
Abstract
In sustainable agriculture, organic farming endorses multifaceted benefits for producing quality crops with biodiversity enrichment by reducing pesticide use and other harmful practices. Organic farming usually sustains species richness, controls pests, enhances pollination, and reduces soil impurities than conventional agriculture. Due to the recognition of such benefits, organic farming is encouraging globally in recent times. Consequently, in promoting organic crop yield, the presence of Coleopteran beetles in organic farms has considerably gained attention. Considering these, the present study concentrates on the faunal diversity and richness of Coleopteran beetles from five organic agricultural farms across central Laos during monsoon and post-monsoon seasons. Trapped beetles were taxonomically identified, grouped according to seasons and fields, and calculated their diversity indices. Identified Coleopteran individuals (n = 1038) belonging to 24 families showed higher diversity indices (Shannon, H′; Simpson, D; evenness, J′) during monsoon than dry seasons. Beetle occurrence frequencies were also recorded higher in monsoon than post-monsoon period. Representatives from Curculionidae, Staphylinidae, and Phalacridae families predominately prevailed across the organic fields in both seasons; however, no significant variations of beetle assemblages were noticed among the farmlands. The present study infers that sustainable organic farming supports the diversity of Coleopteran beetles in the agroecosystems as indicator taxon.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Organic agriculture endorses sustainability compared to traditional farming, though the multifunctional sustainability benefits of organic farms may interfere with the landscape context. Assessment of the landscape context of an organic farm affects its sustainability and also directly influences biodiversity, crop yields, and profitability (Smith et al. 2020). Although organic farms produce lower yields than conventional farms (Seufert et al. 2012), they help biodiversity enrichment (Tuck et al. 2014; Lichtenberg et al. 2017) and species richness (van Elsen 2000). Organic agriculture also ensures benefits in reducing pesticide use and other harmful practices associated with traditional agriculture (Reganold and Wachter 2016). Generally, conventional agricultural intensification decreases biodiversity and ecosystem services. This phenomenon has ubiquitously occurred to any given ecosystem from fields, farms, or landscapes and its surroundings independently or simultaneously (Culman et al. 2010). In contrast to conventional agriculture, organic farming ensures the qualitative production of crops with biodiversity enrichment and more sustainable agricultural output (Inclán et al. 2015). For example, the abundance of beetles may be higher in non-crop habitats than in crop fields, but species diversity will be more in crop fields than in non-crop habitats (Gaines and Gratton 2010). The richness and diversity of ground beetle communities might be similar during the transition to the organic period from conventional cropping; however, beetle communities become change during the established organic phase (Adhikari and Menalled 2020). One of the major causes for the change of beetle assemblage is the predatory disturbances in the farmland (Gaines and Gratton 2010), and thus, organic farming also promotes a higher richness of ground beetle predators (Snyder 2019). The higher species richness could be due to characteristics of the agriculture types making these environments more attractive to diverse insect species (Wickramasinghe et al. 2004; Montañez and Amarillo-Suárez 2014).
Lao PDR is the only landlocked country from South East Asia, compacting very rich in floral and faunal diversity. Traditional organic agricultural farming towards producing cereals, crops, and vegetables is encouraging in the country for sustainable agricultural practices (Yamada et al. 2004). The natural organic resources of Laos encompass vast shares of the economy, and consequently, the rich bioresources of the country provide indispensable benefits to the rural people of society. The government, therefore, promotes sustainable organic agriculture and biodiversity-centered resource sharing for nutritious food and income for people’s healthcare and well-being (Panyakul 2009).
On organic farms, the soil is often amended with composted plants and animal wastes that significantly increase the ground beetle community. Many of these diverse beetle communities act as detritus-feeding arthropod predators and shape the richness of alternative predatory species (Gaines and Gratton 2010; Snyder 2019). In the practice of sustainable agriculture, especially for organic farming, the Coleopteran beetles often support the community structure of the ecosystem by enhancing its diversity, richness, and evenness (Crowder et al. 2010; Mone et al. 2014). Further, biodiversity conservation in agricultural landscapes also provides more species richness and facilitates meta-population processes between habitat patches (Perfecto and Vandermeer 2010). Several studies have reported on organic farming promoting more significant levels of biodiversity that may be more robust to species loss compared to their conventional counterparts (Macfadyen et al. 2009). The Coleopteran beetles are considered the most commonly studied fauna for organic farming for their direct role in soil nutrient recycling and mineral turnover (Hole et al. 2005). It is well reported that Coleopteran beetles are the most widespread insect community for organic farming and endorse added ecosystem services by modifying natural habitats than any other insect fauna (Manning et al. 2016). In addition, they serve as significant biological control agents and, therefore, are considered to some extent as potential bioindicator fauna for agricultural and natural environments (Rodrigues et al. 2013).
