
Abstract
Rapid urbanisation increasingly isolates and exerts pressure on natural wetlands, particularly in the fast-growing developing countries of the tropics, including those of West Africa. Constructed wetlands such as sewage treatment plants, may unintendedly offer wildlife protection due to prohibitive access control and limited use, thereby attracting wary and specialised waterbirds in otherwise heavily disturbed formally protected wetlands with less polluted waterbodies. We present data from a rapid survey on 1-year post-opening colonisation and use of waterbirds in a recently constructed 11 ha restricted-access sewage treatment plant, situated in Ghana’s capital, Accra. During November-December 2013 and January 2014, nine daily counts in each month produced an accumulated count of >4200 observations belonging to 26 species of waterbirds, including several important Afro-Palaearctic and intra-African migrants, hereunder ardeids, piscivorous divers, waterfowl and waders. The distributional patterns of waterbirds clearly reflected local foraging opportunities and water quality parameters in the system of 12 inter-connected waste stabilisation ponds. A nearby semi-natural wetland with cleaner waterbodies, but higher levels of human interference, supported fewer waterbirds, predominantly commensal gregarious species. Our data suggests that strict protection from disturbances outweighs possible negative implications attributed to mere pollution of waterbodies supporting various waterbird guilds, thus highlighting the potential importance of informally protected sewage treatment plants distributed in functional networks, as a complement to designated wetlands. We anticipate that establishing similar or larger plants jointly will improve sewage treatment and waterbird conservation in urban Ghana, and West Africa in general.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Wetlands are high on the conservation agenda across the globe due to decades of unsustainable anthropogenic exploitation and degradation (e.g. Dudgeon et al. 2006; Keddy et al. 2009; Davidson 2014), but also by virtue of their pivotal role in ecosystem functioning and services related to climate change, hereunder flood control (Knapp et al. 2019; Shokoufeh et al. 2021), drought management (Dean et al. 2015; Dixon et al. 2021) and carbon sequestration (Dayathilake et al. 2021). Wetlands are subjected to heavy disturbance regimes, particularly in urban areas, where draining for agriculture, farming, fishing, hunting, livestock rearing, mining, and pollution from sewage encroach upon and degrade waterbodies with associated wildlife (e.g. Schuyt 2005; Verhoeven et al. 2006; Junk et al. 2013; De Troyer et al. 2016). Among the vulnerable wetland fauna are various guilds of waterbirds, including many migrants (Okes et al. 2008; Sutherland et al. 2012; Runge et al. 2015). Conservation efforts to safeguard wetlands include regulations and protection of key sites (Beatty et al. 2014; Kleijn et al. 2014), but such efforts are often hampered by increasing human population and associated economic growth, as well as inadequate management, legislation and law enforcement (O’Connell 2000; Aynalem and Bekele 2008; Gbogbo 2007a; Gbogbo et al. 2008, 2009). Across most developing tropical countries, wetland conservation and management are therefore typically curtailed by unsustainable human activities, particularly in densely populated urban areas of sub-Saharan Africa (Mitchell 2013; Vickery et al. 2014), hereunder West Africa (Adams 1993; Uluocha and Okeke 2004), including Ghana (Gbogbo 2007b; Lamptey and Ofori-Danson 2014).
With the current prospects of unpredictable and unusual rainfall patterns, related to global warming, wetlands, whether natural or man-made, are increasingly a focal integrated part of urban landscape and environmental planning (Erwin 2009; Junk et al. 2013). As such, albeit constituting seriously threatened ecosystem components, rivers, streams, lakes, dams and ponds likewise offer great potentials for mitigation against environmental and economic disasters related to rainstorms and flooding (Douglas et al. 2008; Knapp et al. 2019; Shokoufeh et al. 2021), as well as prolonged periods of drought (Dean et al. 2015; Dixon et al. 2021). Their importance for sustained ecosystem functions and services therefore cannot be over-emphasised, not the least across the African continent burdened by low agricultural production, poor infrastructure, urban congestion and impoverished health facilities (Douglas et al. 2008; Belle et al. 2018; Dixon et al. 2021).
Formally protected wetlands in Africa, including Ramsar sites, are increasingly subjected to over-exploitation and illegal activities (Schuyt 2005; Gbogbo et al. 2008; Finlayson 2012; Junk et al. 2013; Lamptey and Ofori-Danson 2014; Dixon et al. 2021). Waterbirds and their habitats are often threatened by illegal and unsustainable utilisation of natural resources, including fish stock, invertebrates and fuel wood (O’Connell 2000; Willoughby et al. 2001; Gbogbo et al. 2008; Okes et al. 2008). Alongside unsustainable resource extraction that interferes with waterbird foraging and nesting ecology, the avifauna is also indirectly affected by water pollution and draining, as well as directly by hunting, water sports and other recreational activities (O’Connell 2000; Uluocha and Okeke 2004; Mitchell 2013). Thus, informally protected or unmanaged wetlands serve as important habitat supplements and sometimes alternative refuges for waterbirds (Attuquayefio and Gbogbo 2001; Gbogbo 2007b; Harebottle et al. 2008; Gbogbo and Attuquayefio 2010; Murray and Hamilton 2010). Artificial or constructed wetlands (CW) often receive informal protection, and include agro- and aquaculture ponds, irrigation dams, salt ponds, drainage canals (Froneman et al. 2001; Otieno et al. 2015), as well as sewage treatment plants (STP) of various types (Scholz and Lee 2005; Wang et al. 2014). The construction of STPs is steadily expanding in many parts of the temperate zone (Scholz and Lee 2005; Knapp et al. 2019), subtropics (Hsu et al. 2011; Wang et al. 2016; Giosa et al. 2018) and tropics (Murray and Hamilton 2010; Murray et al. 2012, 2014), with a gradual increase in some parts of Africa (Ashkenazi 2001; Kivaisi 2001; Harebottle et al. 2008; Wang et al. 2014; Dean et al. 2015; De Troyer et al. 2016). Even though the waterbird diversity of CWs and STPs may be inferior to designated protected areas such as Ramsar sites (Hamdi and Ismail-Hamdi 2014; Kleijn et al. 2014), substantial evidence suggests that man-made wetlands may increasingly contribute to waterbird conservation in the near future (Murray and Hamilton 2010; Wang et al. 2016; van Biervliet et al. 2020). As such, although often comparatively small, isolated and situated far from other larger protected wetlands, artificial wetland habitats can increasingly serve as replacement or complementary refuges for lost or degraded natural wetland biota, including vulnerable waterbird guilds (Afdhal et al. 2012; Murray et al. 2012; Wiegleb et al. 2017; Giosa et al. 2018). The efficiency and success of STPs for waterbird conservation, however, depend on the degree and extent of applied ecology-based management practices and access control (Ashkenazi 2001; Hsu et al. 2011; Murray et al. 2014; Wang et al. 2016; Wiegleb et al. 2017; van Biervliet et al. 2020).
