
Abstract
To evaluate soil organic carbon (SOC) and total nitrogen (TN) stocks of ash, alder and oak tree stands in Karacabey floodplain forest and adjacent Calabrian pine forest, grassland, cropland and sand dune in relation to soil depths (0-130 cm), a study in three replicate sites for each tree and the adjacent sites was carried out in northwest Turkey. The results indicated that among the tree species, alder stands had the greatest SOC (3.97%) and TN (0.328%) and total accumulation of SOC (405 Mg ha-1) and TN (34.4 Mg ha-1), followed by ash tree (3.11%, 0.302%, 393 Mg ha-1 and 26.2 Mg ha-1 respectively) and oak (2.43%, 0.220%, 293 Mg ha-1 and 28.6 Mg ha-1 respectively). However, the grassland showed the highest cumulative SOC densities within 0-130 cm depth (678 Mg ha-1) compared to the tree species. It also showed higher TN densities (27.5 Mg ha-1) than the ash tree and the Calabrian pine stands, whereas lower than the alder and oak stands. The sand dunes showed the lowest SOC and TN values. Compared to the soil depth of 0–30 cm, mean SOC and TN stored in 30–130 cm soil depth accounted for 58% and 40% in ash stands, 41% and 45% in alder stands, 52% and 56% in oak stands, 57% and 66% in Calabrian pine stands respectively. Thick alluvial soil and dry climate in the region could be responsible for the better root system development, and thus much higher SOC and TN stocks into deeper soil layers.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Increased atmospheric concentration of greenhouse gases (especially carbon dioxide) due to anthropogenic emissions is now widely acknowledged by the scientists as a major cause of climate change (Hertzberg and Schreuder 2016). It has been estimated that CO2 levels are rising at a rate of 2.0 ± 0.1 ppm per year in the last decade (IPCC 2014). The IPCC estimates atmospheric concentration of carbon dioxide will rise to between 540 and 940 ppm by the year 2100. Hence, it is inevitable to reduce the CO2 concentration in the atmosphere. Forests can play an important role in mitigating elevated atmospheric CO2 concentrations and preventing global warming since they are the largest carbon reservoir in terrestrial ecosystems (Schimel et al. 2001; Reichstein and Carvalhais 2019). For that reason, Annex I countries under the United Nations Framework Convention on Climate Change (UNFCCC) should report the national emissions and removal of greenhouse gases associated with the forest sector in the Agriculture, Forestry, and Other Land-Use (AFOLU) (IPCC 2006; UNFCCC COP 15 2009). Many developing countries, including Turkey, have reported on the carbon stocks from all carbon pools. Terrestrial forests ecosystems are mostly subject to many studies due to their important role in the global carbon (C) flux. Although it is estimated that wetlands cover approximately six to nine per cent of the Earth’s surface and contain about 35 per cent of global terrestrial carbon (Kolka et al. 2018), the role of wetlands in carbon sequestration and storage has generally been underestimated and not completely understood. There are still uncertainties about the overall carbon balance in wetland systems, about their existing carbon stocks and even about the global area of wetlands, which have been historically underappreciated. Since the early 1700s, it has been estimated that 87% of global wetland area has been lost (Davidson 2014). Land use change, pollution, water extraction, and landscape modification have threatened global wetland areas (van Asselen et al. 2013). A wetland’s ability to capture carbon can be undermined by such disturbances, but critically, those disturbances can result in microbial breakdown, demineralization and ultimately release of significant amounts of carbon that had already been stored (Atwood et al. 2017). It has been reported that conversion to agricultural land for cropping and grazing can lead to 80–96% reduction in wetland SOC (Sigua et al. 2009).
Some types of wetlands play a particularly important role as carbon stores. These include temperate and tropical peatlands, vegetated inter-tidal wetlands and forested wetlands (floodplain forests). It has been stated that floodplains (approximately 0.5–1% of the global land area) are responsible for a range of 0.5–8% of global SOC stocks (Naiman et al. 2005; Cierjacks et al. 2010; Sutfin et al. 2016; D’Elia et al. 2017).
Forests that grow on the floodplain are called “floodplain forests” to differentiate them from upland forests. About 60% of the wetlands are estimated to be the floodplain forests. Floodplain forests are Europe’s most threatened natural ecosystems. They are considered as ‘priority forest habitat type’ in the Annexe I of the European Habitats Directive. Floodplain forest ecosystems are host to very high diversity of plant species, including trees and shrubs, and home to a wide range of fauna (Daily 1997). Unfortunately, floodplain forests are getting less and less in Europe and 90% of their original area has disappeared and they are in critical condition (Hughes et al. 2003). Floodplain forest ecosystems serve a critical function in the global carbon (C) cycle due to their important role in C sink management relative to other terrestrial ecosystems (Cartisano et al. 2013; Nath et al. 2017). Despite their importance for ecosystem and human services, biomass storage and dynamics in floodplain forest ecosystems remain poorly understood (Melack et al. 2009). The flood regime, or flood pulse, is considered a major driver of spatial variability in C storage and productivity of flooded forests (Junk 1989).
Turkey has some floodplain forests in several regions, especially in the Marmara and Black Sea Regions, but unfortunately only 11,400 ha floodplain forests has remained in Turkey. There are several riverine and floodplain forests in the northern part of Turkey, some of which have already been studied in terms of ecology and biology (Pamay 1967; Çiçek 2002). However, there has been no study available in Turkey comparing soil organic carbon and nitrogen stocks of floodplain forest to surrounding terrestrial forests or to land use types under similar climate conditions. Quantification of the changes in pool size and fluxes of C and nitrogen is fundamental to the understanding of the effects of land-use change on the floodplain forest ecosystem functions. On the other hand, typical soil carbon (C) stocks used in global carbon models only account for the upper 30 cm meter of soil. However, in the literature, there have been evidences that deep floodplain soils may store substantial quantities of C (Sutfin et al. 2016; D’Elia et al. 2017). Especially, limiting carbon stock estimates to the upper soil profile of 0–30 cm vastly underestimates wetland carbon and nitrogen storage capacity. Here we assess deep soil C pools (0-130 cm) associated with an alluvial forested floodplain ecosystem, which is similar to soil depth (0-120 cm) studied in forested wetlands by a number of authors, for example Baties (2011) and Nahlik and Fennessy (2016). Therefore, we set up a detailed study in Bursa Karacabey floodplain forests to investigate the variations in SOC and TN contents and stocks: (1) among tree species in Karacabey floodplain forests composing of ash (Fraxinus angustifolia Vahl.), alder (Alnus glutinosa (L.) Gaertn and oak (Quercus cerris L.) species (2) between the floodplain forests and the surrounding terrestrial environments (Calabrian pine forest, grassland, cropland and sand dune); (3) between soil depths (0–10 cm, 10–20 cm, 20–30 cm, 30–60 cm, 60–100 cm and 100–130 cm).
