
Abstract
Lactic acid is an intermediate-volume specialty chemical, used in the production of biodegradable polymers and other chemicals. Although lactic acid production process is well established, however, the cost of production is very high. Therefore, in this study; starchy biomass (cassava) was hydrolyzed with in-house enzyme cocktail prepared from Aspergillus foetidus MTCC508 and Bacillus subtilis RA10. Process optimization using Taguchi experimental design helped to optimize the most effective ratio of fungal and bacterial amylase for effective saccharification of cassava. A higher sugar yield of 379.63 mg/gds was obtained under optimized conditions, using 30 U/gds of bacterial enzyme and 90 U/gds of the fungal enzyme at pH 4 within 48 h of saccharification. Among 11 lactic acid bacteria isolated, Lactobacillus fermentum S1A and Lactobacillus farraginis SS3A produced the highest amount of lactic acid 0.81 g/g and 0.77 g/g, respectively, from the cassava hydrolysate. The study proved the potential renewable source of cassava biomass as a source for fermentable sugars that can be fermented to lactic acid with high yield. In future, this cost-effective and environmental-friendly bioprocess can be upscaled for industrial lactic acid production.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Lactic acid bacteria (LAB) are one of the most promising contenders for use in future biorefineries to convert plant-derived biomass into value-added compounds such as lactic acid and γ-amino butyric acid. Lactic acid production has received significant attention, because it can be used as a feedstock for the production of poly-lactic acid (PLA), a polymer useful in medical applications as well as for developing environmental-friendly biodegradable plastics, as a substitute for polymer derived from the petrochemical industry (Alexandri et al. 2019). Lactic acid also finds wide applications as a flavor enhancer, preservative, mainly used in food and pharmaceutical industries (Abdel-Rahman et al. 2013). The global lactic acid market size was valued at USD 2.22 billion in 2017, with the overall consumption of 750 kilotons per year and is anticipated to reach up to 1960.1 kilotons by 2025 with a compound annual growth rate (CAGR) of 12.9% from 2015 (Market research survey 2014). Although chemical synthesis of lactic acid is possible, through hydrolysis of lactonitrile by strong acids, this process yields a racemic mixture of the two D and L isomers (Farooq et al. 2012) along with the generation of large amounts of effluent, which is a great environmental concern; hence, fermentation method is generally preferred (Panesar and Kaur 2015). Few amylolytic lactic acid bacteria like Lactobacillus plantarum, Enterococcus faecium, L. lactis and Pediococcus acidolactici are capable to produce lactic acid using the agro-waste as a carbon source, but due to the high load on their metabolic machinery and difference in the temperature of amylolytic saccharification and fermentation, the lactic acid yield is very low in such fermentations (Unban et al. 2017; Xu et al. 2020). Researchers have used the separate hydrolysis and fermentation for production of xanthan gum from potato extract Bhatia et al. (2015). It has been reported that the cost of raw materials for the fermentative production of lactic acid usually accounts for more than 34% of the total manufacturing costs (Vandenberghe et al. 2018). Thus, the efficiency and economics of lactic acid fermentation is still a problem and the availability of a low-cost substrate plays a vital role in the improvement of the overall economics of the process. Renewable materials such as lignocellulose and starch from agricultural residues and forestry resources are generally considered as attractive feedstocks for the production of lactic acid. For industrial production, the lactic acid market has been mainly dependent on the availability and processing of corn, sugarcane, and cassava. Raw sugar, extracted from sugarcane, is one of the main feedstocks used to manufacture lactic acid and its derivatives, but in succession, it creates food crisis and increases the cost of production.
However, the major bottleneck when utilizing renewable materials is the cost of pretreatment and the lack of hydrolytic machinery in lactic acid-producing strains. Starchy substrates are better suited for lactic acid production, but generally require liquefaction at 80 °C and subsequent use of saccharifying enzymes such as α/β-amylase and glucoamylase to generate reducing sugars (Chu-Ky et al. 2016). Generally, amylases are produced by microbial fermentation of starchy materials as a low-cost substrate (Anto et al. 2006; Mukherjee et al. 2009). Rice husk, wheat bran, and potato waste have been used as a low-cost substrate for amylase production either using amylolytic bacteria (Bacillus strains) or fungi (mainly Aspergillus strains) (Baysal et al. 2003; Shukla and Kar 2006; Asgher et al. 2007). Besides cassava, potato starch, wheat straw, paddy straw, corn stalk, sugar cane bagasse, etc. have been used in separate hydrolysis and fermentation (SHF) to produce saccharified hydrolysate which can be further converted to various platform chemicals, exopolysaccharides and biofuel using different microorganisms (Bhatia et al. 2015). Enzyme contributes approximately 30–35% to the total process cost in SHF which can be reduced using in-house enzyme cocktail (Srivastava et al. 2020).
The study was focussed on development of complete process for conversion of cassava biomass to lactic acid using in-house enzyme cocktail of Bacillus subtilis and Aspergillus foetidus for saccharification and newly isolated lactic acid bacteria Lactobacillus fermentum S1A.
Materials and methods
Raw materials and microorganisms
Different starchy materials, i.e., rice bran, cassava (Manihot esculenta, variety-local selection) and soluble starch, used for enzyme production were acquired from the rice processing unit of ICAR-Indian Agricultural Research Institute (IARI), New Delhi, local market of Kerala and Himedia Laboratories, respectively. Two microbes (A. foetidus MTCC 508 and Bacillus amyloliquefaciens MTCC 610) were procured from MTCC, Chandigarh. A bacterial strain, i.e., B. subtilis RA10, one fungus A. niger SH3, and one actinobacterium Streptomyces sp. SSR-198 used in the present study were collected from the culture collection of Division of Microbiology, ICAR-IARI, New Delhi, India. All other chemicals used in the study were purchased from SRL (India) and Sigma Aldrich Chemicals Pvt Ltd, Delhi, India.
