
Abstract
Arbuscular mycorrhizal fungi (AMF) from the rhizosphere of the endemic Laurisilva tree, Picconia azorica, were characterised at two sites in each of two Azorean islands (Terceira and São Miguel). Forty-six spore morphotypes were found, and DNA extraction was attempted from individual spores of each of these. DNA was obtained from 18 of the morphotypes, from which a 1.5 kb long fragment of the nuclear ribosomal RNA gene (SSU-ITS-LSU) was sequenced. A total of 125 AMF sequences were obtained and assigned to 18 phylotypes. Phylogenetic analysis revealed sequences belonging to the families, Acaulosporaceae, Archaeosporaceae, Claroideoglomeraceae, Gigasporaceae and Glomeraceae. Phylotype richness changed between islands and between sampling sites at both islands suggesting that geographical and historical factors are determinant in shaping AMF communities in native forest of Azores. Ecological analysis of the molecular data revealed differences in AMF community composition between islands. In Terceira, the rhizosphere of P. azorica was dominated by species belonging to Acaulosporaceae and Glomeraceae, while São Miguel was dominated by members of Glomeraceae and Gigasporaceae. This is the first molecular study of AMF associated with P. azorica in native forest of the Azores. These symbiont fungi are key components of the ecosystem. Further research is needed to develop their use as promoters of plant establishment in conservation and restoration of such sites.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Picconia azorica (Oleaceae), an endangered endemic woody plant of native Laurisilva in the Azores, is present in coastal and medium altitude forests, coastal cliffs, ravines, lava flows, coastal scrubland (Erica, Morella, mixed) and natural forests (Morella, Laurus) in all the islands except Graciosa (Cardoso et al. 2008; Silva et al. 2009; dos Santos et al. 2010). Used for furniture construction and religious statuary, it has been over-exploited to near-extinction in some islands and is a priority Azorean endemic species for conservation, listed as endangered (EN B1 + 2c) on the IUCN Red List 2004, and in Annex II of the EC Habitats Directive. The tree’s small populations and poor seed production necessitate development of alternative propagation methods to supply saplings for restoration of depleted natural populations, particularly where invasive species preclude natural regeneration.
Arbuscular mycorrhizal fungi (AMF) can be helpful during sapling production, as these ubiquitous fungi form symbiotic associations with a wide diversity of plants, including P. azorica. In these symbiotic associations the obligately biotrophic fungi provide their hosts with minerals, nutrients (e.g., particularly P) and water (Wu et al. 2013), and obtain carbon from the plant (Smith and Read 2008). AMF also have been shown to increase host resistance to biotic and abiotic stresses, including pathogens (Ismail and Hijri 2012), water stress (Boyer et al. 2015) and environmental pollutants (Hildebrandt et al. 2007). Because of these effects on plant and soil performance, AMF can be used to benefit in reclamation and restoration of degraded ecosystems (Ferrol et al. 2004; Middleton et al. 2015). Mycorrhizas therefore should receive special attention in indigenous tree seedling production and restoration.
The diversity of AMF potentially influences plant fitness, community structure, biodiversity and ecosystem variability (van der Heijden et al. 1998). Moreover different AMF can simultaneously colonise a single root segment, and species are functionally diverse (Leake et al. 2004; Helgason et al. 2007; de Oliveira and de Oliveira 2010). To understand ecological plant–fungus interactions in natural ecosystems, it is necessary to identify the fungal partners, and this can be achieved through both morphological and molecular characterization (Helgason et al. 2002; Verbruggen et al. 2010).
There are approximately 300 described species in the Glomeromycota ( http://amf-phylogeny.com and www.indexfungorum.org), although the mycorrhizal status of many of these is still unknown. There are also many more sequences in public databases (GenBank; NCBI, etc.) attributed to glomeromycotan fungi from ‘environmental samples’, which consequently have not been linked with an identified fungal organism, and therefore cannot yet be named to species (Krüger et al. 2009; Öpik et al. 2009; Stockinger et al. 2010). Traditional studies on AMF diversity are based mainly on spore morphology (Walker 1992; Oehl et al. 2010), but identification of AMF spores collected directly from the field is difficult and can be unreliable due to degradation or parasitism by other organism and because of morphological similarity to closely related species. Because AMF are obligate symbionts, they cannot be cultivated in the absence of a phytobiont. Usually, ‘trap plants’ are established in unsterilised soil, or with spores extracted from soil to establish cultures of these fungi (Gilmore 1968). Spores from the resultant cultures are then used for isolation and propagation of single species cultures which can then be used for identification and scientific study. Such cultures can then be used as ‘stock’ to initiate cultures for commercial application. For ecological studies, this indirect culture strategy is time consuming and error-prone, and may reveal only a subset of species present in the studied ecosystem (Jansa et al. 2002).
Molecular techniques make it feasible to identify AMF directly from plant roots, soil and spores. Several PCR-based methods have been developed over recent years, the majority of which target the nuclear ribosomal RNA (rDNA) gene (Schüßler et al. 2001; Clapp et al. 2002; Krüger et al. 2009, 2012). Phylogenetic inferences from rDNA regions can be used for AMF species characterisation and recognition (Stockinger et al. 2009). The ITS region of the nuclear RNA cistron, was proposed as the DNA barcode region for Fungi (Schoch et al. 2012), but it has been shown that for AMF an extended DNA barcode, including both ITS and LSU regions, is needed to discriminate closely related species (Stockinger et al. 2010; Schoch et al. 2012).
Although molecular techniques are gaining prominence, identification approaches based on morphological criteria are still important and can complement molecular methods (Brundrett and Ashwath 2013). Indeed, knowing only the phylotypes without having established cultures of AMF is of little use for application as plant growth promoters. Combining morphological and molecular data provides complementary verification of species (Redecker et al. 2013).