Majority of African studies that assess waterbird diversity and habitat utilisation in STPs are from the southern and eastern regions of the continent, with apparently limited information from the northern, western and central parts (Ashkenazi 2001; Harebottle et al. 2008; Harrison et al. 2010; Wang et al. 2014). Moreover, few studies from Africa document the early phases of STP waterbird colonisation (Ashkenazi 2001; Dean et al. 2015), just as most data collected are from coastal areas (Harrison et al. 2010) or inland plants, integrated with natural wetlands (Harebottle et al. 2008). Ghana, and Accra in particular, has in the past decade initiated new strategies for improved sewage treatment to curb the heavy pollution that threatens many waterbodies in the urban coastal zone (Nixon et al. 2007). Since the early 2000s, this has resulted in the planning and initial construction of improved sewage systems and different types of STPs, one of which is the Legon Sewage Treatment Plant (hereafter called LSTP), situated amidst a densely populated area in the Accra metropolis, which is part of the Accra Sewerage Improvement Project (ASIP).
In this study, we present data on the early-phase (~1 year) colonisation by waterbirds at the LSTP, with emphasis on spatio-temporal abundance, distribution and diversity within various components of a unique inland STP in urban Ghana. The major objectives of this baseline study were to describe and assess the waterbird fauna with regard to the: 1) early inflow and colonisation; 2) spatio-temporal patterns of distribution and diversity; 3) relative conservation importance of migrants and rarities; 4) protection potential for vulnerable and wary species; 5) relative importance of water pollution versus unintended bird protection. We therefore aimed at increasing the general knowledge base on waterbird utilisation of CWs in urban landscapes of West Africa. Our early data also serve to focus attention on the conservation potentials of newly established STPs, as well as highlighting the urgent need for additional and complementary refuges for vulnerable waterbirds, alongside improved sewage management in the West African sub-region.
Methods
Study Area
The LSTP (GPS: 5°39’50 N; 0°11’31 W) is situated ~14 km north of the Accra coastline on a gentle slope (~25 m total elevation gradient) adjacent to the main University of Ghana, Legon campus (Fig. 1). It is bordered by the heavily-trafficated Haatso-Atomic road, thickets and farmlands surrounding the Legon campus, as well as the Legon Botanical Gardens with the integrated >60 yrs. old constructed Vaughan’s Dam, isolated from, but situated <200 m away from the LSTP. This nearby <0.02 km2 semi-natural wetland area comprises ~0.01 km2 of open waterbodies bordered by narrow verges of mixed swampy vegetation zones with dense reed beds, herbs and thickets (Fig. 1), and is supplied by serene small streams that emanate from the adjoining swampy areas of the Legon Botanical Gardens. The avifauna of the Vaughan’s Dam was well-known >20 years prior and up to the survey, and this waterbody was not contaminated by sewage or agricultural effluents, although the algal eutrophication level was moderate with a water transparency estimated at ~0.40-0.50 m during the peak of the major dry and hot season in December-February, 2013-2014 (L.H. Holbech, pers. obs.). Incidentally, the Vaughan’s Dam underwent drastic changes in its human use along with the operation of the LSTP in 2012-2013. Hence, intense anthropogenic disturbances were introduced abruptly in the form of commercial recreational activities, including angling, canoeing, cycling, jogging, play-grounds, group picnics, and large parties with loudspeakers. Moreover, small-scale aquaculture was established, and the old artificial wetland was manipulated by reed removal and thicket thinning, as well as deepening and cleaning of the banks by mechanical excavation (L.H. Holbech, pers. obs.).

a Aerial view of the Legon Sewage Treatment Plant (LSTP), with the surrounding urban areas and the Legon Botanical Gardens to the East; b Perpendicular perspective of the LSTP; images taken with a drone (DJI Mavic Pro®)
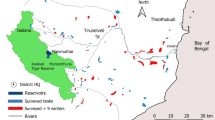
The LSTP measures ~0.11 km2 (~310 × 360 m), of which ~0.06 km2 are open water in the form of ponds lined by rocky banks, and access is restricted by 2.5 m tall barbed wire fencing. It is a stabilisation pond system (Murray et al. 2014; Wang et al. 2014; Dean et al. 2015), constructed in 2010-2012 by an African Development Bank project, and managed by the Accra Metropolitan Assembly under the ASIP. It consists of 12 ponds, with four levels of three similar-sized ponds each (Figs. 1 and 2). The four levels are inter-connected from upper level A to lower level D as: A1-3 = anaerobic ponds, 5 m deep, measuring ~60 × 40 m; B1-3 = facultative ponds, 2.5 m deep, measuring ~75 × 100 m; C1-3 = upper maturation ponds, 1.3 m deep, measuring ~85 × 75 m; D1-3 = lower maturation ponds, 1.3 m deep, measuring ~85 × 65 m (Fig. 2). Water flow is gravitational, with a maximum daily inlet capacity of ~9000 m3, receiving water from the campuses of Legon, the Presbyterian Boys’ Secondary School, and other surrounding state-owned institutions. The small anaerobic ponds are lined with a rubber membrane and flanked by three parallel concrete water-inlet ducts, measuring ~0.9 m wide and ~ 0.9 m deep (Figs. 1 and 2). These ponds were half covered with sludge at the time of our study, and fill up in about 5 years. The large facultative ponds are lined with granite rocks of ~0.1-0.5 m diameter, each having a ~ 1.3 m wide rock intercept situated perpendicularly midway to reach halfway shore to shore (Figs. 1 and 2). The two similar three-set maturation ponds are likewise lined with granite rocks, but lack midway intercepts. At the time of our study, the granite rock linings were partially covered by pioneering sedge grasses, herbs and shrubs characteristic of nutrient-rich loamy soil, typically the Nut sedge grass Cyperus blepharoleptos, the shrub White popinac Leucaena leucocephala, and the herbs Bachelor’s button Gomphrena celosioides, Hogweed Boerhavia diffusa, and Hornbeam-leaved crossberry Grewia carpinifolia, whereas the surroundings were dominated by 1-2 m tall and tough grasses such as Guinea grass Panicum maximum, partly covering the laterite gravels. The vegetation lining the ponds, as well as floating and submerged macrophytes were regularly (i.e. twice yearly) controlled mechanically or by Glyphosate herbicide application, albeit not during our study period. Floating or submerged macrophytes were only present in the C and D ponds; D2 was particularly extensively covered by water lilies (Nymphaea sp.) and reeds (Typha sp.) at the edges. Water clarity was least in the B and C facultative/maturation ponds, indicating explosive algal growth. The LSTP is the largest wetland in a radius of ~10 km from the UGL campus, with the nearest other large constructed wetland ~9 km to the east, namely the ~1.40 km2 Nungua Agriculture Station Dam with its associated marshy areas around the Ayensu River. The Sakumo Lagoon, a Ramsar Site of ~13.40 km2 which joins the Ashaiman Agricultural Dam, is situated ~17 km to the south-east (Fig. 2).

