
Abstract
Herons typically reproduce colonially, either monospecific or mixed, over waterbodies, high in trees or shrubs, mangroves, and islands. Information from neotropical mangroves is lacking concerning heron breeding areas, reproductive biology, and functional relationships with mangroves. Here we compared the reproductive biology of Cattle Egret (Bubulcus ibis), Little Blue Heron (Egretta caerulea), and Snowy Egret (E. thula), with emphasis on breeding season, nesting materials, nest height, clutch size, breeding success, and nest failure in a mangrove ecosystem in Sepetiba Bay, southeastern Brazil. The breeding season lasted from November to mid-February (195 nests). Nests were built mostly with twigs, but also with grasses (17 nests, 9%) and nylon lines and ropes (Cattle Egrets: 16 nests, 28%; Snowy Egrets: 4 nests, 14%). Little Blue Herons nested higher (2.25 ± 0.1 m) than Cattle Egrets (1.97 ± 0.06 m) and Snowy Egrets (1.80 ± 0.1 m). The modal clutch size differed among species, with two eggs in Cattle Egrets (1–7 range) and Little Blue Herons (1–4 range) and three eggs in Snowy Egrets (2–4 range). Breeding success was high both in Cattle Egrets (90%) and Snowy Egrets (89%) but was low in Little Blue Herons (41%). Higher nest failure in Little Blue Herons occurred due to localized predation events in all clumped nests built isolated at the colony edge. Therefore, these predation events suggest that interspecific nest aggregation was important to diminish nest failure. The remaining nests were quite successful, even though brood reduction (common in herons) occurred in most nests.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Herons and egrets (family Ardeidae) are wading birds associated with wetlands worldwide (Sick 1997; Lovette and Fitzpatrick 2016). Most of their activities, including foraging, rest, courtship, and reproduction, occur along the margins of water bodies with nearby vegetation (Grose et al. 2014; Cintra 2015). Egrets are typically just smaller herons (hereafter simply herons), and herons play important roles in trophic relationships in wetlands. They contribute to nutrient enrichment in local food webs by nutrient cycling between wetlands and adjacent terrestrial habitats through prey capture and excretion (Green and Elmberg 2014; Faria et al. 2016). Heron presence in wetlands is associated with several environmental characteristics, including resource availability, vegetation structure and composition, water column depth, and available habitat for nesting (Tavares et al. 2015; Zhang et al. 2018). Thus, for herons, wetland quality influences habitat selection for both foraging and reproduction (Cintra 2015; Almeida et al. 2016).
Mangroves are coastal wetland ecosystems found between terrestrial and marine environments in tropical and subtropical latitudes worldwide (Schaeffer-Novelli 1995). They are more abundant or dense in areas protected from intense wave and tidal forces, such as estuaries, bays, and lagoons, where they provide substrate and habitat for a variety of animals, including herons (Schaeffer-Novelli 1995; Ruiz-Guerra and Echeverry-Galvis 2019). In the eastern seaboard of the Americas, mangroves are found from the United States to southern Brazil, and Brazil has the second largest extent of mangroves in the world (ICMBio 2018). Mangroves are among the most threatened ecosystems worldwide and their loss and degradation pose major threats to bird species dependent on these habitats (Ávila et al. 2019; Martínez et al. 2020). This ecosystem is a critical habitat for many bird species that are associated with them, often for foraging and nesting (Etezadifar and Barati 2013; Ávila 2019).
Reproduction of herons in mangroves is determined mostly by vegetation structure, but also by productivity in surrounding waters (Etezadifar and Barati 2013, 2015; Cox et al. 2019). Vegetation structure and productivity also influence heron coloniality (Kelly et al. 2008; Scherer et al. 2014). Colonial birds may often be bioindicators of local conditions as the size of breeding populations and breeding success vary depending on resources, species aggregation, and predation risk (Kushlan 1993; Kelly et al. 2008; Olguín et al. 2015; Zhao et al. 2017). Colonial breeding may increase nesting success through greater foraging efficiency (due to interspecific interaction) and lower predation rates (Brown and Brown 1987; Branco 2003).
Herons tend to breed in monospecific or mixed colonies in trees and shrubs over waterbodies, in mangroves, and on islands close to feeding areas (Gianuca et al. 2012; Koczur et al. 2018). Some species, such as Cattle Egrets, may breed farther from where they forage to improve their breeding success (Weller 1999; Metallaoui et al. 2020). Reproductive traits, such as clutch size, nest site and structure, and breeding success, in mixed colonies may vary among species due to microclimate conditions and interspecific competition (Johnsgard 2009). Herons usually compete for suitable nest substrates and nest sites within the colony. Nesting trees with higher canopy spread, girth size, and quality protect against predators and heavy weather conditions and provide nesting materials and easy access to nest locations (Kelly and Condeso 2014; Roshnath and Sinu 2017). Nests in central regions of the colony may also decrease predation risk in relation to nests in more peripheral areas (Uzun 2009). Therefore, mixed colonies can provide valuable comparative data about the effects of microclimate variation and interspecific interactions in heron breeding success under similar weather conditions (Martínez et al. 2020).
