
Abstract
Control of biogenic amines (BAs) is important to guarantee the safety of sausage-like fermented meat products. This study investigated the influences of tea polyphenols (TP) and its lipophilic palmitic acid-modified derivatives, palmitoyl-TP (pTP) and palmitoyl-epigallocatechin gallate (pEGCG), on BAs and microbial ecosystem in Chinese sausages. TP, epigallocatechin gallate (EGCG), pTP, and pEGCG all reduced the formation of BAs and N-nitrosodimethylamine at 0.05% (g/g); yet, compared with TP and EGCG, the modified derivatives exhibited stronger action on BAs decreasing (P < 0.05), and pEGCG showed the highest effect (a reduction of total BAs from 376.22 to 168.98 mg/kg compared to control). The improved inhibitory effect of pTP and pEGCG should be attributed to their stronger dual-directional regulation of the bacterial and fungal communities during the natural fermentation of sausage. The modified pTP and pEGCG highly suppressed the growth of Staphylococcus, Candida, and Kurtzmaniella, all of which were positively correlated with BAs formation (all P < 0.05). However, pTP and pEGCG worked more effectively than the unmodified ones to promote Lactobacillus, Lactococcus, and Debaryomyces (all P < 0.05). The results above are significant for the application of palmitoyl-TP and similar TP derivatives in meat products in consideration of food safety.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
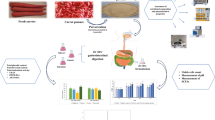
Chinese sausages are popular meat products consumed as side dishes (Zhang et al. 2017). They are commonly naturally fermented under cool conditions without start culture (Sun et al. 2018; Zhang et al. 2017).Lactic acid bacteria, coagulase-negative Staphylococcaceae, and yeasts are the main groups of microorganisms that have an important role in the sensory quality of sausages (Perea-Sanz et al. 2020; Flores and Piornos 2021; Wang et al. 2018). However, some microorganisms (e.g., Enterobacteriaceae) and their metabolites may produce biogenic amines (BAs) and nitrosamines (Halagarda and Wójciak 2022), which pose a health risk for consumers. BAs are usually formed by amino acids through decarboxylation (Biji et al. 2016; Liu et al. 2021; Sivamaruthi et al. 2021) and are commonly found in various fermented meat and fish products (Biji et al. 2016; Sivamaruthi et al. 2021). Excessive levels of BAs in food, including Chinese sausages (Li et al. 2019), are toxic and may lead to changes in food quality; these BAs are hazardous to the nervous and cardiovascular system of humans (Biji et al. 2016). In addition, some BAs are also widely known as import precursors of carcinogenic nitrosamines (Biji et al. 2016). So, regulating the microbial ecosystem and consequently inhibiting BAs accumulation in fermented meat products like Chinese sausages is very important.
Tea polyphenols (TP) are extensively used as antioxidants in the food and medical industry owing to their numerous beneficial health effects, such as scavenging free radicals, as well as anti-inflammatory and anti-carcinogenic properties (Khan and Mukhtar 2018; Deng et al. 2019; Hirasawa and Takada 2004). However, they are hydrophilic and sparingly soluble in lipid medium, which may compromise their effective application in lipophilic meat matrix (Paonessa et al. 2018). In recent years, fatty acid-modified TPs have markedly attracted scholars’ attention due to their excellent antioxidation and liposolubility in lipid medium because they could enter into cells more easily (Paonessa et al. 2018; Chu et al. 2019). Palmitoyl-TP (pTP), with a dominant constituent of palmitoyl-EGCG (pEGCG), is one of the main fatty acid-modified TP recommended as a source of antioxidants by the China Food and Drug Administration. Previous studies on pTP and pEGCG have mainly concentrated on their antioxidant and antiviral properties (de Oliveira et al. 2013; Ying and Fereidoon 2012), but their effect on BAs accumulation by regulating the microbial community structure in sausage fermentation was rarely reported. In this study, we investigated the influences of lipophilic pTP and pEGCG on forming BAs and nitrosamines in Chinese sausages compared to the unmodified ones. Also, the mechanism was discussed by exploring the microbial ecosystem. This study might be significant for controlling BAs and nitrosamines by applying lipophilic TP derivatives in the production of fermented meats.