Map showing the position of the Legon Sewage Treatment Plant (LSTP), with surrounding coastal and inland wetlands within the Accra Plains (top), Southern Ghana (insert). LSTP only drawn approximately to scale (bottom)
Data Collection
Nine systematic bird counts per month, each typically enduring 1.5-2 h, were conducted by two observers from November 2013 to January 2014, a period that constitutes the major dry season (November-February) on the Accra Plains, and coincides with the peak numbers of Afro-Palaearctic waterbird migrants in Ghana (Ntiamoa-Baidu 1991; Lamptey and Ofori-Danson 2014). Given the likelihood that the abundance of some species fluctuated throughout the day, three separate count periods were applied equally on each of the nine census days per month, i.e. in the morning (06 h00-08 h00), midday (12 h00-14 h00), and late afternoon (16 h00-18 h00). We anticipated that this systematic sampling of three time periods, repeated thrice per each month, increased the probability of recording as many species as possible, as well as reflecting the true spatio-temporal abundance at the plant. Each of the three replicate ponds (1-3) at each level (A-D) was considered as a unit, within which all birds were counted, before moving on to the next pond, starting from either A1 or D3 (Fig. 2) on each day, on a day-wise rotational basis. Total counts in each of the 12 ponds were done from vantage points with good visibility and aided by a pair of 10 × 42 binoculars, and occasionally a telescope (20-60×). Care was taken to avoid repeat-counting individuals that moved from pond to pond, particularly for wary and mobile species such as ducks and other gregarious birds. This was done by constantly observing the movements of flushed individuals when cautiously approaching a pond. The A-C ponds could comfortably be surveyed and monitored in this way, whereas D ponds were not visible from the levels of A and B ponds. Most flocks relocated from only one level to the adjacent, and rarely did so more than two levels apart. In the case of large numbers of White-faced Whistling Duck Dendrocygna viduata and other gregarious birds, recounting several times with subtractions and additions or averaging were often deemed necessary; counts of such species were associated with an error of ~10%. Notes on breeding were taken of nests, eggs, juvenile or immature birds. Other vertebrates, invertebrates and characteristic plants observed were also recorded opportunistically.
As the lead author (LHH) had been resident in the Legon campus area for >15 years during the period of 1990-2014, the presence and relative abundance of the avifauna in and around the Vaughan’s Dam and the Legon Botanical Gardens was thoroughly known, and this familiarity was based on years of observations during regular birdwatching and walks (L.H. Holbech, pers. obs.). Similarly, non-systematic observations of nocturnal flight activities of ducks and ardeids were made during occasional birding or regular walks and runs in the vicinity (<0.5-1 km) of the LSTP (L.H. Holbech, pers. obs.). Besides bird counting, on site sampling of physico-chemical water quality parameters in each of the 12 ponds was undertaken on a single day in December 2013, with a Horiba U-52G multi-parameter water quality meter, including pH, turbidity (NTU) and total dissolved oxygen (mg/l).
Data Analysis
The three similar ponds at each level were regarded as replicate units in calculating monthly means ± SD of bird abundance based on counts at all the four pond levels over the three-month study period. We used the exponential of the Shannon-Wiener species diversity index (ExpH’) to calculate total effective species number (Jost 2006) for each pond level based on the three replicate counts for each level over the entire study period. Similarly, to quantitatively assess the overall species similarity among the four pond levels, we used the Morisita-Horn index (CMH), which is an abundance-based index resistant to under-sampling biases, and sensitive to the most abundant species, thus having a good discriminant ability for differences in functional diversity between compared sites (Jost et al. 2011). As such, we were able to assess the spatio-temporal abundance, diversity and similarity of waterbirds distributed at the LSTP. Likewise, for the water quality parameters, the mean ± SD was calculated based on the replicate ponds (1-3) at each of the four levels. These parameters were then related to the abundance and diversity of birds in order to evaluate any apparent implications for the waterbirds present at the four pond levels (A-D). To assess any relationships between each of the three water quality parameters (pH, TDO, turbidity) and the distribution of waterbirds across the four pond levels, we performed Pearson’s r-correlations of the parameters as explanatory variables (i.e. the mean of the three replicate ponds for each pond level) with the total abundance of each of the six foraging guilds counted over the total period of three months, as the respondent variables, and applied the best curve fits for r-values at n = 4.
Each waterbird species was classified into six overall foraging guilds based on information in Ntiamoa-Baidu et al. (1998), as: Guild 1 = Herbivorous waterfowl (ducks); Guild 2 = Visual surface foraging waders; Guild 3 = Tactile surface foraging waders; Guild 4 = Pelagic foraging waders; Guild 5 = Stalking herons; Guild 6 = Diving waterfowl (grebes and cormorants). Based on this classification, the modes of foraging employed by the species with their respective abundances could be compared, in order to assess the habitat and niche provisions provided for by the four levels (ponds A-D). Such assessment is important for understanding the spatial distribution of birds in various types of STPs, and this information can be used to optimize construction design and management practices in order to increase waterbird protection and conservation. Each species was also classified according to migratory status, as Afro-Palaearctic migrant, intra-African migrant, and resident for Ghana (Dowsett-Lemaire and Dowsett 2014), as well as global conservation status following the IUCN Red List criteria (Handbook of the Birds of the World Alive/BirdLife, online data).
The coincidence of the LSTP inception with the drastic environmental changes at the Vaughan’s Dam in 2012-2013, provided us with a unique opportunity to compare the early development of the avifaunal colonisation at the LSTP with the avifaunal changes at the Vaughan’s Dam, before and after these disturbances were introduced. As such, even though the in depth knowledge on the waterbird fauna of the nearby Vaughan’s Dam that LHH had obtained during regular birdwatching in 1990-2014 (L.H. Holbech, pers. obs.), was not an integral part of the systematic LSTP data sampling, we were able to relate the LSTP waterbird composition with the avifaunal changes detected at the Vaughan’s Dam, up to and during 2012-2013. We therefore performed a qualitative comparison (see Discussion) of the waterbird presence and relative abundance, by converting the quantitative LSTP data into five approximate abundance categories that were then related to similar abundance categories based on the data gathered at the Vaughan’s Dam during 1990-2014; namely: ‘Abundant’ = ~20-25 or more birds observed on any visit; ‘Common’ = 10-20 birds observed on ~50-100% of visits; ‘Frequent’ = 5-10 birds observed on ~50-75% of visits; ‘Uncommon’ = 3-5 birds observed on ~25-50% of visits; ‘Rare’ = singletons or a pair observed on ~25% or less of visits (L.H. Holbech, pers. obs.).
Results
Total Abundance of Birds in Relation to Pond Levels
We accumulated a total of 4224 bird observations counted on 27 days over the three-month study period, translating into a mean of 156 birds per day-count (Table 1). The six most abundant species comprised ~82% of individual birds counted, and were in the following order of descending dominance; White-faced Whistling Duck (~20%), Cattle Egret Bubulcus ibis (~17%), Common Sandpiper Actitis hypoleucos (~14%), Black-winged Stilt Himantopus himantopus (~13%), Wood Sandpiper Tringa glareola (~11%) and Spur-winged Lapwing Vanellus spinosus (~7%). Other common species were Little Grebe Tachybaptus ruficollis (~4%), Common Greenshank Tringa nebularia (3%) and Senegal Thick-knee Burhinus senegalensis (3%). Abundance was significantly lower at A ponds (χ2 = 526.3, p < 0.00001, df = 3), and 4-5 times higher at B, C and D ponds, amongst which abundance did not differ significantly (χ2 < 1.3, p > 0.5, df = 2). The most abundant birds at A ponds were Common Sandpiper, Black-winged Stilt, Spur-winged Lapwing and Wood Sandpiper, with a few other waders. Similarly, at B ponds, common and wood sandpipers dominated, together with black-winged stilts, spur-winged lapwings, as well as Senegal thick-knees, common greenshanks and cattle egrets (Table 1). The C and D ponds differed from A and B, having a dominance of particularly White-faced Whistling Duck, as well as relatively higher numbers of cattle egrets and little grebes. Although C and D ponds were very similar, C had particularly high dominance of white-faced whistling ducks, whereas cattle egrets dominated at D (Table 1). Total monthly counts from November 2013 to January 2014 were 1463, 1164 and 1597, respectively, indicating a consistently high presence of most species during the study period. However, the overall abundance was significantly (χ2 = 36.3, p < 0.00001, df = 2) lower in December, primarily attributed to relatively lower numbers of the three most abundant species, White-faced Whistling Duck, Cattle Egret and Common Sandpiper (Table 1). The daily count periods of morning-midday-afternoon did not show any significant differences indicating stable populations throughout the daytime hours. Cattle egrets and white-faced whistling ducks displayed the highest daily and monthly abundance fluctuations during the study period, for which the latter were often flight-active after sunset and throughout the night (L.H. Holbech, pers. obs.).