In this study glomeromycotan fungal communities were surveyed from the rhizosphere of P. azorica by morphological and molecular methods. Native forests from two islands, Terceira and São Miguel, were sampled to observe patterns of distribution of AMF species and phylotypes to provide baseline information for later use in establishing strategies for conservation of P. azorica in particular, and native Azorean forests in general.
2 Material and methods
2.1 Study sites
This study was conducted in two Islands of the Azorean archipelago, São Miguel (4.01 Myr BP) and Terceira (3.52 Myr BP). The sampling localities were from four populations of P. azorica: Terra Brava (TB) and Serreta (TS), from Terceira and Lombo Gordo (SG) and Ribeira Quente (SR) from São Miguel. Terra Brava, located at 38°43′51.87″N; 27°11′36.40″W (Fig. 1), is included in the very wet Laurisilva at 650 m of altitude. This ecosystem is dominated by endemic woody plants, principally Laurus azorica, Ilex perado, Frangula azorica, Vaccinium cylindraceum, Erica azorica, Myrsine africana, and occasionally by Juniperus brevifolia and P. azorica. The Laurisilva of Serreta (38°45′42.54″N; 27°22′03.61″W) is the low altitude type (95 m), characterised by a low diversity of plants, dominated by Morella faya and P. azorica, and occasionally by L. azorica. These forests are located in the most thermophilic areas of Azores and are almost extinct (Dias 1996). The highest canopy is dominated by a dense cover of Pittosporum undulatum, and rarely by L. azorica. This forest is mixed with other invasive woody species including Metrosideros excelsa, Eucalyptus globules, Acacia melanoxylon, Sphaeropteris cooperi, Fuchsia magellanica and Rubus inermis. The herbaceous stratum is dominated by Dryopteris azorica, Hedera helix var. azorica, Smilax aspera and Gomphocarpus fruticosus (Dias 1996). Lombo Gordo (37°47′10.47″N; 25°8′35.94″W) is covered by a coastal scrubland where P. azorica dominates in certain areas, but is mixed with other native and invasive woody species including M. faya, E. azorica, P. undulatum, Arundo donax, Hedychium gardnerianum and Phormium tenax (Martins et al. 2011). Ribeira Quente (37°44′26.65″N; 25°18′7.31″W) is also a coastal scrubland dominated by the endemic plants L. azorica and P. azorica, but also associated with other native and invasive woody species such as P. undulatum, M. Faya, and A. melanoxylon.

Location of Picconia azorica native forest of Terceira (a) (Terra Brava - TB and Serreta - TS) and São Miguel (b) (Lombo Gordo - SG and Ribeira Quente - SR)
3 Sample collection
Ten soil samples with associated roots were collected on each site (Terceira - TB; TS and São Miguel - SG; SR) at each of three different sampling times, September 2012 (autumn 2012), May 2013 (spring 2013) and September 2013 (autumn 2013) resulting in a total of 120 samples. The distance between samples taken on each site was at minimum 25 m, and the distance between sample sites was about 25 km in Terceira, and 30 km in São Miguel. Soil samples, to a depth of up to 30 cm, to ensure feeder roots were collected, were taken as described in Melo et al. (2017). Picconia roots could be recognised by their gross morphology, and other roots in the sample were discarded at the collection location.
4 Establishment of trap cultures
Two open-pot trap cultures were established from soil samples collected at each sampling time, one with 1-week-old Zea mays seedlings (Melo et al. 2014) and another one with a micropropagated P. azorica plant from a clone of the in vitro collection of CBA-UAc, previously acclimated for two months in the greenhouse (Mendonça et al. 2015). Two seedlings of Z. mays or one rooted explant of P. azorica were transplanted to each pot, containing 1.5 kg of fresh soil sampled from each site mixed with autoclaved volcanic soil” bagacina” (2:1 v/v). Ten pots per sampling site were established, resulting in 240 open-pot cultures (4 sites × 10 samples/site × 3 sampling dates × 2 trap cultures). All pots were maintained in a greenhouse for five months, watered every two days solely with distilled water.
5 Spore extraction and morphological identification
Glomeromycotan spores were extracted from 50 g of air-dried soil of each sample from field soil and trap cultures by wet sieving and sucrose centrifugation (Walker 1992), and stored at 4 °C in autoclaved water pending examination. Different spore types were initially separated under a stereo-microscope, and then examined through a compound microscope on slides in a 1:1 mixture of polyvinyl alcohol lacto-glycerol (PVLG) and Melzer’s reagent. They were classified into known species or undetermined spore morphotypes based on colour, size, surface ornamentation, hyphal attachment, reaction to Melzer’s reagent, and wall structure. Once the different spore types had been determined, from a total number of 732 specimens on PVLG slides, they could be identified by their characteristics under a dissecting microscope.
6 Molecular identification
Molecular characterisation, including DNA extraction, PCR, cloning, RFLP, sequencing and phylogenetic analyses are as described in Melo et al. (2017). Sequences have been deposited in GenBank (accession numbers KP756414 to KP756538). One spore of each morphotype was characterised for each different soil sample. Sequences were aligned by hand to the backbone after Krüger et al. (2012). This alignment was used to calculate the full phylogenetic tree with RAxML (Stamatakis 2014) and can be found in the supporting information (Fig. S1). The phylogenetic positions of the individual sequences are termed “phylotypes”.
Additionally, sequences were grouped into operational taxonomical units (OTUs) testing different sequence similarity threshold (97, 98 and 99%) using UPARSE (Edgar 2013) in the SEED workbench (Větrovský and Baldrian 2013). The most abundant sequence per OTU was chosen as representative and these sequences were automatically aligned by PyNAST (Caporaso et al. 2010a) to the phylogenetic backbone alignment in the Qiime pipeline (Caporaso et al. 2010b). Followed by placing them into the phylogenetic tree with the Evolutionary Placement Algorithm (EPA) for short sequence reads (Berger et al. 2011) implemented in QIIME (Fig. S2).