The reproductive biology of herons in mangroves is poorly studied, especially in South America (Xiao et al. 2017; Martínez et al. 2020). Here, we compare the reproductive biology of Cattle Egret (Bubulcus ibis), Little Blue Heron (Egretta caerulea), and Snowy Egret (E. thula) in a mangrove in southeastern Brazil. Specifically, we compare: 1) breeding season, 2) nesting materials, 3) nest height distribution, 4) clutch size, 5) breeding success, and 6) nest failure. We highlight the importance of these results to understand the poorly-known reproductive biology of herons in neotropical mangroves and to support the conservation of mangroves in close proximity to urban/industrial development.
Materials and methods
Study Area
Our study was conducted in Sepetiba Bay (22º53’ to 23º05’S, 44º01’ to 43º33’W), one of the largest extents of mangrove remaining in the state of Rio de Janeiro, southeastern Brazil, with about 38 km² (Rezende et al. 2007; Soares et al. 2011). Average annual precipitation varies from 1,000 mm to 2,230 mm, with a maximum from December to March (summer) and a minimum from June to August (winter). The climate is classified as rainy tropical (tropical humid, Af-Am-Aw) and mild subtropical (Wa) according to the Köppen classification (SEMEADS 2001).
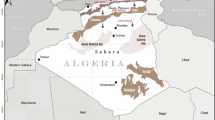
The central region of Sepetiba Bay has a small and dense forest with an almost continuous canopy dominated by white mangrove (Laguncularia racemose), black mangrove (Avicennia germinans, A. schaueriana), and red mangrove (Rhizophora mangle). Its continental side is limited by highways, railways, and urban occupation. Natural flood dynamics are unrestricted by anthropic influence, with freshwater flows towards the intertidal zone. The mangrove is flooded daily by tides. The heron colony was at the southern end of this mangrove, in a forest fragment of 700 m2 dominated by A. schaueriana and few R. mangle (Fig. 1).

Location map showing the study area in Sepetiba Bay, state of Rio de Janeiro, southeastern Brazil and the three-heron mixed colony (hatched region) in the mangrove forest. TLS provided the silhouette illustration
Reproduction and Nest Monitoring
The colony was monitored from November 2014 to March 2015, at three- to seven-day intervals. We started marking nests 20 days after the first egg-laying record to minimize human disturbance. Due to the limited access by foot, we always checked nests when the tide was ≤ 0.8 m as stated in the Itaguaí Port (CHM 2014) tide table. We randomly monitored a subset of nests in the two nest-site groups (edge and interior). All monitored nests were marked and identified to species level to ensure correct identification on subsequent observations. We noted the beginning of nest construction, nesting materials, clutch size, nest height, fledging date, and nest outcome. Nest status (eggs and nestlings) was checked through an extensible telescopic rod with a mirror attached to its end that reached each nest and allowed the view of nest contents. Reproductive periods were classified as incubation period, which began when the first egg was laid and ended when the first egg hatched and nestling period, which began after the first egg hatched and ended when all young fledged (Marini et al. 2009; Fierro-Calderón et al. 2021). Nesting materials were classified as twigs with or without mangrove leaves, grasses, and nylon lines and ropes. They were compared by species as the proportion of nests containing each material of the total number of monitored nests. Nest height was measured with 5 cm precision using a 3.5 m graduated telescopic rod. Nest-site groups were classified as edge (with adjacent nests only on one side, the other side had water) and interior (nests had adjacent nests on any side).
We assumed that all transitions (egg-laying, hatching, failure, and fledging) occurred at the midpoint between nest checks when we did not directly observe them. This assumption was used to estimate the incubation (first egg laid to last egg hatched) and nestling (hatching to fledging) periods. Otherwise, we noted the exact date. Nests were successful if at least one young fledged.
Nest failure was classified as predation if eggshells, dead and damaged nestlings or feathers were found inside and around the nest. Abandonment was assumed if unhatched eggs remained on the nest with no sign of parents after ~ 25 days from egg-laying. Destroyed nests were lost due to weather or tide. Rarely, a nest may have been destroyed by a predator rather than weather, but without evidence, we classified it as destroyed.
Data Analysis
We compared nest height, clutch size, breeding success, and nest survival among species. Nest height was assessed by linear model test using log10-transformed data to meet the assumptions of normality and homogeneity of variances. Clutch size and breeding success were compared using contingency tables and G-test. Breeding success was estimated as the proportion of successful nests of all nesting attempts by species. All analyses were conducted in the R 4.1.0 software environment (R Core Team 2021). The linear model was fitted in stats package (R Core Team 2021) using lm function. G-test was conducted in DescTools package (Signorell et al. 2021) using GTest function. Nest survival was calculated in RMark package (Laake 2013) using mark function. Nest survival analysis requires three dates for each nest: (1) the day the nest was found, (2) the last day the nest was active (with eggs or nestlings), and (3) the last day the nest was checked. The analysis also requires the nest outcome, calculated as a binary variable: successful (1) and failed (0) (Dinsmore et al. 2002). We modeled nest survival as a function of species. Figures were generated in ggplot2 package (Wickham 2016) and QGIS (QGIS.org. 2022).
