
Abstract
Wetlands in mountains are highly dynamic and provide ecosystem services to human wellbeing. Understanding temporal and spatial wetland dynamics is crucial for successful management. This paper presents the reconstruction of a mire evolution at a high-altitude Andean valley in central-western Argentina, in subtropical South America (30°-36° S), during the Mid- and Late Holocene. The research is based on sedimentological and pollen analysis from a sedimentary section of 3.2 m thick exposed at the El Peñón valley. The record begins with an outwash environment after ca. 5700 cal. yrs BP associated with Pteridophytes dominance, followed by the development of a mire environment after ca. 3700 cal. yrs BP associated with Cyperaceae dominance. The environmental and vegetation changes are hypothesised to have occurred in response to a shift from cold to warmer conditions. Colder environments may have occurred again between 800 until < 600 cal. yrs BP, probably associated with the Little Ice Age. Tephra inputs are evidenced in the record from 1200 years BP onwards. However, the results do not show any conclusive evidence about the impact of volcanism in the dynamics of the wetland. High Amaranthaceae proportions would evidence human activities in the high-altitude valleys of the southern Andes, probably for the last 250 cal. yrs BP. This work permitted us to infer the evolution of the El Peñón wetland under multiple concurrent forcing factors from the Mid- Holocene onwards at different temporal scales, i.e. climate during the last 5700 yrs, and volcanism and anthropogenic impacts during the last millennium.
Resumen
Los humedales de montaña son altamente dinámicos y proveen servicios ecosistémicos a la humanidad. La comprensión de la dinámica temporal y espacial de los humedales es crucial para un manejo exitoso. Este trabajo presenta la reconstrucción de la evolución de una vega de montaña durante el Holoceno Medio y Tardío, en un valle andino del centro-oeste de Argentina, región subtropical de América del Sur (30°-36° S). La investigación está basada en los análisis sedimentológico y polínico de una sección sedimentaria de 3,2 m de potencia, expuesta en el valle El Peñón. El registro comienza con un ambiente de planicie glacifluvial posterior a los 5700 años cal. AP, asociado con dominancia de Pteridofitas, seguido por el desarrollo de un ambiente de vega luego de los 3700 años cal. AP, asociado con dominancia de Cyperaceae. Los cambios ambientales y de vegetación habrían ocurrido en respuesta a un cambio de condiciones frías a más cálidas. Condiciones más frías habrían ocurrido nuevamente entre los 800 y < 600 años cal. AP, asociadas probablemente con la Pequeña Edad de Hielo. Se observaron depósitos de cenizas volcánicas en el registro a partir de los 1200 cal. años AP. Sin embargo, los resultados no muestran evidencia concluyente sobre los impactos del volcanismo en la dinámica del humedal. Altas proporciones de Amaranthaceae se vincularían con actividades humanas en estos valles montañosos de Los Andes, probablemente en los últimos 250 cal. años AP. Este trabajo nos permitió inferir la evolución del humedal en el valle El Peñón bajo múltiples y concurrentes factores ambientales desde el Holoceno Medio en adelante en diferentes escalas temporales, i.e. el clima durante los últimos 5700 años, y el volcanismo y el impacto antrópico durante el último milenio.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mountain landscapes are highly dynamic because of an active morphogenesis in response to autogenic and/or allogenic forcing (e.g. tectonisms, volcanism and/or climatic change). Lakes and wetlands, including peatlands/mires, are part of that mountain dynamics and will become widespread as mountain glaciated areas recede (http://www.mountainwaterfutures.ca/wetlands.html, last visited August 2020). Wetlands are considered biodiversity cores and hydrological regulators because of water-quality improvement, flood abatement and carbon management, and are suppliers of ecosystem services contributing to human wellbeing (Malvárez, 1999; Tooth et al., 2015; Clarkson et al., 2013). Despite all these positive aspects, nearly 50% of the wetland areas have been lost worldwide and the condition of remaining wetlands is declining (Clarkson et al., 2013). In such a context, Grenfell et al. (2019) stated that the understanding of the temporal and spatial scales of wetland dynamics is crucial to successful wetland management and restoration. Besides, these authors indicate that long-term landscape processes provide a suitable base with which hydrology interacts, allowing wetland formation.
In the high Andes of central Argentina, in subtropical South America, wetlands are scarce (< 6% of the region) and are developed mostly as high altitude mires and/or muddy environments (barreales) (Benzaquén et al., 2017). The current knowledge about the long-term evolution –centennial and millennial timescales- of these wetland environments is deficient. During the last decades, research work has been carried out in the central-western region of Argentina to detect and analyse Late Quaternary archives that allow disentangling the palaeoenvironmental evolution of fluvial systems and the history of vegetation in central-western Argentina (e.g. Mehl and Zárate, 2012; Mehl and Zárate, 2014; Navarro et al., 2012 and references therein; Rojo et al., 2012, 2018). However, only few studies have been conducted in high Andean peatbogs/mire environments, i.e. Salina 2 and Salado peatbog sites (Markgraf, 1983) and El Peñón valley mire (Espizua, 2005; Espizua and Pitte, 2009). Salina 2 and Salado sites document wetter and colder conditions than present previous to 3000 yrs. BP; then, similar-to-present conditions were established. At El Peñón valley, three Holocene glacier advances were inferred based on geomorphologic and radiocarbon chronologic controls from mire deposits (Espizua 2005; Espizua and Pitte, 2009); two of them inferred between 5000 and 2000 yrs. BP and the youngest one, linked, according to the authors, to the Little Ice Age (LIA).
This paper presents sedimentological and pollen analyses conducted at a sedimentary section (Vega El Peñón -VEP- section site) that records the last 5700 cal. yrs in the El Peñón valley. The aim is to reconstruct the Mid- and Late Holocene environmental evolution of a high-mountain wetland, in a sensitive climatic setting with an active morphodynamics. In this research we assess how sensitive the wetland has been to environmental factors, e.g. climate change, volcanism and / or anthropogenic impacts, and which of them have been the main drivers of the wetland evolution. To address these questions, we firstly present the geological and environmental contexts of the study area. Then, we report the results of the chronology, and sedimentological and pollen records. And finally, we propose a scheme of wetland evolution in which the palaeoenvironmental inferences as well as the scope and limitations of the studied records are discussed.
Study Area
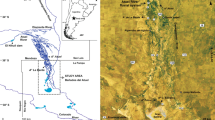
The El Peñón valley (35°15’ S, 70°31’ W) is in the high Andes cordillera (Mendoza province, Argentina) close to the present-day-active Peteroa-El Planchón volcanic complex (Fig. 1) and in the headwaters of the Valenzuela River basin. It is a valley of ~ 3.9 km2 that records glacier variations related to Mid-Holocene neoglacial advances (El Peñón and Peteroa moraines) and to the Little Ice Age (LIA; Amarilla I, II and III lateral moraines set) (Espizua, 2005; Espizua and Pitte, 2009). Currently there is a two ice-tongued glacier of 3.2 km long (Espizua and Pitte, 2009). Besides, an alluvial fan has developed at the lower valley and most of the valley bottom is covered by a mire of soligenous type reaching up to 9 m thick in the upper valley area (Figs. 1, 2a-b). The mire is supported by melting water from ice and snow that promote groundwater circulation and runoff at the valley. The studied section rests inside the latero-frontal moraine arc of the El Peñón moraine and ca. 800 ms upwards from its terminus. It is exposed at the banks of a perennial stream running along a flat floored U shape gully carved on the mire surface (Fig. 2).