Considering the efficacy of Coleopteran beetles in turning over soil micro-nutrients in organic farmlands, substantial knowledge on diversity manifestations and richness strengths of Coleopteran faunal assemblages is essential. Under this understanding, we are interested to know (a) the extent of Coleopteran beetle diversity succeeds in organic farmlands and (b) the variation of beetle diversity amidst monsoon and post-monsoon seasons. Therefore, this paper describes the critical aspects of Coleopteran beetle diversity and richness parameters across different organic agricultural farms from central Laos in two different seasons. Our findings will assist in enhancing the knowledge of biodiversity conservation for sustainable organic farming in such agricultural ecosystems, in general.
Material and methods
Study area
Coleopteran beetles were collected from five organic agricultural fields (study field, n = 5) located in and around Nabong, Laos (18°70′N and 102°47′E) during monsoon (September to November 2018) and post-monsoon seasons (December 2018 to February 2019). The average aerial distance between the two study plots was approximately 300 ± 20 m, and the mean elevations of the studied fields were approximately 170 ± 20 m above sea level. The area for each study field was approximately 74 ± 10 hectares (0.74 ± 0.1 km2). Therefore, from a wide area of Nabong, five organic agricultural plots were selected, each covering an area of approximately three-quarter km2, and the mean distance between the plots was approximately a quarter km. The average temperatures during monsoon and post-monsoon seasons were 28 ± 2 °C and 16 ± 2 °C, respectively. In monsoon, average rainfall during the study period (September to November) was recorded approximately 1600 ± 65 mm, where during the post-monsoon season (December to February), no rainfall was recorded. All beetle samples were collected from the center zones of the integrated crop plantations from each field, and no data were recorded from the peripheral areas of the crop lands. During sample collections from both monsoon and post-monsoon seasons, approximately one-third of the organic agricultural landscapes were covered with paddy plantations, where two-third of the landscape was covered with other crops such as vegetables and fruit orchards.
Trapping and insect collection
Beetles were caught by soda-bottle traps (Steininger et al. 2015) from the fields, followed by labeling and transportation to the laboratory for preservation and further taxonomic identification. The soda-bottle traps were made using transparent plastic water bottles (1.5 l) with one window and were suspended upside down for easy access to the beetles. Propylene glycol (100 ml) was placed inside the bottle, ensuring the beetles from not escaping the trap, and the chemical helps preserve insects for further studies. Three window traps were installed at each field, and each trap was kept at a distance of at least 5 m apart. Ethanol (95%) was used as an attractant for the beetles and was kept inside the traps in tubes. The traps were hanged on wooden poles at approximately 150 cm above the ground. Traps were kept in the field for 6 months (September 2018 to February 2019), and each of them was observed weekly to notice the beetle if trapped any. If a trap detected broken during the experiment, it was replaced immediately with an identical one. A set of 3 bottle traps was placed for a field, and counting of beetles for a trap was done for a month; thus, 15 countings (3 traps × 5 fields) were made per month. With this manipulation, monsoon period (September to November) data was analyzed on assembling 45 countings (15 countings per month × 3 months), and similarly, post-monsoon period (December to February) data was also analyzed on another 45 countings. Therefore, the present study is based on 90 incidents of collected beetle samples across the five studied fields for 6 months.
Trapped beetles were collected weekly in marked vials separately, followed by counting (for the month) and taxonomic identification. Data generated for a month to all fields and a field to all months were tabulated, splitting into two seasonal categories (monsoon and post-monsoon). All beetles were taxonomically identified up to family levels at the laboratory following the techniques set by Beattie and Oliver (1994). All identified samples were housed at the “insect repository” at the Plant Protection Unit, Faculty of Agriculture National University of Laos, Lao PDR.