Isolation of lactic acid bacteria and their molecular characterization
Four different soil samples were collected from the model dairy of Karnal, Haryana, India (29° 41′ 8.2644″ North and 76°59′ 25.9692″ East). Different fermented foods samples were also collected from the native people of Himachal Pradesh, (India) for the isolation of Lactic acid bacteria (LAB). Isolation of lactic acid-producing bacteria through sequential methods including enrichment technique, acid production based qualitative screening, followed by quantitative selection based on lactic acid production. Isolation was done using De Man, Rogosa and Sharpe (MRS) medium with 2% w/v glucose as a carbon source at pH 6.5 ± 0.1. Flasks were incubated at 37 °C for 3–4 days under static conditions. Furthermore, samples were serially diluted and spread plated on MRS agar plates containing glucose as a sole carbon source at 37 °C for 2–3 days. Morphologically different colonies were further purified and subcultured on MRS agar slants with 1% CaCO3 and stored at 4 °C. All the isolates were quantitatively checked for lactic acid production using fermentation medium having Yeast extract 0.75%; Manganese sulfate 20 mg/l; Calcium carbonate 1.5% and glucose 5% at pH 6.0 (Panesar et al. 2010). Samples were withdrawn after every 24 h up to 72 h and lactic acid was quantified by High performance liquid chromatography (HPLC) (Waters pump 515 model) equipped with Photo Diode Array (PDA) and Refractive index (RI) detector. The Aminex HPX-87H column was run with 5 mM H2SO4 as the mobile phase at a flow rate of 0.5 ml/min and the oven temperature was kept at 60 °C.
Molecular identification of selected LA producing bacteria was carried out by PCR based 16 s rDNA amplification technique. DNA was isolated using the Zymo Research Fungal/Bacterial DNA MicroPrep™ isolation kit. Isolated genomic DNA was used as a template for 16S rDNA amplification using a universal forward primer (pA: 5′AGAGTTTGATCCTGGCTCAG3′) and reverse primer (pH: 5′AAGGAGGTGATCCAGCCGCA 3′) (Edwards et al. 1989). The amplified PCR products were purified and sequenced by Agrigenome Pvt. Ltd (India). The phylogenetic tree was constructed by the neighbor-joining method as well as by the maximum likelihood method. Phylogenetic analysis was performed using molecular evolutionary genetics analysis (MEGA) software, version 7.0 (Kumar et al. 2016). The gene sequences were submitted to the NCBI GenBank and accession numbers were obtained.
Compositional analysis of substrates
Starch content was estimated in the substrates-cassava, rice bran, and soluble starch using a modified acid hydrolysis method described by Macrae et al. (1974). Briefly, samples (cassava, rice bran, and soluble starch) were moistened with distilled water and incubated in a water bath at 60 °C for 30 min. The samples were incubated in 95% ethanol to remove excess sugar and digested with 1 ml of 1M HCl. The released sugar was estimated by 3,5-dinitrosalicylic acid (DNSA) method (Miller 1959). Total organic carbon and nitrogen were estimated by the method of Navarro et al. (1993) and the Kjeldahl method (Bremne 1960), respectively.
Enzyme production potential of various microorganisms on different starchy waste
Five different microorganisms as mentioned in an earlier section (raw materials and microorganisms) were screened for their amylolytic enzyme production potential on three different starchy waste materials (Cassava, rice bran, and soluble starch). Enzyme production was carried out under submerged condition using YPS medium (Yeast extract 0.4%, K2HPO4 0.1% and MgSO4·7H2O 0.05%, pH 7.0) supplemented with the starchy substrate (1% w/v) as a sole carbon source, 10% (v/v) bacterial inoculum and incubated at 30 °C for 7 days. Bacterial cultures were incubated under shaking condition, while fungal and actinomycete were incubated under static condition. After 7 days, the crude extracellular enzyme was harvested by centrifugation at 10,000 rpm for 10 min at 4 °C. The supernatant was collected and concentrated using chilled acetone precipitation. The precipitate was further resuspended in citrate buffer (pH 5.0) to make it ten times concentrated and stored at − 20 °C. The crude enzyme was assayed for amylase activity by standard protocol (Colowick et al. 1955).
Amylase activity
The concentrated enzyme extract stored at − 20 °C was assayed for its amylase activity. Amylase (EC 3.2.1.1) was assayed by the method of Colowick et al. (1955) using 1% soluble starch as a substrate in 0.05 M citrate buffer (pH 5.0) at 50 °C. The reducing sugars released were estimated using the DNSA method (Miller 1959). One unit of amylase activity was defined as the amount of enzyme releasing 1 μmol of maltose released per ml per minute from soluble starch under the assay condition.
Temperature and pH optimization of enzymes
The promising microbes (A. foetidus MTCC 508, A. niger SH3 and B. subtilis RA10) and substrates (rice bran, cassava, and soluble starch) combinations (listed in Table 4) were selected based on the enzyme activity. Of the fifteen combinations, seven promising combinations with high amylase activities were investigated for their temperature and pH optima by one factor at a time approach. The optimum pH for amylase activity was studied over the pH range of 3.0–7.0 using 0.05 M phosphate and citrate buffer. The temperature optimum was studied over the range of 30–70 °C. Enzyme activity was assayed by the standard methods as described earlier. Based on the highest activity of bacterial and fungal crude enzyme extract, an enzymatic cocktail was developed for saccharification of starchy substrates. Bacterial and fungal enzyme extract having the highest activity was further analyzed for its thermostability at its temperature optima up to 72 h.