Materials and methods
Materials and chemicals
A mixture of 9 BAs standard in a hydrochloric acid solution containing histamine, phenylethylamine, tyramine, putrescine, cadaverine, tryptamine, spermine, spermidine, octopamine was purchased from Century Aoke Biotechnology Co., Ltd (Beijing, China). TP (99%) was from Macklin (Shanghai, China). EGCG (98%), pTP (> 70%), and pEGCG (> 90%, with mono-palmitoyl-EGCG > 55%) were provided by Changxing Sanju Biotechnology Co., Ltd. (Hangzhou, Zhejiang, China).
Preparation of Chinese sausages
Samples (sausages) were prepared in triplicate according to the process of a previous study (Sun et al. 2018) with slight modifications. Ingredients of the base mixture without starter culture were as follows: back fat 200 g/kg, lean pork 800 g/kg, NaCl 2.5%, NaNO2 0.015%, glucose 1.5%, sterile water 5%, and wine 1.5%. The base mixture was divided into five groups: a control group and four groups mixed with 0.05% (g/g) of TP, EGCG, pTP, and pEGCG powder, respectively. The five groups were cultured at 10 °C for 2 h and then stuffed in a porcine casing. The sausages were fermented at 25 °C and 90–95% RH for 2 days and then ripened at 10 °C and 65–70% RH for 4 weeks. Ripened sausage (500 g) was cut, mashed, and then sampled for analysis.
BAs determination
BAs extraction and derivatization were performed according to the common method (Zhang et al. 2017). 5 g of sausage were homogenized with 20 mL of perchloric acid (0.4 mol/L) and centrifuged (4 °C, 5000 × g, 10 min). One milliliter of the supernatants was reacted at 40 °C for 30 min with 300 μL saturated sodium bicarbonate, 200 μL sodium hydroxide (2 mol/L), and 1 mL dansyl chloride reagent (10 mg/mL). After that, the reaction was terminated with 100 μL ammonium hydroxide (25%). The mixture was adjusted to 5 mL with acetonitrile and filtered (0.22 μm) for analysis.
HPLC system (Agilent 1290 Intifinity) equipped with a C18 column (Eclipse XDB-C18, 4.6 mm × 250 mm, 5 μm) (Aglient Technologies, CA, USA) was used for analysis. The injection volume was 20 μL, and determinations were conducted at 254 nm and 35 °C. Mobile phases (0.8 mL/min) included solvent A [0.05 mol/L acetate buffer (0.1% acetic acid)-acetonitrile (10:90, v/v)], and solvent B [0.05 mol/L acetate buffer (0.1% acetic acid)-acetonitrile (90:10, v/v)]. Gradients were as follows: 0–22 min, 60–85% A; 22–25 min, 85–100% A; 25–32 min, 100% A; 32–37 min, 60% A.
Nitrosamines determination
Samples (5 g) were mixed with an internal standard (50 ng NDMA-d6), and nitrosamines were extracted by QuEChERS for further analysis (Qiu et al. 2017). Nitrosamines were measured using an Agilent 7010B GC/TQ system (Agilent, Delaware, USA) equipped with an HP-5 ms capillary column (30 m × 0.32 mm × 0.25 μm, Agilent, Santa Clara, USA). The injection volume was 2 μL, and helium (99.99%) was used as carrier gas at 1.2 mL/min. The temperature program was as follows: 80 °C, held for 3 min, 80–100 °C at 25 °C/min, 100–110 °C at 10 °C/min, 110–120 °C at 5 °C/min, 120–130 °C at 10 °C/min, 130–280 °C at 40 °C/min, finally held for 5 min. The MS/MS was implemented in multiple reaction monitoring modes with 70 eV electron energy and two or three mass transitions. The ion source temperature was set at 230 °C. Identification of the analytes was confirmed by the qualifier-to-target ion ratios and retention time of target ion, as shown in Supplemental Information (Table S1).
Moisture content, water activity, pH, and microbial counts
Moisture content and pH were analyzed according to AOAC procedures (AOAC 2006). Water activity (aw) was determined using a water activity meter (Rotronic, Bassersdorf, Switzerland). For microbial counts, samples (3 g) were homogenized in 300 mL of sterilized normal saline for 1 min, after which serial tenfold dilutions were prepared. The selective mediums used were as follows: PCA for total aerobic bacteria (TAB) (37 °C, 48 h), MRS for lactic acid bacteria (LAB) (37 °C, 48 h), VRBGA for Enterobacteriaceae at (37 °C, 24 h), and PDA for yeast and mold (37 °C, 24 h). The results were expressed as colony-forming units per gram (log CFU/g).