Data analysis
Shannon’s diversity index (H′), Simpson’s diversity index (D), and evenness index (J) were calculated for the identified counted beetles among months for both monsoon and post-monsoon seasons following the methods described by Pielou (1966), Whittaker (1972), and Adhikari and Menalled (2020). Only the identified beetles were considered for calculations of diversity indices. During the calculation, data from all bottle traps were pooled together for a month from each field. Similarly, data were also pooled from all studied fields in the calculation of the distribution percent of Coleopteran families observed during monsoon and post-monsoon seasons. Data were analyzed and compared among months for calculation of the Bray–Curtis similarity index. The software, R statistic (ver. 3.6.0, R development Core Team 2019), and SPSS (ver. 25.0) were used for statistical data processing and graphical representation. Occurrence frequency (richness) of beetle families was measured by using the formula, occurrence frequency (%) = the number of individuals trapped for a family/total number of trapped isolates × 100 (Magurran 2004). One-way analysis of variance (ANOVA) was done to test the difference of beetle occurrence between seasons. Non-parametric t-tests were performed to analyze the seasonal variance of beetle occurrence frequencies across the fields. Data were log-transformed to meet the assumptions for ANOVA.
Results and discussion
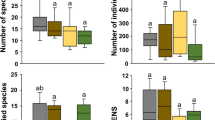
Individuals (n = 1038) belonging to order Coleoptera were collected consecutively for 6 months during monsoon (M) and post-monsoon (PM) seasons across different organic agricultural landscapes (study fields, n = 5) from central Laos. Collected Coleopterans were identified up to the family level, and a total of 24 families were taxonomically identified. Beetles from family Curculionidae were recorded most dominant (M, 33%; PM, 38%) followed by Staphylinidae (M, 21%; PM, 27%) and Phalacridae (M, 12%; PM, 14%), whereas beetles from families Scirtidae, Alleculidae, Carabidae, Mycetophagidae, Passalidae, Cleridae, Dytiscidae, and Helodidae were noticed to be the least abundant representatives in either season (0.17 to 0.22%). Percent distribution of the relative number of beetles for each family for monsoon and post-monsoon seasons is given in Table 1. For occurrence frequencies (%) of the beetles when compared among seasons, it was found that the occurrence trend of beetle families matched with the monsoon to post-monsoon periods (Fig. 1a), though there remained a significant difference of occurrences between the seasons (ANOVA, F1, 718 = 1.22, p < 0.5) (Fig. 1b).

Occurrence frequency (%) of Coleopteran beetle families (n = 24) during monsoon and post-monsoon seasons. Data are pooled from all studied fields (n = 5), (a) individual distribution percent, where a box is representing a single family, (b) inclusive distribution percent, where families are pooled together in a box. Data are shown in median values with interquartile range (box), total range (whiskers), and outliers (circles)
In characterizing the Coleopteran family diversity, our result indicates the Shannon diversity index (H′) reached the maximum onset of the rainy season (September to November) (1.95 to 1.98). After that, with the cessation of rain, H′ gradually decreased with dry-season advancement and reached the lowest during February (1.49). In measuring beetle diversity, the Simpson index (D) also indicated maximum during the monsoon period (0.80) but decreased gradually in the post-monsoon season. Value of family evenness index (J′) also followed a similar trend like H′ and D. Higher J′ was observed during monsoon (0.76) but turned down at the bottom (0.58) with the dry season. Diversity indices for the beetle assemblage in organic farming during monsoon and post-monsoon seasons are presented in Table 2, where the value of the Bray–Curtis similarity index of the beetle community across the seasons is indicated in Table 3. Similarity index trends among monsoon months (Sep to Nov) were paralleled to post-monsoon months (Dec to Feb).
The diversity distribution of Coleopteran beetles from the agricultural landscape presented in the present study agrees with the fact that organic farming has high connectivity with surrounding vegetation of the beetles’ community because plant coverings and species play an influential role in the beetle assemblage (Hutton and Giller 2003). As beetle assemblage and species composition deviate with the fluctuation of environmental conditions corresponding to the seasons, the pattern and functionality of beetle assemblage in organic farms work more potentially during monsoon than during the dry season (Kubo et al. 2013). According to Sawada and Harowatari (2002), Coleopteran diversity is influenced by water precipitations, and its assemblage prefers a more moist and humid environment. In another study, Burgio et al. (2015) also reported that rainfall has a significant role to play with the assemblage of beetle diversity in organic farming. Beetle occurrence to such habitat suggests their bioindicator potentiality for moist habitat. Therefore, as seasonal patterns of beetle communities differ among monsoon and post-monsoon seasons and within the organic farmlands, our result suggests the plasticity of beetle communities’ temporal dynamics in humid environmental tropics agreeing with the report from Wardhaugh et al. (2018).