Extracellular proteome analysis of A. foetidus MTCC508 and B. subtilis RA10
Sample preparation for proteome analysis
The whole secretome of B. subtilis RA10 and A. foetidus MTCC 508 was collected after incubation in the respective medium as described in Sect. 2.4. The crude enzyme collected was further centrifuged and passed through the 0.22 μm syringe filters. Further, the concentrated secretome of B. subtilis RA10 and A. foetidus having protein content ~ 25 μg were denatured in 8 M urea and 10 mM dithiothreitol followed by alkylation with 25 mM iodoacetamide. Proteins were digested using mass spectrometry (Sorimachi et al. 1997) grade trypsin (G-Biosciences, USA) (1:50) for 3 h. The peptides were desalted and dried by vacuum centrifugation and stored at − 80 °C before MALDI analysis.
LC–MALDI MS/MS spectrometry and data analysis
The digested peptides were extracted using 5% acetonitrile and 0.1% trifluoroacetic acid in water and were injected in the chromolithCapronRP-18e (150–0.1 mm) column. The extracted peptides were spotted with the α-cyano-4-hydroxycinnamic acid matrix in a 1:1 ratio and analyzed for peptide mass fingerprinting spectra of MALDI/MS in ABSCIEX Triple TOF plus 5600 system in reflector positive ion mode.
The data were explored using the online MASCOT search engine (Matrix Science, Boston, MA, USA). Searches were conducted against the fungal and bacterial database available in the NCBI database for detailed annotation. The theoretical values of molecular weight and isoelectric point were obtained using the Compute pI/Mw tool (ExPASY; ExPASY Bioinformatics Resource Portal https://www.expasy.org) according to the predicted amino acid sequences. Carbohydrate-active enzyme annotation was done using the tool dbCAN meta server (Zhang et al. 2018).
Optimization of bacterial and fungal enzyme dosages for cocktail development with starchy waste
Two enzyme extracts produced by B. subtilis RA10 and A. foetidus MTCC 508 were selected for cocktail development using Taguchi experimental design. A total of four factors were selected for the optimization including loading of the fungal enzyme, bacterial enzyme loading, substrate, and pH. A standard L16(44) orthogonal array was used to design the experiment, where L is the Latin square, subscript 16 showing the number of the experiment, and (44) represent four factors with their respective four levels (Table 1). All the experiments were carried out in triplicate.
Cassava was used as substrate and saccharification was carried out at 60 °C (as optimized in an earlier experiment) in a water bath up to 48 h under shaking condition (150 rpm). After saccharification, samples were withdrawn to quantify total sugars (glucose, xylose arabinose) by HPLC. Minitab 16.2.1 software was used to evaluate the results by analysis of variance (ANOVA) and response curve to observe the statistical contribution, the ranking of each factor, and their respective levels. The analysis was performed with “bigger is better” quality for a selected response. The optimized condition suggested by the design of the experiment was again revalidated in a separate experiment.
Lactic acid production using the saccharified cassava hydrolysates by L. fermentum S1A and Lactobacillus farraginis SS3A
Among all the seven LAB isolates, two promising bacterial isolates L. fermentum S1A and L. farraginis SS3A were evaluated for lactic acid production from cassava hydrolysate in shake flasks. Since hydrolysate lacks growth factors, yeast extract (1% w/v) and calcium carbonate (2% w/v) to maintain the desirable pH, was supplemented in the cassava hydrolysate. Fermentation was carried out under static condition at 37 °C for 48 h and samples were withdrawn after at 24 and 48 h. All the samples were analyzed by HPLC to quantify the lactic acid produced by the organisms. A control was also kept using cassava hydrolysate as a carbon source at 37 °C and sugar concentration was analyzed.
Quantification of sugars and lactic acid by HPLC
The samples were analyzed for the presence of sugars and l-lactic acid using high performance liquid chromatography (HPLC) Waters 515 (Waters Corporation, Milford, MA, USA) equipped with a Waters 2414 refractive index (RI) and photodiode array (PDA) detector. Sugars were detected by RI (Glucose RT 8.7 min, Xylose RT 9.4 min, and Arabinose 10.2 min) detector and l-lactic acid (RT 12.2 min) was detected by PDA detector. The column Aminex HPX-87H was operated with 5 mM H2SO4 as a mobile phase at a flow rate of 0.5 ml/min. The oven temperature was kept at 60 °C.
Results and discussion
Isolation of lactic acid bacteria and their molecular characterization
A total of forty-five different morphotypes were isolated from soil and fermented food samples. All the isolates were qualitatively screened on CaCO3 supplemented MRS plate for acid production by visualizing a clear zone. Among them, fifteen isolates were able to produce a clear zone around colonies due to the dissolution of CaCO3, which indicates their capability to produce acid. These isolates were further screened for catalase and oxidase production to confirm their identity, because LAB lacks both the enzymes. Of them, five were found to be catalase-positive and two were oxidase-positive. These fifteen bacterial isolates were also screened for their thermotolerance and it was found that all the fifteen isolates were able to grow at 40 °C but when the temperature was raised to 45 °C, bacterial strains SR1 and PP2A failed to show any growth. Thermotolerant lactic acid bacteria are more suitable for simultaneous saccharification and fermentation (SSF) process, where fermentative microorganism need to work at a higher temperature (≥ 50 °C) to sustain the activity over a longer incubation time (Eiteman and Ramalingam 2015). As the halo zone formation on CaCO3 supplemented MRS plate alone cannot be considered as a confirmatory test for the production of lactic acid, all the thirteen isolates were analyzed for their lactic acid production potential with glucose as a substrate, followed by HPLC based quantification. It was found that among the 15 isolates, 11 were producing lactic acid in variable amounts ranging between 1.19 and 13.98 mg/ml. Based on the confirmation of lactic acid production capacity, 11 isolates were selected and identified by 16S rDNA sequencing. The respective sequences were analyzed by BLAST using the NCBI database and submitted to the GenBank (Table 2). Among 11 identified isolates, 7 isolates belong to the order Lactobacillales, family Lactobacillacea and the rest of the isolates, were identified as members from Bacillus. Lactobacillus is a well-known bacterium for lactic acid production, while the thermotolerance and lactic acid production capacity of Bacillus sp. (Patel et al. 2005) and Pediococcus pentosaceus (RamÍRez-ChavarÍN et al. 2010) have been previously described.