DNA extraction, PCR amplification, and sequencing
Approximately 500 mg of sausage samples (each in triplicated) were used for DNA extraction using an OMEGA E.Z.N.A soil DNA kit (Feiyang BIOTECH Co., Ltd, Guangzhou, China) according to the manufacturer’s instructions. DNA quality was monitored by 0.8% agarose gel electrophoresis. The DNA samples were stored at −80 °C for further analysis.
The V3-V4 region of the bacterial 16 S rRNA gene was amplified with primer pairs 338F (5’-ACTCCTACGGGAGGCAGCA-3’) and 806R (5’-GGACTACHVGGGTWTCTAAT-3’). The ITS region of the fungal ITS gene was amplified using primer ITS1F (5’-CTTGGTCATTTAGAGGAAGTAA-3’) and ITS2 (5’-GCTGCGTTCTTCATCGATGC-3’). The PCR amplification was performed according to a previously described method (Yang et al. 2016). Finally, amplicons were pooled in equimolar concentrations and sequenced by an Illumina MiSeq platform and MiSeq Reagent Kit v1 (Illumina, Inc., Santiago, CA, USA) at the Biomarker Technologies Corporation (Beijing, China).
The sequences data were merged, quality filtered, and optimized. High-quality sequences (length > 100 bp, without reads containing base ‘N’, the lowest overlap length < 10 bp) were adopted for further analysis. Chimeras were checked by QIIME Pipeline (Chimera checking sequences with QIIME-Homepage). Operational taxonomic units (OTUs) were determined using the MOTHUR software package v.1.31.2., with a 97% sequence identity threshold. 16S rRNA reads were assigned using 16S rRNA training set 9 in the RDP database with a local BLAST search. A local BLAST search was also used to assign ITS 1 reads to the NCBI GenBank database. The bioinformatics analysis was performed in BMK Cloud (www.biocloud.net). The Shannon (https://mothur.org/wiki/shannon/) and Simpson (https://mothur.org/wiki/simpson/) diversity indices were calculated for analyzing the species diversity, while Chao 1 (https://mothur.org/wiki/chao/) was for the richness. The beta diversity distance measurements, including weighted UniFrac and Bray–Curtis, were visualized via principal coordinate analysis (PCoA). Linear discriminant analysis (LDA) effect size (LEfSe) algorithm (LDA score > 4 and P < 0.05) was used to compare relative abundances of microbial taxa among the five groups. The redundancy analysis (RDA) was performed to visualize the relationship between microbial community and BAs.
The bacterial and fungal sequencing data are publicly available in the National Center for Biotechnology Information (NCBI) under accession numbers PRJNA801771 and PRJNA801773, respectively.
Statistical analysis
All values were expressed as mean ± standard deviation (n = 3). An analysis of variance (ANOVA) with Tukey’s multiple comparisons was performed to investigate the significance of differences by R (version 3.6.1, × 64). Differences were considered to be significant when P < 0.05.
Results and Discussion
BAs and nitrosamines
Six BAs, including putrescine, cadaverine, histamine, tyramine, spermidine, and spermine, were detected in all samples (Fig. 1). The total concentrations of BAs in the case of control, TP, EGCG, pTP, and pEGCG were 376.22, 291.06, 237.44, 210.12, and 168.98 mg/kg, respectively. Among the detected BAs, tyramine had the highest concentration, followed by spermine, cadaverine, putrescine, spermidine, and histamine, similar to the observation of Harbin dried sausages (Sun et al. 2018). The highest concentration of tyramine (164.97 mg/kg) appeared in the control, and it was within the reported range of dried sausages and fermented fish products (Dsd et al. 2021). Putrescine and cadaverine, used as indicators of food hygiene, are commonly found in sausages and aquatic products, and their contents in control were 29.54 mg/kg and 55.92 mg/kg, respectively. Spermine ranged from 57.29 to 102.61 mg/kg, and spermidine was 10.97–18.29 mg/kg. Histamine is usually considered as the most toxic BA; in this study, it ranged from 4.43 to 4.88 mg/kg, which was significantly lower than the limit of 50 mg/kg (Biji et al. 2016; FDA 2001).