Species Richness, Diversity and Similarity in Relation to Pond Levels
We recorded a total of 26 waterbird species during the three-month study period, including 11 waders, eight ardeids, six waterfowl and the Malachite Kingfisher Corythornis cristatus (Table 1). Only seven species were recorded at A ponds, which contrasted with 19 or 20 at B, C and D ponds. Species diversity was highest at C ponds, whereas it was very similar at B and D ponds (Table 1). The distinctively higher species diversity at C ponds is attributable to a higher evenness and lower dominance of the most abundant species (Table 1). Likewise, abundance-based species similarity (CMH) was highest between A and B ponds, with ~86% species similarity (~54% shared species), whereas C and D ponds had ~60% similarity and ~ 81% shared species (Table 2). In contrast, D ponds versus A and B ponds, respectively, displayed relatively low similarities of 15-20%, albeit with ~31-62% shared species. C ponds versus A and B ponds, respectively, had medium similarity of ~31-32%, but with ~42-58% shared species (Table 2). In summary, C and D ponds were most similar with regard to abundances of particular species as well as number of total shared species, with the least similarity between A and D ponds, and moderate similarity between B and C ponds.
Distribution of Foraging Guilds in Relation to Pond Levels and Water Quality
The White-faced Whistling Duck (Guild 1) was absent from A ponds, scarce at B ponds, fairly abundant at D ponds, and most abundant at C ponds (Table 3). Only two other species in Guild 1 were observed; the Common Moorhen Gallinula chloropus, exclusively and equally found in C and D ponds, and a single Knob-billed Duck Sarkidiornis melanotos in January 2014 at B ponds (Table 1). Visual surface foraging waders (Guild 2), as the most diverse foraging guild with nine species, was represented at all levels, but with the highest abundance and diversity at B ponds, a pattern similarly displayed by the two very abundant small sandpipers (Tables 1 and 3). Only one tactile surface foraging species (Guild 3), Little Stint Calidris minuta, was detected, indicating the lack of exposed mud flats during the study period. In contrast, the pelagic foraging waders (Guild 4), notably represented by black-winged stilts, showed high affiliation to B and C ponds, moderate for A ponds, and only occasional at D ponds. The very diverse guild of eight piscivorous stalking or darting ardeids (Guild 5) showed outstandingly high diversity and abundance at C and D ponds (7-8 species respectively), the latter dominated by large cattle egret flocks. This guild, however, was not recorded at A ponds. Similarly, the guild of diving piscivores (Guild 6), represented primarily by the Long-tailed Cormorant Microcarbo africanus and Little Grebe, showed highest affinity for C and D ponds, with a few of the latter species at B ponds, and a complete absence of the guild at A ponds.
The levels of pH showed weak to moderate alkalinity, and there were significant differences among the four pond levels (Kruskal-Wallis H-test = 9.359, p = 0.02488, df = 3), with A ponds lowest, C ponds highest, and B and D ponds similar (Fisher’s LSD test) (Table 3). Total dissolved oxygen (TDO) showed significant differences across the pond levels (Kruskal-Wallis H-test = 8.128, p = 0.04344, df = 3), with C ponds higher than both A and D ponds, whereas B and C ponds, as well as A, B and D ponds were similar (Fisher’s LSD test). Turbidity also varied significantly among the four levels (Kruskal-Wallis H-test = 8.744, p = 0.0329, df = 3), with D ponds significantly lower than A, B and C ponds. However, turbidity levels in the three latter ponds were not significantly different (Fisher’s LSD test). In summary, ponds B and C showed highest levels of both TDO and turbidity, indicating highest levels of algal growth, whereas the anaerobic nature of A ponds was evidenced by very low TDO and moderately high turbidity. The relatively low turbidity and TDO in D ponds indicated lower algal densities.
The respective correlations between each of the three water quality parameters (pH, TDO and turbidity), measured within the four pond levels (A, B, C, D), as against total abundances of each of the six foraging guilds counted over the 3 months, did not show significant relationships for any of the best-fitted trend lines at the 5% level. However, with regard to turbidity versus total abundances of foraging guilds, Guild 2 and 4 showed positive correlations, in contrast to Guild 5 and 6, indicating negative relationships (Fig. 3). These non-significant correlations suggest that piscivorous divers (i.e. cormorants and grebes) or darters (ardeids) are favoured in D ponds with lower turbidity (i.e. higher water clarity), also shown by the significantly higher abundances of those two guilds, particularly in the D ponds (Table 3). In contrast, for Guild 2 and 4, and partly also for Guild 1, the high turbidity, particularly in B and C ponds, appeared to favour the presence of waterbirds foraging either visually on the surface (Guild 2), or by tactile (Guild 4) or filtering (Guild 1) means in the pelagic water zones with high turbidity, attributed to either algal growth or dead organic matter in the suspended sludge of the turbid waterbodies of B and C ponds.

Correlation between overall bird abundance (total accumulated counts over 3 months) versus turbidity (mean turbidity across three replicates for each of the four pond levels, A, B, C, D) at the Legon Sewage Treatment Plant; (a) Guild 2 and 4; (b) Guild 5 and 6
Migratory and Conservation Status of Waterbirds
We recorded a total of 11 Afro-Palaearctic migrants (~42% of overall species richness), including Black-winged Stilt, whose migration status remains uncertain in Ghana (Dowsett-Lemaire and Dowsett 2014). Resident breeders within Ghana numbered at most 17 species (~65%), including Little Egret Egretta garzetta and Intermediate Egret Ardea intermedia, both with uncertain breeding status in Ghana (Dowsett-Lemaire and Dowsett 2014). Four species were intra-African migrants, including Intermediate Egret. Overall, the proportions of resident breeders therefore seemed higher than migrants. Even amongst migrants, Afro-Palaearctics were in the majority. All species recorded had IUCN Red List status of ‘Least Concern’, and except for Knob-billed Duck, all species are commonly recorded in coastal habitats of Ghana (Ntiamoa-Baidu 1991; Lamptey and Ofori-Danson 2014).