Materials and Methods
Study Area
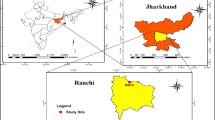
This study was carried out in Karacabey Floodplain Forests (40º23 × 38´´- 40º21 × 43´´N, 28º23 × 02´´- 28º34 × 21´´E) which is the third-largest wetland in Turkey (Fig. 1). Its neighboring Kocaçay Delta in the northwestern Bursa province is home to hundreds of flora and fauna that come alive with rich diversity in the spring and summer months. The Kocaçay Delta covers an area of 42,000 ha in the region. It is formed by the unification of the Susurluk River and Nilüfer Stream as they empty into the Marmara Sea. It has a great importance for the natural life since it has two shallow lagoons namely Dalyan and Arapçiftliği, large sand dunes, swamp, open areas (grasslands, croplands) and floodplain forests (Fig. 1).

The location of the research area (▲) according to the grid system of Turkey (Henderson, 1961), and land-use types (Akay et al. 2017)
The Karacabey floodplain ecosystem is formed by the accumulation of sediments deposited by creeks and streams flowing into the sea. The water level varies based on the rise and fall of groundwater during certain periods throughout the year. Total size of the Karacabey floodplain is approximately 3800 ha. (Akay et al. 2017). It includes a variety of habitats; sand dunes (623 ha), swamp (532 ha), lakes (760 ha), grasslands (390 ha), croplands (545 ha) and floodplain forests (950 ha). The floodplain forests are not only dependent on rainfall and air humidity but more on ground water. Ministry of Forestry and Water Affairs described the area as “the floodplain is similar to the mangrove forests of tropical regions”.
A semi humid climate is generally characteristics to the study region. According to previous year’s meteorological data (2007–2020), mean annual precipitation was 719 mm and mean temperature was 15.5 0 C. Although the most dominant tree species in the floodplain forests are ash (Fraxinus angustifolia Vahl.), alder (Alnus glutinosa (L.) Gaertn., oak (Quercus robur, Quercus cerris, Quercus pubescens), a wide variety of vegetation types is also reported in the Kocaçay Delta by Ursavaş and Keçeli (2018, 2019) mostly dominated with Quercus spp., Carpinus spp., Acer spp., Alnus glutinosa, Salix alba, Castanea sativa, Sambucus nigra, Hedera helix, Populus tremula., Cornus spp., Erica arborea, Ruscus aculeatus, Tilia tomentosa, Fraxinus spp., Pinus spp. Dozens of animal species call Karacabey floodplain forest home, from wild horses and boars to a range of waterfowl including flamingos, black storks and herons. The “flooded forest” attracts ecotourists to the region for bird watching, photography, nature walks and camping (URL-1).
Field Survey and Soil Sampling
Three sampling plots (20 m × 20 m) located approximately 300 m. apart were identified and sampled for three tree species (ash, alder and oak) in Karacabey floodplain forests, for one tree species (Calabrian pine) in the terrestrial forests as well as for grasslands, croplands and sand dunes (Fig. 2). Total subplots were 21.

Soil samples were collected from cropland (a), grassland (b), sand dunes (c), alder (d), ash tree (e), Calabrian pine (f) and oak stands
In the floodplain forest plots, three mature and taller trees in each plot were used to determine mean stand age, stand height and diameter at breast height. Annual growth ring in the trunk of the trees was counted to determine the tree age. A Blume-Leiss altimeter was used to measure the tree heights. A diameter tape was used to measure diameter at the breast height (DBH). Canopy cover was visually decided in each plot and then this determination was corrected by measurements of stem number and DBH. Some information about the studied plots in the floodplain forests is presented in Table 1.
Soil samples were collected in summer (July) 2019, when the soil had minimum moisture and the water table was at the lowest depth (150 cm). The soil samples were collected from 6 different soil depths (0–10 cm, 10–20 cm, 20–30 cm, 30–60 cm, 60–100 cm and 100–130 cm) using a soil auger (5-cm diameter). Composite soil samples were achieved by mixing the soil samples from the same layer in each plot.
A soil profile was also taken in each plot, and soil samples were collected along each profile for bulk density measurements at depths of 5 cm, 15 cm, 25 cm, 45 cm, 80 and 115 cm. All soil samples were placed into plastic bags and brought to the laboratory for chemical and physical analyses.
Analysis of Soil Samples
In order to remove stones, roots, large organic particles and macro fauna, moist soil samples from the field were dried under the laboratory conditions and then crushed by hand and sieved using a less than 2 mm stainless soil sieve. After that they were bulked to give a single representative soil sample for each soil depth.
Soil pH was determined by a combination glass electrode in H2O (soil-solution ratio 1:2.5) (Gülçür 1974). Electrical conductivity (EC) was determined in 1:1 soil water extract by using conductivity meter and expressed as dS/m (Allen 1989). Soil organic matter was determined by the modified Walkley-Black method as described by Kalra and Maynard (1991). Soil texture was determined by Bouyoucos’ hydrometer method (Bouyoucos 1962). Soil bulk density was determined by the undisturbed core sampling method (Black 1965). Percent total pore space was computed from the values of bulk density (BD) and particle density (PD) (assuming a particle density of 2.65 g cm− 3) as described by Hillel (2004). Soil bulk density was used to calculate soil organic C and N stocks.
A CNH-S elementary analyser (Eurovector EA3000-Single) was used to determine mean SOC and TN in the soil samples (Vesterdal and Raulund-Rasmussen 1998). Volume, bulk density, soil carbon and nitrogen content of the soil were used to calculate the SOC and TN pools as Mega gram C or N in per hectare (Mg / ha) (Lee et al. 2009). Dry mass of the soil was found as follows:
Dry soil mass (Mi) = bulk density (BDi) x thickness of the soil depth (Ti) x 104.
Soil organic carbon or nitrogen stock (kg C or N ha-1) in the soil depth was found as follows:
C or N mass to the soil depth (i) = Carbon or nitrogen concentration x Mi.
Statistical Analysis
One-way ANOVA was used to analyze for differences in SOC and TN contents and stocks among the sites (the four tree species, grassland, cropland and sand dune) and among the six soil depths using the SPSS® (software v. 11). For variables whose ANOVA results differed, the Tukey’s mean separation test was performed at a significance level of α = 0.05.
Results
General Soil Properties
Mean values of soil pH, electrical conductivity, moisture, porosity, bulk density, organic matter and soil texture (clay, silt and sand) from four tree species (alder, ash tree, oak and Calabrian pine) and from four land use types (forest, grassland, cropland and sand dune) in relation to soil depths are shown in Table 2. All soil properties varied significantly according to the land use types. However, only electrical conductivity, soil organic matter and bulk density varied significantly with the soil depths (Table 3). For all land use types, electrical conductivity, porosity and soil organic matter decreased with increasing soil depths, whereas soil bulk density increased (Table 2).