Influence of TP, EGCG, pTP, and pEGCG on the contents of biogenic amines (BAs) in Chinese sausages. a Putrescine; b cadaverine; c histamine; d tyramine; e spermidine; f spermine. Usage of TP, EGCG, pTP, or pEGCG was 0.05% (g/g); different lowercase letters (a–e) indicate significant differences in the reduction of BAs contents (P < 0.05)
TP, EGCG, pTP, and pEGCG reduced the contents of BAs in Chinese sausages. The palmitic acid-modified pTP and pEGCG had a stronger inhibitory effect compared to TP and EGCG, while pEGCG showed the highest decrease of 55.08% total BAs at 0.05% of usage (P < 0.05) (Fig. 1). Tyramine was reduced by pTP and pEGCG to < 100 mg/kg as an acceptance dose (Biji et al. 2016; FDA 2001), and pEGCG had the most effective suppression on tyramine (reduced by 61.25% compared to control). Apart from that, pEGCG decreased (P < 0.05 vs. control) putrescine (40.52%), cadaverine (74.02%), histamine (3.89%), spermidine (40.02%), and spermine (44.17%). The observed effect of TP, EGCG, pTP, and pEGCG is possibly caused by their ability to inhibit decarboxylase activity and suppress BAs formation (Gyawali and Ibrahim 2014). In addition, the long palmitoyl hydrocarbon chain endows pTP and pEGCG with strong liposolubility (Pan et al. 2020). The stronger BAs suppression ability of pTP and pEGCG compared with TP and EGCG might originate from their improved liposolubility because they could disperse well in sausage and enter into cells more easily to perform activities (Paonessa et al. 2018; Chu et al. 2019).
A total of 9 nitrosamines were examined in the study, but only NDMA was detected in sausage samples (Fig. 2, Table S1, and Fig. S1). The highest content of NDMA was found in the control group (0.86 μg/kg) (Fig. 2), which was markedly below the limit (10 μg/kg) (FDA 2001). TP, EGCG, pTP, and pEGCG all significantly reduced the formation of NDMA (P < 0.05), confirming the suppressive effects of TP and EGCG on the formation of nitrosamines (Deng et al. 2019). EGCG reduced the content of NDMA by 66.81% (vs. control) and showed the most effective inhibition among the four antioxidants. The concentrations of NDMA in the sausages treated with pTP and pEGCG were reduced by 27.77% and 48.72% (vs. control), respectively. The performance of pTP and pEGCG was weaker than that of TP and EGCG in effecting the accumulation of NDMA (P < 0.05). It was known that polyphenols could block nitrosamine formation by scavenging nitrite (Deng et al. 2019). Modified with palmitic acid reduced the hydrophilicity of TP and EGCG, which may influence its reaction efficiency with sodium nitrite, thereby weakening their inhibitory effect on NDMA formation (Deng et al. 2019; Gyawali and Ibrahim 2014). Overall, palmitic acid-modified TP and EGCG derivatives remained active in inhibiting the formation of NDMA in Chinese sausages.

Influence of TP, EGCG, pTP, and pEGCG on the contents of NDMA in Chinese sausages. Usage of TP, EGCG, pTP, or pEGCG was 0.05% (g/g); different lowercase letters (a–e) indicate significant differences in the reduction of NDMA contents (P < 0.05)
Microbial counts, pH, moisture content, and water activity
The long period of ripening resulted in a moisture content of < 25% in the final products of sausages (Table 1). No significant differences in moisture content were found among groups (all P > 0.05). The values of aw in sausages were very close with relatively small differences and generally consistent with the trend in moisture contents. However, the pH values of Chinese sausages were influenced by the addition of antioxidants. The samples with both palmitic acid-modified TPs and unmodified ones had lower pH values than the control group (P < 0.05), which is consistent with Sun et al. (2018), suggesting that adding cinnamon can effectively decrease the pH of dry sausage.
No viable Enterobacteriaceae was found in all treatments, indicating a sanitary condition in experiments. The counts of total aerobic bacteria (TAB), lactic acid bacteria (LAB), and fungi (yeast and mold) in control were 7.92, 6.00, and 5.74 Log (CFU/g), respectively, which was in agreement with similar studies without starter culture (Sun et al. 2018). Application of TP, EGCG, pTP, and pEGCG regulated the community structure of bacteria and fungi during the natural fermentation of sausage. Overall, they inhibited the biomass of TAB but promoted the multiplication of LAB and fungi (P < 0.05), accompanied by a decrease in product pH (Table 1). Compared to TP and EGCG, pTP and pEGCG had a greater effect on the community structure (P < 0.05); pEGCG significantly regulated the bacteria and fungi counts in sausages (P < 0.05).
