
Abstract
Necrotrophic pathogens experience host-generated oxidative stress during pathogenesis. They overcome such hostile environment by intricate mechanisms which are largely understudied. In this article, reference-based transcriptome analysis of a devastating Ascochyta Blight (AB) disease causing chickpea pathogen Ascochyta rabiei was explored to get insights into survival mechanisms under oxidative stress. Here, expression profiling of mock-treated and menadione-treated fungus was carried out by RNA-Seq approach. A significant number of genes in response to oxidative stress were overrepresented, suggestive of a robust and coordinated defense system of A. rabiei. A total 73 differentially expressed genes were filtered out from both the transcriptomes, among them 64 were up-regulated and 9 were found down-regulated. The gene ontology and KEGG mapping were conducted to comprehend the possible regulatory roles of differentially expressed genes in metabolic networks and biosynthetic pathways. Transcript profiling, KEGG pathway and gene ontology-based enrichment analysis revealed 12 (16.43%) stress responsive factors, 25 (34.24%) virulence associated genes, 10 (13.69%) putative effectors and 28 (38.35%) important interacting proteins associated with various metabolic pathways. In addition, genes with differential expression were further explored for underlying putative pathogenicity factors. We identified five genes ST47_g10291, ST47_g9396, ST47_g10294, ST47_g4395, and ST47_g7191 that were common to stress and fungal pathogenicity. The factors recognized in this work can be used to establish molecular tools to explain the regulatory gene networks engaged in stress response of fungal pathogens and disease management.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fungal diseases in crops pose a significant risk to food safety and the health of the ecosystem (Dean et al. 2012). Upon pathogen attack, high production of nitric oxide (NO) and reactive oxygen species (ROS) commonly referred as oxidative burst, is an earliest response from host which results in localized host cell death. These ROS produced by the consumption of molecular oxygen in different cellular compartments restrict the growth of pathogen during host–pathogen interactions (Asai and Yoshioka 2009; Pietrowska et al. 2015). The initial defence pathway through mitogen-activated protein kinase (MAPK) is known to be involved in signaling during oxidative and nitrosative stress (Asai et al. 2008; Asai and Yoshioka 2009; Sarkar et al. 2014; Lehmann et al. 2015). At higher concentrations, ROS cause damage to biomolecules such as nucleic acid, protein and lipid; whereas at low or moderate concentrations, they act as secondary messenger in intracellular signaling cascades. NO acts as a signaling molecule and regulates many important signaling pathways, including defense responses (Besson-Bard et al. 2008). It appeared in conjunction with ROS as a significant participant in hypersensitive response (HR) and cell death (Yoshioka et al. 2009).
Oxidative stress response is often investigated to understand acclimatization mechanisms of pathogen under fluctuating environments and to elucidate stress signaling and regulatory molecular processes. The biotrophic pathogens derive their nutrition from living host tissue while the necrotrophs kill the host and thrive on dead tissue. The biotrophic pathogens generally suppress host-generated oxidative stress to alleviate host-immune responses while the necrotrophs survive under these extreme conditions and utilize host-generated oxidative stress towards their own benefit (Glazebrook 2005). Hence, despite the presence of robust host defense system the specialized necrotrophs can successfully colonize host (Govrin and Levine 2000) which indicates the involvement of some evolved mechanisms against oxidative stress during infection process. For example, Botrytis cinerea, a necrotrophic fungal pathogen produces ROS scavenging enzymes (catalase and superoxide dismutase) to prevent oxidative damage (Gilad et al. 2000; Mayer et al. 2001). The Magnaporthe oryzae catalase gene (CATB) knockout mutant demonstrates impaired pathogen fitness and decreased pathogenicity (Skamnioti et al. 2007). Moreover, it has been also shown that transcription factor MoAtf1 is essential for oxidative stress response and virulence of M. oryzae (Guo et al. 2010). The rice blast fungus M. oryzae counteracts nitrooxidative stress with the help of Nitronate monooxygenase (NMO) enzymes which maintain the redox balance and promote fungal virulence (Marroquin-Guzman et al. 2017). A gene from B. cinerea encoding flavohemoglobin (BcFHG1) has been reported as a crucial factor in NO neutralization and provides protection from nitrosative conditions (Turrion-Gomez et al. 2010). Taken together, these reports suggest that effective and rapid elimination of ROS and other toxic compounds produced by the plants is essential for the survival of fungal pathogens.
The phytopathogen Ascochyta rabiei (Pass.) Lab. (Teleomorph: Didymella rabiei), a necrotrophic fungus which causes Ascochyta Blight (AB) in chickpea (Cicer arietinum L.), resulting in severe losses in crop productivity (Nizam et al. 2010). The primary characteristics of A. rabiei infection are concentric necrotic lesions on all the above ground parts of plant. In tolerant cultivars, during Chickpea-Ascochyta interaction, the occurrence of a fast oxidative burst followed by HR has been observed (Höhl et al. 1990; Otte and Barz 1996). The availability of A. rabiei genomic resources at NCBI under the bioprojects PRJNA272665 (Verma et al. 2016) and PRJNA510692, and advanced RNA-Seq technologies enabled us to perform reference-based transcriptome analysis of this fungus under control and oxidative stress conditions. We aimed to decipher molecular responses and elucidate genes deployed in oxidative stress responses of A. rabiei. This study provides putative oxidative stress responsive genes which may have contribution in oxidative stress tolerance and fungal pathogenicity.