7 Statistical analysis
The structure of the spore communities in the rhizosphere of P. azorica were analysed from the 18 AMF phylotypes determined. Phylotype richness was calculated as the number of phylotypes per sample in each field site. The sampling followed a hierarchical nested design, with field sites nested within each island. Variation in phylotypes richness between islands and field sites within each island were analysed by Nested ANOVA followed by a Tukey honestly significant difference test at p < 0.05 (MINITAB Inc. 2000). All data were tested for normality and fulfilled the assumptions of Nested ANOVA. To investigate differences in AMF diversity between the four sampling sites we used the phylotype occurrence (number of samples in which each phylotype occurred in each field site) to calculated the Shannon Wiener index (H′) (Magurran 2004). using the software package ‘Species Diversity and Richness IV’ (see http://www.pisces-conservation.com) (Henderson and Seaby 2002). To understand better how the different AMF phylotypes commonly found in rhizosphere of P. azorica were related to habitat, the phylotype occurrence was analysed by correspondence analyses (CA) using CANOCO for Windows (ter Braak and Smilauer 2002).
8 Results
8.1 Glomeromycotan spore types
Forty-six distinct morphotypes from 10 families and 12 genera were detected, including nine different undetermined glomoid morphotypes (Table 1).
8.2 Molecular identification
From the 46 glomeromycotan taxa (distinguishable morphotypes) in the rhizosphere of P. azorica, 18 were molecularly identified, and 125 sequences derived from this study were deposited in the EMBL database (Table 2) which were classified in 18 phylotypes, approximately equating to species.
The 125 AMF sequences from spores were grouped either into 35, 48 or 65 OTUs based on 97, 98 or 99% sequence similarity (Table S1, Fig. S1); or assigned to 18 phylotypes based on phylogenetic analysis (Fig. 2). DNA sequences of 18 glomeromycotan morphotypes (SSU-ITS-LSU) showed similarities to Glomeromycota sequences when BLASTed against the NCBI database, having thus been included in the phylogenetic analysis, and could be linked to 18 phylotypes. For the Acaulosporaceae, sequences of the spore morphotypes Ac. brasiliensis, Ac. koskei, Ac. lacunosa and Ac. laevis were clustered with previously published sequences of these glomeromycotan species with bootstrap supports (BS) of 78, 98, 42 and 100% respectively (Fig. 2). Sequences of the spore morphotype Ac. mellea cluster together with an ‘Ac. mellea’ sequence (FJ009670) and sister to Ac. lacunosa. A BLAST search revealed that ‘Ac. mellea’ (FJ009670) is most probably an Ac. lacunosa, indicating that the published species identification (Yen et al. 2006) is incorrect. The morphotype Ac. cavernata sequences clustered in a separated clade (100% BS) sister to Ac. cavernata BEG33, W3293/Att209-37 and Acaulospora sp. W2941/Att869-3 with 80% BS. Nevertheless, Acaulospora sp.1 and Acaulospora sp.2 each formed a separate clade in Acaulosporaceae sister to the Ac. brasiliensis- Ac. alpina-clade, both supported by 100% BS.

Collapsed maximum likelihood phylogenetic tree, spanning a part of the SSU, the full ITS1-5.8S-ITS2 region and a large part of the nuclear LSU rRNA gene region (ca. 1.5 kb long), including the AMF sequences found in Picconia azorica marked in blue. For clearer presentation, clades were collapsed where possible, BS below 60% are not shown and for main branches they were reduced to polytomies. Full phylogenetic tree is provided as supplementary (Fig. S2). Scale bar shows number of substitution per site
Regarding Archaeosporaceae, the morphotype Archaeospora sp. clustered sister to Archaeospora at the genus-level (79% BS). In Diversisporaceae, the sequences of the morphotype Diversispora sp.1 clustered within Diversispora sp. W3033/Att669-1, while Diversispora sp.2 formed a separated clade in Diversisporaceae supported by 95% BS, sister to Diversispora sp. W3033/Att669-1 (an undescribed species from Iceland) both species clustering with 94% BS into Diversispora.
Relative to Glomeraceae, the morphotype defined as Glomus sp.1 falls within the genus Glomus, but as sister to Gl. macrocarpum with 94% (BS), and in its own clade with 99% BS. Sequences of the morphotype Glomeraceae sp. with 99% BS form a separate branch sister to an uncultured Rhizophagus sp. from a root sample. The morphotype Rhizophagus sp. clustered within a clade together with a Rhizophagus sp. VAh1_2, sister to Rh. fasciculatus, supported by 66% BS.
Sequences of the morphotype Glomus sp.2 formed a separated clade in Glomeraceae supported by 100%, sister to the Gl. macrocarpum. In relation to Claroideoglomeraceae, the sequences of the morphotype Cl. claroideum clustered with 100% BS to Cl. claroideum (AY639184). In Gigasporaceae, sequences of the morphotype Gi. margarita clustered within (60% BS) the Gi. margarita BEG34-clade, while sequences of the morphotype Scutellospora sp. form a separate clade with 96% and 95% BS, sister to Sc. spinosissima W3009/Att664-1, Sc. calospora BEG32 and Sc. aurigloba WUM53 in the Scutellospora (72% BS).
By implementing the different OTUs (97, 98, 99% similarity) into the AMF backbone after Krüger et al. (2012) we could show that, independent of applied threshold, the phylogenetic assignment resulted in the same 16 phylotypes over all threshold-levels analysed. These 16 phylotypes, namely two Diversispora sp., Ac. laevis, Ac. brasiliensis, Ac. lacunosa, Ac. cavernata, Gi. margarita, Scutellospora sp., two Rhizophagus sp., Rh. irregularis, Sclerocystis sp., Gl. macrocarpum, Glomus sp., Cl. claroideum and Ambispora sp., could be restored through all phylogenetic trees (Fig. S1).