Tea polyphenols have broad-spectrum antibacterial properties (Khan and Mukhtar 2018). Therefore, esterification modification could endow these polyphenols with an improvement of such activity (Chu et al. 2019). In fact, they can also promote the proliferation of some probiotics, such as Lactobacillus sp. (Lee et al. 2006). This study achieved a dual-directional regulation by 0.05% (g/g) of TP (or its palmitic acid-modified derivatives) in sausage, including inhibition of TAB and promotion of LAB and fungi. Comparatively, the lipidic derivatives pTP and pEGCG had a stronger ability of such regulation than the unmodified ones when they were used in the natural sausage fermentation.
Bacterial community in sausages
High-throughput sequencing was further applied to evaluate sausage microbial composition and diversity. As presented in Table 2, sausages treated with EGCG, pTP, and pEGCG showed slightly lower richness (Chao 1) than that of control, but the diffrence was not significant (P > 0.05), while the diversity (Simpson and Shannon) of pTP-and pEGCG-treated sausages was higher than that of TP and EGCG groups (all P < 0.05). In Fig. 3a, a noticeable separation was observed among the five groups, and 80.81% of the total variance (PC1 = 63.42%, PC2 = 17.39%) was explained according to the PCoA analysis based on weight UniFrac distance metric. Palmitoyl-TP and pEGCG groups were separated farther from the control, as the PC1 explained 63.42% of the variance. This observation suggested that palmitic acid modification of TP and EGCG might enhance their effects on the bacteria community composition in Chinese sausage fermentation.

Effect of TP, EGCG, pTP, and pEGCG on the bacterial community composition of Chinese sausages at the genus level. a PCoA analysis based on weighted UniFrac distance metrics; b Relative abundance; c RDA analysis. Usage of TP, EGCG, pTP, or pEGCG was 0.05% (g/g)
Figure 3b illustrates the relative abundance of the top 10 genera in all sausages. The bacterial community composition in control was similar to a previous study of Chinese sausage (Wang et al. 2018). Staphylococcus and LAB (Lactococcus, Weissella, and Lactobacillus) were the dominant genera, accounting for 94.58–98.53% of all bacteria genera in the five groups. Application of 0.05% (g/g) of TP, EGCG, pTP, and pEGCG in sausage significantly (P < 0.05) suppressed Staphylococcus growth; meanwhile, they generally promoted the growth of LAB (e.g., Lactococcus and Lactobacillus). In comparison with TP and EGCG, the lipidic pTP and pEGCG were more effective in performing such suppression and promotion (P < 0.05). These observations were consistent with the results of microbial counts (Table 1).
Redundancy analysis (RDA) was performed to further investigate the relationship between microbial community and BAs, as well as NDMA. The correlations between two variables in RDA were determined by the angle between the corresponding vectors (Dai et al. 2021). The sharper the angle, the stronger the positive correlation when angles < 90°; the blunter the angle, the stronger the negative correlation when angles > 90° (Dai et al. 2021). As shown in Fig. 3c, Staphylococcus was positively correlated with NDMA and BAs cadaverine, putrescine, tyramine, spermine, and spermidine, as their angles were lower than 90°. In addition, LAB genera such as Lactobacillus and Lactococcus showed strong negative correlations with NDMA and the BAs as the angles were more than 90°. These observations suggested that both inhibition actions on Staphylococcus and promotion on Lactobacillus and Lactococcus were crucial for decreasing BAs and NDMA in sausage. Staphylococcus is usually found in fermented meat products, and it is important for products' proteolysis, color, and flavor quality (Flores and Piornos 2021; Halagarda and Wójciak 2022). However, Martuscelli et al. (2000) reported that some decarboxylase-positive strains of Staphylococcus sp., such as S. xylosus S167, generate BAs, including tyramine, spermidine, spermine, and tryptamine. In contrast, some LAB strains (e.g., Lactobacillus pentosus R3 and Lactobacillus plantarum) were applied to efficiently inhibit the formation of BAs and nitrosamines (Li et al. 2021; Shao et al. 2021). As Staphylococcus, Lactobacillus, and Lactococcus were the most different biomarkers in the control, pEGCG, and pTP groups, respectively, when LDA score > 4 (all P < 0.05) (Fig. S2), the groups of sausages treated with or without palmitic acid-modified TPs could be distinguished by the dominant genera. The overall results above further demonstrated that the suppression of Staphylococcus and promotion of Lactobacillus and Lactococcus by pTP and pEGCG possibly contributes to inhibiting the formation of BAs and NDMA in Chinese sausage.