The representing beetles from three families like Curculionidae, Staphylinidae, and Phalacridae were recorded highly dominant during monsoon and even in the post-monsoon period along with ten beetle families (viz., Chrysomelidae, Bostrichidae, Tenebrionidae, Histeridae, Anthribidae, Scarabaeidae, Cerambycidae, Laemophloeidae, Erotylidae, Nitidulidae) with moderate appearance. A cluster of another ten families (viz., Scirtidae, Pythidae, Alleculidae, Carabidae, Mycetophagidae, Passalidae, Cleridae, Dytiscidae, Helodidae, Coccinellidae) recorded rarely occurred across the study fields. Predominant occurrence (33–38%) of Curculionidae weevils (snout beetles) across the organic farmlands might be due to the presence of shrubs and fruit orchards around the study areas and possibly due to their wood-boring lifestyle. The appearance of snout weevils in such kinds of habitats was also reported earlier, demonstrating their bioindicator identity. As snout weevils constitute a diverse group of phytophagous insects, many of them are considered harmful pests for agricultural lands and orchard crops (Hurej et al. 2013; Tewari et al. 2014). On studying with bean and triticale, it was further reported that snout weevils might cause severe damage in diverse habitats, including significant damage to narrow-leafed lupin in organic farms (Hurej et al. 2013). Our recorded snout weevils were very tiny in size, possibly to be the bark and ambrosia beetles, and were ubiquitously present across the fields. Since Curculionid beetles are likely to consume a wide range of living plants, their assemblages were probably due to the mixed types of agricultural landscape comprised of natural open land with a large percentage of hardwood surrounding (Oliver and Mannion 2001).
Representatives from Staphylinidae (rove beetles) were common elements of the soil biota, living in the litter and deeper soil layers decay plant debris. They are considered as potential predators from soil-dwelling invertebrates among insects. Although they play a significant role in soil nutrient enhancing by decaying organic matter, their role in plant growth is poorly understood. Staphylinidae was recorded as the second most dominant (21–27%) Coleopterans throughout the monsoon and post-monsoon seasons in our study. One of the principal reasons for their omnipresence in the fields is that rove beetles are good predators and predate many invertebrates from agricultural lands. For example, Dalotia coriaria (Coleoptera: Staphylinidae) predates on Bradysia sp. nr Coprophilia (Diptera: Sciaridae) and occurs widely from almost any type of agricultural landscape (Echegaray et al. 2015). Our finding also corresponds with the fact that organic farming enhances the biodiversity of predators agreeing with the findings of Martins et al. (2013). In another study, Shah et al. (2003) also reported the diversity and abundance of predatory Coleopteran fauna in organic farm assemblages extensively. Ground and rove beetle diversity and abundance were remarkable among the predators (Shah et al. 2003), though their appearances generally depended on environmental factors (Dritschilo and Erwin 1982). The small bronzy or black shiny pollen beetles from family Phalacridae were recorded the third dominating Coleopterans (12–14%) across the studied farms. They visited the flowers of a wide range of ornamental plants and vegetables, fed pollen, and assisted pollination (Gimmel and Aston 2010). Our survey also reflects a good assemblage of pollen beetles (Phalacridae) across the organic farming landscapes.
Among the moderately dominant beneficial families, where occurrence frequencies varied from about 1 to 8%, the representing families more often occurred during monsoon than the dry season. Both the diversity and similarity indices also indicated a similar trend of beetle occurrence in the post-monsoon season. It can be argued that factors like trapping type, trap position, and trapping time of the year might affect catching difference of insects (Hanula et al. 2011; Peuhu et al. 2019), but maintaining all factors indifferent as acceptable as possible, we noticed that beetle variance was due to moisture content of the agroecosystem which was highly related to rainfall. Less than 5% of beetle families in the study sites could be designated as accidental species.
