
Abstract
An endophytic Bacillus amyloliquefaciens subsp. subtilis was isolated from surface sterilized seedlings of tropical corn. To evaluate the hypothesis that this endophyte is defensive in nature and determine the mechanisms of host defense we examined the effects of the endophyte on pathogens and host disease resistance genes. The bacterial endophyte showed antifungal activity against fungal pathogens Fusarium moniliforme, Colletotrichum gloeosporioides and Aspergillus flavus. Plant protection activity was also observed in Indian popcorn seedlings inoculated with the endophyte against F. moniliforme. Gene expression analysis was conducted and demonstrated that up-regulation of several defense genes in corn was greater after 48 hours of pathogen challenge than 24 hours. Plant defense pathways triggered by the endophytic bacterium appeared to be SA (salicylic acid) independent. The abundance of the zm Jasmonic acid-induced gene expression was greater in pathogen challenged plants pre-treated with the endophytic bacterium than in the plants challenged only with pathogen. Pre-treatment with the endophytic bacterium resulted in a more intense induction of maize pathogenesis related protein genes PR-1 and PR-10 during interaction with F. moniliforme compared to plants that were treated only with the fungal pathogen.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Plants have multiple pathways that function in defense against pathogen attack (Uknes et al. 1992; Van Loon and Bakker 2006). The systemic acquired resistance (SAR) pathway may be triggered by induction of plant defense through inoculation with a necrotizing pathogen or application of chemical agents such as salicylic acid (SA), 2–6-dichloroisonicotinic acid (INA), and benzo (1,2,3) thiadiazole-7-carbothioic acid S-methyl ester (BTH) (Uknes et al. 1992). Salicylic acid is a signalling molecule involved in both locally- and systemically-induced disease resistances. SAR is characterized by an early increase of endogenously synthesized SA and enhanced production of pathogenesis related (PR) proteins, specifically PR-1, PR-2, and PR-5, not only at the site of primary infection but also systemically in the uninfected plant tissues (Malamy et al. 1990; Métraux et al. 1990; Uknes et al. 1992). The expression of PR proteins is reported to stimulate pathogens to avoid further infection. Plant growth regulators jasmonic acid and ethylene also play an important role in defense mechanisms. This jasmonic acid/ethylene (JA/ET) is independent of the SA pathway. The exogenous application of these signalling molecules (JA/ET) induces synthesis of antimicrobial proteins such as defensins and thionins (Epple et al.1997). The signalling of JA and ET is also involved in induction of the PR proteins like PR-3, a basic chitinase; PR-4, a chitin-binding protein; and PDF1.2, a member of plant defensins (van Loon and van Stein 1999).
Some non-pathogenic plant-associated bacteria like Pseudomonas spp. and Bacillus spp. induce plant defense pathways that render the host more resistant to further pathogen attack (Ongena and Thonart 2006; Van Loon and Bakker 2006). This pathway of host defense is termed induced systemic resistance (ISR). It is reported that plant growth promoting B. mycoides strain Bac J and B. pumilus strains 203–6 and 203–7 provide ISR in sugar beet by enhanced peroxidase activity and increased production of one chitinase isozyme and two isozymes of β-1,3-glucanase (Bargabus et al. 2002; Bargabus et al. 2004). A non-pathogenic rhizobacterium Pantoea agglomerans isolated from Teosinte (maize ancestor) induces salt tolerance by triggering expression of salt resistance genes in modern maize (Gond et al. 2015a).
Endophytic bacteria colonize internal tissues of plants establishing symbiotic interactions with hosts (White et al. 2014; Hardoim et al. 2015). These microbes may promote plant growth by secreting anti-pest compounds, producing plant growth hormones, increasing nutrient supply, etc. (Ryan et al. 2008). Endophytic actinobacteria isolated from wheat tissues up-regulate defense genes like PR-1 and PR-4 of SAR as well as PDF1.2 and Hel genes of the jasmonic acid/ethylene (JA/ET) pathway in Arabidopsis thaliana (Conn et al. 2008). In a recent study, endophytic Bacillus spp. were reported to produce antifungal lipopeptides and induce host defense gene expression like PR-1 and PR-4 in maize (Gond et al. 2015b). That study demonstrated that the endophytic bacterium which produces antifungal lipopeptides in vitro can also trigger expression of pathogenesis related genes of the host without interaction with pathogens. The aim of the current study is to evaluate the effect of the endophytic bacterium Bacillus amyloliquefaciens subsp. subtilis on salicylic acid dependent and salicylic acid independent pathways of Indian popcorn, a flint maize variety.