Materials and methods
Fungal culture and treatment
The pure cultures of A. rabiei isolate ArDII (Indian Type Culture Collection No. 4638) were maintained in our laboratory. Fungal cultures were inoculated with spore suspensions (1 × 103 spores/ml) and grown on PDB (Potato Dextrose Broth; Difco Laboratories, USA), shaking at 120 rpm at 22 °C in the dark for 4 days. The 4 days grown fungal hyphae were used for exogenous menadione treatment (250 μM, Sigma-Aldrich, USA). Fungal cultures were harvested at 3 h post menadione treatment. Mock-treated samples were used as controls.
RNA extraction and quality control
Three independent biological replicates of each mock-treated and menadione-treated sample were collected and frozen in liquid nitrogen. The total RNA was extracted from tissues using TRI Reagent (Sigma Life Science, USA) following manufacturer’s protocol. The quantity and quality of RNA samples were checked using NanoVue (GE Healthcare, Hong Kong). Agilent 2100 Bioanalyzer and RNA 6000 nanokit (Agilent Technologies, Singapore) were also used to check the quality of all RNA samples. The RNA samples with 260/230 ratio from 2.0 to 2.5, 260/280 ratio from 1.9 to 2.1 and RIN (RNA integrity number) from 8.7 to 9.0, were further considered for the analysis.
Library preparation and high-throughput sequencing
Equal quantities of total RNA samples from three biological replicates were pooled and mRNA was purified using Dynabead mRNA purification kit (Invitrogen) following the manufacturer’s protocol. The two cDNA libraries were prepared using cDNA rapid library preparation kit (Roche Diagnostics) according to manufacturer’s instructions for the GS FLX Titanium series. The SuperScript® double-stranded cDNA synthesis kit (Invitrogen) was used to synthesize double-stranded cDNA. About 600 ng of double-stranded cDNA were nebulized and selected for the fragment length range of 300–800 bp. The unique adapters were ligated to the fragmented cDNA and denatured to produce single-stranded cDNA followed sequencing by emulsion PCR. The quality of both libraries was evaluated using High Sensitivity DNA Kit on Agilent 2100 Bioanalyzer. The cDNA libraries were sequenced with GS FLX Titanium series sequencing reagents and sequencer in two runs (one complete and two half runs). Statistics of high quality data of mock-treated and menadione-treated samples are provided in Table 1.
Pre-processing and reference transcript assembly
Sequences output from the Roche 454 FLX Titanium sequencer in Standard Flowgram Format (sff) were converted into fastq format using open source GitHub tool sff2fastq (https://github.com/indraniel/sff2fastq). The quality of Roche 454 sequence reads was checked and further filtered to minimum quality (Phred score 20) and minimum length (200 bp). Adapter sequences along with low-quality nucleotides bases were trimmed using PRINSEQ tool (https://prinseq.sourceforge.net/) (Schmieder and Edwards 2011). Thus, filtered high-quality reads (20×) were used for further analysis. Subsequently, mapping of trimmed reads were done to the Ascochyta reference genome using the HISAT2 software v2.1.0 (Kim et al. 2015). The high-quality Roche 454 FLX Titanium reads were used for reference guided transcript assembly for the two samples using StringTie v1.3.5 (Pertea et al. 2015). Various other parameters were taken into account as a function, such as total number of contigs (about 200 bp), average contig length and N50 length to optimize best possible transcript length for transcriptome assembly. Contigs clustering using CD-HIT (Li and Godzik 2006) significantly reduced the number of sequences in each set. On a server with 48 cores and 128 GB random access memory, all assemblies were conducted. Overall schematic workflow is graphically represented in Fig. 1.

Schematic work flow diagram for mock and menadione-treated A. rabiei transcriptome assembly and annotation analysis
Read mapping to reference genome
The analysis starts by mapping high-quality reads against the reference genome of Ascochyta to define the positions from which the reads originated. This mapping data enables us to gather subsets of reads for each gene and then assemble and quantify transcripts represented by those reads. The reference genome of A. rabiei (Strain ArDII) (Verma et al. 2016) and the corresponding GTF file was downloaded from NCBI database (https://www.ncbi.nlm.nih.gov/assembly/GCA_001630375.1). The draft genome consists of ~ 34.6 Mb in bp length. The reference GTF file contains information (locus and description) regarding genes, exons and transcripts. There are a total of 10,596 genes, 29,292 exons and 10,596 transcripts as per the GTF file. The high-quality reads were then mapped to A. rabiei genome using HISAT2 software v2.1.0 (Kim et al. 2015) to create alignment in BAM format for each sample. First step is indexing the reference genome using hisat-build (HISAT2 specific indexer program). Then the input reads, in FASTQ format, were given to HISAT2 aligner along with the reference genome index and create splice-site aware alignment.
Differential gene expression analysis
The expression levels of each unigene were measured with FPKM (fragment per kilobase of exon per million fragments mapped) method. Briefly, reads were first aligned against each transcript of A. rabiei using HISAT2 v2.1.0. The abundance of the transcripts was estimated between mock and menadione treated datasets using StringTie (Pertea et al. 2015) and the read count information was used to find out the DEGs between the two groups using the Bioconductor (https://www.bioconductor.org/packages/release/bioc/html/) package DESeq2 v1.24.4 in R environment (https://www.r-project.org/). The package DESeq2 provides methods to test for differential expression in gene expression data by the negative binomial distribution-based model. The resulting false discovery rate was calculated by Benjamini and Hochberg’s algorithm. The log2FC ≥ 2 and log2FC < 2 were considered as criteria to identify up-regulated and down-regulated genes, respectively. Further, transcripts with combination of at least two-fold change difference and p value ≤ 0.05 were considered significantly modulated. The criterion used to identify upregulated and downregulated transcripts along with the significance is provided in Supplementary Table S1.