It was noted that soils in the study sites were alkaline and vary from the forest soils of other sites. Three dominant soil texture were seen as sandy clay, sandy loam and sandy clay loam. The Calabrian pine site had the lowest soil pH (7.98), while the alder site had the highest soil pH (8.78). Electrical conductivity and moisture were also lowest in the Calabrian pine site (0.061 dSm− 1 and 8% respectively), while they were highest in the ash tree soil (1.365 dSm− 1 and 56%). The sand dune site showed the lowest soil organic matter and porosity (0.67% and 28% respectively), whereas it showed the highest bulk density (1.94 g cm− 3). The ash tree site had the highest organic matter (8.66%) and porosity (56%), but lowest soil bulk density (1.26 g cm− 3). The lowest clay (5%) and silt (7%) were noted in the sand dune site, whereas it had the highest sand (89%). The highest clay (28%) was seen in the ash tree site, while the Calabrian pine site had the highest silt (34%) and the lowest sand (46%).
Mean SOC and TN Content
The main effects of the land use types and soil depths on the SOC and TN contents were all significant. Land use type x soil depth interaction was also significant for the SOC and TN contents indicating that it behaved in different ways according to soil depth on different land use types (Table 3).
Among the four tree species, the SOC and TN contents in the alder site were highest (3.97% and 0.328% respectively) followed by the ash tree site (3.11% and 0.302% respectively), the oak site (2.43% and 0.220% respectively), and the Calabrian pine site (1.01% and 0.087% respectively), within 0-130 cm soil depth.
The grassland site showed lower SOC content (2.79%) than the alder and ash tree sites, but higher than the oak and Calabrian pine site. The cropland site also had higher SOC content (1.70%) than the Calabrian pine site, but lower SOC content than the other three tree sites (alder, ash tree and oak). Both the grassland and cropland sites showed higher TN content (0.210 and 0.155% respectively) than the Calabrian pine site, but lower TN content than the other three tree sites. The sand dune site had the lowest SOC (0.53%) and TN (0.022%) content compared to all three tree species, grassland and cropland sites.
Mean SOC content decreased with the soil depth in the three tree sites, the grassland and the sand dunes sites, and the greater content was in the 0–30 cm topsoil (Fig. 3). The distribution of SOC content along the soil profile in the cropland site was, however, stable at the first three soil depths (0–30 cm), and then it sharply decreased in the 30–130 cm. Below 30 cm, mean SOC in the cropland did not show any variation with the soil depth (Fig. 3).

Mean SOC content decreased with the soil depth in the forest stands. Alder stands had the highest SOC followed by ash tree, oak and Calabrian pine stands. Top soil (0–30 cm) had 3- to 4-fold more C content than deeper soil (30–130 cm). The bars are standard deviations, means with the same letters do not differ at alpha = 0.05)
The SOC contents in the first 0–30 cm depth averaged 6.76% in the alder site, 5.39% in the ash tree site, 3.32% in the oak site, 1.53% in the Calabrian pine site, 4.03% in the grassland site, 2.58% in the cropland site and 0.73% in the sand dune site.
A decrease of TN contents from the upsoil to the deeper soil along the profile followed the pattern of SOC (Fig. 4). As noted for the SOC, the distribution of TN in the cropland site was also only stable at the first three soil depths (0–30 cm) and then sharply decreased and stabilized in the bottom soil layers (60–130 cm). The greater TN was also in the upsoil of 0–30 cm. The TN contents in the first 0–30 cm depth averaged 0.539% in the alder site, 0.507% in the ash tree site, 0.292% in the oak site, 0.117% in the Calabrian pine site, 0.315% in the grassland site, 0.237% in the cropland site and 0.031% in the sand dune site.

Similar to mean SOC content, total nitrogen (TN) also decreased with the soil depth in the forest stands. This time, ash tree had the highest TN followed by alder, oak and Calabrian pine. Top soil (0–30 cm) also had 2- to 3-fold more N content than deeper soil (30–130 cm). The bars are standard deviations, means with the same letters do not differ at alpha = 0.05)
Overall, soil C/N ratios in the four forest tree species decreased with the soil depths (Fig. 5). This result was correlated with much higher SOC within the topsoil layers in the forest tree species. There were significant variations in soil carbon to nitrogen ratios among the four trees along the soil profile. In general, within 0–30 cm, the alder and Calabrian pine sites had higher C/N ratio than the oak and ash tree sites. Within 30–100 cm, the alder and oak sites showed higher C/N ratio than the Calabrian pine and the ash tree sites. As seen within 0–30 cm, the alder and Calabrian pine sites had higher C/N ratio than the oak and ash tree sites, within 100–130 cm.

The C–N ratios expressed as SOC/TN decreased with the soil depth in the forest stands. With exception of top soil (0–20 cm) and deep soil (130 cm), alder and oak showed higher C–N ratios than ash tree and Calabrian pine
In the cropland site, the SOC/TN ratio was relatively stable along the profile in the cropland site, while it showed a slight increase to stable level along the soil profile in the grassland site (Fig. 5).
Mean SOC and TN Stocks
The main effects of the land-use types and soil depths on the SOC and TN stocks were all significant. Land use type x soil depth interaction was not significant for the SOC and TN contents indicating that it behaved in similar way according to soil depth on different land-use types (Table 3).
Among the four tree species, the cumulative SOC densities over area within 0-130 cm depth were highest in the alder site (405 Mg ha-1), followed by ash tree site (393 Mg ha-1), oak forest (293 Mg ha-1) and the Calabrian pine site (162 Mg ha-1) (Fig. 6a). The trend of cumulative TN density was very consistent with SOC among the four tree species. The alder site had the highest TN (34.4 Mg ha-1), followed by oak site (28.6 Mg ha-1), ash tree site (26.2 Mg ha-1) and the Calabrian pine site (15.9 Mg ha-1) (Fig. 6b). The differences in SOC and TN stocks among the four tree species were generally significant at each soil depth (Fig. 6ab).

Among the three floodplain tree species, the cumulative SOC and TN densities over area within 0-130 cm depth were highest in the alder, followed by ash tree and oak. Among the land use types, Calabrian pine and sand dunes had lower cumulative SOC and TN densities than the grassland and the floodplain tree species. The error bars indicate the standard deviations of the means
The grassland site showed the highest cumulative SOC densities within 0-130 cm depth (678 Mg ha-1) compared to the three tree sites (Fig. 6a). It also showed higher TN densities (27.5 Mg ha-1) than the ash tree and the Calabrian pine sites, but lower than the alder and oak sites (Fig. 6b). The cropland site had higher cumulative SOC and TN densities (259 Mg ha-1 and 23.2 Mg ha-1 respectively) than the Calabrian pine site, but lower densities than the other three tree sites and the grassland site. The sand dune site had the lowest SOC (63 Mg ha-1) and TN (2.53 Mg ha-1) densities compared to all tree species, grassland and cropland sites.