2 Materials and methods
2.1 Isolation of endophytic bacteria
To obtain endophytic bacteria from the seedlings of tropical corn, seeds were soaked in 4 % NaOCl for 20 min and then washed three times with sterile distilled water and placed on sterile filter paper soaked with water in Petri dish for 5 days. The root and shoot parts of growing seedlings were dissected and again surface sterilized with 4 % NaOCl for 2 min and washed three times with sterile distilled water and placed on yeast extract (1 %) sucrose (1 %) agar (YESA) plates amended with antifungal cyclohexamide. The water after the last wash of the seedlings was also plated on separate culture plates to check the efficacy of surface sterilization. Plates were incubated at room temperature (~21 °C) for growth of bacteria. Based on appearance in culture and sequence analysis, a single bacterial species was obtained from seedlings. The bacterium was subcultured and purified using streak plating for identification. All isolates were stored at −80 °C in the Department of Plant Biology and Pathology, Rutgers University.
2.2 Identification of the endophytic bacterium
The bacterium was identified by 16 s rDNA sequencing from cultures. Total genomic DNA was extracted by the DNA extraction kit (Qiagen, Valencia, California, USA) and the 16 s rDNA sequence was amplified by primers 27f (AGAGTTTGATCMTGGCTCAG) and 1525r (AAGGAGGTGWTCCARCC) in a Biometra thermocycler (Biometra, Gottingen, Germany). The amplification program employed an initial denaturation at 95 °C for 5 min and then 30 cycles for 1 min at 95 °C, 1 min at 55 °C and 1.30 min at 72 °C. Final extension was at 72 °C for 10 min. The PCR product was purified by a PCR purification kit (Qiagen, Valencia, California, USA) and sent to Genewiz Inc. (South Plainfield, New Jersey, USA) for sequencing. The sequence was BLAST searched on the NCBI GenBank website to identify the closest matches.
2.3 Functional characterization of endophytic bacterium
2.3.1 Growth on N-free medium
The bacterium was streaked onto three plates of Norris Nitrogen-Free Agar and kept at room temperature for 48 h. For the control the bacterium was streaked onto three plates containing YESA.
2.3.2 Phosphate solubilisation
Phosphate solubilization capacity of the endophytic bacterium was determined by the plate assay method using Pikovskaya agar (Pikovskaya 1948). The bacterium was inoculated onto the centre of three Pikovskaya agar plates and incubated for one week. The clearing zones around the colonies were the positive indication of phosphate solubilization.
2.3.3 Auxin production
The bacterium was grown in LB broth with tryptophan (100 μg/ml) and inoculated at room temperature for two days at 100 rpm shaking. LB broth with tryptophan (100 μg/ml) was used as control. After incubation, cultures were centrifuged at 8000 rpm for 15 min at 4 °C. Two millilitres of freshly prepared Salkowski reagent (1 ml of 0.4 M FeCl3 in 50 ml of 35 % sulphuric acid) was added to 4 ml of culture supernatant. The reaction mixture was incubated at room temperature for 25 min. Development of pink color indicated the production of IAA.
2.3.4 Chitinase production
The chitinase assay was done using colloidal chitin as a substrate in three plates of chitinase assay medium. Colloidal chitin was prepared according to the method of Berger and Reynolds (1988). The composition of chitinase assay medium was Na2HPO4(6gL−1); KH2PO4 (3 g L−1); NH4Cl (1 g L−1); NaCl (0.5 g L−1); yeast extract (0.05 g L−1); agar (15 g L−1) and colloidal chitin (1%w/v). Colonies showing clearing zones were considered to be chitinase-producers.