Quantitative real-time PCR (qRT-PCR)
For qRT-PCR analysis, total RNA was extracted from three biological replicates as described above and treated with DNase I (Promega, USA) according to manufacturer’s protocol followed by cDNA synthesis. The primers were designed using Primer ExpressH (version 3.0) software (Applied Biosystems) with the default parameters. The primers used for quantitative RT-PCR are shown in Supplementary Table S2. Real-Time PCR was carried on a 7900HT Sequence Detection System using Sequence Detection Systems Software version 2.3 (Applied Biosystems) and Power SYBR Green PCR Master Mix (Applied Biosystems). The default cycling program was used with the following cycling conditions: 2 min at 50 °C, 10 min at 95 °C, and 40 cycles of 15 s at 95 °C and 1 min at 60 °C. Each experiment was performed with three technical replicates. The gene encoding EF1α was used as endogenous control for normalization in all experiments. Expression ratios were calculated from cycle threshold values using the 2−ΔΔCT method.
Similarity Search and functional annotation of DEGs
With a significant e value cut-off of ≤ 1E-06, BLAST algorithm was used to annotate differentially expressed (DE) transcripts (obtained from differential expression analysis) against various databases [non-redundant (nr) protein database of NCBI, UniProt/Swissprot, GO, PFAM and KEGG, KOG] to assign putative function to them on basis of similarity search. Only the best hits were extracted and hits with a 60% sequence similarity were considered as significant. Gene Ontology (GO) (https://www.geneontology.org/)-based annotations for biological process, molecular function and cellular component categories to assign one or more GO terms and their putative functional roles were based on the BLAST hit which was performed from Blast2GO v2.4.2 (https://www.blast2go.com/) (Götz et al. 2008) suite and GOseq R packages (Young et al. 2010). The associated hits were searched for their respective GO terms. Moreover, we used the functionality of Pfam annotations to retrieve the conserved protein families/domains by searching against the Pfam v32.0 protein family database with 17,929 protein entries.
GO and KO annotation-based functional enrichment and pathway analysis
For functional enrichment and biological interpretation of the DEGs, the significantly over-represented GO terms and biological pathways were explored with the pathway analysis tool. The significantly enriched pathways for DEGs in this study were determined by the GO enrichment analysis by the GOseq R packages (Young et al. 2010) and KEGG Orthology-Based Annotation System (KOBAS) (https://kobas.cbi.pku.edu.cn/) (Kanehisa et al. 2008; Mao et al. 2005). The amino acid sequences of enzymes implicated in oxidative stress responses were downloaded from the KEGG for the selected fungal species with sequenced genomes representing both ascomycetes and basidiomycetes.
Prediction of secretory and effector proteins
The translated DEGs of A. rabiei transcriptome were searched for putative secreted proteins using various tools to screen DEGs for secretory nature. We performed predictions using servers such as SignalP4.1 (https://www.cbs.dtu.dk/services/SignalP/) (Petersen et al. 2011) for classical pathway on the basis of N-terminal signal peptide whereas SecretomeP2.0 (https://www.cbs.dtu.dk/services/SecretomeP/) for proteins secreted through non-classical pathway. TargetP1.1 (https://www.cbs.dtu.dk/services/TargetP/) was used for localization analysis. Moreover, EffectorP v1.0 and v2.0 (https://effectorp.csiro.au/) tools were used to predict effector proteins among the DEGs.
Identification of stress and pathogenicity related DEGs
Protein sequences for 1985 reference stress response proteins in fungal species were obtained from Fungal Stress Response Database (FSRD) (https://internal.med.unideb.hu/fsrd2/?p=consortium) which includes 211 GO terms in 6 main stress response categories and functionally characterized stress response proteomes of 15 fungal spp. (Karányi et al. 2013). BLASTP was used to identify and classify putative stress responsive genes in A. rabiei using parameters e value 1e−5. The results were filtered with a threshold cut-off of 40% identity and 80% coverage, and then homologous sequences were extracted and classified. To identify the putative pathogenicity related proteins, all DEGs were searched against Pathogen–Host Interactions (PHI) database (https://www.phi-base.org/) (Urban et al. 2017) which contains 6438 gene accessions based on the experimental evidences from 263 species of microbial pathogens.
PPI network construction-based analysis
Using NetworkAnalyst tool (https://www.networkanalyst.ca) (Xia et al. 2015), the protein–protein interaction (PPI) information were mapped to reveal the interactive links among the DEGs that were likely to play important roles in oxidative stress reaction. The enrichment network map was constructed using a standalone Cytoscape version v3.6.1 (Shannon et al. 2003) for the genes associated with enriched KO and GO-terms. Cytoscape plugin BioGRID (Oughtred et al. 2019) interaction database was used for network construction. Using NetworkAnalyzer in Cytoscape (Shannon et al. 2003), we calculated the topological properties (such as betweenness centrality and degree distribution) of the PPI network. The degree distribution of all network nodes can assist to clarify whether or not a network is scale-free. In protein interaction networks, the densely linked group of proteins could be referred to as hub genes. The enriched pathways of DEGs in significant modules (≥ 5 DEGs) were analyzed with a threshold of p ≤ 0.05. In the PPI network, nodes and edges (lines) represent proteins and their interactions, respectively.
Results
Reference-based transcriptome assembly captures high-quality transcripts
The Roche 454 sequencing run resulted in 565,567 reads for Control (Mock-treated) and 719,459 reads for oxidative stress (Menadione-treated) samples. After quality filtering, adapter trimming and mapping on the A. rabiei reference genome, 81.20% and 83.80% of the filtered reads remained for control and oxidative stress samples, respectively. Their assemblies produced 22,548 contigs (average read depth = 5.6) for control, and 20,369 contigs (average read depth = 5.9) for oxidative stress sample. The statistics of transcript assemblies is provided in Table 2.