adapted from Bruniard (1982). Yellow star: location of the studied sedimentary section; yellow dashed line: Argentina—Chile international border; yellow solid line: contour lines
Location of the study area. El Peñón study site is located in a glacial valley nowadays covered by mire vegetation (red colour in a Sentinel-2 scene from 06–01-2018, RGB 843 bands combination). South America Arid Diagonal location

El Peñón valley. a) View of the upper part of the valley. b) VEP sedimentary section at a gully on the northern margin of the valley. El Peñón moraine is behind
Moisture supply to the area comes from the Pacific Ocean dominantly as winter precipitation as it is located to the west of the South American Arid Diagonal defined by Bruniard (1982) (Fig. 1). More than 80% of the annual precipitation in the region (Viale and Nuñez, 2011) corresponds to snowfalls brought by the extratropical Westerlies with a decreasing gradient to the east (Garreaud et al., 2009; Viale et al., 2019). The average precipitation is ~ 900 mm/yr (Norte, 2000). Mean annual temperature is ~ 10–15° C, with a maximum of 15–20° C (January) and a minimum of ~ 10° C (July) (Hoffman, 1975).
In the study area the Altoandino phytogeographical province (Cabrera, 1976; Méndez, 2011) develops between circa 4500—2300 m a.s.l. and the Patagónica province below 2300 m a.s.l.. Regional Altoandino vegetation is characterised by grassy and shrubby steppes with cushion plants adapted to strong winds, low air temperatures and high solar radiation. Grasslands dominate the Andes mountains and valley’s slopes, with species like Poa ligularis, P. holciformis, Stipa speciosa and several taxa of genus Festuca, associated to other taxa such as Chuquiraga oppositifolia, Lecanophora ameghinoi, Junellia scoparia, Oxalis compacta and Azorella lycopodioides, among others (Cabrera, 1976; Méndez, 2011; Roig, 1972). At higher altitudes, Altoandino grasses dominate as shrubs and other herbs diversity decrease because of harsh environmental conditions (Méndez, 2011). The genus Adesmia is other typical shrubby element of this flora related to Altoandino vegetation, with species such as A. pinifolia, A. schneideri and A. obovata (Morici et al. 2010; Roig et al., 2000). Below 2,300 m a.s.l., the lowland valley is characterised by a grassy steppe associated with shrubs of Patagónica affinity such as Mulinum spinosum, Senna arnottiana, Neosparton aphyllum and Stillingia patagonica. Other common taxa are Schinus fasciculatus, Berberis grevilleana, Junellia juniperina, Ephedra breana, Colliguaja integerrima, Acaena splendens, and different species of the Asteraceae family (Baccharis, Senecio, Nassauvia, Gutierrezia) (Cabrera, 1976; Méndez, 2011; Roig, 1972; Roig et al. 2000). Nowadays, mires in the region are mainly composed of sedges (Cyperaceae), rushes (Juncaceae) and humid-adapted grasses (Poaceae) in the upper and middle areas of valleys (Méndez 2007; Ruthsatz et al. 2020). Other taxa in the mires include Calceolaria filicaulis (ssp. luxurians), Draba australis, and Werneria pygmaea among others. Taxa such as Calandrinia and Arenaria grow in mires but also out of them in rocky and/or moist soils along with Altoandino and Patagónica plant communities (Méndez 2007; Peralta, 1985; Wingenroth, 2012; Zuloaga and Morrone, 1999a, 1999b).
The Valenzuela River basin has been inhabited by hunter-gatherers since ca. 10,000 yrs BP (Salgán et al. 2015; Durán et al. 2016) and pastoral activities, based on European taxa such as goats and sheeps, have been developed in the region in the last 250 years (Durán, 2000).
Materials and Methods
Field work (austral summer 2014) in the El Peñón glacial valley comprised the description of a 3.2 m-thick sedimentary succession (VEP section; 35º14′52.8’’ S, 70º30′13.6″ W, 2456 m. a.s.l.) (Figs. 1, 2b) exposed at a gully, including qualitative grain-size, thickness of deposits, limits, colour in dry samples according to Munsell Color chart (2000), sedimentary structures, geometry and relationships (lateral and vertical) of sedimentary units. Sediment sampling was conducted at intervals according to the main noticeable changes in the described properties (e.g. 10 cm in average).
Grain-size analyses on 30 sediment samples were done with a Malvern Mastersize Hydro 2000 μm laser-based diffractometer (detection range 2000 μm—0.010 μm). Cementing and/or agglutinating substances were previously removed by soaking samples in hydrogen peroxide and 15% hydrochloric acid solutions, rinsed every time with distilled water. Then, sediments were classified following Folk’s proposal (1970). Total organic carbon (TOC) quantification was conducted by the loss-on-ignition technique (LOI) according to standardised procedures (Heiri et al., 2001) and applying a 50% factor of conversion as the amount of organic matter determined by LOI at 550 °C is twice the organic carbon content (Dean, 1999). TOC content was expressed as dry weight percentage.
Sedimentary environment interpretation was made based on different authors’ approaches and concepts (e.g. Bridge and Demicco, 2008; Reading, 1996; Nichols, 2009; Lindsay 2016, 2018); current mire geomorphological features were described following Bower (1960, 1961) and Evans and Warburton (2007).
Sampling for pollen analysis was done at 39 different depth levels and samples stored in two-fold plastic bags. At the laboratory, samples were processed for concentration of pollen grains by standard techniques (Faegri and Iversen, 1989), including hot alkali (KOH; to disaggregate sediment and eliminate humic acids), hot hydrochloric acid (HCl), hydrofluoric acid (HF), and acetolysis mixture. Aliquots of samples were mounted on slices and counted with an optical microscope Nikon Eclipse E200. For the identification of pollen types, the Pollen Reference Collection of the Laboratorio de Palinología del Museo de Historia Natural de San Rafael, and specific atlases (Markgraf and D’Antoni, 1978; Heusser, 1971; Wingenroth and Heusser, 1984) were used. For the pollen sum, a minimum of 200 regional pollen types were counted, excluding Cyperaceae and Pteridophytes as representative of local taxa as well as Nothofagus as extra-regional pollen. A pollen diagram, with taxa expressed as percentages of the pollen sum, was produced with Tilia software (Grimm, 2007) as well as a stratigraphically constrained cluster analysis, using the Edwards and Cavalli-Sforza’s chord distance coefficient of dissimilarity, including only regional terrestrial taxa to facilitate the description and discussion of pollen results.
Three bulk sediment samples from the VEP section were sent for radiocarbon dating in the absence of visible plant macrofossils; organic matter from those bulk sediment samples varied in TOC content from ~ 0.87 to ~ 19%. Previously, Espizua (2005) reported a Mid—Late Holocene interval on another sedimentary section at this mire. Here, reported ages were obtained by accelerator mass spectrometry (AMS) at the NFS-Arizona Laboratory (USA). Calibration of radiocarbon dates and age-depth modelling (linear interpolation between dated levels) were performed using the R clam package (Blaauw, 2010) and the Southern Hemisphere calibration data set (Hogg et al., 2013). The age-depth model was built with the three radiocarbon dates here reported and two radiocarbon dates previously published by Durán et al. (2016) from the same sedimentary section. The segment between 120 and 145 cm was not considered to build the age model because we assumed it was the result of an almost instantaneous deposition (glacio-fluvial deposit).