Results
A total of 195 nests were monitored: 58 Cattle Egrets, 29 Little Blue Herons, and 28 Snowy Egrets. An additional 80 nests were also found, but we were unable to identify the resident. These unidentified nests were only used to estimate overall breeding success and compare success by nest-site group (edge and interior). Nests were found only with eggs (51 Cattle Egret nests, 26 each Little Blue Heron and Snowy Egret nests, total of 103 nests), and with eggs and newly-hatched nestlings (7 Cattle Egrets, 3 Little Blue Herons, and 2 Snowy Egrets, total of 12 nests).
Nesting materials comprised of loosely arranged dry sticks (all nests, all species), grass collected on the beach around the colony (17 nests, 9%), and nylon lines and ropes from fishing lines, nets, and mooring ropes, found in 16 (28%) Cattle Egret and 4 (14%) Snowy Egret nests.
Little Blue Herons nested higher (2.25 ± 0.1 m) than Cattle Egrets (1.97 ± 0.06 m) and Snowy Egrets (1.80 ± 0.1 m) (F2,112 = 5.30, r2 = 0.09, p = 0.006) (Fig. 2). Little Blue Herons constructed both clumped nests (15, 52%) isolated at an edge of the colony and scattered nests throughout the colony (14, 48%).

Boxplot of nest height distributions for Cattle Egret, Little Blue Heron, and Snowy Egret nests in the breeding colony of Sepetiba Bay, southeastern Brazil. The boxplot shows medians, quartiles, 5th and 95th percentiles, raw data, mean values (black dots), and p-values
The modal clutch size differed among species (G = 23.23, df = 12, p = 0.026), with two eggs in Cattle Egrets (1–7 range) and Little Blue Herons (1–4), and three eggs in Snowy Egrets (2–4) (Fig. 3a). Average clutch size was 3.0 ± 0.2 eggs in Cattle Egrets, 2.3 ± 0.1 in Little Blue Herons, and 2.7 ± 0.1 in Snowy Egrets. Cattle Egrets had the largest clutch size, with 6–7 egg clutches (1 nest, 0.9% each) that fledged only 2 young per nest and 5-egg clutches (5 nests, 4.3%) that fledged 2.4 young per nest. Among all species, 2-egg clutches were the most frequent (50 nests, 43.5%) but the least productive (1.28 young per nest).

Reproductive output for Cattle Egret, Little Blue Heron, and Snowy Egret nests in the breeding colony of Sepetiba Bay, southeastern Brazil. (a) Clutch size, demonstrating that Cattle Egrets had the highest variation in clutch size, while Snowy Egrets had the greatest median clutch size. (b) Number of fledglings per nest, showing that all species converged on two young fledged per nest
Egg-laying began in the last half of November and the first nestlings hatched on December 10th (Little Blue Heron), 12th (Cattle Egret), and 13th (Snowy Egret). Most nestlings occurred in December (Little Blue Herons) and January (Cattle Egrets, Snowy Egrets). In February, only two Snowy Egret nests still had nestlings (Fig. 4).

Number of eggs and nestlings of Cattle Egrets, Little Blue Herons, and Snowy Egrets in the breeding colony of Sepetiba Bay, southeastern Brazil
All species fledged young, and around 15 January, a total of 51 nests (57% of all successful nests) had fledged. The last fledglings were from two Snowy Egret nests (5 and 6 February). By mid-February, the colony no longer had nesting birds, and adults and young were observed nearby. By mid-March, juveniles appeared to be scattered and were no longer attended by parents.
Nest failure was due to predation, abandonment, adverse weather, and sibling competition. Both nest-site groups had similar nest failure rates (43% edge, 34% interior, overall 36% failure, G = 1.28, df = 1, p = 0.258). We observed several potential nest predators around the colony, including Black Vulture (Coragyps atratus), White-tailed Hawk (Geranoaetus albicaudatus), Great Kiskadee (Pitangus sulphuratus), and Black-crowned Night-heron (Nycticorax nycticorax). Reptiles and mammals or their footprints were not observed, except for a Teiidae lizard found in one nest. Agonistic interactions were observed between Great Kiskadee and heron nestlings. Breeding success was high both in Cattle Egrets (52 of 58 nests, 90%) and Snowy Egrets (25 of 28 nests, 89%), with lower success in Little Blue Herons (12 of 29 nests, 41%, G = 25.95, df = 2, P < 0.001). Nest survival rates were 99.7% day-1 in Cattle Egrets (SE = 0.0012), 97.1% day-1 in Little Blue Herons (SE = 0.0070, p < 0.001), and 99.7% day-1 in Snowy Egrets (SE = 0.0020). Most nest failures occurred during the incubation period in about half of Little Blue Heron nests (14, 48%), followed by Cattle Egrets (3, 5%), and Snowy Egrets (1, 4%). Nest failure during incubation in Little Blue Herons occurred in the last half of December due to localized predation events in all clumped nests. Brood reduction (fewer young fledged than eggs hatched) occurred in all three species, mostly in Cattle Egrets (27 nests, loss of 1–5 young), followed by Snowy Egrets (14 nests, loss of 1–2 young), and Little Blue Herons (7 nests, loss of 1–2 young). Few nests failed during the nestling period (aside from brood reduction), with only three losses in Cattle Egrets and Little Blue Herons and two losses in Snowy Egrets.