Transcripts differentially expressed during oxidative stress
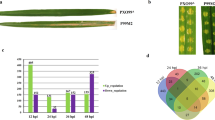
To assess oxidative stress regulated changes in the fungal gene expression both assemblies were taken into account. The differential expression analysis of genes was evaluated through the FPKM values. In this study, all the genes having non-zero FPKM values were taken into consideration for which the box plot has been shown in Fig. 2a. Overall we got 6477 genes (with non-zero FPKM value), out of them a total of 778 genes were down-regulated and 5699 genes were up-regulated. After applying required filters of p value = 1e−05 and absolute log-fold = 2, we could identify 73 genes which showed significant differential expression between the two conditions (Fig. 2b). Among the 73 differentially expressed transcripts analyzed, 9 (12.32%) were significantly down-regulated and 64 (87.67%) were up-regulated (Fig. 2c). For the detection of genes differentially expressed in the two conditions the 73 differentially expressed genes has been plotted as heatmap (Fig. 2d). The resulting list of DEGs and the column description of DESeq2 output is provided in Supplementary Table S3.

FPKM box plot and heatmap of differentially expressed genes (DEGs) of mock-treated (control) and Menadione-treated (oxidative stress) samples of A. rabiei. a Log2(FPKM) values of mock-treated and menadione-treated samples. Tentacles express the range of maximum and minimum value of expression quantity, the boxes express 25–75% of log2(FPKM), and the black lines in the box express the median, b MA plot showing gene expression changes from two different conditions (mock vs. menadione treated) in terms of log fold change on Y-axis and mean of expression counts on X-axis. Points in red represent genes that showed a significant expression difference. c Significantly up and down-regulated DEGs identified under oxidative stress. d Heatmap showing differential gene expression; representing 73 DEGs under control vs. oxidative stress conditions based on gene abundance in samples as a function of expression levels in different FPKM intervals. The X-axis represents samples and the Y-axis represents the number of differential genes
Similarity search and functional annotation of DEGs
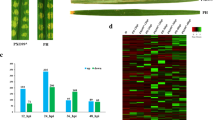
The assembled transcripts of A. rabiei were functionally annotated by similarity search with proteins using various tools. A total of 69 genes (94.52% out of 73 DEGs) were annotated using blastp annotation against the non-redundant (Nr) NCBI database with e value cut-off ≤ 1e−05 and more than 40% similarity whereas 4 of them showed no homology to known sequences. Among these 69 genes, 60 were found hypothetical proteins and only 9 (17.80%) could be assigned with a function. To classify the DEGs for functions, we used Gene Ontology (GO) search and 24 DEGs (out of 73) with at least one GO term for each unigene, (1 GO term for 4 genes and multiple GO terms for 20 genes) were assigned with putative function whereas functions could not be assigned to rest of the 67.12% transcripts. These resulting GO terms were identified under three major categories 21.91% in ‘cellullar component’, 27.39% in ‘molecular function’ and 31.5% unigenes in ‘biological process’ categories. Using Swiss-Prot, we annotated 29 genes (39.72%). The Pfam and Kyoto Encyclopedia of Genes and Genomes Ontology (KO) annotations assigned function to 59 (80.82%) and 27 genes (36.98%), respectively, with more than 40% similarity (Supplementary Table S4). Among all, 11 DEGs could be mapped in common by Nr, Uniprot/Swiss-Prot, GO, Pfam and KOG annotations. The Venn diagram represents the clusters of mapped DEGs of A. rabiei transcriptome against five databases in each intersection (Fig. 3a). After annotation, consistent with previous report (Singh et al. 2012), we found the upregulation of oxidative stress marker genes like Glutathione reductase, peroxidase etc. after menadione-treatment as compared to mock-treated samples (Fig. 3b). To validate the transcriptomic data, we selected 6 genes (1 downregulated and 5 upregulated) out of 73 DEGs and their expression profiles were evaluated using quantitative real-time PCR. These genes encode for Copper ion transmembrane transporter (ST47_g5350), Thioredoxin disulphide reductase (ST47_g10294), Glutathione reductase (ST47_g3798), FMN-binding oxidoreductase (ST47_g426), Zn-binding xidoreductase (ST47_g1601) and Zn-binding oxidoreductase (ST47_g4979) proteins. The samples were collected at 3 h time point after menadione treatment and used for qRT-PCR. The relative-expression results obtained with real-time PCR were consistent with those obtained from the RNA-seq as shown in Fig. 3c.

Intersection of differentially expressed genes identified by multiple tools in downstream analyses and validation. a Venn diagram of annotation against the NCBI non-redundant (Nr), Uniprot/Swiss-Prot, Protein family (Pfam), euKaryotic Ortholog Groups (KOG) databases and Gene Ontology (GO), b Marker genes up-regulated in A. rabiei transcriptome under oxidative stress, c Validation of RNA-seq data using relative qRT-PCR of six genes. The EF1alpha gene of A. rabiei was used as an internal control for normalization. The relative mRNA level was calculated with respect to the control samples. The error bars represent the standard error of means from three biological replicates
Gene ontology and KEGG ontology-based functional enrichment pathway mapping of differentially expressed genes
The functional enrichment analyses of DEGs is based on the GO and KEGG databases. According to the GO functional category enrichment analysis, 22 terms appeared for the upregulated and 2 terms appeared for the downregulated DEGs of oxidative stress vs. mock samples. When the biological processes and molecular function GO categories were taken together, 17 terms were found for upregulated DEGs and one term for downregulated DEG. Among nine downregulated genes, GO terms could be assigned only to the two genes (ST47_g4395 and ST47_g9634) that were assigned GO terms ‘ATP binding’ and ‘inorganic phosphate transmembrane transporter activity’, respectively. In the biological process category, the largest groups were “cellular metabolic process”, “primary metabolic process” and “biosynthetic process” whereas in the molecular function, DEGs with “catalytic activity”, “oxidoreductase activity” and “ion binding” formed the largest groups. This indicates that genes related to “cellular response to stimulus”, “response to external stimulus”, “response to oxidative stress”, “removal of superoxide radicals”, “reactive oxygen species metabolic process” may play important roles in during oxidative stress conditions. Furthermore, the terms “oxidoreductase activity” and “antioxidant activity” were also enriched for up-regulated DEGs in oxidative stress vs. mock-treated combination, suggesting that the large number of ROS scavenging metabolic processes are affected under oxidative stress. The differentially expressed genes representing different GO categories viz. biological process, molecular function and cellular component depicting a broad spectrum of the analyzed transcriptome are shown in Fig. 4. The list of GO domain distribution is provided in Supplementary Table S5.