Discussion
Distributional Patterns of Waterbirds at the LSTP
The LSTP waterbird fauna was represented by six separate foraging guilds, two of which dominated in terms of both species number and abundance, namely visual surface waders (Guild 2) and stalking ardeids (Guild 5), in addition to two non-diverse but abundant guilds, herbivorous waterfowl (Guild 1) and pelagic foraging waders (Guild 4). This guild pattern, with relatively low abundance of diving piscivores (Guild 6) and tactile foraging waders (Guild 3) suggests that turbidity was excessively high and/or that prey availability was limited in water columns and on exposed mudflats. The university community and its co-users of the LSTP went on vacation during the months of December and January (i.e. two of the driest months on the Accra Plains), causing the facility to run at low capacity. It was only during this period of low water levels at C and D ponds that foraging conditions were favourable for Guild 3, which included little stints. Particularly D ponds with the highest water clarity (i.e. low turbidity) supported piscivorous darters and divers (Guild 5 and 6), although C ponds were also frequented relatively often by ardeids, cormorants, grebes and kingfishers. At the time of the study, it was unlikely that fishes may have been introduced by natural means from the nearby small Vaughan’s Dam, although pelagic invertebrates that constitute major food sources for pursuit-diving little grebes may have been abundant (Santoul and Mastrorillo 2004). Likewise, the few long-tailed cormorants suggested that, although these were attracted to the ponds, successful prey captures were most likely large invertebrates (Otieno et al. 2015). Although it is possible that particular pelagic invertebrates (e.g. predaceous diving beetles, Dysticidae) were most abundant in the B and C ponds, the significantly higher turbidity measured there, most likely, may have limited actual prey availability for both diving and darting foragers, as compared to the D ponds with more transparent water, even if the latter were supporting lower prey densities. In summary, the distribution of waterbirds at LSTP largely followed the ecological requirements for each of the six foraging guilds, thus probably reflecting a compromise between prey abundance and availability versus water level (Ntiamoa-Baidu et al. 1998; Gbogbo et al. 2009) and clarity (Holbech et al. 2018), with the latter related to nutrient concentrations and algal blooms (Lamptey and Ofori-Danson 2014).
Significance and Potential Implications of LSTP Data for Waterbird Conservation in Urban Ghana
This study was conducted ~1 year post-opening of the sewage inlet, October 2012, and documented at least 26 waterbird species with average day-counts of ~160 birds, and occasionally up to >220 birds. Notable abundant species were White-faced Whistling Duck (up to ~50-100 at a time), Common Sandpiper, Black-winged Stilt, Wood Sandpiper, Spur-winged Lapwing, Little Grebe, Common Greenshank and Senegal Thick-knee. Many factors may contribute to waterbird colonisation of constructed wetlands, hereunder the size of wetland (Beatty et al. 2014; van Biervliet et al. 2020), the proximity and connectivity to other wetlands (Erwin 2002; Knapp et al. 2019), the surroundings (Dean et al. 2015), food availability (Afdhal et al. 2012; Murray et al. 2014), and protection against disturbances (Harrison et al. 2010; Dean et al. 2015). Nevertheless, it would be inappropriate to explain away the significant bird numbers counted at the LSTP, as attributed to mere random events, particularly as several other formally protected or more natural wetlands of considerable size are within relatively short flight distances, including the 13.40 km2 Sakumo Ramsar site, only 17 km away. Two obvious questions are apparent, though: 1) are other nearby wetlands increasingly less attractive or insufficient to hold increasing waterbird populations?; 2) is the LSTP offering superior protection, and as such, despite its heavier pollution levels, still particularly attractive to wary species? Indeed, both explanations are not mutually exclusive, and probably valid, given the currently challenged status of wetlands in the urban areas of the Accra Metropolis (Gbogbo and Attuquayefio 2010; Lamptey and Ofori-Danson 2014).
For instance, although not completely comparable with regard to size and structure of the LSTP, it is remarkable that the nearby Vaughan’s Dam (hereafter only VD), supplied with rain water from adjoining serene environments, and thus obviously with a superior water quality and clarity as compared to the turbid and highly eutrophicated LSTP ponds, only supported ~5-10 regular or frequent waterbird species, up to and during the same study period (L.H. Holbech, pers. obs.; Table 4). Most plentiful were gregarious, commensal and least wary species, such as the abundant Cattle Egret with a rookery of ~500 birds (L.H. Holbech, pers. obs.), the commonly breeding Long-tailed Cormorant, as well as the frequent-common Squacco Heron Ardeola ralloides and Black-crowned Night Heron Nycticorax nycticorax (Table 4). After 2012-2013, the same period that the operation of the LSTP initiated, the VD had been subjected to intensification of small-scale aquaculture and commercial recreational activities, thus rapidly reducing the numbers of wary waterfowl and wader species, hereunder sandpipers, lapwings, moorhens and jacanas, with a complete absence of ducks and little grebes (Table 4). This semi-natural wetland, both with a higher aesthetic value and better water quality, thus supported far fewer waterbirds than the ‘repugnant’ LSTP, hence most likely attributed to its small size, combined with the much higher anthropogenic disturbance levels, thereby preventing wary species from seeking adequate refuge. Indeed, we find it very likely that many of the formerly more regularly recorded wary waterbirds (e.g. waterfowl) at the VD may have found refuge at the LSTP, which is just a very short flight distance away. Ironically, the attempts to ‘beautify’ and make the VD more attractive to humans also made it more repellent for many waterbirds, which then found refuge at the, for humans, repugnant LSTP. Future studies, involving capturing and colour marking, would evaluate apparent LSTP-VD movements of waterbirds, hereunder to what extent true dispersal or mere commuting are quantitatively involved in any source-sink dynamics between the two adjacent ‘contesting’ wetlands (Erwin 2002; Beatty et al. 2014). We regularly observed cattle egrets, cormorants, night herons and squacco herons commuting between the two CWs, all gregarious species, and colonial breeders at the VD.
The relatively small LSTP pond-bank-verges area (~0.06 km2) implies that the overall average bird abundances translates into a mean density of ~2700 birds/km2 (~27/ha), and occasionally up to ~3500-4000. In comparison the mean density of birds at nearby coastal wetlands, including Ramsar sites, ranges between 300 and 900 birds/km2 (Gbogbo and Attuquayefio 2010), given their much larger areas (>2.50 km2), such as Sakumo Lagoon (13.40 km2) and Densu Delta (46.20 km2). Overall bird density at LSTP was also considerably higher than those recorded for rice fields across West Africa, ranging from 400 to 2000 birds/km2 (Wymenga and Zwarts 2010) or artificial wetlands of Tunisia with >100 birds/km2 (Hamdi and Ismail-Hamdi 2014). However, LSTP-density is similar to densities reported from a South African ~0.53 km2 large wastewater treatment plant of >2600 birds/ km2 (Harebottle et al. 2008). The relatively high density of waterbirds recorded at the LSTP throughout the study period, indicates that despite heavily polluted water bodies, foraging opportunities remain conducive for several waterbird guilds, thus highlighting the attractiveness to waterbirds with regard to abundant food sources. Our results are consistent with small-medium sized STPs in arid areas of Africa providing stable waterbodies even in the driest months (Dean et al. 2015), although small shallow-water ponds with more eutrophic water, and thus higher productivity, often support relatively higher bird densities than larger, oligotrophic low-productivity dams (Afdhal et al. 2012; Hamdi and Ismail-Hamdi 2014).