Comparing the results of the automatically-aligned (EPA tool, OTUs) and manually-aligned (phylotypes) analyses only 16 out of 18 AMF taxa could be restored, and taxonomical classification differs between phylotypes and OTUs. Two main differences were found: Ac. koskei could not be restored from the OTU analysis and clusters together with some other Acaulospora sp. in a single OTU, the Archaeospora sp. (phylotype) also now clusters in Ambispora (OTU). Sequences of both species were difficult to align with already-sequenced AMF (Krüger et al. 2012) and therefore may cluster differently dependent on the alignment method (manual or automatic) (Table S1).
8.3 AMF richness and distribution patterns in the rhizosphere of P. azorica
AMF richness (based on richness of phylotypes) change between Terceira and São Miguel (Nested Anova: F1.2 = 19.73, p < 0.001) and between sites within each island (Nested Anova: F2.4 = 6.21, p = 0.003). The average number of phylotypes was higher in samples of Terceira than is São Miguel (Fig. 3a). The phylotype richness was similar in both sites of Terceira (TB and TS) but different between sampled sites SR and SG on São Miguel (Fig. 3b). The phylotype richness was highest in TB and TS, and lowest in SG (Fig. 3b). Phylotype diversity expressed by Shannon-Wiener index was highest in TB (H′ = 2.43) and TS (H′ = 2.20) followed by SR (H′ = 2.05) and SG (H′ = 2.00).

a Phylotype richness (mean ± SE, N = 60) per 50 g of soil in each island (Terceira; São Miguel); and b Phylotype richness in each field site within each island (mean ± SE, N = 30). Different letters above each bar indicate significant differences in phylotype richness between field sites (Tukey test, P < 0.05)
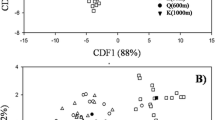
Community composition based on the 18 phylotypes changed between both islands (CA, Fig. 4). The first axis clearly separated the P. azorica forests of Terceira (TB; TS) from the São Miguel forests (SG; SR). The frequency of phylotypes belonging to each AMF clade was different in rhizosphere of P. azorica from that in Terceira and São Miguel (Fig. 5). Species of Acaulosporaceae and Glomeraceae were dominant in the rhizosphere of P. azorica from Terceira, while in the São Miguel rhizosphere, P. azorica was dominated by species belonging to Glomeraceae and Gigasporaceae. Moreover, some AMF phylotypes were found only in a specific field site. Acaulospora sp.1, Acaulospora sp.2 and Rhizophagus sp.1 were morphologically and molecularly identified only in TB and Archaeospora sp. Only in TS.

Correspondence analysis of different native forests from Terceira (TB and TS) and São Miguel (SG and SR) according to the number of AMF phylotypes. Phylotypes are positioned along the first two CA axes, where eigenvalues are 0.572 for CA1 and 0.402 for CA2

Frequency of phylotypes observed in each AMF clades in native forest from Terceira and São Miguel
9 Discussion
Morphological characterisation of the AMF spore community in the rhizosphere soil of Picconia azorica resulted in 46 distinguishable morphotypes, a similar number to the findings of previous studies in the Azores (Melo et al. 2014, 2017). By molecular analysis of representative spores from each morphotype, 18 different phylotypes could be identified, and these reflected the morphotype determination. Eight of these did not cluster with sequence of any described species implying that they were either species that had not yet been sequenced, or were new to science, though these will need to be isolated before they can be linked to appropriate species names. Results were consistent with previous reports, in which the majority of AMF sequence types detected from natural ecosystems showed few sequenced organisms among described and named fungi (Husband et al. 2002; Wubet et al. 2003; Rodríguez-Echeverría and Freitas 2006; Lumini et al. 2010). Secondly, there is high intraspecific variability (up to 24%) in some AMF species with the degree of variability differing among genera (Stockinger et al. 2010; Krüger et al. 2012), and it is not yet clear to what extent this might be reflected in interspecific variation. The number of AMF sequences available in GenBank, for many species, is too small to allow interpretation of the inter- and intraspecific genetic variation of AMF (Wang et al. 2011). For this study, DNA was extracted mainly from spores produced in soil-trap cultures.
Regardless of applied sequence similarity threshold (97, 98, 99%) only 16 out of 18 phylotypes could be restored, when using the AMF database published in Krüger et al. (2012) as backbone. This shows that species delimitation in the Glomeromycota is challenging if not impossible with a single sequence similarity threshold, due to their intraspecific sequence variability (Stockinger et al. 2009, 2010), even if the whole SSU-ITS-LSU rDNA fragment (ca. 1.5 kb) is used for OTU analysis. Nevertheless, molecular characterisation is useful, especially if no conclusive morphological evidence is available (e.g. soil or root analyses without examination of spores), but a robust sequence database from well characterised vouchered fungal specimens is necessary for reliable taxonomical assignment (Stockinger et al. 2010; Krüger et al. 2012, 2015; Schlaeppi et al. 2016).
AMF phylotype richness varied between islands and between sites within each island. Our findings are consistent with our previous work in Juniperus brevifolia forests between Terceira and São Miguel (Melo et al. 2017), suggesting that geographical and historical (i. e., dispersion and speciation) influences are fundamental in determining local diversity of regions that share similar environmental conditions (van der Gast et al. 2011; Bainard et al. 2014). Similar results were also shown in other oceanic islands. Gruenstaeudl et al. (2013) observed that AMF richness and composition associated to Tolpis spp. not only differed between three Canarian islands but also between habitats within each islands. Nam et al. (2015) showed differences in composition of soil fungal communities between Ulleungdo and Dokdo islands.