Gene ontology (GO) functional enrichment analysis of differentially expressed genes
According to the KEGG pathway analysis, we found 19 significantly enriched pathways using the associated KOG annotation terms between mock-treated vs. oxidative stress-treated samples. Moreover, with these associated pathway IDs, 12 enzymes have been found differentially expressed and their effect on corresponding metabolism were also explored. The major DEGs-enriched pathways included, Biosynthesis of ansamycins, Biosynthesis of antibiotics, Carbon fixation in photosynthetic organisms, Pentose phosphate pathway (ST47_g2253), Glutathione metabolism, Metabolism of xenobiotics by cytochrome P450 (ST47_g4543 and ST47_g3798), Sulfur metabolism, Cysteine and methionine metabolism (ST47_g1980), Arachidonic acid metabolism (ST47_g5327), Pyruvate metabolism, Lysine biosynthesis (ST47_g3023), Amino sugar and nucleotide sugar metabolism (ST47_g8615), Selenocompound metabolism (ST47_g10294 and ST47_g10291), Glycine, serine and threonine metabolism (ST47_g10291), Phenylalanine, tyrosine and tryptophan biosynthesis (ST47_g10129), Pyrimidine metabolism (ST47_g6594) and Purine metabolism (ST47_g4305). The combination of the expression patterns of the DEGs and their enrichment outcomes showed that the up-regulation of DEGs and metabolic pathways was predominant, indicating that these components of metabolism play a significant role in response to stress. All extensive pathway maps have been provided in Supplementary data (Table S6, Figure S1 and S2).
Secretory and effector proteins
Fungal pathogens secrete proteins to modulate the immune response/metabolic processes of their host plants. Fungal proteins can also be secreted to suppress the host oxidative stress response and promote pathogen infection. Since these proteins are of secretory nature, therefore, the predicted amino acid sequences of DEGs were screened for the putative secretory proteins using various tools. We performed predictions using servers such as SignalP for proteins secreted through classical pathway, SecretomeP for non-classical pathways and TargetP for localization analysis. Among the DEGs, TargetP predicted 6 secretory proteins whereas SignalP which looks for signal peptide cleavage site at the N-terminal region, could identify only four of them to be secretory through classical secretory pathway. With SecretomeP analysis, a total of eight secretory proteins were predicted which could be secreted through non-classical pathway. Taken together these tools identified six putative secretory proteins (ST47_g9844, ST47_g874, ST47_g9633, ST47_g3245, ST47_g238 and ST47_g1893) which were predicted by at least two tools. Among them, four genes (ST47_g9844, ST47_g874, ST47_g9633 and ST47_g3245) were up-regulated and two genes (ST47_g238 and ST47_g1893) were found down-regulated under menadione induced oxidative stress environment.
Additionally, we utilized EffectorP tool to predict effector proteins among the DEGs. Only 6 (8.2%) and 9 (12.3%) proteins among the 73 DEGs were significantly predicted as effector proteins by EffectorP1.0 and EffectorP2.0, respectively. Out of them one was unique to EffectorP1.0 and 4 were found unique to EffectorP2.0. Hence, we identified at least ten putative effector candidate genes (ST47_g7918, ST47_g10073, ST47_g8510, ST47_g5327, ST47_g4543, ST47_g543, ST47_g8057, ST47_g7659, ST47_g5350, and ST47_g5356) differentially expressed during oxidative stress. Among these, eight genes (ST47_g10073, ST47_g8510, ST47_g5327, ST47_g4543, ST47_g543, ST47_g8057, ST47_g7659 and ST47_g5356) were up-regulated with two genes (ST47_g7918 and ST47_g5350) down-regulated. The eight up-regulated genes are probable to play a vital role in modulating the oxidative stress responses during host colonization and could be used for further research.
Oxidative stress–response and pathogenicity-related proteins
To identify potential stress response proteins, we searched all 73 DEGs against fungal stress database and found hits only for 12 (16.43%) genes with a cut-off e-value of 1e-5 and more than 40% similarity (Supplementary Table S7). This database contains 1985 proteins which are known to respond against various types of stresses such as oxidative stress, heat stress and osmotic stress. We also focused on known pathogenicity-related genes in A. rabiei as previous reports suggest that stress response and fungal pathogenicity are correlated (Lambou et al. 2010; Pradhan et al. 2017; Marroquin-Guzman et al. 2017). Therefore, we used Pathogen–Host Interactions (PHI) database to identify the putative pathogenicity related proteins. All 73 DEGs were searched against PHI database which contains 6438 accessions based on the experimental data from various pathogenic microbes. We found a total 26 DEGs (35.61% of 73) significantly associated with fungal pathogenicity considering single hit for each DEG and more than 40% similarity cut-off (Supplementary Table S4). The stress responsive and virulence associated DEGs are shown in Fig. 5. Thus, we have identified six genes (ST47_g10291, ST47_g9396, ST47_g10294, ST47_g4395, ST47_g7191 and ST47_g8615) which were common between FSRD and PHI databases. Out of these six genes, only ST47_g8615 exhibits unaffected virulence phenotype where as others show reduced virulence phenotype. The rest of genes which were not found in PHI database can be the possible pathogenicity factors for A. rabiei. These candidates can be further characterized in future for their role in oxidative stress tolerance and fungal pathogenicity.