Despite study field and crop variation, we did not find any significant difference (NS) in the variation of occurrence frequencies of beetle families among the fields (one-way ANOVAmonsoon, F4, 355 = 5.88, NS; one-way ANOVApost-monsoon, F4, 355 = 4.24, NS), either in monsoon or post-monsoon periods (Fig. 2a). Therefore, our results do not say anything on the variation of beetle richness across the organic farms which suggest that the surveyed fields were nearly equivalent on faunal assemblage in terms of Coleopteran diversity and occurrences. Further, no significant difference in beetle families occurred in any organic field during monsoon and even in the post-monsoon period (non-parametric t-test; field 1: t = − 0.619, df = 139, p (2-tailed) = 0.537; field 2: t = − 0.507, df = 138, p (2-tailed) = 0.613; field 3: t = − 0.597, df = 137, p (2-tailed) = 0.551; field 4: t = − 0.529, df = 134, p (2-tailed) = 0.598; and field 5: t = − 0.510, df = 139, p (2-tailed) = 0.611) (Fig. 2b). This evidence might also be a sign of identical nature of organic fields.

Variation of Coleopteran beetle occurrence frequencies (%). Data presented at log10 scale, (a) non-significant (NS) difference of beetle occurrence (one-way ANOVAmonsoon, F4, 355 = 5.88, NS; one-way ANOVApost-monsoon, F4, 355 = 4.24, NS) across fields both in monsoon and post-monsoon periods, (b) non-significant (even letters) difference (non-parametric t-test) of beetle occurrences in a field for both seasons
Considering the assessment from Crowder et al. (2010), species evenness might be considered a vital response variable in the present study as our result indicates stable evidence of beetle assemblage across the study fields. Our surveyed landscape was surrounded by natural open land with a large percentage of hardwood, natural forest, and livestock farming areas. Although landscape characteristics (Bommarco 1998; Östman et al. 2001) as well as surrounding landscape structure (Purtauf et al. 2005) are essential for species diversity and richness for different groups of organisms, including Coleopteran insects, seasonal variation of temperature and rainfall play a significant role in species diversity, richness, and evenness for sustainable organic farming (Marino and Landis 1996; Jonsen and Fahrig 1997; Svensson et al. 2000; Crowder et al. 2010; Pramual and Wongpakam 2010). Beetle diversity indices in our study also echo with this fact that temperature and rainfall during monsoon favor significantly for their assemblage.
The information presented in the present study is suggesting the contributory role of Coleopteran beetles for organic farming, and henceforth, it is often considered that they might be a possible solution for biodiversity conservation in intensive agricultural landscapes (Mone et al. 2014). On the other hand, the rarely occurred Coleopteran families, where occurrence frequencies were restricted to < 1%, were fragmentarily appeared in the landscape, suggesting the necessities for their conservation, restoration, and management in such habitats for sustainable organic farming. In addition, the rare Coleopteran families are essential for predicting the relationship between insect diversity and ecosystem functions; therefore, the importance of their conservation is imperative (Haines-Young and Potschin 2010).
Conclusion
The outcome of the present study demonstrates the role of the Coleopteran beetle community as an enhancer in organic farming for both the monsoon and post-monsoon seasons. Our study also shows that beetle diversity in organic farms might contribute to the pest-predator balance in the landscape structure. Since organic farming leads to a high diversity of Coleopteran beetles, and some representatives among those insects exemplify good biological indicators for such farms, preserving beetle communities is imperative. The present study concludes that sustainable organic farming supports the diversity of Coleopteran beetles as a significant representative taxon among insect communities. Though it is reasonably realized that some Coleopteran beetles are harmful pests for certain crops but help organic farming by improving soil conditions through nutrient recycling, hence, more scientific reviews can be done on how harmful they are and how well they can play a beneficial role in organic farming.
Data availability
All sample materials are kept at “insect repository” at the Plant Protection Unit, Faculty of Agriculture National University of Laos, Lao PDR.
Code availability
Not applicable.