Results
Chronology and TOC
The sedimentary organic matter content at the VEP section site was radiocarbon dated at 5691 (5336–5917) cal. yrs BP at a depth of 290 cm depth, and at 1488 (1356–1691) and 1221 (993–1362) cal. yrs BP at 210 cm and 200 cm depth, respectively (Table 1). Two youngest ages, 607 (549–667) and 594 (480–723) cal. yrs BP, were obtained from bulk sedimentary organic matter at 160 cm and 115 cm depth (Table 1). The age-depth model performed (Fig. 3) is valid from 300 to 115 cm depth. The topmost sediment layers of the sequence (115–0 cm) werenot considered in the age-depth model because no radiocarbon ages are available and the uppermost level (0 cm) could not be considered as the present-time. This latter uncertainty is because the occurrence of a gully incision, sometime after ca. 600 yrs. BP (633 ± 47 14C yrs. BP).

Age-depth model of VEP sedimentary section. The slump (excised between 145 and 116 cm) corresponds to a fluvio-glacial deposit. Horizontal grey lines indicate tephra layers (T1-T5). Ages after the youngest radiocarbon date (594 cal. yrs BP) are not considered reliable because of gully incision (see the text for explanation)
The TOC content exhibits variable values along the analysed section (Fig. 4b). TOC content is lower (i.e. < 3.68%) both in the bottom (3.2—2.17 m depth) and upper (1.47 – 0 m depth) parts of the section. However, the intermediate section records a trend of increasing TOC content that begins at a depth of 2.10 m, going from 9% up to 19%.

VEP sedimentary section. a) Sedimentary section and calibrated weighted average ages. b) TOC % derived from LOI. c) View of the VEP sedimentary section; the five calibrated radiocarbon dates in this study are indicated
Sedimentary Section
The analysed section records between 3.2 m to 2.25 m depth very coarse silty fine sand sediments (Fig. 4a, b). An irregular but continuous oxidation limit is observed at the top of this deposit indicating water table fluctuations; at the time field work was conducted deposits were waterlogged at 3.10 m depth. Sandy matrix gravel deposits underlie the silty sand deposit.
From 2.25 m to 1.95 m depth, there is a fine sandy very coarse silt laminated layer with clear and smooth limits (Fig. 4a, b). Lamination is marked by alternated millimetre-thick dark brown and greyish brown sediment laminas. Upwards, there is a silty deposit (ca. 25-cm-thick), with very coarse silty fine sand at the lower part and a tephra (ash; grain size < 2 mm) layer (T1) of 2–3 cm-thick interbedded at ~ 200 cm depth (Fig. 4a).
From 1.95 m to 1.45 m, sandy very coarse silt and silty fine sand layers with common Fe-oxide specks are covered by a ~ 2 cm-thick volcanic ash layer (T2; 165 cm depth). On top, there is a laminated deposit of very coarse silty fine sand (165–150 cm depth) with common bioturbations and Fe-oxide specks (3% to ~ 15% in the upper part); an interbedded volcanic ash layer of 5–8 cm-thick (T3; ~ 155 cm depth), including scarce lapilli (grain size > 2 mm), is present (Fig. 4a). In turn, this deposit is covered, with a clear and smooth contact, by a medium silt fine layer, with common oxidised roots (150–145 cm depth) and common Fe-oxide specks at the uppermost part.
From 1.45 m to 1.20 m depth, a matrix- supported conglomerate (fine to medium pebbles in a rusty coarse sandy to very fine pebble matrix) is overlayed by a centimetre-laminated sandy very coarse silt deposit with common Fe-oxide in the upper part (Fig. 4b); a light-colour volcanic ash layer of ~ 2 cm-thick (T4; 110 cm depth) with lapilli is interbedded.
From 1 to 0 m depth, the sedimentary deposits include loose silty fine and coarse sand with variable colours. A 13-cm-thick layer of volcanic ash (T5) is interbedded at a depth of 77 cm. Some levels, (e.g. at 30 cm and 50 cm depth), exhibit common Fe-oxidation marks and minuscule vegetal rests. The topmost part (20 cm) presents loose poorly sorted sand (Fig. 4a) with abundant roots from the vegetation in the current mire environment.
Recorded Pollen
The pollen content preserved in the VEP section site encompassing the last 5700 cal. yrs reflects vegetation communities in the mire, inside the valley, almost exclusively represented by Cyperaceae and Pteridophytes. It also records Altoandino grasslands from the surrounding slopes mainly represented by Poaceae and shrubs and herbs such as Asteraceae family, Ephedra, Adesmia, Calandrinia, Arenaria, Apiaceae, Brassicaceae, Malvaceae, Rosaceae, Amaranthaceae, Wendtia, Nassauvia, and Oxalidaceae (Fig. 5). Other taxa such as Mutisiae, Schinus, Senna, Lycium, Colliguaja, Rhamnaceae, Gomphrena, and Proteaceae are present in few samples with very low abundances and do not show major changes in the pollen record. A similar record is shown by extra-regional pollen taxa such as arboreal Maytenus, Nothofagus, Podocarpaceae, Drimys and Myrtaceae as they are expressed with very low values these taxa do not show major changes throughout the pollen zones. They indicate the influence of extra-regional vegetation because of wind long transportation.

Pollen diagram from VEP sedimentary section. Cluster analysis (right) is based on regional terrestrial taxa (yellow, green and orange colours). Depth and age scales are indicated in the vertical axis (left). Red horizontal lines indicate T1-T5 tephras
Four pollen assemblage zones, 1 to 4 from bottom to top, were identified by cluster analysis. They are dominated by Poaceae, associated mainly with Cyperaceae and Pteridophytes as secondary taxa in abundance, and with minor proportions of Asteraceae Subf. Asteroideae, Nassauvia, Calandrinia, Amaranthaceae, Arenaria, Asteraceae Subf. Cichorioideae, Oxalidaceae, Wendtia, and Ephedra (Fig. 5).
Zone 1 (290—253 cm; 5500—3700 cal. yrs BP)
It is characterised by Poaceae (70–80%) and by a maximum content of Pteridophytes (including Azolla, 10–15%). Also, Asteraceae Subf. Cichorioideae and Ephedra are present and associated with their highest values in the VEP section (Fig. 5). Other pollen taxa are present in traces, i.e. Nassauvia, Asteraceae Subf. Asteroideae, Adesmia, Arenaria, Apiaceae, Malvaceae, and Cyperaceae.
Zone 2 (253 – 173 cm; 3700—800 cal. yrs BP)
Poaceae keeps high proportions (70%) but Asteraceae Subf. Cichorioideae and Ephedra decline. Calandrinia appears and reaches its highest values in the profile (Fig. 5). Arenaria also registers its maximum values in the profile at the top of the zone. Nassauvia maintains low values as in the previous zone, but Asteraceae Subf. Asteroideae increases (< 10%) in the middle of the zone and then decreases abruptly to traces to the upper part of the zone. Brassicaceae (including Menonvillea) appear with low values (< 3%). Oxalidaceae appear and register trace values. Cyperaceae increases gradually until reaching its highest proportions (60%) to the top of the zone (Fig. 5). On the contrary, Pteridophytes decrease consistently through the zone and are only present in trace amounts at the zone's upper boundary.
Zone 3 (173—80 cm; 800—< 600 cal. yrs BP)
It shows a slight increase in Poaceae (80%) and a sharp depletion in Calandrinia and Arenaria. Nassauvia and Asteraceae Subf. Asteroideae increases a little after the middle of the zone, and then diminishes to the top. Asteraceae Subf. Cichorioideae, Ephedra, and Apiaceae register traces. Similarly, Adesmia appears in traces but restricted to the middle of the zone. Brassicaceae and Malvaceae continue with very low abundances and disappear to the top. Although with fluctuating proportions, Cyperaceae shows a trend of decreasing values (from 60 to 40%) to the top of this zone (Fig. 5). Pteridophytes record traces in all this zone.