2.3.5 Antifungal activity
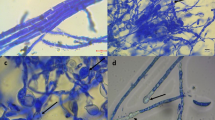
The dual culture assay was done for antifungal activity of the bacterium. The fungal pathogens Fusarium moniliforme, Colletotrichum gloeosporioides and Aspergillus flavuswere collected from stored samples in the Department of Plant Biology and Pathology, Rutgers University. The selection of these fungi was based on their pathogenicity against maize. A small disc of fungus was placed at the edge of each of three plates of potato dextrose agar (PDA), and after two days the bacterium on a 7 mm diam. Agar disc was placed opposite the growing fungal colony (Fig. 1). Inhibitory activity was determined after one week at lab ambient temperature (~21 °C) by assessing presence of radial growth inhibition. The radial growth inhibition was measured by subtracting the radial growth of mycelia towards bacterium side from radial growth of mycelia opposite to bacterium side from the centre of fungal disc. The data are averages of three replicates.

Inhibitory activity of Bacillus amyloliquefaciens subsp. subtilis against fungal pathogens
2.4 Corn seedling treatment with the endophytic bacterium and fungal pathogen
Roots of six 5-day-old Indian popcorn seedlings were dipped in 5-ml of bacterial suspension (~106 cells/ml) for 2.5 h and incubated for 24 h at room temperature under aseptic conditions. The controls were treated with sterile saline water. After 24 h, 1-ml of spore suspension of F. moniliforme (~104 spores/ml) was sprayed onto each seedling separately. Plant defense gene expression study was done after 24 and 48 h of F. moniliforme inoculations respectively.
2.5 RT-qPCR for expression of defense genes in corn seedlings by the endophytic bacterium
Induction of plant defense genes by the endophytic bacterium during interaction with the fungal pathogen was examined using relative quantification (RQ) in qPCR. Total RNA was extracted from 100 mg of fresh tissues after 24 and 48 h of the F. moniliforme inoculation of seedlings that were either pre-treated with the endophytic bacterium or water. The ‘only water’ treatment was used as a control. RNA was extracted by RNeasy Plant Minikit (Qiagen, Valencia, California, USA) according to the manufacturer’s protocol. The concentration and purity of total RNA was measured by a NanoDrop® ND1000 spectrophotometer. Real-Time One-Step RT-PCR was performed on Step One Plus Cycler (Applied Biosystems) by QuantiTect SYBR Green RT-PCR kit (Qiagen, Valencia, California, USA) according to the manufacturer’s protocol. The reverse transcription of RNA was set at 50 °C for 30 min. The PCR steps were initial activation of Taq polymerase at 95 °C for 15 min and cycling 94 °C for 15 s, annealing 60 °C for 30 s, extension 72 °C for 30 s for 40 cycles followed by melt curve analysis. The defense genes (zmPR1, zmPR10, zm salicylic acid induced gene and zm jasmonic acid induced gene) of the Indian popcorn were identified from the Maize genetics and genomics database (http://www.maizegdb.org/sequence.php). The selection of these genes was based on their expression against fungal pathogens and their role in SAR and ISR. The endogenous control gene was maize actin zmAct 1. The Primers for these genes were designed by ‘Primer Express' software (Table 1). The significance of the differences in gene expression was calculated by ‘Student’s T-test’.
2.6 Inoculation of bacterium and antifungal activity in Arabidopsis sp.
Seeds of a wild-type strain of Arabidopsis thaliana Col-0 were surface sterilized with 4 % NaOCl for 2 min and washed three times with sterile distilled water. The seeds were suspended in a one-day-old bacterial suspension for 2 h. The control seeds were treated with only the bacterial growth medium (LB broth). Seedlings were grown in sterile soil in magenta boxes under a 12-h alternating light/dark cycle (florescent lighting) at laboratory ambient temperature (~21 °C) and watered periodically using sterilized demineralized water to maintain soil moisture during the course of the experiment. After one week, seedlings were sprayed with a spore suspension of F. moniliforme. Seedlings were grown two additional weeks prior to comparing treatments.