Overall, 176 fledglings were produced (106 in Cattle Egrets nests, 22 in Little Blue Herons, and 48 in Snowy Egrets, Fig. 3b). Thus, of 116 breeding Cattle Egret adults, 106 young were produced (92% replacement, 63% of eggs laid); of 58 Little Blue Heron adults, 22 young were produced (38% replacement, 34% of eggs laid); and of 56 Snowy Egret adults, 48 young were produced (86% replacement, 63% of eggs laid).
Discussion
We found lower breeding success in Little Blue Heron nests (41%, 34% of eggs laid) compared to greater success in the other colony nesting species (~ 90% in both Cattle Egrets and Snowy Egrets). Local predation events may explain its lower breeding success as all Little Blue Heron clumped nests failed in the last half of December. Therefore, interspecific aggregation within a breeding colony may play an important role to decrease predation risk. In general, nests had relatively high success, with brood reduction occurring in 52% of Cattle Egret, 58% of Little Blue Heron, and 56% of Snowy Egret nests. Despite these losses, all three species are relatively long-lived, with longevity records of 11–16 years (Clapp et al. 1982), so lifetime breeding success should be much greater than simple replacement at these rates, including the Little Blue Herons. Although its proximity to urban areas increased the use of anthropogenic debris as nesting materials, the overall high breeding success of most nests can place this mangrove as an important breeding site for herons.
The breeding season begins in late spring and continues through summer in the southern hemisphere. In our breeding colony, the breeding season started later and ended sooner than is typical for herons in temperate North America, where they begin nesting in late winter and continue through late summer (Hancock and Kushlan 2010). In Sepetiba Bay, breeding is timed for the rainy season, which is similar to other locations in the USA, Mexico, Cuba, Argentina, and southern Brazil (Ávila et al. 2003; Mellink and Riojas-López 2008; Josens et al. 2009; Gianuca et al. 2012; Bisinela et al. 2014; Baker and Dieter 2015). Just a short distance farther south, however, in the state of São Paulo, Little Blue Herons began breeding a month sooner than Sepetiba Bay (Olmos and Silva e Silva 2002). Climate is very similar in the two mangroves (though somewhat warmer in Sepetiba Bay), and this suggests that factors affecting breeding patterns may be subtle. Some evidence suggests that breeding season length may increase with colony size (Shirai 2013), but both breeding season length and colony size may be associated with other variables.
In tropical systems, breeding season length can allow multiple broods per year as a strategy to increase annual breeding success (Roper 2005). However, here we observed only one brood during the breeding season. Nests that started later may have been after a previous nest failure within the breeding season, but this would only be likely for Little Blue Herons as breeding success was much greater in the other two species. A single brood per breeding season is typical of herons in temperate latitudes (Johnsgard 2009), so perhaps the limits on breeding season length here do not allow multiple nests per pair unless they were replacement nests after failure (Parsons and Master 2000). Herons remaining in the vicinity of their nesting colonies for up to two months after breeding seems to be typical (Browder 1973; Erwin et al. 1996; Cox et al. 2019).
Nests were similar to those reported elsewhere (Sick 1997; Lovette e Fitzpatrick 2016). They were mostly built on Avicennia schaueriana with few nests (mainly Little Blue Herons) on Rhizophora mangle. However, the use of nylon lines and ropes was unusual and only occurred in two of the three herons. The anthropogenic nest materials in our breeding colony could be related to its close location to fishing communities, urban populations, and important Brazilian harbors. Anthropogenic debris as nesting materials is increasing in birds nesting in coastal wetlands (Brentano et al. 2020; Thompson et al. 2020). Their use can lead to injury or death of adults and chicks through entanglement and ingestion (Witteveen et al. 2017; Ryan 2018). Thus, management of fishing waste and other anthropogenic debris, such as initiatives to collect and recycle nylon lines and ropes, should be conducted to decrease the risk of heron mortality (Yorio et al. 2022).
Nest-site selection is paramount and requires suitable substrates to ensure breeding success (Roshnath and Sinu 2017; Metallaoui et al. 2020). In mixed nesting colonies, the nest-site selection is usually related to bird size (larger species dominate) and arrival order (first to arrive get the best nest sites) at the colony (Kim and Koo 2009). However, Cattle Egrets tend to select nest sites based on other variables rather than simply height (Burger and Gochfeld 1990; Scherer et al. 2014). Here, although Little Blue Heron nests were placed higher than Cattle Egrets in the nesting trees, we did not observe a strong vertical nest stratification as all species were similar in size (smallest Cattle Egrets − 334 g, largest Little Blue Herons − 399 g, Ávila 2011) and arrived about the same time. This may suggest that suitable conditions for breeding favor the synchronous arrival of all three species, and they independently select a nest site without much interference from the others.