Selection of substrate based on starch content
Starch is the most valuable carbohydrate and as per FAO statistics (FAO) in 2017 (FAO Food and Agriculture Organization of the United Nations. http://www.fao.org), the world’s total production of starchy biomass including rice, wheat, potatoes, and cassava reached 319.786 million tonnes. Hence, cassava, rice bran, and soluble starch (commercial substrate) were used for enzyme production and lactic acid production. All the three substrates were analyzed for starch, total organic carbon (TOC), total nitrogen (N), phosphorous (P), and potassium (K) content. Although botanically, starch is not present in rice bran during the process of milling, rice endosperm gets broken, which contributes to starch content (Fabian et al. 2011). Its quantity varies according to the degree of milling, generally 5–35% starch content could be detected in rice bran (Friedman 2013; Saunders 1985). Comparatively, cassava contains starch content up to 32–42% (Ozoegwu et al. 2017; Wei et al. 2015). Soluble starch had the highest starch content (78.89%) followed by cassava (40.05%) and rice bran (36.21%). Similarly, in terms of total organic carbon content, soluble starch had the highest amount of TOC (57.64%) followed by cassava and rice bran (Table 3). Although, soluble starch had the highest starch content, all-natural starchy substrates have different amylose:amylopectin ratio ranges from 15:85 to 35:65. The difference in mechanical structure tends to induce different hydrolases which can be helpful for making the enzymatic cocktail more diverse and feasible to hydrolyze complex starchy biomass (Cano et al. 2014). As soluble starch and rice bran, are commercial product with many other industrial applications, is a major impeding factor for commercial lactic acid production. Hence, cassava was selected as the substrate for further studies on lactic acid production.
Thermostable amylase production on different starchy waste from various microorganism
Starch consists of mainly α-glucan in the form of amylose and branched amylopectin. The complex and branched structure of starch makes it difficult to be hydrolyzed into its monomeric subunit glucose (Tester et al. 2007). The potential of microbial amylases has been explored widely for saccharification of starchy biomass and in general, bacteria produce α/β amylases, while fungi produce glucoamylase. Both enzymes are needed for the complete hydrolysis of starch. Hence, for this study, different amylolytic microbes (A. foetidus, A. niger SH3, B. subtilis RA10, B. amyloliquifaciens, Streptomyces sp. SSR-198) were tested for their amylase producing capability using starchy biomass, i.e., rice bran, cassava, and soluble starch as substrates (Table 4). As the expression of the inducible enzymes is very much dependent specifically on the substrate (Abd-Elhalem et al. 2015; Balkan and Ertan 2007), the extracellular activity was determined in all the combinations. It was observed that Streptomyces sp. SSR-198 produced substantially less amount of amylase from all the three substrates (maximum 4.25 U/ml on soluble starch). Based on amylase activity in other microbes, it was concluded that A. foetidus MTCC 508 (12.76 U/ml), A. niger SH3 (9.98 U/ml) and B. subtilis RA10 (11.74 U/ml) were better producers of amylases with rice bran as a carbon source. In general, both fungi and bacteria were found to produce more amylase with rice bran than other substrates which might be due to higher nitrogen content in the bran (1.92%) compared to other two substrates. Similar results were reported by some researchers, who have reported that natural starch sources were better inducers for amylase production (Abd-Elhalem et al. 2015; Najafi et al. 2005; Saxena and Singh 2011). The selected crude enzymes produced using rice bran as substrate were analyzed for their pH and temperature optima.
Temperature and pH optima of selected amylase
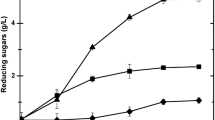
In industries, liquefaction of starch at high temperature is the first requisite before saccharification by bacterial and fungal amylases. These steps increase the overall cost of the process, so the search for thermotolerant amylases fulfils the increasing demand of the biotechnological industries (Suganthi et al. 2015). Temperature is the most important factor which markedly influences the enzyme activity, and the temperature optima of all the selected enzyme preparations were found to be 60 °C. It was also observed that the highest enzyme activity of 17.93 U/ml and 39.81 U/ml were recorded in A. foetidus MTCC 508 and B. subtilis RA10, respectively, at 60 °C, when rice bran was used as a substrate (Fig. 1A). Similarly, pH optima of both the organisms were observed to be pH 5.0 (Fig. 1B). Hence, both the extracts were used for cocktail development, as the fungal strain is also known to produce glucoamylases and bacteria produce α/β amylases.

Temperature [A] and pH [B] optimization of enzyme extracted from different microbial sources grown on various types of starchy biomass for amylase activity
Both the bacterial and fungal enzyme extracts were checked for their thermostability at 60 °C and it was found that residual activity of bacterial extract was stable up to 72 h. Residual activity of fungal amylase slightly decreased by 80% at 48 h and then it remained stable up to 72 h (Supplementary Fig. 1). Bacterial strains produce α or β-amylases and the synergistic combination of both the amylases is essential for effective hydrolysis of starchy biomass like cassava.