Zone 4 (80—0 cm; < 600 cal. yrs BP)
Poaceae decreases to its lowest values (60%) along the VEP section, and Calandrinia, Oxalidaceae, and Amaranthaceae reach up to 10%. Brassicaceae and Rosaceae show peaks at the base of the zone and then disappear to the top. Nassauvia and Wendtia also increase from the middle part to the top of the zone. Arenaria continues with low percentages as previously. Ephedra only registers traces. Ateraceae Subf. Cichorioideae, Malvaceae and Phacelia appear in traces to the top of the zone. Cyperaceae show fluctuating values between 20 and 40%, and Pteridophytes increase subtly from the middle to the top of the zone (Fig. 5).
Discussion
Mid- to Late Holocene Wetland Evolution Inferred at the El Peñón Valley
The sedimentary succession at the VEP section site records at least three phases of wetland evolution during the Mid- to Late Holocene. A first phase (~ 5700 yrs. BP—~ 3700 yrs. BP) is recorded at the lower part of the Sect. (3–῀2.5 m) by sandy glacio-fluvial deposits covered by silty coarse sandy fluvial sediments of a likely outwash environment in which shallow streams and water bodies may have developed. The highest abundance of Pteridophytes registered in this phase (Pollen zone 1, Fig. 5) could mainly be the result of taxa as Azolla spp. that live floating in shallow waters (Arana et al. 2011) associated with the outwash environment. Other taxa such as Cheilanthes spp., currently recorded outside the valley's floor growing between rocks´ fissures (Wingenroth, 1992), could also have contributed to some extent to the pollen record. By this time, Espizua (2005) proposed a neoglacial advance (Mid- Holocene) recorded throughout the El Peñón moraine. However, Rodbell et al. (2009) questioned Espizua's (2005) proposal by stating that numerical ages for the El Peñón moraine, as well as those from moraines at the neighbouring El Azufre glacial valley (Espizua, 2005), are minimum-limiting ages which bolster the notion of a region-wide Mid- Holocene glacial advance. They considered it conceivable to expect a Lateglacial age for the oldest moraine in each valley because of the lack of maximum-limiting age constraints.
The vegetation of surrounding slopes was dominated by the Poaceae family. Probably, the Altoandino grassland, dominated today by grasses taxa (Poa holciformis), was the main contributor to the pollen record (Fig. 5). However, some local influence of humid-adapted grasses should not be discarded. Other few herbs and shrubs are present in low values (Asteroideae and Cichorioideae subfamilies, and Ephedra) or traces (Nassauvia, Apiaceae, Arenaria, and Malvaceae). This low plant diversity and Poaceae dominance is interpreted as harsh environmental conditions as those that predominate at higher altitudes nowadays. Ephedra pollen, from a gymnosperm wind-pollinated taxon from Patagónica vegetation at lower altitudes, would have reached the area by long-distance wind transportation, as it was suggested by Markgraf (1983) in similar depositional environments of the region.
Then, a second phase (~ 3700 yrs. BP—~ 594 yrs. BP) in the evolution of the VEP section site corresponds to the onset of mire sedimentation. Thus, the middle and upper part of the VEP section (῀2.5 m–0 m, Fig. 4a) is featured by mire deposits accumulated in a low-energy, shallow environment, interpreted from intermingled fine sand, sandy silty and silt layers with TOC content reaching up to the highest values of the record (18.9%). Levels with relatively higher TOC content indicate organic matter accumulation under redox conditions favouring its preservation at the mire environment. This geomorphic situation probably favoured the gradual but major change in vegetation, as indicated in the pollen record. Cyperaceae appeared at ~ 3700 and started to replace Pteridophytes slightly but with a steady increase until ~ 1300 cal. yrs BP. Then, after a sharper rise, they dominate completely between 1300 and 800 cal. yrs BP.
The regional grassland would have not changed significantly in this phase as Poaceae declined subtly in this lapse from 80 to 70% (Pollen zone 2). At the same time, a major representation of Patagónica-Altoandino taxa (e.g., such as Calandrinia and Arenaria), concomitantly, with Ephedra and Asteraceae Subf. Cichorioideae declination, would indicate higher presence of shrubs in the area.
After 607 cal. yrs BP and before 594 cal. yrs BP mire sedimentation was interrupted, at this portion of the valley, by a reactivation of fluvial aggradation with a higher energy stream able to transport and accumulate a sandy gravel load (matrix- supported conglomerate deposit, section depth of 120–145 cm). This event was dated younger than 1123 cal. yrs BP at the Espizua’s section in the mire (2005) where it was recorded with a similar thickness (80–104 cm depth). The ages obtained at the VEP section site now constrain this glacio-fluvial event between 594 (480–723) and 607 (549–667) cal. yrs BP. In relation to this, Espizua (2005) proposed Late Holocene glacial advances occurred at the valley, one of them (Amarrilla I-III moraines), undated, linked to the LIA. Likely the analysed glacio-fluvial deposit could be related to an outwash environment linked to some of these three LIA advances in the El Peñón valley.
After the matrix- supported conglomerate deposit, the mire taxa (Cyperaceae 60%) show a fluctuating tendency to decline (Pollen zone 3). Probably, instability in the valley floor as a consequence of higher energy runoff could have caused this Cyperaceae fluctuation. At the same time, the increment of grassland elements (Poaceae reach 80%) and the concomitant decrease in Patagónica-Altoandino elements (e.g. Calandrinia and Arenaria) could be interpreted in a similar way than Pollen zone 1, i.e. a higher representation of grassland from the surrounding slopes implying harsher environmental conditions. However, it should not be ruled out that Poaceae contribution could have been partially provided by growing grasses in the mire, as it is seen nowadays (Wingenroth, 1992).
Accumulation of tephra layers T1-T3 (tephra fallouts or tephra re-worked) also characterises the second phase since 1221 cal. yrs BP onwards. T1 tephra, around 1221 cal. yrs BP (Pollen zone 2, Fig. 5), coincides with a peak in the TOC content and the increase in Cyperaceae pollen which suggests a positive impact of T1 in the mire plants. Taxa growing outside the mire such as Asteraceae and Oxalidaceae show a decrease in abundance which, on the contrary, could indicate a negative effect.
The end of this phase is somewhat uncertain because of the lack of numerical ages. Considering glacial evidence, it could be in coincidence with the end of the LIA, the event associated by Espizua and Pitte (2009) with a neoglacial advance in the valley between ca. 450 and 250 years BP.
A third phase (< ~ 594 cal. yrs BP) in the valley evolution started at the very Late Holocene. A series of parallel downstream gullies were carved exposing the mire and fluvio-glacial deposits of the last 5700 yrs (Fig. 3) in the VEP section site. As a result, mire groundwater could have drained along the new gullies and a likely slower accumulation rate could have characterised the mire formation until present. The lack of a higher resolution at the uppermost metre of the VEP section implies uncertainty about deposition since around 594 (480–723) cal. yrs BP to the present-day.