The percentage contributions of the SOC and TN stocks in the 0-30-cm, 30–60 cm, 60–100 cm and 100–130 cm soil layer are shown in Fig. 7. Among the four tree species, the upper 30 cm alone contributed 59%, 48%, 43% and 42% for SOC in the alder, oak, ash tree and Calabrian pine sites respectively, while 60%, 55%, 44% and 34% for TN in the ash tree, alder, oak and Calabrian pine sites respectively. As for the grassland, cropland and sand dune sites, the upper 30 cm alone contributed 28%, 49% and 68% for SOC respectively, while 57%, 51% and 72% for TN respectively.

The percentage contributions of the SOC and TN stocks continually decreased with the soil depths. Among the four tree species, the contribution of upper 30 cm alone varied from 42% (Calabrian pine) to 59% (alder) for SOC and from 34% (Calabrian pine) to 60% for TN (ash tree). The error bars indicate the standard deviations of the means
The percentage contributions of the SOC and TN stocks in the alder and oak sites and the sand dune site continually decreased with the soil depths (Fig. 7ab). Within 30–60 cm, the contribution percentages also sharply decreased in the other tree species, the grassland and the cropland sites. However, below 60 cm, the contribution percentages were either stable or slightly increased in ash tree, the Calabrian pine the grassland and the cropland sites (Fig. 7ab).
Mean SOC, TN and stocks had a strong relationship with soil properties (Table 4), except for soil pH, soil texture. Mean SOC exhibited a positive significant relation with total N, porosity, SOM, electrical conductivity and soil moisture but showed negative correlation with soil bulk density.
Discussion
The results from this study indicated that soil properties (physical and chemical) analysed in this study significantly differed among the four land use types (forest land, grassland, cropland and sand dune) and between the forest tree species (ash tree, alder, oak and Calabrian pine) in Karacabey floodplain forests. Soil properties also showed significant differences with soil depths except for soil pH, moisture and soil textures (clay, silt and sand) (Table 3). Land use type x soil depth interaction was only significant for the porosity, SOC and TN contents indicating that they behaved in different ways according to soil depth on different land use types.
Effects of Soil Depth on Soil Properties
In general, the soil properties including porosity, soil organic matter, soil organic carbon and total nitrogen contents and stocks decreased with increasing soil depths, whereas only soil bulk density increased (Table 2). As stated in many studies, less organic matter and weight of the overlying horizons can mostly result in higher soil bulk density in the inner soil layers (Grüneberg et al. 2014). In this study, mean SOC, TN and stocks showed a decrease with soil depth with more content near the soil surfaces. It seemed that the availability of more organic matter from aboveground vegetation (trees, grasses, bushes) contributed to have more SOC, TN and stocks in the topsoil. Especially, forest trees can increase root turnover and continuously provide plant litters in the upper layers (Kimmins 2004), which enhance the SOC and TN (Wu et al. 1993). Similar result was also noted by Soleimani et al. (2019) which is in agreements with our results.
Effects of Land Use Type on General Soil Properties
Several researchers have examined the soil physical and chemical properties in various terrestrial landscapes such as upland forests, grasslands, and agricultural areas (Lepcha and Devi 2020; Francaviglia et al. 2017; Zhang et al. 2014). In Turkey, most studies showed a significant difference in the soil general properties (for example pH, texture, moisture, bulk density) with land-use type (Sariyildiz et al. 2016; Göl and Yılmaz, 2017; Kucuk et al. 2019), but the others found no significant variations in soil properties with land-use types (Evrendilek et al. 2004; Korkonc 2014). However, studies focusing on forested wetland landscapes are limited. Under the forested wetland landscapes, our results indicated a significant difference in the soil general properties with land-use type (Table 3).
We found significant variations in the soil general properties among the three floodplain forest trees (alder, ash tree and oak). In general, alder and ash tree species showed similar soil properties compared to oak tree which had lower soil properties except for the bulk density and sand content (Table 2). However, compared the floodplain forest tree species to the upland tree species (the Calabrian pine site) and other land use types (grassland, cropland and sand dune sites), the three floodplain tree species clearly had higher pH, EC, moisture, organic matter, porosity and sand, but lower silt than the upland Calabrian pine forest (Table 2). Soil bulk density was lower in alder and ash tree site than that in oak and Calabrian pine site, while clay content was higher in ash tree and Calabrian pine sites than that in alder and oak sites. Similar results were reported by Tecimen and Kavgaci (2010) who studied some soil and forest floor characteristics of floodplain forest, thermophile forest and sand dune at Igneada floodplain forest in Turkey. They found that the floodplain forests had higher sand (64%), clay (20.1%), organic carbon (5.619%) and total nitrogen (0.213%) than the thermophile forest (52.4%, 12.6%, 4.191%, 0.154% respectively), while it had lower soil bulk density (907.8 g l-1) than that in the thermophile forest (970.7 g l-1). The sand dunes had the highest sand content (91%) and BD (1257 g l-1), while it showed the lowest clay (5.1%), organic carbon (0.478%) and total nitrogen (0.062%).
The upland Calabrian pine site also showed lower general soil properties compared to the grassland and cropland sites except for silt content which was higher in the Calabrian pine site. On the other hand, the lowest organic matter, porosity, clay and silt contents were noted in the sand dune sites, whereas it had the highest bulk density and sand content.
The differences in soil bulk density of the land use types in this study could be attributed to the variation in the soil texture in the study sites (Dumig et al. 2006; Zhang et al. 2014; Francaviglia et al. 2017; Lepcha and Devi 2020). Additionally, compaction of the cropland soils by the continuous use of machinery could further contribute to the higher bulk density of subsoil. Loss of organic matter and a decline of soil aggregation by the cultivation were reported as causes of the increased bulk density (Lal 1987).
Effects of Land Use Type on SOC and TN Contents and Stocks
Several researchers have studied the soil capacity to store OC and TN in various terrestrial environments (e.g., forests, prairies, and farmland) (Kondo et al. 2017; Don et al. 2007) and measured their variability based on land use types (Wiesmeier et al. 2015; Rodríguez-Murillo 2001). Most studies have generally focused on the changes in SOC and TN of the topsoil layer (0–30 cm), which store the highest SOC and the greatest microbial activity (Umrit et al. 2014; Hao et al. 2015). On the other hand, inconsistent results for the influence of land-use change on SOC and TN have been previously reported (Binkley et al. 2004; Specht and West 2003). Conversion from natural forest or perennial grassland to agricultural land was reported to decrease SOC by 20–43% (Wei et al. 2014). Similarly, conversion of forest to grassland induce large variation in SOC dynamics, leading to carbon losses of 10–55% (Wei et al. 2014; Perrin et al. 2014; Yang et al. 2015).