The 26 waterbird species comprised six distinctive foraging guilds that utilised water columns, banks, and associated grounds and grassland verges. Apart from favourable foraging prospects, this diverse avian assemblage witnesses that fence-enclosure and staff surveillance of the plant provided sufficient protection for waterbirds against various human interferences. In contrast, both formally protected Ramsar sites (e.g. Sakumo and Densu Delta) and unprotected coastal wetlands in the Accra Metropolis are prone to high levels of natural resource exploitation (Attuquayefio and Gbogbo 2001; Lamptey and Ofori-Danson 2014), thus negatively affecting waterbird food resources such as fish and crabs (Gbogbo et al. 2008). Furthermore, direct disturbances from hunting (Afdhal et al. 2012), predation from carnivores (Dean et al. 2015; Knapp et al. 2019), and other activities that distress wary species such as waterfowl have a negative impact (Gbogbo 2007b; Gbogbo et al. 2008). We therefore believe that the combined effect of a nutrient rich and safe environment (Dean et al. 2015) are prime factors for the rapid waterbird colonisation at LSTP. Constructed wetlands of this nature, even when located inland (Dean et al. 2015), may have positive implications for waterbird conservation in the urban areas of Ghana and West Africa in general, notwithstanding the poor conservation status of many formally protected wetlands in densely populated regions of Ghana (Attuquayefio and Gbogbo 2001; Nixon et al. 2007; Lamptey and Ofori-Danson 2014) and the sub-region (Adams 1993; Uluocha and Okeke 2004; Mitchell 2013).
The ASIP, under the Accra Metropolitan Assembly, initially planned for an additional similar STP adjoining the Densu Delta Ramsar site, but further constructions ceased prior to the LSTP completion. Therefore, LSTP is presently the only available waste stabilisation pond system in Accra and coastal Ghana, whilst other mechanical STPs have also been constructed. Based on the rapid colonisation, consistent and significant use of the whole plant by a diverse waterbird assemblage, we anticipate that the proliferation of similar STPs in urban and peri-urban Ghana, most likely will contribute positively to waterbird conservation, as well as minimising the negative effects of sewage discharge into natural waterbodies and the sea (Murray et al. 2014; De Troyer et al. 2016). Likewise, monitoring and research on waterbirds and their ecology at STPs, will provide more knowledge on the conservation impacts that these artificial wetlands have in rapidly growing coastal (Ashkenazi 2001; Harrison et al. 2010) or inland (Afdhal et al. 2012; Dean et al. 2015) urban zones, hereunder in West Africa. We therefore recommend a rapid expansion of STPs in Ghana and West Africa, and highlight the importance of future monitoring and assessment of the impacts such plants have on the abundance, diversity and distribution of waterbirds. State-owned lands, currently reserved for public green spaces and parks, including cemeteries and other recreational sites without housing potentials, could be earmarked for STP developments of variable size and inter-connectivity (Erwin 2002; Beatty et al. 2014). Areas adjoining governmental institutions and facilities, including the military, aviation in-flight zones, power lines and plants could also be integrated in networks of informally protected areas of CWs (Beatty et al. 2014), hereunder STPs (De Troyer et al. 2016; Knapp et al. 2019). Additionally, focus should be on the ecological and management implications of STPs for biodiversity complexity (Wiegleb et al. 2017), hereunder vulnerable species (Ashkenazi 2001; van Biervliet et al. 2020), intra- and interspecific interactions (Afdhal et al. 2012; Murray et al. 2014), including agonistic and interference competition (Harrison et al. 2010). Future important research aspects may include species’ susceptibility to microbial diseases (Murray et al. 2014), the potentials for heavy metal bioaccumulation (López-Perea et al. 2019), and for zoonosis transmission. Finally, we recommend research into STP management practices that reconcile effective water treatment with habitats that support a balanced and diverse waterbird fauna (Harrison et al. 2010; Dean et al. 2015; Knapp et al. 2019; van Biervliet et al. 2020).
Conclusion
We demonstrate that a relatively small constructed urban wetland situated far from the coast or other nearby large inland wetlands was subjected to rapid colonisation by a diverse assemblage of both migrant and resident waterbirds. Waterbirds benefitted from the rich food sources of invertebrates and plankton, in combination with informal protection by virtue of the restricted nature of the managed STP. We anticipate that the abundance and species diversity of waterbirds will increase as other trophic levels and community components of the biota matures, and waterbirds habituate to this protected wetland oasis amidst an otherwise densely populated urban coastal zone of West Africa. However, the persistence of diverse bird populations at similar STPs will depend on careful habitat management. Future long-term studies and population monitoring that encompasses the full calendar year could decipher such interactive relationships.
Data Availability
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Adams WM (1993) Use of wetlands and sustainable development in West Africa. The Geographical Journal 159(2):209–218 https://www.jstor.org/stable/3451412
Afdhal B, Charfi-Cheikhrouha F, Moali A (2012) Tunisian man-made wetlands as alternative habitats for waterbirds and their role for conservation. African Journal of Ecology 51:154–163. https://doi.org/10.1111/aje.12022
Ashkenazi S (2001) Long-term trends in the breeding populations of waterbirds (1951-1985) at a sewage treatment plant. Ostrich 72(1-2):10–19. https://doi.org/10.2989/00306520109485277
Attuquayefio DK, Gbogbo F (2001) Prospects of conserving wetlands along the Mukwe lagoon at Nungua in the Greater Accra region of Ghana. West African Journal of Applied Ecology 2:65–75. https://doi.org/10.4314/wajae.v2i1.45564
Aynalem S, Bekele A (2008) Species composition, relative abundance and distribution of bird fauna of riverine and wetland habitats of Infranz and Yiganda at southern tip of Lake Tana, Ethiopia. Tropical Ecology 49(2):199–209 ISSN 0564-3295. https://tropecol.com/pdf/open/PDF_49_2/11%20Aynalem.pdf
Beatty WS, Kesler DC, Webb EB, Raedeke AH, Naylor LW, Humburg DD (2014) The role of protected area wetlands in waterfowl habitat conservation: implications for protected area network design. Biological Conservation 176:144–152. https://doi.org/10.1016/j.biocon.2014.05.018
Belle JA, Andries J, Nacelle C (2018) Managing wetlands for disaster risk reduction: a case study of the eastern Free State, South Africa. Jamba: J Disas Risk Stud 10(1):a400. https://doi.org/10.4102/jamba.v10i1.400
Davidson NC (2014) How much wetland has the world lost? Long-term and recent trends in global wetland area. Marine and Freshwater Research 65(10):934–941. https://doi.org/10.1071/MF14173