References
Adhikari S, Menalled FD (2020) Supporting beneficial insects for agricultural sustainability: the role of livestock-integrated organic and cover cropping to enhance ground beetle (Carabidae) communities. Agronomy 10(8):1210. https://doi.org/10.3390/agronomy10081210
Beattie AJ, Oliver I (1994) Taxonomic minimalism. Trends in Evolution and Ecology 9(488):490
Bommarco R (1998) Reproduction and energy reserves of a predatory carabid beetle relative to agroecosystem complexity. Ecol Appl 8(3):846–853. https://doi.org/10.1890/1051-0761(1998)008[0846:RAEROA]2.0.CO;2
Burgio G, Sommaggio D, Marini M, Puppi G, Chiarucci A, Landi S, Masetti A (2015) The influence of vegetation and landscape structural connectivity on butterflies (Lepidoptera: Papilionoidea and Hesperiidae), carabids (Coleoptera: Carabidae), syrphids (Diptera: Syrphidae), and sawflies (Hymenoptera: Symphyta) in Northern Italy farmland. Environ Entomol 44(5):1299–1307. https://doi.org/10.1093/ee/nvv105
Crowder DW, Northfield TD, Strand MR, Snyder WE (2010) Organic agriculture promotes evenness and natural pest control. Nature 466(7302):109–112. https://doi.org/10.1038/nature09183
Culman SW, Young-Mathews A, Hollander AD, Ferris H, Sánchez-Moreno S, O’Geen AT, Jackson LE (2010) Biodiversity is associated with indicators of soil ecosystem functions over a landscape gradient of agricultural intensification. Landscape Ecol 25(9):1333–1348. https://doi.org/10.1007/s10980-010-9511-0
Dritschilo W, Erwin TL (1982) Responses in abundance and diversity of cornfield carabid communities to differences in farm practices. Ecology 63(4):900–904
Echegaray EA, Cloyd RA, Nechols JR (2015) Rove beetle (Coleoptera: Staphylinidae) predation on Bradysia sp. nr. coprophilia (Diptera: Sciaridae). J Entomol Sci 50(3):225–237. https://doi.org/10.18474/JES14-38.1
Gaines HR, Gratton C (2010) Seed predation increases with ground beetle diversity in a Wisconsin (USA) potato agroecosystem. Agr Ecosyst Environ 137(3–4):329–336. https://doi.org/10.1016/j.agee.2010.03.003
Gimmel ML, Aston P (2010) New records of the family Phalacridae from Hong Kong (Coleoptera: Cucujoidea). Hong Kong Entomological Society 2(1):11–12
Haines-Young R, Potschin M (2010) The links between biodiversity, ecosystem services and human well-being. Ecosystem Ecology: a new synthesis 1:110–139
Hanula JL, Ulyshen MD, Horn S (2011) Effect of trap type, trap position, time of year, and beetle density on captures of the redbay ambrosia beetle (Coleoptera: Curculionidae: Scolytinae). J Econ Entomol 104(2):501–508. https://doi.org/10.1603/EC10263
Hole DG, Perkins AJ, Wilson JD, Alexander IH, Grice PV, Evans AD (2005) Does organic farming benefit biodiversity? Biol Cons 122(1):113–130. https://doi.org/10.1016/j.biocon.2004.07.018
Hutton SA, Giller PS (2003) The effects of the intensification of agriculture on northern temperate dung beetle communities. J Appl Ecol 40(6):994–1007. https://doi.org/10.1111/j.1365-2664.2003.00863.x
Hurej M, Twardowski JP, Kozak M (2013) Weevil (Coleoptera: Curculionidae) assemblages in the fields of narrow-leafed lupin sown as pure stand and intercropped with spring triticale. Žemdirbyste-Agriculture 100:393–400. https://doi.org/10.13080/z-a.2013.100.050
Inclán DJ, Cerretti P, Gabriel D, Benton TG, Sait SM, Kunin WE, Marini L (2015) Organic farming enhances parasitoid diversity at the local and landscape scales. J Appl Ecol 52(4):1102–1109. https://doi.org/10.1111/1365-2664.12457
Jonsen ID, Fahrig L (1997) Response of generalist and specialist insect herbivores to landscape spatial structure. Landscape Ecol 12:185–197. https://doi.org/10.1023/A:1007961006232
Kubo JS, Torgersen CE, Bolton SM, Weekes AA, Gara RI (2013) Aquatic insect assemblages associated with subalpine stream segment types in relict glaciated headwaters. Insect Conservation and Diversity 6(3):422–434. https://doi.org/10.1111/j.1752-4598.2012.00210.x