Forested wetlands are usually not considered when assessing opportunities for managing ecosystems to enhance terrestrial C and N storage. For example, nationally in Turkey, soil organic carbon stocks have been estimated to be 55.68 Mg C ha-1 for forests, 49.77 Mg C ha-1 for grassland, 35,96 Mg C ha-1 for cropland and 49,71 Mg C ha-1 for wetlands (Agriculture and Forest Ministry Soil Organic Carbon Project, 2008). This approach assumes that wetland forests do not have substantially different soil organic carbon than terrestrial forests. However, this present study has shown that the floodplain forests and adjacent grassland sites or even cropland site generated from the floodplain forest can store more carbon stocks in soils than the terrestrial forest ecosystems.
We measured and accounted for deeper SOC and TN in this study. Accounting for the SOC and TN stocks of deeper soil layers in our study represents this ecosystem service that forested wetlands provide. Especially, limiting carbon stock estimates to the upper soil profile of 0–30 cm vastly underestimates wetland carbon and nitrogen storage capacity. Hansen and Nestlerode (2014) reflected this in their study where they reported soil carbon densities to a depth of 10–15 cm in the Gulf of Mexico coastal region of 34–47 Mg C ha− 1. However, by assessing soils to 120 cm, Nahlik and Fennessy (2016) showed for the coastal areas that SOC was greater than 340 ± 94 Mg C ha− 1. Batjes (2011) reported for the forested wetland sites under native vegetation (0 to 120 cm depth) that SOC stocks were 135 Mg ha-1 and 74 Mg ha-1 for warm temperate-moist and –dry climate regions respectively. We found that the highest SOC stocks to a depth of 130 cm was in the grassland site, followed by alder > ash tree > oak > cropland > Calabrian pine > sand dune sites (Fig. 5a). The highest percent of TN stocks was found in alder site, followed by oak > grassland > ash tree > cropland > Calabrian pine > sand dune sites (Fig. 5b). In general, the higher SOC and TN under the floodplain forest trees and grassland can be attributed to the addition of organic matter to the surface soil from above biomass and fine root density of naturally grown grasses and shrubs in grasslands or animal grazing (Wickland et al. 2013). Grazing on the open areas within the floodplain forests is widespread (Liu et al. 2009; Oates et al. 2008; Yao et al. 2010; Wickland et al. 2013). Our results show that grassland (natural/fallow) is more beneficial to surface OC storage than the terrestrial forest, forested wetlands or cultivated cropland. Previous studies (Lugo and Brown 1993; Yeasmin et al. 2020; Jin et al. 2014; Don et al. 2011; Chen et al. 2007; Guo and Gifford 2002) also revealed similar results with much higher SOC under tropical grassland than the adjacent forests. Tate et al. (2000) reported that OC storage in grassland soil was 13% higher than in forest and crop land soil. A review by Conant et al. (2001) stated that SOC increased nearly 70% with the transformation of native rain forests to grassland. On the other hand, our results have shown that conversion of the floodplain forest to the cropland resulted in a significant reduction in SOC and TN stock (alder site 405 Mg C ha− 1 and 34.4 Mg N ha− 1 → cropland site 259 Mg C ha− 1 and 23.2 Mg N ha− 1).
Higher total N in soils of the alder site could be related to the nitrogen-fixing ability of this tree species. Rothe et al. (2002) showed that soil total nitrogen content was increased with the presence of nitrogen fixing species. Higher SOC and stocks in the floodplain forests of our study are in agreement with the previous studies which showed that forested wetland areas, for example riparian zones, can significantly store more SOC than near terrestrial forests (Kern 1994; Davis et al. 2004; Ricker et al. 2013). The upper 1.30 cm of floodplain forest soils from our study contained 405 Mg C ha− 1 while the average upland SOC pool was only 162 Mg C ha− 1 (Fig. 5a). Floodplain forest ecosystems are more humid and distinctively biodiverse than the adjacent Calabrian pine forest. Soil moisture and soil clay showed a positive correlation with SOC in our study indicating that wet and fine textured soils typically contain more organic carbon. Additionally, in the study area, it can be seen that organic matters on the upslopes can be delivered by hillslope erosion and debris flows, which bury organic matter under mineral sediment in the floodplain forests. A number of researchers also stated that in riparian areas, the mosaic of organic carbon distribution is due to the erosional and depositional disturbances, and partly through the redistribution of litter and POM by fluvial processes (Pinay et al. 2002; McClain et al. 2003; Hall et al. 2009; Ramos Scharrón et al. 2012).
Conclusion
The results of the present study from Karacabey floodplain region have shown that land use types, tree species, and soil depth significantly vary soil organic carbon and total nitrogen stocks. Floodplain forest ecosystems with thick litter layers and naturally grown grasses and shrubs in grasslands with the fine root density tend to increase SOC and TN, and result in storing more carbon and nitrogen and also provide better soil health and fertility for the restoration. However, soil fertility decreases in the sand dune sites due to less organic matter inputs, and in the cropland sites due to intensive land management practices. The results have also revealed that relative to the conventional soil carbon and nitrogen studies of 30-cm depth in terrestrial forest ecosystems, alluvial hydric floodplain soils contain significantly higher SOC and TN at the depths of 30 to 130 cm than the adjacent non hydric floodplain soils. These results indicate that deeper SOC and TN storage capacity of floodplain forests should be taken into an account when floodplain forest C and N stocks are estimated. As shown in our study, below soil layers of the floodplain forests can have substantial quantities of C and so the restoration and protection of floodplain forests should promote active C sequestration in the region.
Data Availability
All data produced from this study are available from Temel Sariyildiz (temel.sariyildiz@btu.edu.tr) on reasonable request.
Code Availability
Not applicable.