Showing number of DEGs involved in stress response and pathogenicity along with putative pathogenicity determinants
Gene-set enrichment analysis and protein–protein interaction (PPI) network of the DEGs
We also examined the prospective associations of the 73 DEGs with the help of protein–protein interaction network analysis. To define functional interaction (FI) networks, we mapped these genes to the Saccharomyces cerevisiae interactome data set because there is no functional interaction network for A. rabiei oxidative stress proteins. Prior to mapping, the IDs of the 73 DEGs were converted to gene symbols for the respective encoded proteins; however, only 28 genes were assigned symbols (Supplementary Table S8). We used the algorithm of edge betweenness to define the network's functional modules. The number of genes included by shared GO terms was used to predict important pathways and subsequent crosstalk between these pathways. We found that some DEGs had few interactions with other proteins. The obtained PPI network based on the 73 DEGs between two groups is represented in Fig. 6. The interactions included direct (physical) and indirect (functional) associations. The top hub proteins were ST47_g3798 (ArGLR1), ST47_g10129 (ArTRP5), ST47_g10294 (ArTRR1), ST47_g10291 (ArCYS3), ST47_g426 (ArOYE2), ST47_g1359 (ArOYE3) and ST47_g4979 (ArYML131W). Interestingly, most of these key proteins are related to oxidative stress response whereas ArOYE3 and ArYML131W exhibit oxidoreductase activity. These oxidoreductases might be involved in maintaining cellular redox homeostasis.

Protein–protein interaction network of the proteins found differentially expressed in mock-treated vs. oxidative stress-treated sample, based on interaction evidence. The highlighted circle area denotes Red: down-regulated, Green: up-regulated, Yellowish-green: highly up-regulated DEGs. Common DEGs among FSRD and PHI database are shown as purple circles where green outline represents up-regulated and red outline shows down-regulated DEGs
Discussion
Increased concentrations of ROS in cells result in protein, lipid and DNA damage. Thus, cells have endogenous antioxidant mechanisms to neutralize ROS, and these systems are critical to maintain appropriate functioning of the cells. Reduced glutathione (GSH) is one of the key cellular antioxidants, most effectively regenerated by glutathione reductase and reduced nicotinamide adenine dinucleotide phosphate. In fungal biology, stress-related developmental and physiological processes are important but they are yet to be explored. The amount of articles that aim to decipher the notable fungal stress tolerance is steadily growing.
Our study utilized the Roche 454 reads for RNA-seq analysis whose depth coverage is sufficient to identify the average and highly expressed genes while the FPKM analysis normalized the minor variation in mapping depth of control and menadione sample reads to the A. rabiei transcriptome. Although matured Illumina sequencing technology has decreased RNA-seq costs with increased read-depth, as compared to Roche 454, for identifying the genes expressed at low-levels. RNA-seq analysis in the present study, identified several differentially expressed genes (DEGs) associated with oxidative stress response characteristics. These DEGs were found to be engaged in biological functions such as oxidation–reduction, maintenance of redox cellular homeostasis, DNA repair and detoxification of ROS. RNA-seq analysis indicated higher up-regulation of four genes (ST47_g9844, ST47_g3798, ST47_g785 and ST47_g6429). We found that the genes encoding oxidoreductase, lyase and ATPase are upregulated in response to oxidative environment which indicates their role in stress adaptation. Previous studies on PKS1, CBL1 and BcatrB from Cochliobolus sativus, F. oxysporium and B. cinerea, respectively suggest that these enzymes are required for fungal virulence and pathogenesis (Yang et al. 1996; Schoonbeek et al. 2001; Seong et al. 2005). Among the DEGs, we identified putative effector proteins which were upregulated under oxidative stress in A. rabiei. Such proteins modulate the host immune machinery and are crucial for successful host colonization. Moreover, Pep1 effector from Ustilago maydis has been shown to suppress plant immunity by the inhibition of host peroxidases (Hemetsberger et al. 2012). Recently, the AvrBsT effector of Xanthomonas campestris pv. vesicatoria has been shown to cause H2O2 burst and HR cell death in Capsicum annuum (Kim et al. 2013).
Out of 7 hub proteins identified, ST47_g3798 (ArGlr1), a major important interacting node protein which interacts with multiple proteins, is well-known to play critical role during adaptation to oxidative and nitrosative stress and thereby helps in colonisation of the host (Tillmann et al. 2015). Our findings emphasize the significance of maintaining redox homeostasis and the glutathione redox system seems to play a major role in this fungal pathogen. All these processes require energy and a transketolase encoding gene, ST47_g2253, was found to be upregulated during the oxidative stress survival mechanism of A. rabiei. In KEGG pathway analysis, it was classified under the term ‘Carbon fixation in photosynthetic organisms’ which seems interesting as this enzyme function in the pentose phosphate pathway in all organisms and the Calvin cycle of photosynthesis. Although, the carbon fixation pathway of plants/photosynthesis (CFPP)-associated enzymes have been found in other heterotrophic phytopathogenic fungus such as Sclerotinia sclerotiorum genomes (Lyu et al. 2015). In conclusion, here we report the comparative transcriptomic landscape of gene expression in phytopathogenic fungus A. rabiei under oxidative environment. Our data suggests that besides activating antioxidant metabolism and stress response genes, necrotrophic pathogen can significantly modulate the various metabolic and biosynthetic processes under oxidative stress. The genes which are highly upregulated under oxidative stress conditions are plausible to play critical role in fungal survival during host-induced oxidative stress and cell death. Secretary proteins identified through current study could be the possible effectors which are deployed to manipulate immune system of host. This study delivers putative candidate genes (Fig. 7) to further study their possible role in oxidative stress tolerance and fungal virulence. Taken together, the current investigation opens up additional possibilities for systematic functional analysis of new ROS scavenging machinery and understanding of oxidative stress regulators of various fungal phytopathogens.