Fungal community in sausages
Yeast has an important role in forming the flavor and aroma of sausage due to its lipolytic and proteolytic activity (Perea-Sanz et al. 2020). However, these biochemical reactions may potentially produce BAs. In this study, six fungal genera, including Debaryomyces, Kurtzmaniella, Candida, Penicillium, Meyerozyma, and Wickerhamomyces, were identified in all samples, five of which were yeasts (Fig. 4a). The predominant genus identified was Debaryomyces, accounting for 86.07–97.36% of the total genera. The presence of EGCG, pTP, and pEGCG in sausage significantly (P < 0.05) decreased the fungal diversity in association with the lower Simpson and Shannon values than that of the control group (Table 2). The PCoA of fungi (Fig. 4b) showed a weaker separation of the five groups than that of bacteria. Nevertheless, pTP and pEGCG were closely clustered and were noticeably separated from other groups, indicating that palmitic acid modification could remarkably change the effects of tea polyphenols on fungi. The effect of TP on the fungal community was weaker than those of EGCG, pTP, and pEGCG (P < 0.05). However, EGCG, pTP, and pEGCG significantly improved the growth of Debaryomyces while inhibiting the growth of Candida and Kurtzmaniella (P < 0.05) (Fig. 4a). The relative abundances of Candida and Kurtzmaniella were reduced by 69.04% and 98.51% from pEGCG compared to control, respectively. The inhibition on yeast genera was consistent with a previous study (Hirasawa and Takada 2004), where tea polyphenols showed suppression on some noxious yeasts (e.g., Candida. albicans).

Effect of TP, EGCG, pTP, and pEGCG on the fungal community composition of Chinese sausages at the genus level. a Relative abundance; b PCoA analysis based on Bray–Curtis distance metrics; c RDA analysis. Usage of TP, EGCG, pTP, or pEGCG was 0.05% (g/g)
Figure 4c shows that Debaryomyces were strongly negatively correlated with NDMA and BAs cadaverine, putrescine, tyramine, spermine, and spermidine, while Candida and Kurtzmaniella were positively correlated with them. As the dominant fungus in sausage, some strains of Debaryomyces (e.g., D. hansenii H525) were found to effectively degrade histamine, tyramine, and phenylethylamine with high peroxisomal amine oxidase activity (Bäumlisberger et al. 2015). These results indicate that, besides regulating the bacterial community, pTP and pEGCG might also inhibit the formation of BAs and NDMA in sausage by promoting Debaryomyces and suppressing Candida and Kurtzmaniella.
Conclusions
The palmitic acid-modified pTP and pEGCG were found to be more effective than TP and EGCG (P < 0.05) in reducing the formation of BAs in Chinese sausage due to their lipidic structure. This inhibitory effect should be attributed to a dual-direction regulation of promotion and suppression of the microbial community in Chinese sausage. The pTP and pEGCG could promote the growth of Debaryomyces, Lactobacillus, and Lactococcus; meanwhile, they could suppress the proliferation of Staphylococcus, Candida, and Kurtzmaniella. As a result, such dual-direction regulation on the microbial community could remarkably decrease the contents of BAs and NDMA in Chinese sausages. These results may give a reference for applying pTP, pEGCG, and similar fatty acid-modified polyphenols in fermented meat products to regulate the microbial community and inhibit the accumulation of BAs.
Data availability
The data used and/or analysed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable.