References
Akay AE, Gencal B, Taş İ (2017) Spatiotemporal change detection using landsat imagery: the case study of Karacabey flooded forest, Bursa, Turkey. ISPRS Annals of the Photogrammetry, Remote Sensing and Spatial Information Sciences, Volume IV-4/W4, 4th International GeoAdvances Workshop, 14–15 October, Safranbolu, Karabuk, Turkey
Allen SE (1989) Chemical Analysis of Ecological Materials. Blackwell Scientific Publications, Oxford
Atwood TB, Connolly RM, Almahasheer H, Carnell PE, Duarte CM, Lewis CJE et al (2017) Global patterns in mangrove soil carbon stocks and losses. Nat Clim Change 7:523–528. https://doi.org/10.1038/nclimate3326
Batjes NH (2011) Soil organic carbon stocks under native vegetation - Revised estimates for use with the simple assessment option of the Carbon Benefits Project system. Agric Ecosyst Environ 142(3–4):365–373. https://doi.org/10.1016/j.agee.2011.06.007
Binkley D, Kaye J, Barry M, Michael GR (2004) First rotation changes in soil carbon and nitrogen in a Eucalyptus plantation in Hawaii. Soil Sci Soc Am J 68:1713–1719. https://doi.org/10.2136/sssaj2004.1713
Black CA (1965) Methods of soil analysis. Am Soc Agron 1:1570–1572
Bouyoucos GJ (1962) Hydrometer method improved for making particle size analysis of soils. Agron J 54:464–465
Cartisano R, Mattioli W, Corona P, Mugnozza GS, Sabatti M, Ferrari B (2013) Assessing and mapping biomass potential productivity from poplar dominated riparian forests: A case study. Biomass Bioenerg 54(7):293–302. https://doi.org/10.1016/j.biombioe.2012.10.023
Chen L, Gong J, Fu B, Huang Z, Huang Y, Gui L (2007) Effect of land use conversion on soil organic carbon sequestration in the loess hilly area, loess plateau of China. Ecol Res 22(4):641–648. https://doi.org/10.1007/s11284-006-0065-1
Cierjacks A, Kleinschmit B, Babinsky M, Kleinschroth F, Markert A, Menzel M, Ziechmann U, Schiller T, Graf M, Lang F (2010) Carbon stocks of soil and vegetation on Danubian floodplains. J Plant Nutr Soil Sci 173:644–653. https://doi.org/10.1002/jpln.200900209
Conant RT, Paustian K, Elliott ET (2001) Grassland management and conversion into grassland: Effects on soil carbon. Ecol Appl 11:343–355. https://doi.org/10.1890/1051-0761
Çiçek E (2002) Stand structures and necessary silvicultural treatments on bottomland forest of Süleymaniye-Adapazari. Dissertation, University of İstanbul
Daily GC (1997) Nature’s Services: Societal Dependence on Natural Ecosystems. Island, Washington, DC
Davidson NC (2014) How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar Freshw Res 65:934–941. https://doi.org/10.1071/MF14173
Davis AA, Stolt MH, Compton JE (2004) Spatial distribution of soil carbon in southern New England hardwood forest landscapes. Soil Sci Soc Am J 68:895–903. https://doi.org/10.2136/sssaj2004.8950
D’Elia AH, Liles GC, Viers JH, Smart DR (2017) Deep carbon storage potential of buried floodplain soils. Sci Rep 7:8181. https://doi.org/10.1038/s41598-017-06494-4
Don A, Schumacher J, Scherer-Lorenzen M, Scholten T, Schulze E-D (2007) Spatial and vertical variation of soil carbon at two grassland sites: Implications for measuring soil carbon stocks. Geoderma 141:272–282. https://doi.org/10.1016/j.geoderma.2007.06.003
Don A, Schumacher J, Freibauer A (2011) Impact of tropical land-use change on soil organic carbon stocks - a meta-analysis. Global Change Biol 17(4):1658–1670. https://doi.org/10.1111/j.1365-2486.2010.02336.x
Dumig A, Schad P, Kohok M, Beyerlein P, Schwimmer W, Koger-Knabner I (2006) A mosaic of nonallophanic Andosols, Umbrisols and Cambisols on rhyodacite in the southern Brazilian highlands. Geoderma 145:158–173. https://doi.org/10.1016/j.geoderma.2008.01.013
Evrendilek F, Celik I, Kilic S (2004) Changes in soil organic carbon and other physical soil properties along adjacent Mediterranean forest, grassland, and cropland ecosystems in Turkey. J Arid Environ 59:743–752. https://doi.org/10.1016/j.jaridenv.2004.03.002
Francaviglia R, Renzi G, Doro L, Parras-Alcantara L, Lozano-Garcia B, Ledda L (2017) Soil sampling approaches in Mediterranean agro-ecosystems. Influence on soil organic carbon stocks. CATENA 158:113–120. https://doi.org/10.1016/j.catena.2017.06.014
Hall R, Baker M, Arp C, Koch B (2009) Hydrologic control of nitrogen removal, storage and export in a mountain stream. Limnol Oceanogr 54:2128–2142. https://doi.org/10.4319/lo.2009.54.6.2128
Hansen VD, Nestlerode JA (2014) Carbon sequestration in wetland soils of the northern Gulf of Mexico coastal region. Wetl Ecol Manag 22:289–303. https://doi.org/10.1007/s11273-013-9330-6
Hao S, Zhou P, Zhang YZ, Kuzyakov Y, Zhou Q, Tida G et al (2015) Loss of labile organic carbon from subsoil due to land-use changes in subtropical China. Soil Biol Biochem 88:148–157. https://doi.org/10.1016/j.soilbio.2015.05.015
Hertzberg M, Schreuder H (2016) Role of atmospheric carbon dioxide in climate change. Energy Environ 27(6–7):785–797. https://www.jstor.org/stable/90006767
Hillel D (2004) Introduction to Environmental Soil Physics. Elsevier Academic Press
Hughes F, Richards K, Girel J, Moss T, Muller E, Nilsson C, Rood S (2003) The Flooded Forest: Guidance for Policy Makers and River Managers in Europe on the Restoration of Floodplain Forests. The FLOBAR2 Project. Department of Geography, University of Cambridge, Cambridge
Göl C, Yılmaz H (2017) The effect of land use type / land cover and aspect on soil properties at the Gökdere catchment in northwestern Turkey. Å umarski list 141(9–10):459–468. https://doi.org/10.31298/sl.141.9-10.2
Grüneberg E, Ziche D, Wellbrock N (2014) Organic Carbon Stocks and Sequestration Rates of Forest Soils in Germany. Global Change Biol 20:2644–2662. https://doi.org/10.1111/gcb.12558
Gülçür F (1974) Physical and chemical analyzing methods of soil properties. University of İstanbul. Issue No: 201 İstanbul, Turkey
Guo LB, Gifford RM (2002) Soil carbon stocks and land use change: a meta analysis. Global Change Biol 8(4):345–360. https://doi.org/10.1046/j.1354-1013.2002.00486.x
IPCC (2006) IPCC Guidelines for National Greenhouse Gas Inventory. Hayama, Japan, IPCC/IGES
IPCC (2014) In: Pachauri RK, Meyer LA (eds) Climate Change 2014. Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team. IPCC, Geneva, Switzerland, p 151