LC–MALDI MS/MS spectrometry and data analysis
Based upon the highest enzyme activity, one bacterial (B. subtilis RA10 grown on rice bran) and fungus (A. foetidus MTCC 508 grown on rice bran) were selected for cocktail development. The complete protein profiles of both the secretomes were analyzed using the LC–MS/MS based proteomic approach. The secretome of A. foetidus MTCC 508 and B. subtilis RA10 was induced with 1% rice bran as a sole carbon source which sequentially leads to the production of extracellular amylolytic enzymes (Supplementary Table S1).
Secretome analysis of B. subtilis RA10
In the case of B. subtilis RA10, a total of 310 proteins were detected. These proteins were identified through database searches (NCBInr). The glycosyl hydrolase family proteins present in the secretome were identified based on Carbohydrate-Active Enzyme (CAZy) database. The molecular weight and Isoelectric point (pI) of proteins identified by LC–MS/MS have been indicated in Supplementary Table 2. The results suggested that the molecular weights of the proteins were in the range of 1.7–612 kDa along with isoelectric points ranging between 3.8 and 10.61. About 65% of the proteins detected were acidic (pI 3–7). Among all the secreted proteins, 14 were detected as α- amylases. Along with these 14 α-amylases, other proteins belonging to the glycosyl hydrolases family included 23 cellulases, 8 xylanases, 2 mannosidases, and 1 galactanases. Other than glycosyl hydrolases, different proteins present in secretome were 9 proteases, 5 peptidases, 4 phytases, 3 levansucrase, 9 dehydrogenases, 3 catalases, 4 esterases, 1 chitosanase, 95 structural, and 36 hypothetical proteins (Fig. 2). The α-amylases present in the secretome belongs to the GH13 family having a CBM36 module (Supplementary Table 1). This group of enzymes shares several characteristics such as a (β/α)8 barrel structure, the hydrolysis or formation of glycosidic bonds in α-conformation, and several conserved amino acid residues in the active site. The cellulase present in the secretome belongs to three different GH families GH1, GH5, and GH16 having carbohydrate binding module CBM3 and CBM5 (Fig. 2). In the secretome, one endo-beta-1,4-galactanase belonging to GH53 was also detected, which is specifically produced in the bacterium B. licheniformis (Jers et al. 2017). The vast variety of hydrolytic enzymes detected in the secretome of B. subtilis displayed its unique and potential hydrolytic capacity. It also proves that the extracted hydrolases from B. subtilis can not only be used for starch hydrolysis but also can be used for cellulose or hemicellulose saccharification.

Functional classification of proteins detected in the B. subtilis RA10 secretome analysis by LC–MS/MS
Secretome analysis of A. foetidus MTCC 508
In the case of A. foetidus MTCC 508, a total of 122 proteins were detected. These proteins were identified through database searches (NCBInr). The molecular weights of proteins identified by LC–MS/MS along with their isoelectric point have been shown in Supplementary Table 1. The results suggest that the molecular weights of the proteins were in the range of 9.4–325 kDa and their isoelectric points ranged between 3.5 and 11.35. In the secretome, glucoamylase was detected which is specific to fungi for degradation of starchy biomass. The glucoamylase detected in secretome belongs to GH15 family and CBM20 (Supplementary Table 2). A similar glucoamylase was detected and characterized in A. niger which shows two unique binding sites for SBS (starch-binding site) and cyclodextrin glycosyltransferase (Sorimachi et al. 1997; Melikoglu et al. 2013). Along with glucoamylases, 2 xylanases, 6 catalases, 2 dehydrogenases, other structural and hypothetical proteins were also detected (Fig. 3). Surprisingly, B. subtilis RA10 was found to produce a more versatile array of enzymes, as compared to fungus A. foetidus MTCC 508. However, both endoamylase and glucoamylase synergy is required for conversion of starch to glucose, so crude enzyme extract from both B. subtilis RA10 and A. foetidus MTCC 508 were used for saccharification of the starchy biomass.

Functional classification of proteins detected in the A. foetidus MTCC 508 secretome analysis by LC–MS/MS
Enzyme cocktail development and saccharification optimization
For complete hydrolysis of cassava, a synergistic combination of glucoamylase and α/β-amylase is required. Therefore, the saccharification of cassava was optimized using a combination of fungal and bacterial enzymes, in terms of their dose. Other saccharification parameters were also optimized in the same experiment with cocktail development, for the release of maximum sugar from cassava. A statistical Taguchi experimental design was used to optimize four different factors (loading of the substrate, bacterial enzyme, fungal enzyme, and pH), to maximize the sugar release during the saccharification of cassava without liquefaction. The generated model combined with a set of 16 experiments were performed using cassava as a substrate and saccharification was performed for 48 h at 60 °C under shaking conditions (150 rpm) (Table 5).