3 Results
3.1 The endophytic bacterium and its functional characteristics
The endophytic bacterium isolated from tropical corn seedlings was rod-shaped, Gram-positive and identified as Bacillus amyloliquefaciens subsp. subtilis based on its 16 s rDNA sequence homology to strains from NCBI GenBank. The bacterium was able to grow on a nitrogen-free medium, indicating nitrogen fixing capacity. In dual culture assay, the bacterium was able to inhibit the growth of all filamentous fungi tested (Fig. 1, Table 2) while chitinase activity was not found on colloidal chitin agar (Table 2). Secretion of auxin was found in this bacterium while phosphate solubilisation capacity was not indicated on Pikovskaya agar.
3.2 Effect of the endophytic bacterium on the Indian popcorn defense against fungal pathogen
The Indian popcorn seedlings treated with B. amyloliquefaciens subsp. subtilis or water did not show any disease symptoms after 3 days of observations. The growth of the cottony mycelium of F. moniliforme was found on the seedlings pre-treated with only water (Fig. 2). The seedlings pre-treated with B. amyloliquefaciens subsp. subtilis appeared healthy after 48 h of the F. moniliforme treatment (Fig. 2).

Indian popcorn seedlings after 48 h of fungus inoculation (W-water; B-Bacillus amyloliquefaciens subsp. subtilis; F-Fusarium moniliforme; B + F-Bacterium and fungus); Arrow indicates fungal growth
3.3 Induction of defense gene expression in Indian popcorn by endophytic bacterium
The pattern of gene expression was determined after 24 and 48 h of pathogen treatment by transcript analysis using qRT-PCR. Among four genes studied, only zmPR-10 was expressed in seedlings either treated with bacterium or pathogen after 24 h of treatment with greater expression in pathogen-treated seedlings pre-treated with the bacterium (Fig. 3). After 48 h from pathogen inoculation, the expression of zmPR-1, zmPR-10 and zm Jasmonic Acid induced genes were significantly greater in bacterium pre-treated plants than ‘only pathogen’ treated plants (Fig. 4). The expression of zmPR-1 was down-regulated in pathogen challenged plants and highly up-regulated in bacterium treated plants. The results show that the bacterium did not have any effect on the expression of the zm Salicylic Acid induced gene. The ‘only fungal pathogen’ treatment induced expression of the zm Salicylic Acid gene compared to the water treated control plants. The zm Jasmonic Acid induced gene was down regulated in ‘only bacterium’ treated plants compared to the water control plants but highly up-regulated in pathogen-treated plants pre-treated with the bacterium (Fig. 4).

Induction of defense gene expression in Indian popcorn seedlings by Bacillus amyloliquefaciens subsp. subtilis after 24 h of F. moniliforme inoculation showing up-regulation of zmPR-10 gene in fungus treated plants pre-treated with bacterium (RQ-Relative quantification)

Induction of defense gene expression in Indian popcorn seedlings by Bacillus amyloliquefaciens subsp. subtilis after 48 h of F. moniliforme inoculation showing up-regulation of zmPR-1, zmPR-10 and JA induced genes in fungus treated plants pre-treated with bacterium (RQ-Relative quantification)
3.4 Effect of endophytic bacterium on Arabidopsis defense against fungal pathogen
The Arabidopsis seedlings treated with B. amyloliquefaciens subsp. subtilis were free from fungal attack. The cottony mycelia of F. moniliforme were growing on the soil surface of both bacteria-treated and untreated Arabidopsis seedlings but the bacterium-treated seedlings were healthy and flowering (Fig. 5). All untreated control seedlings were infected with the fungal pathogen and exhibited necrosis symptoms.