The average clutch size ranged from 2.3 in Little Blue Herons to 3.0 in Cattle Egrets. Avian clutch size tends to be greater in more temperate regions (4.5 eggs) and decreases at lower, tropical, latitudes (2 eggs) (Ricklefs 1980; Jetz et al. 2008). Herons also follow this pattern, for example, Black-crowned Night Herons (Nycticorax nycticorax) reached an average clutch size of 3.5 in Korea (37°N) (Jungsoo and Tae-Hoe 2007) and 2.4 in Brazil (2°S) (Martínez et al. 2020). Therefore, clutch sizes found in our colony are similar to other herons nesting in tropical wetlands (e.g., Olmos and Silva e Silva 2002; Gómez et al. 2006; Olguín et al. 2015).
Larger clutches may fledge more young but are less frequent. In our colony, 5-egg clutches were more productive (2.4 fledglings per nest) and less frequent (4.3%), except for 7 − 6 clutches, which produced only 2 fledglings. On the other hand, 2-egg clutches were less productive (1.3 fledglings per nest) and more frequent (43.5%). Several hypotheses have been proposed to explain this pattern as higher clutch size may be constrained by nest size (Slagsvold 1982), number of brood patches (Hills 1980), food availability (Lack 1968), individual condition (Perrins and Moss 1975), predation risk (Lack 1968; Slagsvold 1982), parental effort (Lindén and Møller 1989), sibling competition (Mock 1987), and parasite load (Moss and Camin 1970). In herons, larger clutches may represent insurance against egg infertility or loss (Martínez et al. 2020), and asynchronous hatching can favor sibling competition and aggression (Mock 1987), both leading to brood reduction in larger clutches.
Avian breeding success tends to be lower in tropical latitudes due to greater nest predation (Ricklefs 1969; Martin 1995). However, herons had higher success (~ 90% in both Cattle Egrets and Snowy Egrets) when compared to other subtropical locations in Brazil, such as Cattle Egrets with 63% success and Snowy Egrets with 50% in one reservoir environment (Petry and Fonseca 2005). Indeed, the breeding success of Cattle Egrets and Snowy Egrets in our colony was quite similar to northern latitudes where predation rates are lower (Baker and Dieter 2015). While breeding in mixed colonies may often result in reduced breeding success for some species, we found no evidence for interspecific interactions that would have reduced breeding success. Indeed, interspecific interactions may have favored higher breeding success in most species. Nest failure in herons often occurs during the incubation period (Cupul-Magaña 2004; Olguín et al. 2015), even with reduced predation risk provided by mixed colonies (Branco 2003). Nest failure may be higher at the edge than at the interior of the colony due to more exposure to predators and weather conditions (Uzun 2009). However, we did not find this pattern in our colony, yet Little Blue Heron clumped nests built isolated from the other two species at the colony edge suffered higher nest predation. Therefore, interspecific aggregation of nests may play a more important role than nest sites to prevent nest failure.
Conclusion
Cattle Egrets, Little Blue Herons, and Snowy Egrets formed a mixed breeding colony of 230 pairs with high breeding success in a mangrove of Sepetiba Bay, southeastern Brazil. Little Blue Herons had lower success due to the loss of all clumped nests. These localized predation events may indicate that interspecific nest aggregation may reduce nest failure in mixed colonies. Overall, nests were very successful, even though brood reduction (common in herons) occurred in most nests. Thus, this colony in Sepetiba Bay produces more young than a simple replacement of their parents, and this productive colony may be an important source to provide individuals for other, less successful, colonies elsewhere.
Data Availability
The datasets generated in this study are available from the corresponding author on reasonable request.
Code Availability
Not applicable.
References
Almeida BA, Silva CB, Gimenes MR, Anjos L (2016) Waterbirds in a floodplain: influence of spatial and environmental factors through time. Rev Bras Ornitol. https://doi.org/10.1007/BF03544360
Ávila DD (2011) Patrones de crecimiento postnatal en ocho especies de garzas (Ciconiiformes: Ardeidae). Rev Biol Trop 59(2):771–787
Ávila DD, Rodríguez A, Rodríguez P, Jiménez A (2003) Reproducción de la garza ganadera (Bubulcus ibis) en la Ciénaga de biramas, Cuba. J Caribb Ornithol 16(1):45–54
Ávila DD, Bertinez EAC, Ferrer-Sanchez Y (2019) Modelación del efecto de la reproducción colonial de Bubulcus ibis (Aves: Ardeidae) en el flujo de biomasa y energía de los manglares costeros. Rev Biol Trop. https://doi.org/10.15517/rbt.v67i4.34822