Abbreviations
- BAs:
-
Biogenic amines
- TP:
-
Tea polyphenols
- EGCG:
-
Epigallocatechin gallate
- pTP:
-
Palmitoyl-TP
- pEGCG:
-
Palmitoyl-EGCG
- NDMA:
-
N-Nitrosodimethylamine
- TAB:
-
Total aerobic bacteria
- LAB:
-
Lactic acid bacteria
- OTUs:
-
Operational taxonomic units
- NCBI:
-
National center for biotechnology information
- PCoA:
-
Principal coordinate analysis
- LDA:
-
Linear discriminant analysis
- LEfSe:
-
Linear discriminant analysis effect size algorithm
- RDA:
-
Redundancy analysis
References
AOAC (2006) Official methods of analysis, 14th edn. Association of official analytical chemist, Washington, DC
Bäumlisberger M, Moellecken U, König H, Claus H (2015) The potential of the yeast Debaryomyces hansenii H525 to degrade biogenic amines in food. Microorganisms 3:839–850. https://doi.org/10.3390/microorganisms3040839
Biji KB, Ravishankar CN, Venkateswarlu R, Mohan CO, Gopal TK (2016) Biogenic amines in seafood: a review. J Food Sci Technol 53:2210–2218. https://doi.org/10.1007/s13197-016-2224-x
Chu TC, Lee LH, Aponte T, Lopez S, Hsu S (2019) Sporicidal activity of novel formulations containing lipophilic epigallocatechin-3-gallate and natural ingredients. Microbiol Infect Dis 3:1–6. https://doi.org/10.33425/2639-9458.1070
Dai WL, Gu SQ, Xu MJ, Wang WJ, Yao HZ, Zhou XX, Ding YT (2021) The effect of tea polyphenols on biogenic amines and free amino acids in bighead carp (Aristichthys nobilis) fillets during frozen storage. LWT-Food Sci Technol 150:111933. https://doi.org/10.1016/j.lwt.2021.111933
de Oliveira A, Adams SD, Lee LH, Murray SR, Hsu SD, Hammond JR, Dickinson D, Chen P, Chu TC (2013) Inhibition of herpes simplex virus type 1 with the modified green tea polyphenol palmitoyl-epigallocatechin gallate. Food Chem Toxicol 52:207–215. https://doi.org/10.1016/j.fct.2012.11.006
Deng S, Jin J, He Q (2019) Inhibitory effect of epigallocatechin gallate, epigallocatechin, and gallic acid on the formation of N-nitrosodiethylamine in vitro. J Food Sci 84:2159–2164. https://doi.org/10.1111/1750-3841.14737
Dsd B, Lja D, Lm D, Ea C, Fd D, Bdm A (2021) Survey of multiple biogenic amines and correlation to microbiological quality and free amino acids in foods. Food Control 120:107497. https://doi.org/10.1016/j.foodcont.2020.107497
Flores M, Piornos JA (2021) Fermented meat sausages and the challenge of their plant-based alternatives: a comparative review on aroma-related aspects. Meat Sci 182:108636. https://doi.org/10.1016/j.meatsci.2021.108636
Food and Drug Administration (FDA) (2001) Fish and fishery products hazards and controls guide. department of health and human services, Food and Drug Administration, Center for Food Safety and Applied Nutrition, Washington DC
Gyawali R, Ibrahim SA (2014) Natural antimicrobial/antioxidant agents in meat and poultry products as well as fruits and vegetables: a review. Food Control 46:412–429. https://doi.org/10.1080/10408398.2016.1194256
Halagarda M, Wójciak KM (2022) Health and safety aspects of traditional european meat products. A Rev Meat Sci 184:108623. https://doi.org/10.1016/j.meatsci.2021.108623
Hirasawa M, Takada K (2004) Multiple effects of green tea catechin on the antifungal activity of antimycotics against Candida albicans. J Antimicrob Chemother 53:225–229. https://doi.org/10.1093/jac/dkh046
Khan N, Mukhtar H (2018) Tea polyphenols in promotion of human health. Nutrients 11(1):39. https://doi.org/10.3390/nu11010039
Lee HC, Jenner AM, Low CS, Lee YK (2006) Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res Microbiol 157:876–884. https://doi.org/10.1016/j.resmic.2006.07.004
Li L, Zou D, Ruan LY, Wen ZY, Chen SW, Xu L, Wei XT (2019) Evaluation of the biogenic amines and microbial contribution in traditional Chinese sausages. Front Microbiol 10:872. https://doi.org/10.3389/fmicb.2019.00872
Li S, Du X, Feng L, Mu G, Tuo Y (2021) The microbial community, biogenic amines content of soybean paste, and the degradation of biogenic amines by Lactobacillus plantarum HM24. Food Sci Nutr 9:6458–6470. https://doi.org/10.1002/fsn3.2528