The most representative mire vegetation started to recover moderately (Cyperaceae 30%; Pollen zone 4, Fig. 5). Concomitantly, the minimal representation of Poaceae (60%) is associated with the opposite trend in Amaranthaceae, Calandrinia, Oxalidaceae, Nassauvia and Wendtia. The increase of shrubs, with the abrupt rise of Amaranthaceae in particular, would indicate human impacts. Pre-Hispanic societies in central Chile made use of Amaranthaceae family plants likely as food supply (Belmar et al., 2010; Planella et al., 2014). Besides, since 250 cal. yrs BP, Spanish agricultural colonies in Chile (e.g. Talca and Curicó) could have moved European cattle to this high-altitude valley during warm summer seasons (Prieto and Abraham, 1998; Durán, 2000).
This third phase of development also records tephra layers T4 and T5, either by tephra fallout or tephra re-working. In particular, T5 tephra (13-cm-thick) coincides with the limit between Pollen zones 3 and 4, suggesting that pyroclastic sediments may have affected the vegetation after its deposition. However, it is not possible to discern if this event would have caused the decline of Poaceae and/or the increment of Cyperaceae and others shrubs (i.e. Amaranthaceae, Wendtia) and herbs (i.e. Oxalidaceae, Calandrinia, Nassauvia).
To sum up, since 5700 cal. yrs BP until 3700 cal. yrs BP the VEP section site suggests aggradation in an outwash environment associated with colder than present vegetation representatives, as the withdrawal of El Peñón glacier front left ice-free valley surfaces (Fig. 6a). Then, since 3700 cal. yrs BP the environment changed to a low-energy mire associated with similar to present vegetation inside and outside the wetland (Fig. 6b). At around 594 cal. yrs BP glacier dynamic and volcanic factors would have provoked instability in the valley floor and mire deterioration (Fig. 6c). Finally, after < 594 cal. yrs BP in the VEP wetland site an incision event interrupted the aggradation, and exposed the VEP sedimentary section at a gully. The mire vegetation recovered moderately. Human disturbance, evidenced by domesticated plants (as Amaranthaceae), would have occurred particularly after 250 cal. yrs BP (Fig. 6d).

Inferred Mid– to Late Holocene environmental evolution scheme from the VEP section at the El Peñón valley. Four main phases have been identified (a-d) based on the sedimentology and pollen analysis
Regional Environmental Correlations
The onset of mire growth after the Lateglacial in the El Peñón valley could have been delayed by arid conditions during the Early Holocene, implying a change from arid to wetter conditions towards the Middle Holocene (Rodbell et al. 2009). This scenario is coincident with the severe aridity and warmer-than-present conditions inferred between 7500 and 5700 cal. yrs BP from the Aculeo lake pollen record (Villa-Martínez et al., 2003), and in others lakes in Central Chile, i.e. Tagua-Tagua (Valero-Garcés et al. 2005) and Laguna del Maule (Frugone-Álvarez et al., 2020). In the El Peñón valley, absolute ages for the early Holocene have not been documented yet.
In the VEP section site between ca. 5700 and ca. 3700 cal. yrs BP, the documented outwash environment and the low vegetation diversity (almost dominated by Pteridophytes) as expected in the context of the unglaciated valley’s surfaces as well as the surroundings valley slopes covered mainly by grasses and others few shrubs and herbs (adapted higher-altitude communities), probably were reflecting wetter and colder than present conditions. In this regard, no evidence of arid conditions are recorded as it has been proposed for the Mid- Holocene in central-west Argentina and the Andean Puna (Gil et al. 2005; Yacobaccio and Morales, 2005). More studies are necessary to find Mid- Holocene records that clarify environmental conditions in the region.
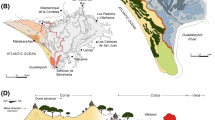
Colder and wetter conditions similar to that inferred at VEP section site between 5700 and 3700 cal. yrs BP are documented in other sedimentary records of the region, i.e. the Salado peatbog (-34°11’S/-69°32’W, 3100 m a.s.l.) (Markgraf, 1983), the Agua Buena alluvial fan (-34°50’S/-69°56’W, 2100 m a.s.l.) (Navarro et al. 2010), and El Sosneado lake (-34°51’S/ 69°55’W, 2100 m a.s.l.) (Navarro et al. 2012)(Fig. 7). The Salado and Agua Buena localities record the end of wetter conditions at ca. 3000 yrs BP and 2800 yrs BP, respectively. On the contrary, El Sosneado lake shows this change at ca. 1900 cal. yrs BP. On the western side of the Andes belt (Chile), colder and wetter conditions have also been inferred between 5700 and 3200 yrs BP, i.e. Aculeo lake (Jenny et al. 2002; Villa-Martínez et al., 2003); these authors interpreted a progressive increase in precipitation in relation to a weakened South Pacific high-pressure cell with intensified Westerlies in the subtropical region of Chile (Jenny et al. 2002). Accordingly, intensified westerlies would have been the main source of humidity at the El Peñón valley during the 5700–3700 cal. yrs BP period.

Correlation scheme of the studied section with Mid- to Late Holocene records in the region, including three sites at the western side of the Andes cordillera Chile (Aculeo, Maule and Chepical lakes) and four at the eastern side (Sosneado lake, Agua Buena alluvial and peat bog sequence and the Salado and El Peñón mire)
The change from an outwash to a mire environment at the VEP section site suggests a shift to warmer and/or drier conditions compared with previous times (Pollen zone 1). The initial stage of mire formation occurs in other Andean highland settings of Mendoza around 3000–4000 yrs BP, i.e. Salado peatbog at ~ 3000 cal. yrs BP (Markgraf, 1983), Agua Buena alluvial fan at ~ 4100 cal. yrs BP (Navarro et al. 2010), and Vega Las Herraduras at ~ 3200 cal. yrs BP (Srur, 2002). This condition is associated with the beginning of a similar-to-present climate (summer rains in the lowlands and more favourable temperatures in the uplands) at this subtropical region at about 3000 yrs by Markgraf (1983). A second neoglacial advance at ~ 2800 cal. yrs BP proposed in the El Peñón valley (Espizua, 2005), implying colder than the present environment, is not documented in our study. Consequently, this scenery could have implied lesser precipitation in the study area compared with the previous period, probably associated with lower influence of westerlies. This contrasts with Jenny et al. (2002) who suggest higher influence of westerlies for the last 3200 yrs BP. Alternatively, in the El Peñon valley temperature would have played a more important role in the balance of humidity as suggested by Markgraf (1983), compared with sites in the western Andes slopes (Jenny et al. 2002; Villa-Martínez et al., 2003.
The change in vegetation between 800 cal. yrs BP and some moment after 600 cal. yrs BP (pollen zone 3) documented in the VEP section site and the associated outwash environment linked to the LIA advance in the El Peñón valley equates the trend of the low-resolution pollen record at Laguna del Maule, Andean highlands of Chile (Carrevedo et al., 2015). This record documents a LIA-like cooling event between 550 and 100 cal. yrs BP (pollen zone LEM-2) (Fig. 7). Besides, at Aculeo lake, a high-resolution pigment-based reconstruction of summer temperatures for central Chile also documents a clear climatic signal related to the LIA-like cooling between 620 and 250 cal. yrs BP (von Gunten et al., 2009). Similarly, the high-altitude Laguna Chepical multiproxy (pollen, diatom and charcoal particles) lake cores (32° S) in Chile, documents the cold period associated with the LIA between 550 and 100 years cal BP (Martel-Cea et al., 2016). Hence, the LIA phase registered in Chile is partially coincident with the period registered in the VEP section site, reinforcing our inference about the LIA event occurrence in the El Peñón valley.