Jin Z, Dong Y, Wang Y et al (2014) Natural vegetation restoration is more beneficial to soil surface organic and inorganic carbon sequestration than tree plantation on the Loess Plateau of China. Sci Total Environ 485–486:615–623. https://doi.org/10.1016/j.scitotenv.2014.03.105
Junk WJ (1989) Flood tolerance and tree distribution in Central Amazonian floodplains. In: Nielsen LB, Nielsen IC, Balslev H (eds) Tropical Forests: Botanical Dynamics, Speciation and Diversity. Academic Press, London
Kalra YP, Maynard DG (1991) Methods Manual for Forest Soil and Plant Analysis. Forestry Canada, Northern Forestry Publications. Alberta, Canada
Kern JS (1994) Spatial patterns of soil organic carbon in the contiguous United States. Soil Sci Soc Am J 58:439–455. https://doi.org/10.2136/sssaj1994.03615995005800020029x
Kimmins JP (2004) Forest ecology: a foundation for sustainable forest management and environment ethics in forestry, 3rd edn. Prentice Hall, Upper Saddle River, NJ
Kolka R, Trettin C, Tang W, Krauss K, Bansal S, Drexler J, Wickland K, Chimner R, Hogan D, Pindilli EJ, Benscoter B, Tangen B, Kane E, Bridgham S, Richardson C (2018) : Chap. 13: Terrestrial wetlands. In Second State of the Carbon Cycle Report (SOCCR2): A Sustained Assessment Report [Cavallaro N, Shrestha G, Birdsey R, Mayes MA, Najjar RG, Reed SC, Romero-Lankao P, Zhu Z (eds.)]. U.S. Global Change Research Program, Washington, DC, USA, pp. 507–567, https://doi.org/10.7930/SOCCR2.2018.Ch13
Kondo M, Saitohb TM, Satoa H, Ichiia K (2017) Comprehensive synthesis of spatial variability in carbon flux across monsoon Asian forests. Agric For Meteorol 232:623–634. https://doi.org/10.1016/j.agrformet.2016.10.020
Korkonc SY (2014) Effect of afforestation on soil organic carbon and other soil properties. CATENA 123:62–66. https://doi.org/10.1016/j.catena.2014.07.009
Kucuk M, Yener I, Duman A (2019) Effects of vegetation cover/land use and slope aspect on surface soil properties near the copper smelter factory in Murgul, Turkey. Appl Ecol Environ Res 17(5):12305–12321. https://doi.org/10.15666/aeer/1705_1230512321
Lal R (1987) Tropical Ecology and Physical Edaphology. John Willy and Sons, New York, USA
Lepcha NT, Devi NB (2020) Effect of land use, season, and soil depth on soil microbial biomass carbon of Eastern Himalayas. Ecol Process 9:65. https://doi.org/10.1186/s13717-020-00269-y
Lee J, Hopmans JW, Rolston DE, Baer SG, Six J (2009) Determining soil carbon stock changes: Simple bulk density corrections fail. Agric Ecosyst Environ 134:251–256. https://doi.org/10.1016/j.agee.2009.07.006
Liu C, Hoist J, Yao Z, Bruggemann N, Butterbach-Bahl K, Han S, Han X, Tas B, Susenbeth A, Zheng X (2009) Growing season methane budget of an Inner Mongolian steppe. Atmos Environ 43(19):3086–3095. https://doi.org/10.1016/j.atmosenv.2009.03.014
Lugo AE, Brown S (1993) Management of tropical soils as sinks or sources of atmospheric carbon. Plant Soil 149(1):27–41. https://doi.org/10.1007/BF00010760
McClain ME, Boyer EW, Dent CL, Gergel SE, Grimm NB, Groffman PM, Hart SC, Harvey JW, Johnston CA, Mayorga E, McDowell WH, Pinay G (2003) Biogeochemical hot spots and hot moments at the interface of terrestrial and aquatic. Ecosyst 6:301–312. https://doi.org/10.1007/s10021-003-0161-9
Melack JM, Novo EMLM, Forsberg BR, Piedade MT, Maurice L (2009) Floodplain ecosystem processes. In: Keller M, Bustamante MG, Dias PS (eds) Amazonia and global change. American Geophysical Union, Washington, D.C., pp 525–541
Nahlik AM, Fennessy MS (2016) Carbon storage in US wetlands. Nat Commun 7:13835. https://doi.org/10.1038/ncomms13835
Naiman RJ, Decamps H, McClain ME, Likens GE (2005) Riparian Ecology, Conservation, and Management of Streamside Communities. Academic Press, Burlington, MA
Nath S, Nath AJ, Sileshi GW, Das AK (2017) Biomass stocks and carbon storage in Barringtonia acutangula floodplain forests in North East India. Biomass Bioenergy 98:37–42. https://doi.org/10.1016/j.biombioe.2017.01.014
Oates LG, Jackson ARD, Allen-Diaz B (2008) Grazing removal decreases the magnitude of methane and the variability of nitrous oxide emissions from spring-fed wetlands of a California oak savanna. Wetlands Ecol Manage 16:395–404. https://doi.org/10.1007/s11273-007-9076-0
Pamay B (1967) Studies on silvicultural analysis of Demirköy-İğneada floodplain forests and silvicultural measures to be taken to make them efficient. Publication by General Directorate of Forestry. (in Turkish) No:43, İstanbul
Perrin AS, Fujisaki K, Petitjean C, Sarrazin M, Godet M, Garric B et al (2014) Conversion of forest to agriculture in Amazonia with the chop-and-mulch method: Does it improve the soil carbon stock? Agric Ecosyst Environ 184:101–114. https://doi.org/10.1016/j.agee.2013.11.009
Pinay G, Clément JC, Naiman RJ (2002) Basic principles and ecological consequences of changing water regime for nitrogen cycling in fluvial systems. Environ Manage 30:481–491. https://doi.org/10.1007/s00267-002-2736-1
Ramos Scharrón CE, Castellanos EJ, Restrepo C (2012) The transfer of modern organic carbon by landslide activity in tropical montane ecosystems. J Geophys Res Biogeosci 117(3):1–18. https://doi.org/10.1029/2011JG001838
Reichstein M, Carvalhais N (2019) Aspects of Forest Biomass in the Earth System: Its Role and Major Unknowns. Surv Geophys 40:693–707. https://doi.org/10.1007/s10712-019-09551-x
Ricker MC, Stolt MH, Donohue SW, Blazejewski GA, Zavada MS (2013) Soil organic carbon pools in riparian landscapes of southern New England. Soil Sci Soc Am J 77:1070–1079. https://doi.org/10.2136/sssaj2012.0297
Rodríguez-Murillo JC (2001) Organic carbon content under different types of land use and soil in Peninsular Spain. Biol Fertil Soils 33:53–61. https://doi.org/10.1007/s003740000289
Rothe A, Cromack K Jr, Resh SC, Makineci E, Son Y (2002) Soil carbon and nitrogen changes under Douglas-fir with and without red alder. Soil Sci Soc Am J 66:1988–1995. https://doi.org/10.2136/sssaj2002.1988
Sariyildiz T, Savacı G, Kravkaz İS (2016) Effects of tree species, stand age and land-use change on soil carbon and nitrogen stock rates in northwest of Turkey. iForest - Biogeosc For 9:165–170. https://doi.org/10.3832/ifor1567-008