ANOVA based resolution of the data was performed by Minitab software and the model was found to be significant (p < 0.05). The recommended and predicted combination of factors, along with respective levels for maximum sugar release, included 90 U/gds of fungal enzyme loading and 30 U/gds of bacterial enzyme loading, cassava biomass (10%), and pH of 4. ANOVA was performed to observe the statistical significance of each factor towards improving the saccharification efficiency (Table 6) which helped to identify the best bacterial and fungal enzyme loading concentration for cocktail development. ANOVA revealed that all the factors showed significant contributions for high sugar release, as the sum of the square of error is lowest as compared with the selected factors. This optimized condition (10% substrate loading, 30 U/gds bacterial enzyme, 90 U/gds fungal enzyme, and pH 4) was experimentally validated resulting in the highest sugar release of 379.63 mg/gds (37.63 mg/ml). Previously, researchers have discussed that the amyloglucosidase from fungal source contains a starch-binding domain (SBD), and exo-enzyme system alone, hence, the starch digestion rate is slower as the available substrate concentration is restricted to the non-reducing end groups of the starch chain (Presecki et al. 2013; Sorndech et al. 2015; Dehabadi et al. 2011). Therefore, the synergism of endo- and exo-enzymes is important for the continuous glucose release. Moreover, the presence of other hydrolytic enzymes like cellulases in the secretome of both the microorganisms helped to release the glucose from cellulose portion of the biomass. Zhang et al. (2013) also displayed that the efficiency of the mixed-enzyme system was more than twice as that of the corresponding value for the exo-enzyme system alone. The saccharification when continued further, released 540 mg/gds of sugars within 72 h.
Lactic acid production using saccharified hydrolysates by L. fermentum S1A and L. farraginis SS3A
Based on the previous experiment, among the 7 isolates, two were producing high titers of lactic acid in synthetic media. These two isolates L. fermentum S1A and L. farraginis SS3A were evaluated for lactic acid production in fermentation medium using cassava hydrolysate as a sole carbon source. It was observed that strains S1A and SS3A yielded 22.74 mg/ml (0.81 g/g) and 21.47 mg/ml (0.77 g/g) l-lactic acid, respectively, within 48 h of fermentation. Table 7 summarizes the comparison of previous reports on the lactic acid production from starchy biomass. This comparison revealed that the yield of lactic acid is comparable with the previous reports and can be further improved by optimizing the fermentation condition. The complete process of lactic acid production is also schematically presented in Fig. 4.

Schematic representation of lactic acid production from cassava biomass
Conclusion
The present work was focussed on the optimization of holistic bioprocess required for lactic acid production from cassava biomass using in-house amylolytic enzyme. From the fungal and bacterial sources explored for higher titer of amylase, A. foetidus MTCC 508 and B. subtilis RA10 were selected. The proteome profile of the selected microoragisms indicated the presence of essential enzymes like glucoamylase and amylase for the complete hydrolysis of starch. Statistical optimization of the enzyme and substrate loading was done to select the best parameter combination that can release maximum glucose (540 mg/gds) from starch. Thermotolerant lactic acid bacteria L. fermentum S1A and L. farraginis SS3A with high lactic acid production potential were characterized and used for lactic acid production from saccharified cassava hydrolysate with a yield of 0.81 g/g and 0.77 g/g, respectively, of substrate. However, further refinement of the process is needed by devising a modified reactor which may be operated under anaerobic conditions to maximize lactic acid yield.
References
Abd-Elhalem BT, El-Sawy M, Gamal RF, Abou-Taleb KA (2015) Production of amylases from Bacillus amyloliquefaciens under submerged fermentation using some agro-industrial by-products. Ann Agric Sci 60(2):193–202. https://doi.org/10.1016/j.aoas.2015.06.001
Abdel-Rahman MA, Tashiro Y, Sonomoto K (2013) Recent advances in lactic acid production by microbial fermentation processes. Biotechnol Adv 31(6):877–902. https://doi.org/10.1016/j.biotechadv.2013.04.002
Alexandri M, Schneider R, Mehlmann K, Venus J (2019) Recent advances in d-lactic acid production from renewable resources: case studies on agro-industrial waste streams. Food Technol Biotechnol 57(3):293–304. https://doi.org/10.17113/ftb.57.03.19.6023
Anto H, Trivedi U, Patel K (2006) α-Amylase production by Bacillus cereus MTCC 1305 using solid-state fermentation. Food Technol Biotech 44(2):241–245
Asgher M, Asad MJ, Rahman SU, Legge RL (2007) A thermostable α-amylase from a moderately thermophilic Bacillus subtilis strain for starch processing. J Food Eng 79(3):950–955. https://doi.org/10.1016/j.jfoodeng.2005.12.053
Balkan B, Ertan F (2007) Production of α-Amylase from Penicillium chrysogenum under solid-state fermentation by using some agricultural by-products. Food Technol Biotechnol 45(4):439–442
Baysal Z, Uyar F, Aytekin Ç (2003) Solid state fermentation for production of α-amylase by a thermotolerant Bacillus subtilis from hot-spring water. Process Biochem 38(12):1665–1668. https://doi.org/10.1016/S0032-9592(02)00150-4