Dayathilake D, Lokupitiya E, Wijeratne V (2021) Estimation of soil carbon stocks of urban freshwater wetlands in the Colombo Ramsar wetland city and their potential role in climate change mitigation. Wetlands 41:29. https://doi.org/10.1007/s13157-021-01424-7
De Troyer N, Mereta ST, Goethals PLM, Boets P (2016) Water quality assessment of streams and wetlands in a fast growing east African city. Water 8(4):123. https://doi.org/10.3390/w8040123
Dean WRJ, Milton SJ, Forsyth HP, Tissiman DR (2015) Fluctuations in bird numbers on sewage treatment ponds in an arid environment, South Africa. Ostrich 86(1-2):145–153. https://doi.org/10.2989/00306525.2015.1029559
Dixon A, Wood A, Hailu A (2021) Wetlands in Ethiopia: lessons from 20 years of research, policy and practice. Wetlands 41:20. https://doi.org/10.1007/s13157-021-01420-x
Douglas I, Alam K, Maghenda M, Mcdonnell Y, Mclean L, Campbell J (2008) Unjust waters: climate change, flooding and the urban poor in Africa. Environment and Urbanization 20(1):187–205. https://doi.org/10.1177/0956247808089156
Dowsett-Lemaire F, Dowsett RJ (2014) The birds of Ghana - a handbook and atlas. Tauraco Press, Liège, p 714
Dudgeon D, Arthington AH, Gessner MO, Kawabata Z-I, Knowler DJ, Lévêque C, Naiman RJ, Prieur-Richard A-H, Soto D, Stiassny MLJ, Sullivan CA (2006) Freshwater biodiversity: importance, threats, status and conservation challenges. Biological Reviews 81:163–182. https://doi.org/10.1017/S1464793105006950
Erwin MR (2002) Managing wetlands for waterbirds: Integrated approaches. Waterbirds 25(Spec Publ 2):5–12 https://www.jstor.org/stable/1522447
Erwin KL (2009) Wetlands and global climate change: the role of wetland restoration in a changing world. Wetlands Ecology and Management 17:71. https://doi.org/10.1007/s11273-008-9119-1
Finlayson CM (2012) Forty years of wetland conservation and wise use. Aquatic Conservation: Marine and Freshwater Ecosystems 22(2):139–143. https://doi.org/10.1002/aqc.2233
Froneman A, Mangnall MJ, Little RM, Crowe TM (2001) Waterbird assemblages and associated habitat characteristics of farm ponds in the Western cape, South Africa. Biodiversity and Conservation 10:251–270. https://doi.org/10.1023/A:1008904421948
Gbogbo F (2007a) Impact of commercial salt production on wetland quality and waterbirds on coastal lagoons in Ghana. Ostrich 78(1):81–87. https://doi.org/10.2989/OSTRICH.2007.78.1.12.56
Gbogbo F (2007b) The importance of unmanaged coastal wetlands to waterbirds at coastal Ghana. African Journal of Ecology 45:599–606. https://doi.org/10.1111/j.1365-2028.2007.00778.x
Gbogbo F, Attuquayefio DK (2010) Issues arising from changes in waterbird population estimates in coastal Ghana. Bird Populations 10:79–87 https://www.birdpop.org/docs/journals/Volume-10/BPJ10-09_Gbogbo_and_Attuquayefio.pdf
Gbogbo F, Oduro W, Oppong SK (2008) Nature and pattern of lagoon fisheries resource utilisation and their implications for waterbird management in coastal Ghana. African Journal of Aquatic Science 33(3):211–222. https://doi.org/10.2989/AJAS.2008.33.3.3.615
Gbogbo F, Oduro W, Oppong SK (2009) Response of waterbird species to fluctuating water levels in tropical coastal wetlands. African Journal of Ecology 48:637–643. https://doi.org/10.1111/j.1365-2028.2009.01156.x
Giosa E, Mammides C, Zotos S (2018) The importance of artificial wetlands for birds: a case study from Cyprus. PLoS One 13(5):e0197286. https://doi.org/10.1371/journal.pone.0197286
Hamdi N, Ismail-Hamdi S (2014) Testing whether artificial wetlands as habitats for waterbirds are good alternatives to natural wetlands: a case study of dams located in northern Tunisia. Vie et Milieu 64:47–57 ISSN 0240-8759. https://www.cabdirect.org/cabdirect/abstract/20153373721
Harebottle DM, Williams AJ, Weiss Y, Tong GB (2008) Waterbirds at Paarl waste water treatment works, South Africa, 1994–2004: seasonality, trends and conservation importance. Ostrich 79(2):47–163. https://doi.org/10.2989/OSTRICH.2008.79.2.4.578
Harrison JA, Williams AJ, MacIver M (2010) Breeding site selection by colonial waterbirds given various combinations of constructed or natural alternatives over a 10-year period. Ostrich 81(3):197–203. https://doi.org/10.2989/00306525.2010.519535
Holbech LH, Gbogbo F, Aikins TK (2018) Abundance and prey capture success of common terns (Sterna hirundo) and pied kingfishers (Ceryle rudis) in relation to water clarity in south-east coastal Ghana. Avian Research 9:25. https://doi.org/10.1186/s40657-018-0116-7
Hsu C-B, Hsieh H-L, Yang L, Wu S-H, Chang J-S, Hsiao S-C, Su H-C, Yeh C-H, Ho Y-S, Lin H-J (2011) Biodiversity of constructed wetlands for wastewater treatment. Ecological Engineering 37(10):1533–1545. https://doi.org/10.1016/j.ecoleng.2011.06.002
Jost L (2006) Entropy and diversity. Oikos 113(2):363–375. https://doi.org/10.1111/j.2006.0030-1299.14714.x
Jost L, Chao A, Chazdon RL (2011) Compositional similarity and beta (ß) diversity. In: Magurran AE, McGill BJ (eds) Biological diversity - frontiers in measurement and assessment, 1st edn. Oxford University Press, New York, pp 66–84
Junk WJ, An S, Finlayson CM, Gopal B, Květ J, Mitchell SA, Mitsch WJ, Robarts RD (2013) Current state of knowledge regarding the world’s wetlands and their future under global climate change: a synthesis. Aquatic Sciences 75:151–167. https://doi.org/10.1007/s00027-012-0278-z
Keddy PA, Fraser LH, Solomeshch AI, Junk WJ, Campbell DR, Arroyo MTK, Alho CJR (2009) Wet and wonderful: the world’s largest wetlands are conservation priorities. BioScience 59(1):39–51. https://doi.org/10.1525/bio.2009.59.1.8
Kivaisi AK (2001) The potential for constructed wetlands for wastewater treatment and reuse in developing countries: a review. Ecological Engineering 16:545–560. https://doi.org/10.1016/S0925-8574(00)00113-0
Kleijn D, Cherkaoui I, Goedhart PW, Hout J, Lammertsma D, Fuller R (2014) Waterbirds increase more rapidly in Ramsar-designated wetlands than in unprotected wetlands. Journal of Applied Ecology 51(2):289–298. https://doi.org/10.1111/1365-2664.12193
Knapp S, Schmauck S, Zehnsdorf A (2019) Biodiversity impact of green roofs and constructed wetlands as progressive eco-technologies in urban areas. Sustainability 11(20):5846. https://doi.org/10.3390/su11205846
Lamptey AM, Ofori-Danson PK (2014) Review of the distribution of waterbirds in two tropical coastal Ramsar lagoons in Ghana, West Africa. West African Journal of Applied Ecology 22(1):77–91 eISSN: 0855-4307. https://www.ajol.info/index.php/wajae/article/view/108001
López-Perea JJ, Laguna C, Jiménez-Moreno M, Martín-Doimeadios RCR, Feliu J, Mateo R (2019) Metals and metalloids in blood and feathers of common moorhens (Gallinula chloropus) from wetlands that receive treated wastewater. Science of the Total Environment 646:84–92. https://doi.org/10.1016/j.scitotenv.2018.07.265
Mitchell SA (2013) The status of wetlands, threats and the predicted effect of global climate change: the situation in sub-Saharan Africa. Aquatic Sciences 75:95–112. https://doi.org/10.1007/s00027-012-0259-2