Schimel DS, House JI, Hibbard KA et al (2001) Recent patterns and mechanisms of carbon exchange by terrestrial ecosystems. Nature 414:169–172. https://doi.org/10.1038/35102500
Sigua GC, Coleman SW, Albano J (2009) Beef cattle pasture to wetland reconversion: impact on soil organic carbon and phosphorus dynamics. Ecol Eng 35:1231–1236. https://doi.org/10.1016/j.ecoleng.2009.05.004
Soleimani A, Hosseini SM, Bavani ARM, Jafari M, Francaviglia R (2019) Influence of land use and land cover change on soil organic carbon and microbial activity in the forest of northern Iran. CATENA 177:227–237. https://doi.org/10.1016/j.catena.2019.02.018
Specht A, West PW (2003) Estimation of biomass and sequestered carbon on farm forest plantations in northern New South Wales, Australia. Biomass Bioenergy 25:363–379. https://doi.org/10.1016/S0961-9534(03)00050-3
Sutfin NA, Wohl EE, Dwire KA (2016) Banking carbon: a review of organic carbon storage and physical factors influencing retention in floodplains and riparian ecosystems. Earth Surf. Proc. Land. 41:38–60. https://doi.org/10.1002/esp.3857
Tate KR, Scott NA, Ross DJ, Parshotam A, Claydon JJ (2000) Plant effects on soil carbon storage and turnover in a montane beech (Nothofagus) forest and adjacent tussock grassland in New Zealand. Soil Res 38(3):685–698. https://doi.org/10.1071/SR99092
Tecimen HB, Kavgacı A (2010) Comparison of soil and forest floor properties of floodplain and surrounding forests in Igneada, Turkey. J Environ Biol 31:129–134
Umrit G, Cheong RN, Gillabel J, Merckx R (2014) Effect of conventional versus mechanized sugarcane cropping systems on soil organic carbon stocks and labile carbon pools in Mauritius as revealed by 13 C natural abundance. Plant Soil 379:177–192. https://doi.org/10.1007/s11104-014-2053-5
UNFCCC COP 15 (2009) Available online: https://unfccc.int/process-and-meetings/conferences/past-conferences/copenhagen-climate-change%20conference-december-2009/cop-15. Accessed on 18 December 2021
Ursavaş S, Keçeli T (2018) Weissia multicapsularis, a rare moss species new to Turkey and Asia. Plant Biosyst 153(5):669–672. https://doi.org/10.1080/11263504.2018.1536086
Ursavaş S, Keçeli T (2019) The Moss Flora of Kocaçay Delta (Karacabey-Bursa) Floodplain Forests in Turkey. Anatol Bryol 5(1):22–34. https://doi.org/10.26672/anatolianbryology.517253
URL-1 :https://www.dailysabah.com/gallery/life/turkeys-karacabey-wetland%20home-to-rich-biodiversity
van Asselen S, Verburg PH, Vermaat JE, Janse JH (2013) Drivers of wetland conversion: a global meta-analysis. PLoS ONE 8(11):e81292. https://doi.org/10.1371/journal.pone.0081292
Vesterdal L, Raulund-Rasmussen K (1998) Forest floor chemistry under seven tree species along a soil fertility gradient. Can J For Res 28:1636–1647. https://doi.org/10.1139/x98-140
Wei XR, Shao MA, Gale W, Li LH (2014) Global pattern of soil carbon losses due to the conversion of forests to agricultural land. Sci Rep 4:4062. https://doi.org/10.1038/srep04062
Wickland KP, Krusche AV, Kolka RK, Kishimoto-Mo AW, Chimner R, Ogle SM, Srivastava N(2013) Inland wetland mineral soils. 2013 Supplement to the 2006 IPCC guidelines for national greenhouse gas inventories: wetlands, 5.1–5.34. https://digitalcommons.mtu.edu/michigantech-p/514
Wiesmeier M, Munro S, Barthold F, Steffens M, Schad P, Kögel-Knabner I (2015) Carbon storage capacity of semi-arid grassland soils and sequestration potentials in northern China. Glob Chang Biol 21:3836–3845. https://doi.org/10.1111/gcb.12957
Wu J, Brookes PC, Jenkinson DS (1993) Formation and destruction of microbialbiomass during decomposition of glucose and ryegrass in soil. Soil Biol Biochem 25:1435–1441. https://doi.org/10.1016/0038-0717(93)90058-J
Yang SH, Zhang HT, Chen JY (2015) The spatial variability of soil organic carbonin plain-hills transition belt and its environmental impact. China Environ Sci 35:3728–3736 (in Chinese)
Yao Z, Wolf B, Chen W, Butterbach-Bahl K, Brüggemann N, Wiesmeier M, Dannenmann M, Blank, B, Zheng X (2010) Spatial variability of N2O, CH4 and CO2 fluxes within the Xilin River catchment of Inner Mongolia, China: a soil core study. Plant Soil 331: 341-359. https://doi.org/10.1007/s11104-009-0257-x
Yeasmin S, Jahan E, Molla A, Islam AKMM, Anwar P, Rashid HO, Chungopast S (2020) Effect of Land Use on Organic Carbon Storage Potential of Soils with Contrasting Native Organic Matter Content. Int J Agron Volume 2020 Article ID 8042961:9pages. https://doi.org/10.1155/2020/8042961
Zhang L, Zhao R, Xie Z (2014) Response of soil properties and C dynamics to land-use change in the west of Loess Plateau. Soil Sci Plant Nutr 60(4):586–597. https://doi.org/10.1080/00380768.2014.922407
Acknowledgements
We would like to thank:
The Scientific and Technological Research Council of Turkey (TUBITAK) for its financial support to carry out this research.
General Directorate of Forestry, Department of International Relations, Education and Research for granting permission to work in Karacabey Floodplain Forest protected by the International Ramsar Agreement.
Funding
This research was funded by The Scientific and Technological Research Council of Turkey (TUBITAK) with the project number 119O263.
Author information
Authors and Affiliations
Contributions
T.S. designed the study, T.S., D.O.A., S.P. and M.T. collected soil samples; S.P., D.O.A. and M.T. measured stand characteristics; D.O.A. and M.T. prepared the soil samples for analysis. T.S. and D.O.A. prepared the tables and figures. T.S. wrote the manuscript. All the authors contributed to editing and agreed on the final version.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
All authors have read this manuscript and consent to its publication.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sariyildiz, T., Aygun, D.O., Parlak, S. et al. Effects of Land Use Types and soil Depths on Soil Organic Carbon and Total Nitrogen Stocks of Karacabey Floodplain Forests in Northwest Turkey. Wetlands 42, 102 (2022). https://doi.org/10.1007/s13157-022-01612-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-022-01612-z