Bahkali AH, Hyde KD (2012a) Medicinal mushrooms in supportive cancer therapies: an approach to anti-cancer effects and putative mechanisms of action. Fungal Divers 55:1–35. doi:10.1007/s13225-012-0151-3
De Silva DD, Rapior S, Hyde KD, Bahkali AH (2012b) Medicinal mushrooms in prevention and control of diabetes mellitus. Fungal Divers 56:1–29. doi:10.1007/s13225-012-0187-4
De Silva DD, Rapior S, Sudarman E, Stadler M, Xu JC, Alias SA, Hyde KD (2013) Bioactive metabolites from macrofungi: ethnopharmacology, biological activities and chemistry. Fungal Divers 62:1–40. doi:10.1007/s13225-013-0265-2
Donk MA (1960) The generic names proposed for Polyporaceae. Persoonia 1:173–302
Fiasson JL, Niemelä T (1984) The Hymenochaetales: a revision of the European poroid taxa. Karstenia 24:14–28
Guindon S, Gascuel O (2003) A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Syst Biol 52:696–704. doi:10.1080/10635150390235520
Harjula R (1980) Mirau and his practice. A study of ethnomedicinal repertoire of a Tanzanian herbalist. Tri-Med Books Ltd., London
Jeon H, Hwang SG, Jung YH, Yang HS, Sung NY, Lee JL, Park DK, Yoo YC (2011) Inhibitory effect of oral administration of sangwhang mushroom (Phellinus linteus) grown on germinated brown rice on experimental lung metastasis and tumor growth in mice. Food Sci Biotechnol 20:209–214. doi:10.1007/s10068-011-0028-3
Jeong WJ, Lim YW, Lee JS, Jung HS (2005) Phylogeny of Phellinus and related genera inferred from combined data of ITS and mitochondrial SSU rDNA sequences. J Microbiol Biotechnol 15:1028–1038
Karsten PA (1879) Symbolae ad mycologiam Fennicam 6. Meddelanden af Societas pro Fauna et Flora Fennica 5:15–46
Katoh K, Misawa K, Kuma K, Miyata T (2002) MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res 30:3059–3066
Katoh K, Toh H (2008) Recent developments in the MAFFT multiple sequence alignment program. Brief Bioinform 9:286–298. doi:10.1093/bib/bbn013
Larsen MJ, Cobb-Poulle LA (1990) Phellinus (Hymenochaetaceae). A survey of the world taxa. Synopsis Fungorum 3:1–206
Patouillard NT (1907) Champignons nouveaux du Tonkin. Bulletin de la Société Mycologique de France 23:69–79
Pattengale ND, Alipour M, Bininda-Emonds ORP, Moret BME, Stamatakis A (2010) How many bootstrap replicates are necessary? J Comput Biol 17:337–354. doi:10.1089/cmb.2009.0179
Petersen JH (1996) Farvekort. The Danish Mycological Society’s colour-chart. Foreningen til Svampekundskabens Fremme, Greve
Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19:1572–1574
Ryvarden L (1983) Type studies in the Polyporaceae 14. Species described by N. Patouillard, either alone or with other mycologists. Occas Pap Farlow Herbarium Cryptogam Bot 18:1–39
Ryvarden L (2002) Studies in neotropical polypores 17. New neotropical Inonotus species. Synopsis Fungorum 15:70–80
Ryvarden L (2005) The genus Inonotus, a synopsis. Synopsis Fungorum 21:1–149
Silvestro D, Michalak I (2012) raxmlGUI: a graphical front-end for RAxML. Org Divers Evol 12:335–337. doi:10.1007/s13127-011-0056-0
Stamatakis A (2006) RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22:2688–2690. doi:10.1093/bioinformatics/btl446
Sun J, Chen QJ, Zhu MJ, Wang HX, Zhang GQ (2014) An extracellular laccase with antiproliferative activity from the sanghuang mushroom Inonotus baumii. J Mol Catal B Enzym 99:20–25. doi:10.1016/j.molcatb.2013.10.004
Tian XM, Yu HY, Zhou LW, Decock C, Vlasák J, Dai YC (2013) Phylogeny and taxonomy of the Inonotus linteus complex. Fungal Divers 58:159–169. doi:10.1007/s13225-012-0202-9
Vilgalys R, Hester M (1990) Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J Bacteriol 172:4238–4246
Vlasák J, Li HJ, Zhou LW, Dai YC (2013) A further study on Inonotus linteus complex (Hymenochaetales, Basidiomycota) in tropical America. Phytotaxa 124:25–36. doi:10.11646/phytotaxa.124.1.3
Wagner T, Fischer M (2002) Proceedings towards a natural classification of the worldwide taxa Phellinus s.l. and Inonotus s.l., and phylogenetic relationships of allied genera. Mycologia 94:998–1016
White TJ, Bruns T, Lee S, Taylor JW (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: a guide to methods and applications. Academic, New York, pp 315–322
Wu SH, Dai YC, Hattori T, Yu TW, Wang DM, Parmasto E, Chang HY, Shih SY (2012) Species clarification for the medicinally valuable ‘sanghuang’ mushroom. Bot Stud 53:135–149
Zhou LW, Qin WM (2012) Inonotus tenuicontextus sp. nov. (Hymenochaetaceae) from Guizhou, Southwest China with a preliminary discussion on the phylogeny of its kin. Mycol Prog 11:791–798. doi:10.1007/s11557-011-0792-8
Acknowledgments
Special thanks are due to Karl-Henrik Larsson (O, Norway) and Tsutomu Hattori (FFPRI, Japan) for loan of specimens and culture. This research was financed by the National Natural Science Foundation of China (Project No. 31200015).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Zhou, LW., Vlasák, J., Decock, C. et al. Global diversity and taxonomy of the Inonotus linteus complex (Hymenochaetales, Basidiomycota): Sanghuangporus gen. nov., Tropicoporus excentrodendri and T. guanacastensis gen. et spp. nov., and 17 new combinations. Fungal Diversity 77, 335–347 (2016). https://doi.org/10.1007/s13225-015-0335-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13225-015-0335-8