Murray CG, Hamilton AJ (2010) Perspectives on wastewater treatment wetlands and waterbird conservation. Journal of Applied Ecology 47:976–985. https://doi.org/10.1111/j.1365-2664.2010.01853.x
Murray CG, Loyn RH, Kasel S, Hepworth G, Stamation K, Hamilton AJ (2012) What can a database compiled over 22 years tell us about the use of different types of wetlands by waterfowl in south-eastern Australian summers? Emu 112:209–217. https://doi.org/10.1071/MU11070
Murray CG, Kasel S, Szantyr E, Barratt R, Hamilton AJ (2014) Waterbird use of different treatment stages in waste-stabilisation pond systems. Emu 114:30–40. https://doi.org/10.1071/MU12121
Nixon SW, Buckley BA, Granger SL, Entsua-Mensah M, Ansa-Asare O, White MJ, McKinney RA, Mensah E (2007) Anthropogenic enrichment and nutrients in some tropical lagoons of Ghana, West Africa. Ecological Applications 17(5):Supplement:S144–Supplement:S164. https://doi.org/10.1890/05-0684.1
Ntiamoa-Baidu Y (1991) Seasonal changes in the importance of coastal wetlands in Ghana for wading birds. Biological Conservation 57(2):139–158. https://doi.org/10.1016/0006-3207(91)90135-V
Ntiamoa-Baidu Y, Piersma T, Wiersma P, Poot M, Battley P, Gordon C (1998) Habitat selection, daily foraging routines and diet of waterbirds in coastal lagoons in Ghana. Ibis 140:89–103. https://doi.org/10.1111/j.1474-919X.1998.tb04545.x
O’Connell M (2000) Threats to waterbirds and wetlands: implications for conservation, inventory and research. Wildfowl 51:1–15 https://wildfowl.wwt.org.uk/index.php/wildfowl/article/view/1076
Okes NC, Hockey PAR, Cumming GS (2008) Habitat use and life history as predictors of bird responses to habitat change. Conservation Biology 22(1):151–162. https://doi.org/10.1111/j.1523-1739.2007.00862.x
Otieno NE, Mutati AS, Akoth C, Ogwanjg D, Mwinami T, Alaro P, Njoka J (2015) Role of invertebrate prey abundance on waterbird distribution across rice field growth stages in Western Kenya. Waterbirds 38(1):47–57. https://doi.org/10.1675/063.038.0107
Runge CA, Watson JEM, Butchart SHM, Hanson JO, Possingham HP, Fuller RA (2015) Protected areas and global conservation of migratory birds. Science 350(6265):1255–1258. https://doi.org/10.1126/science.aac9180
Santoul F, Mastrorillo S (2004) Gravel pits as new wetlands for the Little grebe Tachybaptus ruficollis. Vie et Milieu 54(1):31–36 https://hal.sorbonne-universite.fr/hal-03217988/document
Scholz M, Lee B-h (2005) Constructed wetlands: a review. International Journal of Environmental Studies 62(4):421–447. https://doi.org/10.1080/00207230500119783
Schuyt KD (2005) Economic consequences of wetland degradation for local populations in Africa. Ecological Economics 53:177–190. https://doi.org/10.1016/j.ecolecon.2004.08.003
Shokoufeh S, Almuktar SAAAN, Scholz M (2021) Impact of climate change on wetland ecosystems: a critical review of experimental wetlands. Journal of Environmental Management 286:112160. https://doi.org/10.1016/j.jenvman.2021.112160
Sutherland WJ, Alves JA, Amano T, Chang CH, Davidson NC, Max Finlayson C, Gill JA, Gill RE, González PM, Gunnarsson TG, Kleijn D, Spray CJ, Székely T, Thompson DBA (2012) A horizon scanning assessment of current and potential future threats to migratory shorebirds. Ibis 154:663–679. https://doi.org/10.1111/j.1474-919X.2012.01261.x
Uluocha NO, Okeke IC (2004) Implications of wetlands degradation for water resources management: lessons from Nigeria. GeoJournal 61:151–154. https://doi.org/10.1007/s10708-004-2868-3
van Biervliet O, McInnes RJ, Lewis-Phillips J, Tosney J (2020) Can an integrated constructed wetland in Norfolk reduce nutrient concentrations and promote in situ bird species richness? Wetlands 40:967–981. https://doi.org/10.1007/s13157-019-01247-7
Verhoeven JTA, Arheimer B, Yin C, Hefting MM (2006) Regional and global concerns over wetlands and water quality. Trends in Ecology & Evolution 21(2):96–103. https://doi.org/10.1016/j.tree.2005.11.015
Vickery JA, Ewing SR, Smith KW, Pain DJ, Bairlein F, Škorpilová J, Gregory RD, Fox T (2014) The decline of afro-Palaearctic migrants and an assessment of potential causes. Ibis 156:1–22. https://doi.org/10.1111/ibi.12118
Wang H, Wang T, Zhang B, Li F, Toure B, Omosa IB, Chiramba T, Abdel-Monem M, Pradhan M (2014) Water and wastewater treatment in Africa - current practices and challenges. Clean: Soil, Air, Water 42:1029–1035. https://doi.org/10.1002/clen.201300208
Wang R, Wu F, Chang Y, Yang X (2016) Waterbirds and their habitat utilization of artificial wetlands at Dianchi Lake: implication for waterbird conservation in Yunnan–Guizhou Plateau Lakes. Wetlands 36:1087–1095. https://doi.org/10.1007/s13157-016-0823-y
Wiegleb G, Dahms H-U, Byeon WI, Choi G (2017) To what extent can constructed wetlands enhance biodiversity? International Journal of Environmental Science and Development 8(8):561–569. https://doi.org/10.18178/ijesd.2017.8.8.1016
Willoughby N, Grimble R, Ellenbroek E, Danso E, Amatekpor J (2001) The wise use of wetlands: identifying development options for Ghana’s coastal Ramsar sites. Hydrobiologia 458:221–234. https://doi.org/10.1023/A:1013158329107
Wymenga E, Zwarts L (2010) Use of rice fields by birds in West Africa. Waterbirds 33(special Publication 1):97–104. https://doi.org/10.1675/063.033.s107
Acknowledgements
Firstly, great thanks to Mr. Alfred Ali Nuoh, Centre for African Wetlands, University of Ghana, who assisted in the identification and counting of birds at the LSTP. Secondly, we are grateful to the Managing Director, Mr. Deo Datus Appiah, Val Terra Ltd. (Tema), in liaison with the Accra Metropolitan Assembly, for granting permit, as well as encouragement of the work. Also great thanks to an anonymous editorial assistant, and two reviewers for their constructive criticism and editing of the manuscript. Finally, thanks to Dr. William Gblerkpor for assistance with drone imaging at the LSTP.
Code Availability
Not applicable.
Funding
This study did not receive any funding from any source or institution.
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study, and read and approved the final manuscript; LHH conceptualised, designed and supervised the study, participated in the initial data collection, performed the majority of data analysis, initiated the draft, and prepared the final manuscript; CCC conducted the majority of data collection, and made contributions to material preparation and data analysis.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Ethics Approval
Prior consent and permissions were obtained from respective authorities.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Holbech, L.H., Cobbinah, C.C. Pollution or Protection - What Early Survey Data Shows on Rapid Waterbird Utilisation of a Newly Established Sewage Treatment Plant in Urban Ghana, West Africa. Wetlands 41, 110 (2021). https://doi.org/10.1007/s13157-021-01510-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-021-01510-w