Understanding AMF distributional patterns is important due to their relevant role in ecosystem functioning, and because of the current environmental threats to AMF diversity (Turrini and Giovannetti 2012). Geological history and age play an important role in patterns of biological species richness and endemism in the Azores, and should be taken into account when evaluating local and regional patterns of diversity (Borges and Brown 1999; Borges and Hortal 2009). Evidence from arthropods suggest that recent islands (e.g. Pico) have a lower diversity of native species than older islands, due to a more homogeneous landscape. However, sampling sites from the older island (São Miguel) did not assemble a higher AMF richness than the sampling sites from the younger island (Terceira). Geological age alone cannot be used to explain the glomeromycotan richness patterns in P. azorica forests between Terceira and São Miguel. This result is consistent with diversity of arthropods in native forest of Azores (Laurisilva) which is likely to be influenced, not only by physical factors of the fragment of native forest such as the isolation, geological age and area of the islands themselves, but also by the fragmentation and shrinkage that have shaped the geography directly over the last 550 years (Gaspar et al. 2008).
The composition and dominant taxa of the AMF community in the P. azorica rhizosphere differed between Terceira and São Miguel. The correspondence analysis provided evidence that the AMF communities had become grouped according to the different sites – e. g., the communities at TB tended to cluster with those at TS, whereas those from ST and SG clustered together. Similar differences have been shown to be related to soil characteristics, such as moisture content (Wolfe et al. 2007; Shukla et al. 2013), structure (Lekberg et al. 2007; AL-Ghamdi and Jais 2012), fertility (Egerton-Warburton et al. 2007; Alguacil et al. 2008; Schreiner and Mihara 2009), and disturbance (Alguacil et al. 2008; Schnoor et al. 2011; Trejo et al. 2016), and such factors can influence composition and distribution of the AMF community (Schneider et al. 2015). Levels of organic matter, and total N, were significantly higher in native forest of Terceira than in those of São Miguel, whereas pH and available P were lower (Melo et al. 2017). Some AMF respond differently to soil disturbances, for example Hart and Reader (2002) found that species from ‘Glomaceae’ (actually, these included glomoid-spored groups now in two different families, Glomeraceae and Claroideoglomeraceae) were much less resilient to soil disturbances than species from Gigasporaceae, possibly because the former are said mainly to colonised roots by hyphae and the latter by spores. De la Providencia et al. (2015) also showed that species of Diversispora, Claroideoglomus and Rhizophagus, (glomoid-spored species, one from the Diversisporales and two from the Glomerales) dominated an extreme petroleum hydrocarbon-polluted location, and they suggested that their greater capacity to tolerate the presence of oil-related organic pollutants could be explained by their life history strategies.
The native forests of the Azores archipelago had been mostly destroyed since human occupation and converted into agricultural land (Martins 1993; Borges et al. 2008; Triantis et al. 2010) and in São Miguel, most indigenous forest was already seriously fragmented through agriculture and plantation of Cryptomeria japonica. Some evidence has shown how human-introduced invasive species can influence AMF communities (Moora et al. 2014; Shah et al. 2015). Mummey and Rillig (2006) found lower AMF taxon diversity among invasive species when compared with native vegetation. Also Violi et al. (2008) showed that conversion of mature (indigenous) forest to pine, and thus to predominance of an ectomycorrhizal host, clearly reduced AMF spore density. Conversion of temperate forest to avocado plantations and maize fields in central México had considerable repercussions on AMF composition and richness (González-Cortés et al. 2012). These studies show that widespread non-native plant invasions can alter AMF communities. If action is not taken to control invasive plant species, distributions of indigenous AMF species are very likely to be disrupted.
10 Conclusion
This is the first molecular study of AMF associated with P. azorica in native forest of the Azores. Our results suggest that, rather than plant identity, factors such as geographical region and conversion of native forest are probably instrumental in shaping AMF communities in Laurisilva ecosystems. Among the glomeromycotan spores, sequences of eight morphotypes did not cluster with sequence of any described species, thus possibly representing undescribed fungi. Establishment of the fungi in culture is needed to discover if these newly identified fungal types possess useful functional traits, but so far isolation attempts have failed. Work is proceeding to try to isolate these organisms, and to assess their potential utility for Laurisilva ecosystem restoration in the Azores.
References
AL-Ghamdi AA, Jais HM (2012) Interaction between soil textural components, flavonoids in the roots and mycorrhizal colonization in ‘Juniperus procera in Saudi Arabia. Afr J Microbiol Res 7:996–1001
Alguacil MM, Lumini E, Roldan A et al (2008) The impact of tillage practices on arbuscular mycorrhizal fungal diversity in subtropical crops. Ecol Appl 18:527–536