The LIA globally cold event (Bradley et al., 2003) is suggested to have finished in this subtropical region either at ca. 250 cal. yrs BP or at ca. 100 cal. yrs BP as indicated at Aculeo or Maule and Chepical records, respectively. We favour Aculeo’s ages at ca. 250 cal. years BP because it reinforces the idea that people introducing cattle to the El Peñón valley and hunting (Durán, 2000) could have been favoured by the end of the LIA. However, the LIA event in El Peñón valley would have initiated hundreds of years before (ca. 800 cal. yrs BP) than those reported in the sites mentioned. In the northern hemisphere, dated records of ice-cap growth from Arctic Canada and Iceland show that the beginning of the LIA would have been triggered by volcanism (Miller et al. 2012). In such a context, T1-T5 ash deposits recorded in the VEP section site could have played a role in triggering the cold period earlier than in other sites; but the low resolution of our data prevents a deeper analysis. Additional independent high-resolution proxies are necessary to disentangle the signal and duration of this climate event in this area of the Andes highlands.
As regards the development of gullies in the El Peñón valley sometime after 594 cal. yrs BP, similar incision events have been documented regionally, as in the case of the Andes piedmont in Mendoza and central Argentina (Zárate, 2002; Quattrocchio et al., 2008; Mehl and Zárate, 2008; Mehl, 2010; Zárate and Mehl, 2011). The climate and human disturbance, or a combination of both, have been proposed as controlling factors, although the causes remain unsolved (Mehl and Zárate, 2012).
Conclusions
The VEP sedimentary succession permitted reconstructing Mid- to Late Holocene palaeoenvironmental and palaeoclimatic conditions in the Andean highlands and at the subtropics of southern South America. During the last 5700 cal. yrs, the VEP section site evidences climatic fluctuations with a trend from cold to warm present conditions and an environment where water was mostly present. After 3700 cal. yrs BP, environmental conditions favoured Cyperaceae appearance and the setting of the mire, marking the beginning of the similar-to-present environmental conditions.
Other than climate, factors such as volcanism and human disturbance may have also controlled, secondarily, the wetland evolution. Volcanism has been recorded repeatedly since 1220 cal. yrs BP. Although some tephra levels seem to have favoured the development of some taxa and the declination of others, the low resolution of pollen record pre and post tephras hampers a deeper analysis and suggests the need to conduct more high-resolution studies to evaluate the impact of volcanic eruptions in the wetland. Regarding human disturbance, people would have also produced a significant impact in the mire by introducing exotic plants by pre-hispanic people, and/or to a lesser extent summer cattle grazing by Spaniard colonies during the last ~ 250 cal. yrs.
The palaeoenvironmental scenario depicted in this work indicates that at least three forcing factors have influenced the region since the Mid- Holocene onwards at different temporal scales. Consequently, it promotes further higher resolution analyses in this unexplored sensitive mire ecosystem and similar settings in the high southern Andes. Insights on how volcanic eruptions and human impact, in addition to climate, influence wetlands could help to better assess future human management for wetlands conservation worldwide.
Availability of data and material
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Arana M, Bianco C, Martínez Carretero E, Oggero A (2011) Licofitas y helechos de Mendoza. Multequina 20. ISSN: 0327–9375. https://www.redalyc.org/articulo.oa?id=42824203011 last-accessed 18/08/2020
Blaauw M (2010) Methods and code for ‘classical’ age-modelling of radiocarbon sequences. Quat Geochronol 5(5):512–518
Belmar C, Quiróz L, Reyes V (2010) ¿Las comunidades alfareras iniciales de la Zona Central son sólamente cazadoras-recolectoras? Una pregunta enunciada desde el registro carpológico del sitio Estación Quinta Normal, Línea 5 del Metro de Santiago. Actas del XVI Congreso Nacional de Arqueología Chilena. Tomo II: 1179–1189. Santiago.
Benzaquén L, Blanco DE, Bo R, Kandus P, Lingua G, Minotti P, Quintana R. (Eds.) (2017) Regiones de Humedales de la Argentina. Ministerio de Ambiente y Desarrollo Sustentable, Fundación Humedales / Wetlands International, Universidad Nacional de San Martín y Universidad de Buenos Aires, pp 333. ISBN: 978–987–29811–6–7
Bower MM (1960) Peat erosion in the Pennines. Advances of Science 64:323–331
Bower MM (1961) The erosion of blanket peat in the Southern Pennines. East Midlands Geographer 2(13):22–33
Bradley RS, Hughes MK, Diaz HF (2003) Climate in Medieval Time. Science 302:404–405
Bridge J, Demicco R (2008) Earth surface processes, landforms and sediment deposits. Cambridge University Press, Cambridge, p 815
Bruniard E (1982) La diagonal árida Argentina: un límite climático real. Rev Geogr 95:5–20
Cabrera AL (1976) Regiones fitogeográficas Argentinas. In: Kugler WF (ed) Enciclopedia Argentina de Agricultura y Jardinería. Buenos Aires: ACME: 1–85 pp.
Carrevedo ML, Frugone M, Latorre C, Maldonado A, Bernárdez P, Prego R, Cárdenas D, Valero-Garcés B (2015) A 700-year record of climate and environmental change from a high Andean lake: Laguna del Maule, central Chile (36°S). The Holocene 25(6):956–972
Clarkson BR, Ausseil AE, Gerbeaux P (2013) Wetland ecosystem services. In: Dymond JR (ed) Ecosystem services in New Zealand – conditions and trends. Manaaki Whenua Press, Lincoln, New Zealand, pp 192–202
Dean WE (1999) The carbon cycle and biogeochemical dynamics in lake sediments. J Paleolim 21:375–393
Durán V (2000) Poblaciones indígenas de Malargüe. Su arqueología e historia. Serie Libros del CEIDER. Tomo 1. Facultad de Filosofía y Letras. Universidad Nacional de Cuyo. Mendoza.
Durán V, Winocur D, Stern C, Garvey R, Barberena R, Peña Monné J, Benítez A (2016) Impacto del volcanismo y glaciarismo holocénicos en el poblamiento humano de la cordillera sur de Mendoza (Argentina): una perspectiva geoarquelógica Revista Intersecciones en Antropología. Volumen Especial 4: 33–46. Facultad de Ciencias Sociales. Universidad Nacional del Centro de la Provincia de Buenos Aires. Olavarría.
Espizua LE (2005) Holocene glacier chronology of Valenzuela Valley, Mendoza Andes Argentina. Holocene 15(7):1079–1085
Espizua LE, Pitte P (2009) The Little Ice Age glacier advance in the Central Andes (35°S), Argentina. Palaeogeogr Palaeoclimatol Palaeoecol 281:345–350
Evans M, Warburton J (2007) Geomorphology of upland peat: erosion, form and landscape change. Blackwell Publishing, Oxford
Faegri K, Iversen J (1989) Textbook of Pollen Analysis. 4th Edition. (ed K Faegri, PE Kaland and K Krzywinski). John Wiley & Sons: Chichester.