Lichtenberg EM, Kennedy CM, Kremen C, Batary P, Berendse F, Bommarco R, Bosque-Pérez NA, Carvalheiro LG, Snyder WE, Williams NM, Winfree R (2017) A global synthesis of the effects of diversified farming systems on arthropod diversity within fields and across agricultural landscapes. Glob Change Biol 23(11):4946–4957. https://doi.org/10.1111/gcb.13714
Macfadyen S, Gibson R, Polaszek A, Morris RJ, Craze PG, Planqué R, Memmott J (2009) Do differences in food web structure between organic and conventional farms affect the ecosystem service of pest control? Ecol Lett 12(3):229–238. https://doi.org/10.1111/j.1461-0248.2008.01279.x
Magurran AE (2004) Measuring biological diversity. Blackwell Science, Oxford
Manning P, Slade EM, Beynon SA, Lewis OT (2016) Functionally rich dung beetle assemblages are required to provide multiple ecosystem services. Agr Ecosyst Environ 218:87–94. https://doi.org/10.1016/j.agee.2015.11.007
Martins ICF, Cividanes FJ, Ide S, Haddad GQ (2013) Diversity and habitat preferences of Carabidae and Staphylinidae (Coleoptera) in two agroecosystems. Brigantia 71(4):471–480. https://doi.org/10.1590/S0006-87052013005000009
Marino PC, Landis DA (1996) Effect of landscape structure on parasitoid diversity and parasitism in agroecosystems. Ecol Appl 6(1):276–284. https://doi.org/10.2307/2269571
Mone S, Kusha KM, Jathanna D, Ali M, Goel A (2014) Comparison of insect biodiversity between organic and conventional plantations in Kodagu, Karnataka, India. J Threat Taxa 6(9):6186–6194. https://doi.org/10.11609/JoTT.o3778.6186-94
Montañez MN, Amarillo-Suárez Á (2014) Impact of organic crops on the diversity of insects: a review of recent research. Revista Colombiana De Entomología 40(2):131–142
Oliver JB, Mannion CM (2001) Ambrosia beetle (Coleoptera: Scolytidae) species attacking chestnut and captured in ethanol-baited traps in middle Tennessee. Environ Entomology 30(5):909–918. https://doi.org/10.1603/0046-225X-30.5.909
Östman Ö, Ekbom B, Bengtsson J, Weibull AC (2001) Landscape complexity and farming practice influence the condition of polyphagous carabid beetles. Ecol Appl 11(2):480–488. https://doi.org/10.1890/1051-0761(2001)011[0480:LCAFPI]2.0.CO;2
Panyakul V (2009) Organic agriculture in Laos PDR: overview and development options. Earth Net Foundation/Green Net. International Trade Center’s project “Support to trade promotion and export development in Laos PDR” Project number LAO/61/89
Perfecto I, Vandermeer J (2010) The agroecological matrix as alternative to the land-sparing/agriculture intensification model. Proc Natl Acad Sci 107(13):5786–5791. https://doi.org/10.1073/pnas.0905455107
Peuhu E, Thomssen PM, Siitonen J (2019) Comparison of three trap types in sampling saproxylic beetles living in hollow urban trees. J Insect Conserv 23(1):75–87. https://doi.org/10.1007/s10841-018-0115-3
Pielou EC (1966) The measurement of diversity in different types of biological collections. J Theor Biol 13:131–144. https://doi.org/10.1016/0022-5193(66)90013-0
Pramual P, Wongpakam K (2010) Seasonal variation of black fly (Diptera: Simuliidae) species diversity and community structure in tropical streams of Thailand. Entomological Science 13(1):17–28. https://doi.org/10.1111/j.1479-8298.2009.00354.x
Purtauf T, Roschewitz I, Dauber J, Thies C, Tscharntke T, Wolters V (2005) Landscape context of organic and conventional farms: influences on carabid beetle diversity. Agr Ecosyst Environ 108(2):165–174. https://doi.org/10.1016/j.agee.2005.01.005
Reganold JP, Wachter JM (2016) Organic agriculture in the twenty-first century. Nature Plants 2(2):1–8. https://doi.org/10.1038/nplants.2015.221
R Development Core Team (2019) R: a language and environment for statistical computing. Vienna (Austria): R Foundation for Statistical Computing. https://www.R-project.org/. Accessed 31 Oct 2019
Rodrigues MM, Uchoa MA, Ide S (2013) Dung beetles (Coleoptera: Scarabaeoidea) in three landscapes in Mato Grosso do Sul. Brazil Brazilian Journal of Biology 73(1):211–220. https://doi.org/10.1590/S1519-69842013000100023