Bainard LD, Bainard JD, Hamel C, Gan Y (2014) Spatial and temporal structuring of arbuscular mycorrhizal communities is differentially influenced by abiotic factors and host crop in a semi-arid prairie agroecosystem. FEMS Microbiol Ecol 88:333–344. doi:10.1111/1574-6941.12300
Berger SA, Krompass D, Stamatakis A (2011) Performance, accuracy, and web server for evolutionary placement of short sequence reads under maximum likelihood. Syst Biol 60:291–302. doi:10.1093/sysbio/syr010
Borges PAV, Brown VK (1999) Effect of island geological age on the arthropod species richness of Azorean pastures. Biol J Linn Soc 66:373–410
Borges PAV, Hortal J (2009) Time, area and isolation: factors driving the diversification of Azorean arthropods. J Biogeogr 36:178–191
Borges PAV, Ugland KI, Dinis FO, Gaspar C (2008) Insect and spider rarity in an oceanic island (Terceira, Azores): true rare and pseudo-rare species. In: Fattorini S (ed) Insect ecology and conservation. Research Signpost, Kerala (India) pp 47–70
Boyer LR, Brain P, Xu X-M, Jeffries P (2015) Inoculation of drought-stressed strawberry with a mixed inoculum of two arbuscular mycorrhizal fungi: effects on population dynamics of fungal species in roots and consequential plant tolerance to water deficiency. Mycorrhiza 25:215–227. doi:10.1007/s00572-014-0603-6
Brundrett MC, Ashwath N (2013) Glomeromycotan mycorrhizal fungi from tropical Australia III. Measuring diversity in natural and disturbed habitats. Plant Soil 370:419–433. doi:10.1007/s11104-013-1613-4
Caporaso JG, Bittinger K, Bushman FD et al (2010a) PyNAST: a flexible tool for aligning sequences to a template alignment. Bioinformatics 26:266–267. doi:10.1093/bioinformatics/btp636
Caporaso JG, Kuczynski J, Stombaugh J et al (2010b) QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7:335–336. doi:10.1038/nmeth.f.303
Cardoso P, Gaspar C, Pereira LC et al (2008) Assessing spider species richness and composition in Mediterranean cork oak forests. Acta Oecol 33:114–127. doi:10.1016/j.actao.2007.10.003
Clapp JP, Rodriguez AR, Dodd JC (2002) Glomales rRNA gene diversity – all that glistens is not necessarily glomalean? Mycorrhiza 12:260–270
de la Providencia IE, Stefani FOP, Labridy M, et al (2015) Arbuscular mycorrhizal fungal diversity associated with Eleocharis obtusa and Panicum capillare growing in an extreme petroleum hydrocarbon-polluted sedimentation basin. FEMS Microbiol Lett 362:fnv081. doi: 10.1093/femsle/fnv081
de Oliveira AN, de Oliveira LA (2010) Influence of edapho-climatic factors on the sporulation and colonization of arbuscular mycorrhizal fungi in two Amazonian native fruit species. Braz Arch Biol Technol 53:653–661. doi:10.1590/S1516-89132010000300021
Dias E (1996) Ecologia e sintaxonomia das florestas naturais. Ph.D. Dissertation, University of the Azores
dos Santos R, Girardi CG, Pescador R, Stürmer SL (2010) Effects of arbuscular mycorrhizal fungi and phosphorus fertilization on post vitro growth of micropropagated Zingiber officinale Roscoe. Rev Bras Ciênc Solo 34:765–771. doi:10.1590/S0100-06832010000300018
Edgar RC (2013) UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat Methods 10:996–998. doi:10.1038/nmeth.2604
Egerton-Warburton LM, Johnson NC, Allen EB (2007) Mycorrhizal community dynamics following nitrogen fertilization: a cross-site test in five grasslands. Ecol Monogr 77:527–544
Ferrol N, Calvente R, Cano C et al (2004) Analysing arbuscular mycorrhizal fungal diversity in shrub-associated resource islands from a desertification-threatened semiarid Mediterranean ecosystem. Appl Soil Ecol 25:123–133. doi:10.1016/j.apsoil.2003.08.006
Gaspar C, Borges PA, Gaston KJ (2008) Diversity and distribution of arthropods in native forests of the Azores archipelago. Arquip Life Mar Sci 25:01–30
Gilmore AE (1968) Phycomycetous mycorrhizal organisms collected by open-pot culture methods. Hilgardia 39:87–105
González-Cortés JC, Vega-Fraga M, Varela-Fregoso L et al (2012) Arbuscular mycorrhizal fungal (AMF) communities and land use change: the conversion of temperate forests to avocado plantations and maize fields in central Mexico. Fungal Ecol 5:16–23. doi:10.1016/j.funeco.2011.09.002
Gruenstaeudl M, Santos-guerra A, Hawkes CV, Jansen RK (2013) Molecular survey of arbuscular mycorrhizal fungi associated with Tolpis on three Canarian islands (Asteraceae). Vieraea 41:199–218
Hart MM, Reader RJ (2002) Host plant benefit from association with arbuscular mycorrhizal fungi: variation due to differences in size of mycelium. Biol Fertil Soils 36:357–366. doi:10.1007/s00374-002-0539-4
Helgason T, Merryweather JW, Denison J et al (2002) Selectivity and functional diversity in arbuscular mycorrhizas of co-occurring fungi and plants from a temperate deciduous woodland. J Ecol 90:371–384
Helgason T, Merryweather JW, Young JPW, Fitter AH (2007) Specificity and resilience in the arbuscular mycorrhizal fungi of a natural woodland community. J Ecol 95:623–630. doi:10.1111/j.1365-2745.2007.01239.x
Henderson PA, Seaby RMH (2002) Species diversity and richness, version 3.0. Pisces Conservation Ltd., www.pisces-conservation.com. (http://www.pisces-conservation.com). Accessed 27 Aug 2016
Hildebrandt U, Regvar M, Bothe H (2007) Arbuscular mycorrhiza and heavy metal tolerance. Phytochemistry 68:139–146. doi:10.1016/j.phytochem.2006.09.023
Husband R, Herre EA, Turner SL et al (2002) Molecular diversity of arbuscular mycorrhizal fungi and patterns of host association over time and space in a tropical forest. Mol Ecol 11:2669–2678
Ismail Y, Hijri M (2012) Arbuscular mycorrhisation with Glomus irregulare induces expression of potato PR homologues genes in response to infection by Fusarium sambucinum. Funct Plant Biol 39:236. doi:10.1071/FP11218