Bhatia SK, Kumar N, Bhatia RK (2015) Stepwise bioprocess for exopolysaccharide production using potato starch as carbon source. 3 Biotech 5:735–739. https://doi.org/10.1007/s13205-014-0273-2
Bremne J (1960) Method, determination of nitrogen in soil by the Kjeldahl. J Agric Sci 55:11–33. https://doi.org/10.1017/S0021859600021572
Cano A, Jiménez A, Cháfer M, Gónzalez C, Chiralt A (2014) Effect of amylose:amylopectin ratio and rice bran addition on starch films properties. Carbohydr Polym 111:543–555. https://doi.org/10.1016/j.carbpol.2014.04.075
Chu-Ky S, Pham T-H, Bui K-LT, Nguyen T-T, Pham K-D, Nguyen H-DT, Luong H-N, Tu V-P, Nguyen T-H, Ho P-H, Le T-M (2016) Simultaneous liquefaction, saccharification and fermentation at very high gravity of rice at pilot scale for potable ethanol production and distillers dried grains composition. Food Bioprod Process 98:79–85. https://doi.org/10.1016/j.fbp.2015.10.003
Colowick SP, Kaplan NO, McCormick DB, Wright LD (1955) Methods in enzymology, vol 1. Academic press, New York
Dehabadi VA, Opwis K, Gutmann J (2011) Combination of acid-demineralization and enzymatic desizing of cotton fabrics by using industrial acid stable glucoamylases and α-amylases. Starch Stärke 63(12):760–764. https://doi.org/10.1002/star.201100028
Edwards U, Rogall T, Blöcker H, Emde M, Böttger EC (1989) Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res 17(19):7843–7853. https://doi.org/10.1093/nar/17.19.7843
Eiteman MA, Ramalingam S (2015) Microbial production of lactic acid. Biotechnol Lett 37(5):955–972. https://doi.org/10.1007/s10529-015-1769-5
ExPASY Bioinformatics Resource Portal https://www.expasy.org/. Accessed Apr 2019
FAO Food and Agriculture Organization of the United Nations. http://www.fao.org. Accessed Feb 2019
Fabian C, Ayucitra A, Ismadji S, Ju Y-H (2011) Isolation and characterization of starch from defatted rice bran. J Taiwan Inst Chem Eng 42(1):86–91. https://doi.org/10.1016/j.jtice.2010.03.013
Farooq U, Anjum F, Zahoor T, Rahman S, Randhawa M, Ahmed DA, Akram K (2012) Optimization of lactic acid production from cheap raw material: sugarcane molasses. Pak J Bot 44:333
Freire AL, Ramos CL, Schwan RF (2015) Microbiological and chemical parameters during cassava based-substrate fermentation using potential starter cultures of lactic acid bacteria and yeast. Food Res Int 76:787–795. https://doi.org/10.1016/j.foodres.2015.07.041
Friedman M (2013) Rice brans, rice bran oils, and rice hulls: composition, food and industrial uses, and bioactivities in humans, animals, and cells. J Agric Food Chem 61(45):10626–10641. https://doi.org/10.1021/jf403635v
Jers C, Strube ML, Cantor MD, Nielsen BKK, Sørensen OB, Boye M, Meyer AS (2017) Selection of Bacillus species for targeted in situ release of prebiotic galacto-rhamnogalacturonan from potato pulp in piglets. Appl Microbiol Biotechnol 101(9):3605–3615. https://doi.org/10.1007/s00253-017-8176-x
Kumar S, Stecher G, Tamura K (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol 33(7):1870–1874. https://doi.org/10.1093/molbev/msw054
MacRae J, Smith D, McCready R (1974) Starch estimation in leaf tissue—a comparison of results using six methods. J Sci Food Agric 25(12):1465–1469. https://doi.org/10.1002/jsfa.2740251206
Market Research Survey (2014) Lactic Acid And Poly Lactic Acid (PLA) Market analysis by application (packaging, agriculture, transport, electronics, textiles) and segment forecasts to 2020. Grand View Research Inc
Melikoglu M, Lin CSK, Webb C (2013) Stepwise optimisation of enzyme production in solid state fermentation of waste bread pieces. Food Bioprod Process 91(4):638–646. https://doi.org/10.1016/j.fbp.2013.04.008
Miller G (1959) Determination of reducing sugar by DNS method. Anal Chem 31:426–428
Mukherjee AK, Borah M, Rai SK (2009) To study the influence of different components of fermentable substrates on induction of extracellular α-amylase synthesis by Bacillus subtilis DM-03 in solid-state fermentation and exploration of feasibility for inclusion of α-amylase in laundry detergent formulations. Biochem Eng J 43(2):149–156. https://doi.org/10.1016/j.bej.2008.09.011
Najafi MF, Deobagkar D, Deobagkar D (2005) Purification and characterization of an extracellular α-amylase from Bacillus subtilis AX20. Protein Expr Purif 41(2):349–354. https://doi.org/10.1016/j.pep.2005.02.015
Navarro A, Cegarra J, Roig A, Garcia D (1993) Relationships between organic matter and carbon contents of organic wastes. Bioresour Technol 44(3):203–207. https://doi.org/10.1016/0960-8524(93)90153-3
Ozoegwu C, Eze C, Onwosi C, Mgbemene C, Ozor P (2017) Biomass and bioenergy potential of cassava waste in Nigeria: estimations based partly on rural-level garri processing case studies. Renew Sust Energ Rev 72:625–638. https://doi.org/10.1016/j.rser.2017.01.031
Panesar PS, Kaur S (2015) Bioutilisation of agro-industrial waste for lactic acid production. Int J Food Sci Technol 50(10):2143–2151. https://doi.org/10.1111/ijfs.12886
Panesar PS, Kennedy JF, Knill CJ, Kosseva M (2010) Production of L (+) lactic acid using Lactobacillus casei from whey. Braz Arch Biol Technol 53(1):219–226. https://doi.org/10.1590/S1516-89132010000100027
Patel MA, Ou MS, Ingram LO, Shanmugam KT (2005) Simultaneous saccharification and co-fermentation of crystalline cellulose and sugar cane bagasse hemicellulose hydrolysate to lactate by a thermotolerant acidophilic Bacillus sp. Biotechnol Progr 21(5):1453–1460. https://doi.org/10.1021/bp0400339