Key regulators of oxidative stress response
Data availability and materials
All data analysed during this study are included in this published article and its additional files. The raw RNA-sequencing data are deposited in the NCBI GenBank (www.ncbi.nlm.nih.gov) under study accessions SRX5765333 and SRX5765332, which can be retrieved under the BioProject accession number PRJNA539845.
References
Asai S, Yoshioka H (2009) Nitric oxide as a partner of reactive oxygen species participates in disease resistance to necrotrophic pathogen Botrytis cinerea in Nicotiana benthamiana. Mol Plant Microbe Interact 22(6):619–629
Asai S, Ohta K, Yoshioka H (2008) MAPK signaling regulates nitric oxide and NADPH oxidase-dependent oxidative bursts in Nicotiana benthamiana. Plant Cell 20:1390–1406
Besson-Bard A, Pugin A, Wendehenne D (2008) New insights into nitric oxide signaling in plants. Annu Rev Plant Biol 59:21–39
Dean R, Van Kan JA, Pretorius ZA, Hammond-Kosack KE, Di Pietro A, Spanu PD, Rudd JJ, Dickman M, Kahmann R, Ellis J, Foster GD (2012) Top 10 fungal pathogens in molecular plant pathology. Mol Plant Pathol 13(4):414–430
Gilad NL, Bar-Nun N, Noy T, Mayer AM (2000) Enzymes of Botrytis cinerea capable of breaking down hydrogen peroxide. FEMS Microbiol Lett 190:121–126
Glazebrook J (2005) Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu Rev Phytopathol 43:205–227
Götz S, García-Gómez JM, Terol J, Williams TD, Nagaraj SH, Nueda MJ, Robles M, Talón M, Dopazo J, Conesa A (2008) High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res 36:3420–3435
Govrin E, Levine A (2000) The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr Biol 10:751–757
Guo M, Guo W, Chen Y, Dong S, Zhang X et al (2010) The basic leucine zipper transcription factor Moatf1 mediates oxidative stress responses and is necessary for full virulence of the rice blast fungus Magnaporthe oryzae. Mol Plant Microbe Interact 23:1053–1068
Hemetsberger C, Herrberger C, Zechmann B, Hillmer M, Doehlemann G (2012) The Ustilago maydis effector Pep1 suppresses plant immunity by inhibition of host peroxidase activity. PLoS Pathog 8(5):e1002684
Höhl B, Pfautsch M, Barz W (1990) Histology of disease development on resistant and susceptible cultivars of chickpea (Cicer arietinum L.) inoculated with spores of Ascochyta rabiei. J Phytopathol 129:31–45
Kanehisa M, Araki M, Goto S, Hattori M, Hirakawa M, Itoh M, Katayama T, Kawashima S, Okuda S, Tokimatsu T, Yamanishi Y (2008) KEGG for linking genomes to life and the environment. Nucleic Acids Res 36:D480–D484
Karányi Z, Holb I, Hornok L, Pócsi I, Miskei M (2013) FSRD: fungal stress response database. Database (Oxford) 2013:bat037. https://doi.org/10.1093/database/bat037
Kim NH, Kim BS, Hwang BK (2013) Pepper arginine decarboxylase is required for polyamine and γ-aminobutyric acid signaling in cell death and defense response. Plant Physiol 162(4):2067–2083
Kim D, Langmead B, Salzberg SL (2015) HISAT: a fast spliced aligner with low memory requirements. Nat Methods 12(4):357–360
Lambou K, Lamarre C, Beau R, Dufour N, Latge JP (2010) Functional analysis of the superoxide dismutase family in Aspergillus fumigatus. Mol Microbiol 75:910–923
Lehmann S, Serrano M, L’Haridon F, Tjamos SE, Metraux JP (2015) Reactive oxygen species and plant resistance to fungal pathogens. Phytochemistry 112:54–62
Li W, Godzik A (2006) Cd-hit: a fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 22(13):1658–1659
Lyu X, Shen C, Xie J et al (2015) A “footprint” of plant carbon fixation cycle functions during the development of a heterotrophic fungus. Sci Rep 5:12952
Mao D, Luo Y, Cheng M, Zhang J (2005) Functional grouping of yeast genes via biclustering microarray data. Front Biosci 10:2669–2675
Marroquin-Guzman M, Hartline D, Wright JD, Elowsky C, Bourret TJ, Wilson RA (2017) The Magnaporthe oryzae nitrooxidative stress response suppresses rice innate immunity during blast disease. Nat Microbiol 2:17054
Mayer AM, Staples RC, Gil-ad NL (2001) Mechanisms of survival of necrotrophic fungal plant pathogens in hosts expressing the hypersensitive response. Phytochemistry 58:33–41
Nizam S, Singh K, Verma PK (2010) Expression of the fluorescent proteins DsRed and EGFP to visualize early events of colonization of the chickpea blight fungus Ascochyta rabiei. Current Genetics 56(4):391–399
Otte O, Barz W (1996) The elicitor-induced oxidative burst in cultured chickpea cells drives the rapid insolubilization of two cell wall structural proteins. Planta 200:238–246
Oughtred R, Stark C, Breitkreutz BJ, Rust J, Boucher L, Chang C, Kolas N, O’Donnell L, Leung G, McAdam R, Zhang F, Dolma S, Willems A, Coulombe-Huntington J, Chatr-Aryamontri A, Dolinski K, Tyers M (2019) The BioGRID interaction database: 2019 update. Nucleic Acids Res 47:D529–D541
Pertea M, Pertea GM, Antonescu CM, Chang TC, Mendell JT, Salzberg SL (2015) StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat Biotechnol 33:290–295