Jansa J, Mozafar A, Anken T et al (2002) Diversity and structure of AMF communities as affected by tillage in a temperate soil. Mycorrhiza 12:225–234. doi:10.1007/s00572-002-0163-z
Krüger M, Stockinger H, Krüger C, Schüßler A (2009) DNA-based species level detection of Glomeromycota: one PCR primer set for all arbuscular mycorrhizal fungi. New Phytol 183:212–223. doi:10.1111/j.1469-8137.2009.02835.x
Krüger M, Krüger C, Walker C et al (2012) Phylogenetic reference data for systematics and phylotaxonomy of arbuscular mycorrhizal fungi from phylum to species level. New Phytol 193:970–984. doi:10.1111/j.1469-8137.2011.03962.x
Krüger M, Teste FP, Laliberté E et al (2015) The rise and fall of arbuscular mycorrhizal fungal diversity during ecosystem retrogression. Mol Ecol 24:4912–4930. doi:10.1111/mec.13363
Leake J, Johnson D, Donnelly D et al (2004) Networks of power and influence: the role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning. Can J Bot 82:1016–1045. doi:10.1139/b04-060
Lekberg Y, Koide RT, Rohr JR et al (2007) Role of niche restrictions and dispersal in the composition of arbuscular mycorrhizal fungal communities. J Ecol 95:95–105. doi:10.1111/j.1365-2745.2006.01193.x
Lumini E, Orgiazzi A, Borriello R et al (2010) Disclosing arbuscular mycorrhizal fungal biodiversity in soil through a land-use gradient using a pyrosequencing approach. Environ Microbiol. doi:10.1111/j.1462-2920.2009.02099.x
Magurran AE (2004) Measuring biological diversity. Blackwell Science, Oxford
Martins AM (1993) The Azores-westernmost Europe: where evolution can be caught red-handed. Mus Mun Funchal 2:181–198
Martins J, Moreira O, Silva L, Moura M (2011) Vegetative propagation of the endangered Azorean tree, Picconia azorica. Arquip Life Mar Sci28:39–46
Melo CD, Walker C, Rodríguez-Echeverría S et al (2014) Species composition of arbuscular mycorrhizal fungi differ in semi-natural and intensively managed pastures in an isolated oceanic island (Terceira, Azores). Symbiosis 64:73–85. doi:10.1007/s13199-014-0303-1
Melo CD, Luna S, Krüger C et al (2017) Arbuscular mycorrhizal fungal community composition associated with Juniperus brevifolia in native Azorean forest. Acta Oecol. doi:10.1016/j.actao.2016.12.006
Mendonça D, Luna S, Bettencourt S et al (2015) In vitro propagation of Picconia azorica (Tutin) Knobl. (Oleaceae) an Azorean endangered endemic plant species. Acta Physiol Plant 37:47. doi:10.1007/s11738-015-1797-8
Middleton EL, Richardson S, Koziol L, et al (2015) Locally adapted arbuscular mycorrhizal fungi improve vigor and resistance to herbivory of native prairie plant species. Ecosphere 6:art276. doi: 10.1890/ES15-00152.1
Minitab I (2000) Minitab: release 13 for Windows. Minitab Inc., State College
Moora M, Davison J, Öpik M et al (2014) Anthropogenic land use shapes the composition and phylogenetic structure of soil arbuscular mycorrhizal fungal communities. FEMS Microbiol Ecol 90:609–621. doi:10.1111/1574-6941.12420
Mummey DL, Rillig MC (2006) The invasive plant species Centaurea maculosa alters arbuscular mycorrhizal fungal communities in the field. Plant Soil 288:81–90. doi:10.1007/s11104-006-9091-6
Nam Y-J, Kim H, Lee J-H et al (2015) Metagenomic analysis of soil fungal communities on Ulleungdo and Dokdo Islands. J Gen Appl Microbiol 61:67–74. doi:10.2323/jgam.61.67
Oehl F, Laczko E, Bogenrieder A et al (2010) Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol Biochem 42:724–738. doi:10.1016/j.soilbio.2010.01.006
Öpik M, Metsis M, Daniell TJ et al (2009) Large-scale parallel 454 sequencing reveals host ecological group specificity of arbuscular mycorrhizal fungi in a boreonemoral forest. New Phytol 184:424–437. doi:10.1111/j.1469-8137.2009.02920.x
Redecker D, Schüßler A, Stockinger H et al (2013) An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 23:515–531. doi:10.1007/s00572-013-0486-y
Rodríguez-Echeverría S, Freitas H (2006) Diversity of AMF associated with Ammophila arenaria ssp. arundinacea in Portuguese sand dunes. Mycorrhiza 16:543–552. doi:10.1007/s00572-006-0070-9
Schlaeppi K, Bender SF, Mascher F et al (2016) High-resolution community profiling of arbuscular mycorrhizal fungi. New Phytol 212:780–791. doi:10.1111/nph.14070
Schneider KD, Lynch DH, Dunfield K et al (2015) Farm system management affects community structure of arbuscular mycorrhizal fungi. Appl Soil Ecol 96:192–200. doi:10.1016/j.apsoil.2015.07.015
Schnoor TK, Lekberg Y, Rosendahl S, Olsson PA (2011) Mechanical soil disturbance as a determinant of arbuscular mycorrhizal fungal communities in semi-natural grassland. Mycorrhiza 21:211–220. doi:10.1007/s00572-010-0325-3
Schoch CL, Seifert KA, Huhndorf S et al (2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proc Natl Acad Sci 109:6241–6246
Schreiner RP, Mihara KL (2009) The diversity of arbuscular mycorrhizal fungi amplified from grapevine roots (Vitis vinifera L.) in Oregon vineyards is seasonally stable and influenced by soil and vine age. Mycologia 101:599–611. doi:10.3852/08-169
Schüßler A, Schwarzott D, Walker C (2001) A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol Res 105:1413–1421
Shah MA, Beaulieu M-E, Reshi ZA et al (2015) A cross-city molecular biogeographic investigation of arbuscular mycorrhizas in Conyza canadensis rhizosphere across native and non-native regions. Ecol Process. doi:10.1186/s13717-015-0034-0
Shukla A, Kumar A, Jha A et al (2013) Soil moisture levels affect mycorrhization during early stages of development of agroforestry plants. Biol Fertil Soils 49:545–554. doi:10.1007/s00374-012-0744-8