Effect of B. amyloliquefaciens subsp. subtilis on pathogenesis of F. moniliforme on Arabidopsis seedlings (a- Control; b-Treated with B. amyloliquefaciens subsp. subtilis)
4 Discussion
Endophytic bacteria may play roles in various plant growth promotion activities including plant protection against phytopathogens. The infection of plants by endophytic bacteria may occur in the rhizosphere or the phyllosphere (Rosenblueth and Martinez Romero, 2004). Some endophytic bacteria transmit through seeds of the host and form systemic infections that may colonize all parts of plants (Coombs and Franco 2003; Hallmann et al. 1997). The endophytic bacterium isolated in this study from tropical corn seedlings was identified as Bacillus amyloliquefaciens subsp. subtilis (Table 2). The most common genera reported as endophytic bacteria from maize varieties are Bacillus spp., Burkholderia spp., Herbaspirillum spp., Enterobacter spp., Klebsiella spp., and actinobacteria Arthrobacter spp. and Microbacterium spp. (Johnston-Monje and Raizada 2011; Zinniel et al. 2002; Chelius and Triplett 2000; McInroy and Kloepper 1995; Gond et al. 2015b). Bacillus amyloliquefaciens occurs as a common plant growth promoting rhizobacterium (PGPR) as well as an endophyte which secretes various kinds of antifungal molecules, including lipopeptides (White et al. 2014; Gond et al. 2015b). The strain of B. amyloliquefaciens subsp. subtilis isolated in this study showed strong activity against fungal pathogens and was able to grow on nitrogen-free media, and further was shown to reduce acetylene in culture (data not shown) indicating nitrogen fixing capacity (Table 2). This suggests that B. amyloliquefaciens subsp. subtilis promotes plant growth not only by secreting anti-pest compounds but also may increase a host’s nitrogen supply (Szilagyi-Zecchin et al. 2014). This bacterium showed strong antifungal activity, but did not show chitinase secretion, suggesting that antifungal activity may be due to lipopeptides or similar inhibitors (Gond et al. 2015b). Phosphate solubilisation was not seen in this bacterium, perhaps because it grows within host tissues; populations of phosphate solubilising bacteria are frequently in greater abundance in rhizospheric soil than other habitats (Katznelson et al. 1962; Raghu and MacRae 1966).
We observed antifungal activity of B. amyloliquefaciens subsp. subtilis in dual culture assays; and further we demonstrated that the bacterium could protect plants of Indian popcorn from fungal pathogens (Fig. 2). The growth of fungal mycelia was not observed on bacterium treated seedlings. Similarly, an endophytic Pseudomonas sp. strain PsJN gave enhanced protection against Verticillium sp. in tomato (Sharma and Nowak 1998) and Botrytis cinerea in grapevine (Barka et al. 2000, 2002). The early response of plants in microbe-plant interactions is frequently release of signalling molecules including salicylic acid (SA) and jasmonic acid (JA). These signal molecules are involved in induction of expression of defense genes (Mur et al. 2006; Vlot et al. 2009). The expression of signalling genes either for SA or JA was not detected after 24 h of pathogen challenge. It appears that F. moniliforme resistance generation in seedlings is SA independent because expression of the SA gene was down-regulated in ‘bacterium treated’ and ‘bacterium and fungus treated’ seedlings (Fig. 4). In contrast to this the abundance of the JA gene transcript was highest after 48 h of the pathogen challenged seedlings that were pre-treated with B. amyloliquefaciens subsp. subtilis (Fig. 4). In a similar study, endophytic Streptomyces sp. strain EN27mediated resistance in Arabidopsis seedlings against the fungal pathogen F. oxysporum via a NPR1-dependent pathway but also required SA and was JA/ET independent (Conn et al. 2008). The expression of endogenous JA is involved in a variety of critical functions, including fruit ripening, senescence, tuber formation, tendril coiling, pollen formation, and defense responses against pests and pathogens (Creelman and Mullet 1997). The over-expression of pathogenesis related genes zmPR-1 and zmPR-10 was also observed in pathogen challenged seedlings pre-treated with B. amyloliquefaciens subsp. subtilis (Fig. 4). Actinobacteria have been shown to up-regulate both SAR and JA/ET pathway defense genes, but defense responses by P. fluorescens WC417r or LSW17S and Bacillus spp. were found to be SA independent and were rather found to be associated with JA and ET (Kloepper et al. 2004; Pieterse et al. 1996 and 1998; Conn et al. 2008). The induction of ISR in Arabidopsis thaliana by root-colonizing strains of Pseudomonas fluorescens was shown to be ethylene- and JA-dependent but SA-independent (Knoester et al. 1999; Ton et al. 2002; Iavicoli et al. 2003).