Baker NJ, Dieter CD (2015) Reproductive success of colonial tree-nesting waterbirds in Prairie Pothole wetlands and rivers throughout northeastern South Dakota. Am Midl Nat. https://doi.org/10.1674/0003-0031-174.1.132
Bisinela GC, Andrade TR, Cremer MJ (2014) Ecologia reprodutiva de Nyctanassa violacea, Egretta thula e Egretta caerulea no ninhal do rio Pedreira, Santa Catarina. Biotemas. https://doi.org/10.5007/2175-7925.2014v27n3p109
Branco JO (2003) Reproduction of Sterna hirundinacea Lesson and S. eurygnatha Saunders, along Santa Catarina coast, Brazil. Rev Bras Zool. https://doi.org/10.1590/S0101-81752003000400017
Brentano R, de Brum AC, Montone RC, Petry MV (2020) Incidence of anthropogenic material in Sula leucogaster nests in a distant archipelago of Brazil. Mar Pollut Bull. https://doi.org/10.1016/j.marpolbul.2019.110815
Browder JA (1973) Long distance movements of cattle egrets. Bird Band. https://doi.org/10.2307/4511956
Brown CR, Brown MB (1987) Group-living in cliff swallows as an advantage in avoiding predators. Behav Ecol Sociobiol. https://doi.org/10.1007/BF02395437
Burger J, Gochfeld M, Waterbirds (1990) https://doi.org/10.2307/1521583
CHM (2014) Centro de Hidrografia da Marinha. Tábuas de Maré. Marinha do Brasil. https://www.marinha.mil.br/chm/tabuas-de-mare. Accessed 2 February 2021
Cintra R (2015) Spatial distribution and composition of waterbirds in relation to limnological conditions in the Amazon basin. https://doi.org/10.1007/s10750-014-2148-2. Hydrobiologia
Clapp RB, Klimkiewicz MK, Kennard JH (1982) Longevity records of north-american birds - Gaviidae through Alcidae. J Field Ornithol 53:81–124
Cox WA, Bankovich B, Malachowski K, Schwarzer AC, Paul A, Rachal M, Lorenz JJ, Meyer KD, Kent GM (2019) Nest site selection by reddish egrets in Florida. J Wildl Manage. https://doi.org/10.1002/jwmg.21577
Cupul-Magaña FG (2004) Observaciones sobre la anidación de tres especies de ardeidos en el estero Boca Negra. Jalisco México Huitzil 5(1):7–11
Dinsmore SJ, White GC, Knopf FL (2002) Advanced techniques for modeling avian nest survival. https://doi.org/10.1890/0012-9658(2002)083. [3476:ATFMAN]2.0.CO;2 Ecology
Erwin RM, Haig JG, Stotts DB, Hatfield JS (1996) Reproductive success, growth and survival of Black Crowned Night Heron (Nycticorax nycticorax) and Snowy Egret (Egreta thula) chicks in coastal Virginia. Auk. https://doi.org/10.2307/4088940
Etezadifar F, Barati A (2013) Nest-site selection of western reef Heron (Egretta gularis) in relation to mangrove (Avicennia marina) structure in the Persian Gulf: implication for management. https://doi.org/10.1016/j.foreco.2013.07.060. For Ecol Manag
Etezadifar F, Barati A (2015) Factors affecting offspring growth and daily nest survival rates in the coastal breeding western reef Heron (Egretta gularis) in the Persian Gulf. https://doi.org/10.1111/maec.12147. Mar Ecol
Faria FA, Silva-Costa A, Gianuca D, Bugoni L (2016) Cocoi Heron (Ardea cocoi) connects estuarine, coastal, limnetic and terrestrial environments: an assessment based on conventional dietary and stable isotope analysis. Estuar Coast. https://doi.org/10.1007/s12237-016-0073-5
Fierro-Calderón K, Loaiza‐Muñoz M, Sánchez‐Martínez MA, Ocampo D, David S, Greeney HF, Londoño GA (2021) Methods for collecting data about the breeding biology of neotropical birds. J Field Ornithol. https://doi.org/10.1111/jofo.12383
Gianuca D, Gianuca AT, Vooren CM (2012) Abundance, breeding and food of the little blue heron Egretta caerulea (Aves, Ardeidae) in the Patos lagoon estuary, a recently colonized area in southern Brazil. Iheringia Ser Zool. https://doi.org/10.1590/S0073-47212012000100003
Gómez J, Gil-Delgado JA, Monrós JS (2006) Breeding success of a colony of Boat-billed Herons Cochlearius cochlearius (Ciconiiformes: Ardeidae) in pasturelands of Costa Rica. Rev Biol Trop 54(4):1131–1134
Green AJ, Elmberg J (2014) Ecosystem services provided by waterbirds. Biol Rev. https://doi.org/10.1111/brv.12045
Grose AV, Cremer MJ, Moreira N (2014) Reprodução de aves aquáticas (Pelicaniformes) na Ilha do Maracujá, estuário da Baía da Babitonga, litoral norte de Santa Catarina. Biotemas. https://doi.org/10.5007/2175-7925.2014v27n2p117
Hills S (1980) Incubation capacity as a limiting factor of a shorebird clutch size. Am Zool 20:774
ICMBio (2018) Atlas dos Manguezais do Brasil. Instituto Chico Mendes de Conservação da Biodiversidade, Brasília
Jetz W, Sekercioglu CH, Böhning-Gaese K (2008) The worldwide variation in avian clutch size across species and space. PLoS Biol. https://doi.org/10.1371/journal.pbio.0060303
Johnsgard PA (2009) Birds of the great plains: family Ardeidae (herons and bitterns). In: Johnsgard PA (rev) Birds of the great plains. Digital Commons, Nebraska. http://digitalcommons.unl.edu/bioscibirdsgreatplains/12. Accessed 12 January 2021