Petersen TN, Brunak S, von Heijne G, Nielsen H (2011) SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Methods 8(10):785–786
Pietrowska E, Rozalska S, Kazmierczak A, Nawrocka J, Malolepsza U (2015) Reactive oxygen and nitrogen (ROS and RNS) species generation and cell death in tomato suspension cultures-Botrytis cinerea interaction. Protoplasma 252(1):307–319
Pradhan A, Herrero-de-Dios C, Belmonte R, Budge S, Lopez Garcia A, Kolmogorova A et al (2017) Elevated catalase expression in a fungal pathogen is a double-edged sword of iron. PLoS Pathog 13(5):e1006405
Sarkar TS, Biswas P, Ghosh SK, Ghosh S (2014) Nitric oxide production by necrotrophic pathogen Macrophomina phaseolina and the host plant in charcoal rot disease of Jute: complexity of the interplay between necrotroph-host plant interactions. PLoS ONE One 9(9):17
Schmieder R, Edwards R (2011) Quality control and preprocessing of metagenomic datasets. Bioinformatics 27(6):863–864
Schoonbeek H, Del Sorbo G, De Waard MA (2001) The ABC transporter BcatrB affects the sensitivity of Botrytis cinerea to the phytoalexin resveratrol and the fungicide Fenpiclonil. Mol Plant Microbe Interact 14(4):562–571
Seong K, Hou Z, Tracy M, Kistler HC, Xu JR (2005) Random insertional mutagenesis identifies genes associated with virulence in the wheat scab fungus Fusarium graminearum. Phytopathology 95(7):744–750
Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T (2003) Cytoscape: a software environment for integrated models of bimolecular interaction networks. Genome Res 13(11):2498–2504
Singh K, Nizam S, Sinha M, Verma PK (2012) Comparative transcriptome analysis of the necrotrophic fungus Ascochyta rabiei during oxidative stress: Insight for fungal survival in the host plant. PLoS ONE One 7(3):e33128
Skamnioti P, Henderson C, Zhang Z, Robinson Z, Gurr SJ (2007) A novel role for catalase B in the maintenance of fungal cell-wall integrity during host invasion in the rice blast fungus Magnaporthe grisea. Mol Plant Microbe Interact 20:568–580
Tillmann AT, Strijbis K, Cameron G, Radmaneshfar E, Thiel M, Munro CA et al (2015) Contribution of Fdh3 and Glr1 to glutathione redox state, stress adaptation and virulence in Candida albicans. PLoS ONE One 10(6):e0126940
Turrion-Gomez JL, Eslava AP, Benito EP (2010) The flavohemoglobin BCFHG1 is the main NO detoxification system and confers protection against nitrosative conditions but is not a virulence factor in the fungal necrotroph Botrytis cinerea. Fungal Genet Biol 47:484–496
Urban M, Cuzick A, Rutherford K, Irvine A, Pedro H, Pant R, Sadanadan V, Khamari L, Billal SK, Mohanty S, Hammond-Kosack KE (2017) PHI-base: a new interface and further additions for the multi-species pathogen–host interactions database. Nucleic Acids Res 45:D604–D610
Verma S, Gazara RK, Nizam S, Parween S, Chattopadhyay D, Verma PK (2016) Draft genome sequencing and secretome analysis of fungal phytopathogen Ascochyta rabiei provides insight into the necrotrophic effector repertoire. Sci Rep 6:24638
Xia J, Gill EE, Hancock RE (2015) NetworkAnalyst for statistical, visual and network-based meta-analysis of gene expression data. Nat Proto 10(6):823–844
Yang G, Rase MS, Turgeon BG, Yode OC (1996) A polyketide synthase is required for fungal virulence and production of the polyketide T-Toxin. Plant Cell 8:2139–2150
Yoshioka H, Asai S, Kobayashi M (2009) Molecular mechanisms of generation for nitric oxide and reactive oxygen species, and role of the radical burst in plant immunity. Mol Cells 28:321–329
Young MD, Wakefield MJ, Smyth GK, Oshlack A (2010) Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol 11:R14
Acknowledgements
This work is supported by external funds from Department of Biotechnology, Government of India (Project numbers: BT/PR7164/PBD/16/1016/2012 and BT/AGR/CG-Phase II/01/2014).
Author information
Authors and Affiliations
Contributions
PKV conceptualized the idea of manuscript and supervised all the experiments. RM, YS, MS, KS and PM performed all the analysis, interpreted the results and formulated the manuscript. SKS and KP edited, and updated the manuscript. PKV, KS, KK and SV contributed critically by revising the draft provided valuable suggestions, and updating the manuscript for publication.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest. We have also followed the accepted principles of ethical and professional conduct and no animals or humans are involved in this research.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Maurya, R., Singh, Y., Sinha, M. et al. Transcript profiling reveals potential regulators for oxidative stress response of a necrotrophic chickpea pathogen Ascochyta rabiei. 3 Biotech 10, 117 (2020). https://doi.org/10.1007/s13205-020-2107-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13205-020-2107-8