In this study after 24 h of pathogen challenge, only the zmPR-10 gene was induced in seedlings pre-treated with B. amyloliquefaciens subsp. subtilis (Fig. 3). The expression of zmPR-10 was also greater in pathogen challenged seedlings pre-treated with bacterium compared to only pathogen challenged seedlings (Fig. 4). This suggests a role of the zmPR-10 protein in protection of corn seedlings against the fungal pathogen F. moniliforme. The expression of zmPR-10 protein is also induced during kernel development of maize and protects against Aspergillus flavus (Chen et al. 2006). The expression of PR-10 protein has also been reported in rice plants to protect against infection by the pathogen Magnaporthe grisea and in cowpea to protect against infection by the rust Uromyces vignae (McGee et al. 2001; Mould et al. 2003). The up-regulation of the zmPR-1 gene was observed after 48 h in ‘pathogen challenged seedlings pre-treated with bacterium’ and seedlings ‘treated with bacterium’ while down-regulated in seedlings only challenged with the fungal pathogen (Fig. 4). The up-regulation of the zmPR-1 gene was also reported in maize seedlings treated with endophytic B. subtilis by Gond et al. (2015b).
Plant protection activity of B. amyloliquefaciens subsp. subtilis was also seen in the model plant Arabidopsis thaliana Col-0 against F. moniliforme (Fig. 5). The endophytic bacterium was able to protect A. thaliana Col-0 plants, while the fungal pathogen was seen to grow on the soil surface only (Fig. 5). In an earlier study an endophytic strain of B. subtilis isolated from tropical maize was found to suppress the growth of seed mycoflora of Arabidopsis thaliana (Gond et al. 2015b). Endophytic Bacillus species not only act against fungal pathogens but also protect plants against bacterial diseases. A strain of Bacillus subtilis (Lu144) was found to reduce incidence of bacterial wilt of mulberry (Ji et al. 2008).
5 Conclusions
Our results demonstrate the role of endophytic microorganisms in enhancing plant growth and development. B. amyloliquefaciens subsp. subtilis induces Indian popcorn plants to synthesize defense molecules against fungal pathogen F. moniliforme. The defense pathway triggered by B. amyloliquefaciens subsp. subtilis in Indian popcorn seedlings was jasmonic acid dependent. The expression of pathogenesis-related genes like PR-1 and PR-10 was highly induced by B. amyloliquefaciens subsp. subtilis in response to the pathogen. The capacity of B. amyloliquefaciens subsp. subtilis to protect plants from disease was also demonstrated in Arabidopsis thaliana Col-0 against the fungal pathogen F. moniliforme.