Liu C, Zhu T, Song H, Niu C, Wang J, Zheng F, Li Q (2021) Evaluation and prediction of the biogenic amines in Chinese traditional broad bean paste. J Food Sci Technol 58:2734–2748. https://doi.org/10.1007/s13197-020-04781-z
Martuscelli M, Crudele MA, Gardini F, Suzzi G (2000) Biogenic amine formation and oxidation by Staphylococcus xylosus strains from artisanal fermented sausages. Lett Appl Microbiol 31:228–232. https://doi.org/10.1046/j.1365-2672.2000.00796.x
Pan LH, Wu XL, Luo SZ, He HY, Luo JP (2020) Effects of tea polyphenol ester with different fatty acid chain length on camellia oil-based oleogels preparation and its effects on cookies properties. J Food Sci 85:2461–2469. https://doi.org/10.1111/1750-3841.15341
Paonessa R, Nardi M, Gioia MLD, Olivito F, Procopio A (2018) Eco-friendly synthesis of lipophilic EGCG derivatives and antitumor and antioxidant evaluation. Nat Prod Commun 13:1117–1122. https://doi.org/10.1177/1934578X1801300905
Perea-Sanz L, López-Díez JJ, Belloch C, Flores M (2020) Counteracting the effect of reducing nitrate/nitrite levels on dry fermented sausage aroma by Debaryomyces hansenii inoculation. Meat Sci 164:108103. https://doi.org/10.1016/j.meatsci.2020.108103
Qiu Y, Chen JH, Yu W, Wang P, Rong M, Deng H (2017) Contamination of Chinese salted fish with volatile N-nitrosamines as determined by QuEChERS and gas chromatography-tandem mass spectrometry. Food Chem 232:763–769. https://doi.org/10.1016/j.foodchem.2017.04.055
Shao XF, Xu BC, Zhou H, Chen CG, Li PJ (2021) Insight into the mechanism of decreasing N-nitrosodimethylamine by Lactobacillus pentosus R3 in a model system. Food Control 121:107534. https://doi.org/10.1016/j.foodcont.2020.107534
Sivamaruthi BS, Kesika P, Chaiyasut C (2021) A narrative review on biogenic amines in fermented fish and meat products. J Food Sci Technol 58:1623–1639. https://doi.org/10.1007/s13197-020-04686-x
Sun Q, Zhao X, Chen H, Zhang C, Kong B (2018) Impact of spice extracts on the formation of biogenic amines and the physicochemical, microbiological and sensory quality of dry sausage. Food Control 92:190–200. https://doi.org/10.1016/j.foodcont.2018.05.002
Wang X, Zhang Y, Ren H, Zhan Y (2018) Comparison of bacterial diversity profiles and microbial safety assessment of salami, Chinese dry-cured sausage and Chinese smoked-cured sausage by high-throughput sequencing. LWT-Food Sci Technol 90:108–115. https://doi.org/10.1016/j.lwt.2017.12.011
Yang L, Yang HL, Tu ZC, Wang XL (2016) High-throughput sequencing of microbial community diversity and dynamics during douchi fermentation. PLoS ONE 11:e0168166. https://doi.org/10.1371/journal.pone.0168166
Ying Z, Fereidoon S (2012) Lipophilised epigallocatechin gallate (EGCG) derivatives and their antioxidant potential in food and biological systems. Food Chem 131:22–30. https://doi.org/10.1016/j.foodchem.2011.07.089
Zhang QQ, Jiang M, Rui X, Li W, Chen XH, Dong MS (2017) Effect of rose polyphenols on oxidation, biogenic amines and microbial diversity in naturally dry fermented sausages. Food Control 78:324–330. https://doi.org/10.1016/j.foodcont.2017.02.054
Acknowledgements
The authors gratefully acknowledge the financial support provided by The Key Food Programme of Sichuan Province (No. 20ZDYF1250).
Funding
The authors gratefully acknowledge the financial support provided by The Key Food Programme of Sichuan Province (No. 20ZDYF1250).
Author information
Authors and Affiliations
Contributions
QZ: Conceptualization, Investigation, Writing-original draft. MM: Investigation, Software. BBT: Visualization. QH: Supervision, Writing-Reviewing & Editing.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Consent to participate
All authors have seen and agreed with the contents of the manuscript.
Consent for publication
All authors are aware of its submission to JFST.
Ethical approval
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Zhou, Q., Mo, M., Tang, B. et al. A comparative study of tea polyphenols and its palmitic acid-modified derivatives: their effects on the microbial ecosystem and biogenic amines in Chinese sausage. J Food Sci Technol 60, 1772–1781 (2023). https://doi.org/10.1007/s13197-023-05717-z
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13197-023-05717-z