Silva L, Martins M, Maciel G, Moura M (2009) Flora Vascular dos Açores - Prioridades em Conservação. Azorean Vascular Flora - Priorities in Conservation. Bilingue. Associação Ecológica Amigos dos Açores & CCPA, Ponta Delgada
Smith ES, Read DJ (2008) Mycorrhizal symbiosis. Academic, New York
Stamatakis A (2014) RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30:1312–1313. doi:10.1093/bioinformatics/btu033
Stockinger H, Walker C, Schüßler A (2009) “ Glomus intraradices DAOM197198”, a model fungus in arbuscular mycorrhiza research, is not Glomus intraradices. New Phytol 183:1176–1187. doi:10.1111/j.1469-8137.2009.02874.x
Stockinger H, Krüger M, Schüßler A (2010) DNA barcoding of arbuscular mycorrhizal fungi. New Phytol 187:461–474. doi:10.1111/j.1469-8137.2010.03262.x
ter Braak CJF, Smilauer P (2002) CANOCO reference manual and CanoDraw for Windows user’s guide: software for canonical ordination, version 4.5. Microcomputer power, Ithaca, NewYork
Trejo D, Barois I, Sangabriel-Conde W (2016) Disturbance and land use effect on functional diversity of the arbuscular mycorrhizal fungi. Agrofor Syst 90:265–279. doi:10.1007/s10457-015-9852-4
Triantis KA, Borges PAV, Ladle RJ et al (2010) Extinction debt on oceanic islands. Ecography 33:285–294. doi:10.1111/j.1600-0587.2010.06203.x
Turrini A, Giovannetti M (2012) Arbuscular mycorrhizal fungi in national parks, nature reserves and protected areas worldwide: a strategic perspective for their in situ conservation. Mycorrhiza 22:81–97. doi:10.1007/s00572-011-0419-6
van der Gast CJ, Gosling P, Tiwari B, Bending GD (2011) Spatial scaling of arbuscular mycorrhizal fungal diversity is affected by farming practice. Environ Microbiol 13:241–249. doi:10.1111/j.1462-2920.2010.02326.x
van der Heijden MG, Klironomos JN, Ursic M et al (1998) Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396:912–927
Verbruggen E, Röling WFM, Gamper HA et al (2010) Positive effects of organic farming on below-ground mutualists: large-scale comparison of mycorrhizal fungal communities in agricultural soils. New Phytol 186:968–979. doi:10.1111/j.1469-8137.2010.03230.x
Větrovský T, Baldrian P (2013) Analysis of soil fungal communities by amplicon pyrosequencing: current approaches to data analysis and the introduction of the pipeline SEED. Biol Fertil Soils 49:1027–1037. doi:10.1007/s00374-013-0801-y
Violi HA, Barrientos-Priego AF, Wright SF et al (2008) Disturbance changes arbuscular mycorrhizal fungal phenology and soil glomalin concentrations but not fungal spore composition in montane rainforests in Veracruz and Chiapas, Mexico. For Ecol Manag 254:276–290. doi:10.1016/j.foreco.2007.08.016
Walker C (1992) Systematics and taxonomy. Agronomie 12:887–897
Wang Y, Huang Y, Qiu Q et al (2011) Flooding greatly affects the diversity of Arbuscular Mycorrhizal fungi communities in the roots of wetland plants. PLoS One 6:e24512. doi:10.1371/journal.pone.0024512
Wolfe BE, Mummey DL, Rillig MC, Klironomos JN (2007) Small-scale spatial heterogeneity of arbuscular mycorrhizal fungal abundance and community composition in a wetland plant community. Mycorrhiza 17:175–183. doi:10.1007/s00572-006-0089-y
Wu Q-S, Srivastava AK, Zou Y-N (2013) AMF-induced tolerance to drought stress in citrus: a review. Sci Hortic 164:77–87. doi:10.1016/j.scienta.2013.09.010
Wubet T, Weiß M, Kottke I et al (2003) Molecular diversity of arbuscular mycorrhizal fungi in Prunus africana, an endangered medicinal tree species in dry Afromontane forests of Ethiopia. New Phytol 161:517–528. doi:10.1046/j.1469-8137.2003.00924.x
Acknowledgements
Special thanks to Luís Vasco Nunes of Regional Institute of Agrarian Planning for providing the aerial photographs. Many thanks to Paulo Borges of cE3c- Azorean Biodiversity Group for providing the software packages Species Diversity and Richness IV for the paper, and also for the work reviewed here, and also Reinaldo Pimentel of Azorean Biodiversity Group (CITA-A) for providing the imaging software. We gratefully acknowledge financial support for this research from the Portuguese Fundação para a Ciência e a Tecnologia (PTDC/AGR-ALI/122152/2010, SFRH/BPD/78059/2011 and UID/GEO/04035/2013) and Fundo Regional para a Ciência e Tecnologia – Governo dos Açores (M317/F/010A/2009 and M312/F/041/2011). C. Krüger was supported by the long-term research development project RVO 67985939.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Figure S1
Maximum likelihood consensus and Evolutionary Placement Algorithm (EPA) phylogenetic trees calculated by RaxML. Grouped into A) backbone phylogenetic tree and B) EPA trees achieved by applying different sequence similarity threshold 97, 98 and 99% (from left to right) using UPARSE. AMF sequences found in Picconia azorica roots marked in blue. Scale bar shows number of substitution per site. (PDF 164 kb)
Figure S2
Full maximum likelihood phylogenetic tree, spanning a part of the SSU, the full ITS1-5.8S-ITS2 region and a large part of the LSU rRNA gene regions (ca. 1.5 kb long), including the AMF sequences found in Picconia azorica roots marked in blue. Scale bar shows number of substitution per site. (GIF 43 kb)
Table S1
List of all detected OTUs calculated with SEED. Corresponding sequences are linked to the different OTUs achieved by the sequence similarity thresholds 97, 98 and 99%, in separated tabs. (XLS 39 kb)
Rights and permissions
About this article

Cite this article
Melo, C.D., Luna, S., Krüger, C. et al. Communities of arbuscular mycorrhizal fungi under Picconia azorica in native forests of Azores. Symbiosis 74, 43–54 (2018). https://doi.org/10.1007/s13199-017-0487-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13199-017-0487-2