References
Bargabus RL, Zidack NK, Sherwood JW, Jacobsen BJ (2002) Characterization of systemic resistance in sugar beet elicited by a non-pathogenic, phyllosphere-colonizing Bacillus mycoides, biological control agent. Physiol Mol Plant Pathol 61:289–298
Bargabus RL, Zidack NK, Sherwood JW, Jacobsen BJ (2004) Screening for the identification of potential biological control agents that induce systemic acquired resistance in sugar beet. Biol Control 30:342–350
Barka EA, Belarbi A, Hachet C, Nowak J, Audran JC (2000) Enhancement of in vitro growth and resistance to grey mould of Vitis vinifera co-cultured with plant-promoting rhizobacteria. FEMS Microbiol Lett 186:91–95
Barka EA, Gognies S, Nowak J, Audran JC, Belarbi A (2002) Inhibitory effect of endophyte bacteria on Botrytis cinerea and its influence to promote the grapevine growth. Biol Control 24:135–142
Berger LR, Reynolds DM (1988) Colloidal chitin preparation. Methods Enzymol 161:140–142
Chelius MK, Triplett EW (2000) Immunolocalization of dinitrogenase reductase produced by Klebsiella pneumoniae in association with Zea mays. L Appl Environ Microbiol 66:783–787
Chen ZY, Brown RL, Rajasekaran K, Damann KE, Cleveland TE (2006) Identification of a maize kernel pathogenesis related protein and evidence for its involvement in resistance to Aspergillus flavus infection and aflatoxin production. Phytopathology 96:87–95
Conn V, Walker A, Franco CM (2008) Endophytic Actinobacteria induce defense pathways in Arabidopsis thaliana. Mol Plant-Microbe Interact 21(2):208–218
Coombs JT, Franco CMM (2003) Visualization of an endophytic Streptomyces species in wheat seed. Appl Environ Microbiol 69:4260–4262
Creelman RA, Mullet JE (1997) Biosynthesis and action of jasmonates in plants. Annu Rev Plant Physiol Plant Mol Biol 48:355–381
Epple P, Apel K, Bohlmann H (1997) Overexpression of an endogenous thionin enhances resistance of Arabidopsis against Fusarium oxysporum. Plant Cell 9:509–520
Gond SK, Bergena MS, Torresa MS, Helsel Z, White JF (2015a) Induction of salt tolerance and up-regulation of aquaporin genes in tropical corn by rhizobacterium Pantoea agglomerans. Lett Appl Microbiol 60:392–399
Gond SK, Bergena MS, Torresa MS, White JF (2015b) Endophytic Bacillus spp produce antifungal lipopeptides and induce host defense gene expression in maize. Microbiol Res 17:79–87
Hallmann J, Quadt-Hallmann A, Mahaffee WF, Kloepper JW (1997) Bacterial endophytes in agricultural crops. Can J Microbiol 43:895–914
Hardoim PR, van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A, Döring M, Sessitsch A (2015) The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol Rev 79:293–320
Iavicoli A, Boutet E, Buchala A, Metraux JP (2003) Induced systemic resistance in Arabidopsis thaliana in response to root inoculation with Pseudomonas fluorescens CHA0. Mol Plant-Microbe Interact 16:851–858
Ji X, Lu G, Gai Y, Zheng C, Mu Z (2008) Biological control against bacterial wilt and colonization of mulberry by an endophytic Bacillus subtilis strain. FEMS Microbiol Ecol 65:565–573
Johnston-Monje D, Raizada MN (2011) Conservation and diversity of seed associated endophyesin Zea across boundaries of evolution, ethnography and ecology. PLoS One 6(6):e20396
Katznelson H, Peterson EA, Rovatt JW (1962) Phosphate dissolving microoganisms on seed and in the root zone of plants. Can J Bot 40:1181–1186
Kloepper JW, Ryu CM, Zhang S (2004) Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 94:1259–1266
Knoester M, Pieterse CM, Bol JF, van Loon LC (1999) Systemic resistance in Arabidopsis induced by rhizobacteria requires ethylene dependent signalling at the site of application. Mol Plant-Microbe Interact 12:720–727
Malamy J, Carr JP, Klessig DF, Raskin I (1990) Salicylic acid: a likely endogenous signal in the resistance response of tobacco to viral infection. Science 250:1002–1004
McGee JD, Hamer JE, Hodges TK (2001) Characterization of a PR-10 pathogenesis-related gene family induced in rice during infection with Magnaporthe grisea. Mol Plant-Microbe Interact 14:877–886
McInroy JA, Kloepper JW (1995) Survey of indigenous bacterial endophytes from cotton and sweet corn. Plant Soil 173:337–342