Folk RL, Andrews PB, Lewis DW (1970) Detrital sedimentary rock classification and nomenclature for use in New Zealand. N Z J Geol Geophys 13:937–968
Frugone-Álvarez M, Latorre C, Barreiro-Lostres F, Giralt S, Moreno A, Polanco-Martínez J, Maldonado A, Carrevedo ML, Bernárdez P, Prego R, Delgado Huertas A, Fuentealba M, Valero-Garcés B (2020) Volcanism and climate change as drivers in Holocene depositional dynamic of Laguna del Maule (Andes of central Chile – 36◦ S. Climate of the past 16:1097–1125
Garreaud RD, Vuille M, Compagnucci R, Marengo J (2009) Present-day South American climate. Palaeogeogr Palaeoclimatol Palaeoecol 281:180–195
Grenfell S, Grenfell M, Ellery W, Job N, Walters D (2019) A Genetic Geomorphic Classification System for Southern African Palustrine Wetlands: Global Implications for the Management of Wetlands in Drylands. Front Environ Sci 7:174. https://doi.org/10.3389/fenvs.2019.00174
Gil A, Zárate MA, Neme G (2005) Mid-Holocene paleoenvironments and the archeological record of southern Mendoza, Argentina. Quatern Int 132:81–94
Grimm E (2007) TGView-Tilia Software Version 1.0.1. Illinois State Museum, Research and Collection Center, Springfield, Illinois USA.
Heiri O, Lotter AF, Lemcke G (2001) Loss on ignition as a method for estimating organic and carbonate content in sediments: reproductibility and comparability of results. J Paleolimnol 25:101–110
Heusser CJ (1971) Pollen and Spores of Chile. University of Arizona Press, Tucson.Hoffman, JAJ (1975) Atlas climatológico de América del Sur. OMM-WMO-UNESCO, Hungría.
Hogg AG, Hua Q, Blackwell PG, Buck CE, Guilderson TP, Heaton TJ, Niu M, Palmer JG, Reimer PJ, Reimer RW, Turney CSM, Zimmerman SRH (2013) SHCal13 Southern hemisphere calibration, 0–50,000 years cal BP. Radiocarbon 55:1889–1903
Jenny B, Valero-Garce B, Villa-Martinez R, Urrutia R, Geyh M, Veit H (2002) Early to mid-Holocene aridity in central Chile and the southern westerlies: the Laguna Aculeo record (34S). Quatern Res 58:160–170
Lindsay R (2016) Peatland (mire types): based on origin and behavior of water, peat genesis, landscape position, and climate. In: C.M. Finlayson et al. (eds.), The Wetland Book. Springer: Dordrecht. DOI: https://doi.org/10.1007/978-94-007-6173–5_279–1.
Lindsay R (2018) Peatland Classification. In: C. M. Finlayson et al. (eds.), The Wetland Book. Springer Science+Business Media B.V. DOI: https://doi.org/10.1007/978-90-481-9659-3_341.
Malvárez AI (1999) Tópicos sobre humedales subtropicales y templados de Sudamérica. Oficina Regional de Ciencia y Técnica para América Latina y el Caribe. Programa Hombre y Biosfera-Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura, pp. 228. ISBN 92–9089–064–9.
Markgraf V (1983) Late and postglacial vegetational and paleoclimatic changes in subantarctic, temperate and arid environments in Argentina. Palynology 7:43–70
Markgraf V, D’Antoni LH (1978) Pollen Flora of Argentina. University of Arizona Press, Tucson
Martel-Cea A, Maldonado A, Grosjean M, Alvial I, de Jong R, Fritz SC, von Gunten L (2016) Late Holocene environmental changes as recorded in the sediments of high Andean Laguna Chepical, Central Chile (32°S; 3050 m a.s.l.). Palaeogeogr Palaeoclimatol Palaeoecol 461:44–54. https://doi.org/10.1016/j.palaeo.2016.08.003
Medina ME, Grill S, Fernandez AL, López MA (2017) Anthropogenic pollen, foraging, and crops during Sierras of Córdoba Late Prehispanic Period (Argentina)
Mehl AE, Zárate MA (2008) Sucesiones aluviales del Pleistoceno tardío-Holoceno del valle de Quehué, La Pampa, Argentina. XII Reunión Argentina de Sedimentología (XII RAS). Capital Federal. Buenos Aires, Argentina, del 3 al 6 de junio de 2008. Acta de resúmenes, pp. 115
Mehl AE (2010) Ambientes aluviales del Pleistoceno tardío-Holoceno y Holoceno del Valle de Uco, cuenca del río Tunuyán. In: Gil A, Neme G, Zárate M (Eds.), Paleoambientes y ocupaciones del centro-oeste de Argentina durante la transición Pleistoceno-Holoceno y Holoceno. Sociedad Argentina de Antropología, Buenos Aires, pp. 11- 40
Mehl AE, Zárate MA (2012) Late Pleistocene and Holocene environmental and climatic conditions in the eastern Andean piedmont of Mendoza (33°–34°S, Argentina). J S Am Earth Sci 37:41–59
Mehl AE, Zárate MA (2014) Late Glacial-Holocene climatic transition record at the Argentinian Andean piedmont between 33 and 34◦ S. Climate of the past 10:863–876
Méndez E (2007) La vegetación de los Altos Andes. II Las vegas del flanco oriental del Cordón del Plata. Mendoza Argentina Bol Soc Argent Bot 42:273–294
Méndez, E (2011) La vegetación de Los altos Andes. El flanco oriental del Cordón del Portillo (Tunuyán, Mendoza, Argentina)
Miller GH, Geirsdóttir A, Zhong Y, Larsen DJ, Otto-Bliesner BL, Holland MM, Bailey DA, Refsnider KA, Lehman SJ, Southon JR, Anderson C, Björnsson H, Thordarson T (2012) Abrupt onset of the Little Ice Age triggered by volcanism and sustained by sea-ice/ocean feedbacks. Geophys Res Lett 39:L02708. https://doi.org/10.1029/2011GL050168
Morici EFA, Prina A, Alfonso GL, Muiño W (2010) Flora y vegetación del valle superior del río Atuel (Mendoza-Argentina). Bol Soc Argent Bot 45(1–2):109–118
Munsell Soil Color Charts: Munsell Color x-rite, Revised and washable version, Grand Rapids, X-Rite Incorporated, Michigan, USA, 2000
Navarro D, Mehl AE, Zárate MA, Páez MM (2010) Mid-late Holocene environments of Agua Buena locality (34°50′S; 69°56′W), Mendoza, Argentina. In: Proceedings of the PAGES 1st Young Scientists Meeting: Retrospective View on Our Planet's Future. IOP Conference Series: Earth and Environmental Science. vol. 9 (1). Institute of Physics Publishing, UK, pp. 012014. On-line: 1755–1315. http://iopscience.iop.org/1755-1315/9/1/012014.
Navarro D, Rojo LD, De Francesco C, Hassan G (2012) Paleoecología y reconstrucciones paleoambientales en Mendoza durante el holoceno. In: G. Neme y A. Gil (comps.), Ecología humana y ambiente en el sur de Mendoza. Sociedad Argentina de Antropología
Nichols G (2009) Sedimentology and Stratigraphy, 2nd edn. Wiley-Blackwell, Chichester, p 419
Norte (2000). Mapa Climatológico de Mendoza. In: Abraham, M.E., Martínez, F.M. (Eds.), Recursos y problemas ambientales de zona árida. Primera parte: Provincias de Mendoza, San Juan y La Rioja, Tomo I: Caracterización ambiental. IADIZA, Mendoza, pp. 29–48.