Josens ML, Pretelli MG, Escalante AH (2009) Censos de aves acuáticas en sus colonias reproductivas en lagunas del sudeste de la provincia de Buenos Aires. Hornero 24(1):7–12
Jungsoo K, Tae-Hoe K (2007) Clutch size, reproductive success, and growth rate of Black-Crowned Night Herons Nycticorax nycticorax. Waterbirds. https://doi.org/10.1675/1524-4695(2007)030[0129:CSRSAG]2.0.CO;2
Kelly JP, Stralberg D, Etienne K, Mccaustland M (2008) Landscape influence on the quality of heron and egret colony sites. https://doi.org/10.1672/07-152.1. Wetlands
Kim J, Koo TH (2009) Nest site characteristics and reproductive parameters of grey herons Ardea cinerea in Korea. Zool Stud 489(5):657–664
Koczur LM, Ballard BM, Green MC, Hewitt DG, Henke SE (2018) Breeding ecology and habitat use of North America’s rarest Ardeidae: the Reddish Egret Egretta rufescens. Avian Conserv Ecol. https://doi.org/10.5751/ACE-01258-130210
Kushlan JA (1993) Colonial waterbirds as bioindicators of environmental change. Colon Waterbirds. https://doi.org/10.2307/1521444
Hancock JA, Kushlan JA (2010) The Heron Handbook, 2nd edn. Harper and Row, New York
Laake JL (2013) RMark: an R interface for analysis of capture-recapture data with MARK. https://apps-afsc.fisheries.noaa.gov/Publications/ProcRpt/PR2013-01.pdf. Accessed 22 September 2021
Lack D Ecological adaptations for breeding in birds. Chapman, LondonLindén M, Møller AP (1968) (1989) Cost of reproduction and covariation of life history traits in birds. Trends Ecol Evol. https://doi.org/10.1016/0169-5347(89)90101-8
Lovette IJ, Fitzpatrick JW (2016) Handbook of bird biology. John Wiley & Sons Ltd, Cornell Lab of Ornithology, Ithaca
Marini MA, Matos NO, Borges FJA, Silveira MB (2009) Reproductive biology of Elaenia cristata (Aves: Tyrannidae) in the Cerrado of Central Brazil. Neotrop Biol Conserv 4(1):3–12
Martin TE (1995) Avian life history evolution in relation to nest sites, nest predation, and food. Ecol Monogr. https://doi.org/10.2307/2937160
Martínez C, Miranda AC, Ruiz X (2020) Breeding biology and brood reduction of herons and Ibis in a northern brazilian mangrove swamp: eggs do not starve. https://doi.org/10.1675/063.043.0106. Waterbirds
Mellink E, Riojas-López ME (2008) Waterbirds (other than Laridae) nesting in the middle section of Laguna Cuyutlán, Colima, México. Rev Biol Trop 56(1):391–397
Metallaoui S, Dziri H, Bousseheba A, Heddam S, Chenchouni H (2020) Breeding ecology of the cattle egret (Bubulcus ibis) in Guerbes-Sanhadja wetlands of Algeria. Reg Stud Mar Sci. https://doi.org/10.1016/j.rsma.2019.100979
Mock DW (1987) Siblicide, parent-offspring conflict, and unequal parental investment by egrets and herons. Behav Ecol Sociobiol. https://doi.org/10.1007/BF00292177
Moss WW, Camin JH (1970) Nests parasitism, productivity and clutch size in Purple Martins. Science. https://doi.org/10.1126/science.168.3934.1000
Olguín P, Beltzer A, Giraudo A, Regner S, Juani M, Vianco M, Mariano E, Lorenzón R, Leon E (2015) Reproductive biology of Striated Heron (Butorides striata) in Argentina. https://doi.org/10.1675/063.038.0402. Waterbirds
Olmos F, Silva e Silva R (2002) Breeding biology of Litle Blue Heron (Egretta caerulea) in southeastern Brazil. Ornitol Neotrop 13:17–30
Parsons KC, Master TL (2000) Snowy Egret (Egretta thula), version 2.0. In: Poole AF, Gill FB (ed) The birds of North America. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bna.489. Accessed 5 February 2021
Perrins CM, Moss D (1975) Reproductive rate in the great tit. J Anim Ecol. https://doi.org/10.2307/3712
Petry MA, Fonseca VSDS (2005) Breeding success of the colonist species Bubulcus ibis (Linnaeus, 1758) and four native species. Acta Zool. https://doi.org/10.1111/j.1463-6395.2005.00200.x
QGIS.org (2022) QGIS Geographic Information System. QGIS Association. http://www.qgis.org. Accessed 5 February 2021
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. Accessed 20 September 2021
Rezende CE, Lacerda LD, Ovalle ARC, Silva LFF (2007) Dial organic carbon fluctuations in a mangrove tidal creek in Sepetiba bay, Southeast Brazil. Braz J Biol. https://doi.org/10.1590/S1519-69842007000400012
Ricklefs RE (1969) An analysis of nesting mortality in birds. Smithson Contrib Zool. https://doi.org/10.5479/si.00810282.9
Ricklefs RE (1980) Geographical variation in clutch size among passerine birds: Ashmole’s hypothesis. Auk. https://doi.org/10.1093/auk/97.1.38
Roper JJ (2005) Try and try again: nest predation favors persistence in a neotropical bird. Ornitol Neotrop 16:253–262
Roshnath R, Sinu PA (2017) Nesting tree characteristics of heronry birds of urban ecosystems in peninsular India: implications for habitat management. Curr Zool. https://doi.org/10.1093/cz/zox006