Russo L, Stehouwer R, Heberling JM, Shea K (2011) The composite insect trap: an innovative combination trap for biologically diverse sampling. PLoS ONE 6(6):e21079. https://doi.org/10.1371/journal.pone.0021079
Sawada Y, Hirowatari T (2002) A revision of the genus Acrotrichis Motschulsky (Coleoptera: Ptiliidae) in Japan. Entomological Science 5:77–101
Seufert V, Ramankutty N, Foley JA (2012) Comparing the yields of organic and conventional agriculture. Nature 485(7397):229–232. https://doi.org/10.1038/nature11069
Shah PA, Brooks DR, Ashby JE, Perry JN, Woiwod IP (2003) Diversity and abundance of the Coleopteran fauna from organic and conventional management systems in southern England. Agric for Entomol 5(1):51–60. https://doi.org/10.1046/j.1461-9563.2003.00162.x
Smith OM, Cohen AL, Reganold JP, Jones MS, Orpet RJ, Taylor JM, Kennedy CM (2020) Landscape context affects the sustainability of organic farming systems. Proc Natl Acad Sci 117(6):2870–2878. https://doi.org/10.1073/pnas.1906909117
Snyder WE (2019) Give predators a complement: conserving natural enemy biodiversity to improve biocontrol. Biol Control 135:73–82. https://doi.org/10.1016/j.biocontrol.2019.04.017
Steininger MS, Hulcr J, igut M, Lucky A, (2015) Simple and efficient trap for bark and ambrosia beetles (Coleoptera: Curculionidae) to facilitate invasive species monitoring and citizen involvement. J Econ Entomol 108(3):1115–1123. https://doi.org/10.1093/jee/tov014
Svensson B, Lagerlöf JG, Svensson B (2000) Habitat preferences of nest-seeking bumblebees (Hymenoptera: Apidae) in an agricultural landscape. Agr Ecosyst Environ 77(3):247–255
Tewari S, Leskey TC, Nielsen AL, Piñero JC, Rodriguez-Saona CR (2014) Use of pheromones in insect pest management, with special attention to weevil pheromones. In: Integrated pest management. Academic Press, pp 141–168. https://doi.org/10.1016/B978-0-12-398529-3.00010-5
Tuck SL, Winqvist C, Mota F, Ahnström J, Turnbull LA, Bengtsson J (2014) Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J Appl Ecol 51(3):746–755. https://doi.org/10.1111/1365-2664.12219
van Elsen T (2000) Species diversity as a task for organic agriculture in Europe. Agr Ecosyst Environ 77(1–2):101–109. https://doi.org/10.1016/S0167-8809(99)00096-1
Wardhaugh CW, Stone MJ, Stork NE (2018) Seasonal variation in a diverse beetle assemblage along two elevational gradients in the Australian Wet Tropics. Sci Rep 8(1):1–12. https://doi.org/10.1038/s41598-018-26216-8
Whittaker RH (1972) Evolution and measurement of species diversity. Taxon 21(2/3):213
Wickramasinghe LP, Harris S, Jones G, Vaughan Jennings N (2004) Abundance and species richness of nocturnal insects on organic and conventional farms: effects of agricultural intensification on bat foraging. Conserv Biol 18(5):1283–1292
Yamada K, Yanagisawa M, Kono Y, Nawata E (2004) Use of natural biological resources and their roles in household food security in northwest Laos. Japanese Journal of Southeast Asian Studies 41(4):426–443
Acknowledgements
We want to thank the plant protection unit at NUoL, Laos, for field collections and species identification. The authors are thankful to Joseph Thomas Milburn for reviewing the text.
Author information
Authors and Affiliations
Contributions
Both AD and BC conceptualized the idea and BC and SS designed the experiment. Both BC and SS conducted the survey and sample collection, where AD and BC analyzed the data. All graphical illustrations were prepared by AD. Both BC and SS drafted the manuscript initially, and later, AD revised the draft finally.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
All authors are providing consent for publication of this manuscript.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chouangthavy, B., Sanguansub, S. & Das, A. Sustainable organic farming supports diversity of Coleopteran beetles as a good indicator taxon: a case study from central Lao PDR. Org. Agr. 11, 615–624 (2021). https://doi.org/10.1007/s13165-021-00367-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13165-021-00367-x