Peralta I (1985) Sinopsis de las especies de Calandrinia (Portulacaceae) de los Andes mendocinos. Bol Soc Argent Bot 25(3–4):511–537
Planella MT, Falabella F, Belmar C, Quiróz L (2014) Huertos, chacras y sementeras. Plantas cultivadas y su participación en los desarrollos culturales de Chile Central. Rev Española Antropol Am 44(2):495–522
Prieto MR, Abraham EM (1998) Historia ambiental del sur de Mendoza (siglos XVI al XIX) Los factores críticos de un proceso de desertificación. Bamberger Geographische Schriften Bd 15(8):277–297
Quattrocchio ME, Borromei AM, Deschamps CM, Grill SC, Zavala CA (2008) Landscape evolution and climate changes in the Late Pleistocene-Holocene, southern Pampa (Argentina): evidence from palynology, mammals and sedimentology. Quatern Int 181:123–138
Reading HG (1996) Sedimentary Environments: Processes, Facies and Stratigraphy, 3rd Edition. ISBN: 978–0–632–03627–1. Wiley-Blackwell, 704 pp
Rodbell DT, Smith J, Mark B (2009) Glaciation in the Andes during the Lateglacial and Holocene. Quatern Sci Rev 28:2165–2212
Roig F (1972) Bosquejo fisonómico de la vegetación de la provincia de Mendoza. Boletín De La Sociedad Argentina De Botánica 8:49–80
Roig FA, Martinez Carretero E, Mendez E (2000) Vegetación de la Provincia de Mendoza. In: Abraham, E.M. y F. Rodriguez Martínez (Eds.), Recursos y Problemas Ambientales de la Zona Árida. Primera parte: provincias de Mendoza, San Juan y La Rioja. Tomo I: Caracterización ambiental: 63–66. IADIZA, Mendoza
Rojo LD, Mehl A, Paez MM, Zárate MA (2012) Mid- to Late Holocene pollen and alluvial record of the arid Andean piedmont between 33°- 34° S, Mendoza, Argentina: inferences about floodplain evolution. J Arid Environ 77:110–122
Rojo LD, Mehl A, Zárate MA, Chivas A, García A (2018) Late Pleistocene and Holocene vegetation changes in the arid Andean piedmont of central Argentina revealed by sedimentary stable carbon isotopes and C/N ratios. Palaeogeogr Palaeoclimatol Palaeoecol 495:205–213
Ruthsatz B, Schittek K, Backes B (2020) The vegetation of cushion peatlands in the Argentine Andes and changes in their floristic composition across a latitudinal gradient from 39°S to 22°S. Phytocoenologia 50(3):249–278
Salgán L, Garvey R, Neme G, Gil A, Giesso M, Glascock M, Durán V (2015) Las Cargas: Characterization of a Southern Andean Obsidian Source and Its Prehistoric Use. Geoarchaeology 30(2):139–150
Srur A (2002) Condiciones paleoambientales durante el Holoceno tardío en la Precordillera (Mendoza). Tesis de Grado. Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata
Tooth S, Ellery F, Grenfell M, Thomas A, Kotze D, Ralph T (2015) Wetlands in drylands research network (10 reasons why the Geomorphology of wetlands is important) http://wetlandsindrylands.net/wp-content/uploads/2015/10/10-Reasons- Geomorphology-of-Wetlands-NEAR-FINAL-PRINTER-FRIENDLY.pdf.
Valero-Garcés BS, Jenny B, Rondanelli M, Delgado-Huertas A, Burns SJ, Veit H, Moreno A (2005) Palaeohydrology of Laguna de Tagua Tagua (34 300 S) and moisture fluctuations in Central Chile for the last 46 000 yr. Journal of Quaternary Sciences 20(7–8):625–641
Viale M, Nuñez MN (2011) Climatology of Winter Orographic Precipitation over the Subtropical Central Andes and Associated Synoptic and Regional Characteristics. J Hydrometeorol 12:481–507
Viale M, Bianchi E, Cara L, Ruiz LE, Villalba R, Pitte P, Masiokas M, Rivera J, Zalazar L (2019) Contrasting climates at both sides of the Andes in Argentina and Chile. Front Environ Sci 7:69. https://doi.org/10.3389/fenvs.2019.00069
Villa-Martínez R, Villagrán C, Jenny B (2003) The last 7500 cal yr B.P. of westerly rainfall in Central Chile inferred from a high-resolution pollen record from Laguna Aculeo (34°S). Quatern Res 60:284–293
Von Gunten L, Grosjean M, Rein B et al (2009) A quantitative high-resolution summer temperature reconstruction based on sedimentary pigments from Laguna Aculeo, central Chile, back to AD 850. The Holocene 19(6):873–881
Yacobaccio HD, Morales M (2005) Mid-Holocene environment and human occupation of the Puna (Susques, Argentina). Quatern Int 132(1):5–14
Wingenroth M (1992) La quebrada Benjamín Matienzo, su naturaleza presente y pasada. Ediciones Culturales de Mendoza, Mendoza
Wingenroth M (2012) Ecosistemas presentes y pasados en la Quebrada Benjamín Matienzo (32º 35’ - 32º 50’ l. s. y 70º 06’ l. o.) Cordillera de los Andes, Mendoza. Argentina. Ameghiniana 69(3):436–456
Wingenroth M, Heusser CJ (1984) Polen en la alta cordillera. IANIGLA, Mendoza
Zárate MA (2002) Geología y Estratigrafía del Pleistoceno tardío-Holoceno en el piedemonte de Tunuyán-Tupungato, Mendoza, Argentina. In: Ostera HA, Panarello HO (eds) Cabaleri N, Cingolani C, Linares E, López de Luchi MG. ACTAS II XV Congreso Geológico Argentino, El Calafate, pp 615–620
Zárate MA, Mehl AE (2011) Evolución geomorfológica holocena de la cuenca media del río Atuel, Mendoza, Argentina. XVIII Congreso Geológico Argentino. Neuquén. Actas (CD-ROM), Temas Generales, Estratigrafía y Sedimentología
Zuloaga FO, Morrone O (eds) (1999a) Catálogo de las plantas vasculares de la República Argentina. II. Acanthaceae-Euphorbiaceae (Dicotyledoneae). Monogr Syst Bot Missouri Bot Gard 74:1–621
Zuloaga FO, Morrone O (1999b) Catálogo de las plantas vasculares de la República Argentina. II. Fabaceae-Zygophyllaceae (Dicotyledoneae). Monogr Syst Bot Missouri Bot Gard 74:623–1269
Acknowledgements
We acknowledge Gendarmería Nacional for allowing us access to El Peñón valley as well as the authorities from the International Centre for Earth Sciences (ICES) for granting access to the El Peñón refuge in 2014. Alejandra Guerci is thanked for helping with initial field work. Thanks are due to Dr. Jay-Henrik May for an early review of the manuscript.
Funding
It was provided through projects from Universidad Nacional de Cuyo (SeCTyP 2011–2013, 06/M051) and Agencia Nacional de Promoción Científica y Tecnológica (PICT-2009–101) from Argentina.
Author information
Authors and Affiliations
Contributions
LDR contributed to the conception and design of the study, data collection and pollen analysis, and wrote the manuscript, reviewed and edited the paper. AEM contributed to the study conception and design, analysis of sedimentological data, and co-wrote, reviewed and edited the paper. MP contributed topollen analysis. VD and RB contributed to the discussion about anthropogenic impact. All authors read and approved the final manuscript. VD was in charge of funding acquisition and administration.
Corresponding author
Ethics declarations
Conflicts of interest
The authors have no conflicts of interest to declare that are relevant to the content of this manuscript.
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Code availability
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Rojo, L.D., Mehl, A.E., Pietrelli, M. et al. Mid- to Late Holocene Environmental Evolution of a High Mountain Wetland in the Subtropical Andes Cordillera of Argentina. Wetlands 42, 32 (2022). https://doi.org/10.1007/s13157-022-01549-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-022-01549-3