Métraux JP, Signer H, Ryals J, Ward E, Wyss-Benz M, Gaudin J, Raschdorf K, Schmid E, Blum W, Inverardi B (1990) Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science 250:1004–1006
Mould MJ, Xu T, Barbara M, Iscove NN, Heath MC (2003) CDNAs generated from individual epidermal cells reveal that differential gene expression predicting subsequent resistance or susceptibility to rust fungal infection occurs prior to the fungus entering the cell lumen. Mol Plant-Microbe Interact 16:835–845
Mur LA, Kenton P, Atzorn R, Miersch O, Wasternack C (2006) The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol 140:249–262
Pieterse CM, van Wees SC, Hoffland E, van Pelt JA, van Loon LC (1996) Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell 8:1225–1237
Pieterse CM, van Wees SC, van Pelt JA, Knoester M, Laan R, Gerrits H, Weisbeek PJ, van Loon LC (1998) A novel signalling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell 10:1571–1580
Raghu K, MacRae IC (1966) Occurrence of phosphate-dissolving microorganisms in the rhizosphere of rice plants and in submerged soils. J Appl Bacteriol 29:582–586
Rosenblueth M, Martinez Romero E (2004) Rhizobium etli maize populations and their competitiveness for root colonization. Arch Microbiol 181:337–344
Ryan RP, Germaine KJ, Franks A, Ryan DJ, Dowling DN (2008) Bacterial endophytes: recent developments and applications. FEMS Microbiol Lett 278:1–9
Sharma VK, Nowak J (1998) Enhancement of verticillium wilt resistance in tomato transplants by in vitro co-culture of seedlings with a plant growth-promoting rhizobacterium (Pseudomonas sp. Strain PsJN). Can J Microbiol 44:528–536
Szilagyi-Zecchin VJ, Ikeda AC, Hungria M, Adamoski D, Kava-Cordeiro V, Glienke C, Galli-Terasawa LV (2014) Identification and characterization of endophytic bacteria from corn (Zea maysL) roots with biotechnological potential in agriculture. AMB Express 4:26. doi:10.1186/s13568-014-0026-y
Ton J, Van Pelt JA, Van Loon LC, Pieterse CM (2002) Differential effectiveness of salicylate-dependent and jasmonate/ethylenedependent induced resistance in Arabidopsis. Mol Plant-Microbe Interact 15:27–34
Uknes S, Mauch-Mani B, Moyer M, Potter S, Williams S, Dincher S, Chandler D, Slusarenko A, Ward E, Ryals J (1992) Acquired resistance in Arabidopsis. Plant Cell 4:645–656
van Loon LC, van Stein EA (1999) The families of pathogenesis related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol Mol Plant Pathol 55:85–97
Vlot AC, Dempsey DA, Klessig DF (2009) Salicylic acid, a multifaceted hormone to combat disease. Annu Rev Phytopathol 47:177–206
White JF, Torres MS, Sullivan RF, Jabbour RE, Chen Q, Tadych M, Irizarry I, Bergen MS, Havkin-Frenkel D, Belanger FC (2014) Microscopy research and technique: occurrence of Bacillus amyloliquefaciens as a systemic endophyte of vanilla orchids. Microsc Res Tech 77(11):874–885
Zinniel DK, Lambrecht P, Harris NB, Feng Z, Kuczmarski D, Higley P, Ishimaru CA, Arunakumari A, Barletta RG, Vidaver AK (2002) Isolation and characterization of endophytic colonizing bacteria from agronomic crops and prairie plants. Appl Environ Microbiol 68:2198–2208
Acknowledgments
We are thankful to Department of Plant Biology and Pathology, Rutgers University for providing necessary facilities. SKG acknowledges UGC, India for providing Raman Post Doctoral fellowship No.F 5-31/2013(lC) for the year (2013-14) to visit USA and to administration, Visva-Bharati for providing leave and necessary facilities. RNK expresses his thankfulness to Department of Botany, BHU and DST (SB/EMEQ121/2014 dated 21/07/2014) for facilities and financial support. Authors would like to thanks Dr. Michael D. Pierce for assistance on RT-qPCR. We are grateful for financial assistance from: the New Jersey Agricultural Experiment Station, USDA NIFA Multistate Project W3147, the John and Christina Craighead Foundation, and the Rutgers Center for Turfgrass Science.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Gond, S.K., Bergen, M.S., Torres, M.S. et al. Effect of bacterial endophyte on expression of defense genes in Indian popcorn against Fusarium moniliforme . Symbiosis 66, 133–140 (2015). https://doi.org/10.1007/s13199-015-0348-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13199-015-0348-9