Ruiz-Guerra C, Echeverry-Galvis M (2019) Prey consumed by wading birds in mangrove swamps of the caribbean coast of Colombia. J Nat Hist. https://doi.org/10.1080/00222933.2019.1667037
Ryan PG (2018) Entanglement of birds in plastics and other synthetic materials. Mar Pollut Bull. https://doi.org/10.1016/j.marpolbul.2018.06.057
Schaeffer-Novelli Y (1995) Manguezal: ecossistema entre a terra e o mar. Caribbean Ecological Research, São Paulo
Scherer JFM, Scherer AL, Petry MV (2014) Vertical nest stratification of four heron species in southern Brazil. Stud Neotrop Fauna Environ. https://doi.org/10.1080/01650521.2014.921976
SEMEADS (2001) Bacias hidrográficas e recursos hídricos da macrorregião ambiental 2 – bacia da Baía de Sepetiba, vol 4. Secretaria de Meio Ambiente e Desenvolvimento Sustentável do Estado do Rio de Janeiro, Rio de Janeiro
Shirai T (2013) Colony development and density-dependent processes in breeding grey herons. Int J Zool. https://doi.org/10.1155/2013/404065
Sick H (1997) Ornitologia Brasileira. Nova Fronteira, Rio de Janeiro
Signorell A (2021) et mult al. DescTools: tools for descriptive statistics. R package version 0.99.47. https://cran.r-project.org/package=DescTools. Accessed 20 September 2021
Slagsvold T (1982) Clutch size in birds: the nest predation hypothesis. Oecologia. https://doi.org/10.1007/BF00378388
Soares MLG, Almeida PMM, Cavalcanti VF, Estrada GCD, Santos DMC (2011) Vulnerabilidade dos manguezais da região metropolitana do Rio de Janeiro face às mudanças climáticas. In: Nobre C, Young A (eds) Megacidades, vulnerabilidades e mudanças climáticas: região metropolitana do Rio de Janeiro. Instituto Pereira Passos, Rio de Janeiro, pp 258–289
Tavares DC, Guadagnin DL, Moura JF, Siciliano S, Merico A (2015) Environmental and anthropogenic factors structuring waterbird habitats of tropical coastal lagoons: implications for management. https://doi.org/10.1016/j.biocon.2015.02.027. Biol Conserv
Thompson DL, Ovenden TS, Pennycott T, Nager RG (2020) The prevalence and source of plastic incorporated into nests of five seabird species on a small offshore island. Mar Pollut Bull. https://doi.org/10.1016/j.marpolbul.2020.111076
Uzun A (2009) Do the height and location of black-crowned night Heron (Nycticorax nycticorax) nests affect egg production and breeding success? Waterbirds. https://doi.org/10.1675/063.032.0217
Weller MW (1999) Wetland birds: habitat resources and conservation implications. Cambridge University Press, New York
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer-Verlag, New York
Witteveen M, Brown M, Ryan PG (2017) Anthropogenic debris in the nests of kelp gulls in South Africa. https://doi.org/10.1016/j.marpolbul.2016.10.052. Mar Pollut Bull
Xiao H, Hu Y, Lang Z, Fang B, Guo W, Zhang QI, Pan X, Lu X (2017) How much do we know about the breeding biology of bird species in the world? J Avian Biol. https://doi.org/10.1111/jav.00934
Yorio P, Suárez N, Ibarra C, Gonzalez P, Canti S, Kasinsky T, Marinao C (2022) Anthropogenic debris in Kelp Gull and other seabird nests in northern Patagonia, Argentina. https://doi.org/10.1016/j.marpolbul.2022.113404. Mar Pollut Bull
Zhang M, Hong YM, Castley JG, Zou FS, Zhang Q, Fan HM, Sot-Chan, Leong KF (2018) Habitat features rather than competition explain the distribution and co-occurrence of Ardeidae in a highly urbanized landscape. https://doi.org/10.1675/063.041.0106. Waterbirds
Zhao Z, Li Q, Ni C, Zhang L (2017) Non-destructive bioindicator of little egret (Egretta Garzetta) to assess the pollution of highly toxic organic pollutants in poyang lake wetland. https://doi.org/10.1007/s13157-017-0978-1. Wetlands
Acknowledgements
We thank Denise M Braz for assistance with vegetation identification. James J. Roper provided additional constructive comments. Miguel  Marini revised a version of the manuscript.
Funding
This study was supported in part by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) (Financing Code 001) and by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (Proc. 170041/2018-2).
Author information
Authors and Affiliations
Contributions
The study design and fieldwork were performed by RBGC and IF. RBGC and TLS conducted the data analyses. The manuscript was written by RBGC and subsequently revised by all other authors.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Ethics Approval
The present study was carried out under permit from the ICMBio/SISBIO (license 26290-1).
Consent to Participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Cabral, R.B.G., da Silva, T.L. & Ferreira, I. Reproductive Biology of A Three-heron Mixed Colony in a Neotropical Mangrove Forest. Wetlands 43, 27 (2023). https://doi.org/10.1007/s13157-023-01677-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-023-01677-4