Presecki AV, Blazevic ZF, Vasic-Racki D (2013) Complete starch hydrolysis by the synergistic action of amylase and glucoamylase: impact of calcium ions. Bioprocess Biosyst Eng 36(11):1555–1562. https://doi.org/10.1007/s00449-013-0926-2
RamÍRez-ChavarÍN NL, Wacher-Rodarte C, PÉRez-Chabela ML (2010) Characterization and identification of therrmotolerant lactic acid bacteria isolated from cooked sausages as bioprotective cultures. J Muscle Foods 21(3):585–596. https://doi.org/10.1111/j.1745-4573.2009.00206.x
Saunders R (1985) Rice bran: composition and potential food uses. Food Rev Int 1(3):465–495. https://doi.org/10.1080/87559128509540780
Saxena R, Singh R (2011) Amylase production by solid-state fermentation of agro-industrial wastes using Bacillus sp. Braz J Microbiol 42(4):1334–1342. https://doi.org/10.1590/S1517-83822011000400014
Shukla J, Kar R (2006) Potato peel as a solid state substrate for thermostable α-amylase production by thermophilic Bacillus isolates. World J Microbiol Biotechnol 22(5):417–422. https://doi.org/10.1007/s11274-005-9049-5
Sorimachi K, Gal-Coëffet M-FL, Williamson G, Archer DB, Williamson MP (1997) Solution structure of the granular starch binding domain of Aspergillus niger glucoamylase bound to β-cyclodextrin. Structure 5(5):647–661. https://doi.org/10.1016/S0969-2126(97)00220-7
Sorndech W, Meier S, Jansson AM, Sagnelli D, Hindsgaul O, Tongta S, Blennow A (2015) Synergistic amylomaltase and branching enzyme catalysis to suppress cassava starch digestibility. Carbohydr Polym 132:409–418. https://doi.org/10.1016/j.carbpol.2015.05.084
Srivastava N, Mishra PK, Upadhyay SN (2020) 10—enzymatic conversion of lignocellulosic biomass for biofuel production: an overview. In: Srivastava N, Mishra PK, Upadhyay SN (eds) Industrial enzymes for biofuels production. Elsevier, Oxford, pp 205–216. https://doi.org/10.1016/B978-0-12-821010-9.00010-3
Suganthi C, Mageswari A, Karthikeyan S, Gothandam KM (2015) Insight on biochemical characteristics of thermotolerant amylase isolated from extremophile bacteria Bacillus vallismortis TD6 (HQ992818). Microbiology 84(2):210–218. https://doi.org/10.1134/S0026261715020162
Tester RF, Karkalas J, Qi X (2007) Starch structure and digestibility Enzyme-Substrate relationship. Worlds Poult Sci J 60(2):186–195. https://doi.org/10.1079/WPS200312
Trakarnpaiboon S, Srisuk N, Piyachomkwan K, Yang ST, Kitpreechavanich V (2017) L-Lactic acid production from liquefied cassava starch by thermotolerant Rhizopus microsporus: characterization and optimization. Process Biochem 63:26–34. https://doi.org/10.1016/j.procbio.2017.08.019
Unban K, Kanpiengjai A, Takata G, Uechi K, Lee WC, Khanongnuch C (2017) Amylolytic enzymes acquired from L-lactic acid producing Enterococcus faecium K-1 and improvement of direct lactic acid production from cassava starch. Appl Biochem Biotechnol 183:155–170. https://doi.org/10.1007/s12010-017-2436-1
Vandenberghe LPS, Karp SG, de Oliveira PZ, de Carvalho JC, Rodrigues C, Soccol CR (2018) Chapter 18—solid-state fermentation for the production of organic acids. In: Pandey A, Larroche C, Soccol CR (eds) Current developments in biotechnology and bioengineering. Elsevier, Oxford, pp 415–434. https://doi.org/10.1016/B978-0-444-63990-5.00018-9
Vishnu C, Seenayya G, Reddy G (2002) Direct fermentation of various pure and crude starchy substrates to L (+) lactic acid using Lactobacillus amylophilus GV6. World J Microbiol Biotechnol 18:429–433. https://doi.org/10.1023/A:1015526221744
Wei M, Zhu W, Xie G, Lestander TA, Xiong S (2015) Cassava stem wastes as potential feedstock for fuel ethanol production: a basic parameter study. Renew Energy 83:970–978. https://doi.org/10.1016/j.renene.2015.05.054
Xu Y, Zhou T, Tang H, Li X, Chen Y, Zhang L, Zhang J (2020) Probiotic potential and amylolytic properties of lactic acid bacteria isolated from Chinese fermented cereal foods. Food Control 111:107057. https://doi.org/10.1016/j.foodcont.2019.107057
Zhang B, Dhital S, Gidley MJ (2013) Synergistic and antagonistic effects of α-amylase and amyloglucosidase on starch digestion. Biomacromol 14(6):1945–1954. https://doi.org/10.1021/bm400332a
Zhang H, Yohe T, Huang L, Entwistle S, Wu P, Yang Z, Busk PK, Xu Y, Yin Y (2018) dbCAN2: a meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res 46(W1):W95–W101. https://doi.org/10.1093/nar/gky418
Acknowledgements
Authors are thankful to Indian Council of Agricultural Research-National Agricultural Science Fund, India (ICAR-NASF), ICAR-IARI for funding the research activities for lactic acid project (Grant no. NASF/AE-6017/20216-17). AKC is grateful to the CAPES-Brazil for the financial assistance (Process USP number 15.1.1118.1.0).
Author information
Authors and Affiliations
Contributions
AS, KP: performed the experiments, writing the draft manuscript; SS: Planning of this work, performed HPLC experiments, editing of manuscript; SKK conceived of the presented idea; LN supervised the findings of this work, editing of manuscript; AKC Carry out LCMS experiment; PKSN Statistical analysis.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest.
Ethical statement
This article does not contain any studies involving animals performed by any of the authors.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Sharma, A., Pranaw, K., Singh, S. et al. Efficient two-step lactic acid production from cassava biomass using thermostable enzyme cocktail and lactic acid bacteria: insights from hydrolysis optimization and proteomics analysis. 3 Biotech 10, 409 (2020). https://doi.org/10.1007/s13205-020-02349-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13205-020-02349-4