
Abstract
Ecosystem-atmosphere exchanges of carbon dioxide (CO2) and water vapor were investigated in a moist mountain meadow (Loney Meadow) at 1822 m MSL in the Sierra Nevada, California, USA. An eddy covariance (EC) tower was deployed for most of the snow-free period from May to September 2016. The meadow ecosystem progressed from a strong sink of CO2 in the peak of the growing season under saturated to wet soil conditions (−18.51 gC m−2 d−1) to a weak source (2.97 gC m−2 d−1) following a rapid decline in soil moisture as runoff decreased. The variability of Net Ecosystem Exchange (NEE) over diurnal, synoptic and seasonal timescales was dominated by Gross Primary Production (GPP) which ranged from 43 gC m−2 d−1 during the peak of the growing season to 19 gC m−2 d−1 during senescence. Ecosystem respiration was small in magnitude and variability compared to GPP. Approximations of annual NEE for the meadow ranged from −285 to −450 gC m−2 yr−1, which is high compared to grasslands, and more similar to mature wetland or forest ecosystems. At diurnal and synoptic scales, CO2 flux was driven most strongly by photosynthetically active radiation (PAR), while seasonally, the ecosystem was linked closely to changes in soil moisture. Light-use and water-use efficiencies of the meadow ecosystem were high compared with those found in most other ecosystems using comparable observations. These results suggest meadows have the potential to be large sinks of atmospheric CO2 and that their ability to do so is sensitive to water table height. This is important for understanding the future of carbon sequestration in mountain meadows in the context of changing hydroclimates and different land management decisions that impact meadow hydrology.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mountain meadows are important environmental systems that form in topographical depressions and low-gradient valley bottoms, often containing shallow groundwater, finely textured and organically rich soils and abundant plants dominated by hydric to mesic herbaceous species. In the Sierra Nevada (SN) range of California and Nevada, USA, Viers et al. (2013) estimated that there were more than 17,000 meadows covering nearly 78,000 ha of land. Although they account for a relatively small percentage of land cover (~0.01%), most tributaries pass through multiple sequences of meadows within SN watersheds, which have important water quality and discharge controls.
There is a significant amount of diversity among SN meadows resulting in a wide range of hydrogeomorphic types ranging from peatlands and depressions to more riparian systems (Weixelman et al. 2011). Hydrologic inputs to local meadow aquifers include springs from geologic aquifers, hillslope runoff and surface streams (Loheide et al. 2009). Ecological and rangeland classifications are strongly influenced by hydrology, with wetland obligate or facultative plant assemblages dependent on water table depth and the amount of time water covers the meadow surface (Ratliff 1985; Allen 1987; Dwire et al. 2006). A shallow groundwater table sustaining high soil moisture levels through much of the growth period is the most important factor in maintaining the characteristic herbaceous communities found in wet meadows (Fites-Kaufman et al. 2007; Loheide et al. 2009). With seasonal runoff influenced greatly by snowmelt, SN meadows produce strong seasonal hydrologic and ecologic cycles, behaving much like seasonal wetlands (Ratliff 1985; Loheide et al. 2009).
Vegetation in undegraded meadows typically consists of hydric and mesic species such as perennial grasses, wet sedges, forbs, and other herbaceous species (Ratliff 1985; Allen 1987; Lowry et al. 2011; Maher 2015). The spatial patterns of vegetation within individual meadows are indicative of an extensive land-water ecotone driven by soil moisture gradients (Kondolf et al. 1996).
Ecosystems in meadows with significant riparian and hillslope water sources exhibit strong seasonality with annual growth cycles that begin in the late spring due to snowmelt-induced runoff, senesce from mid to late summer as water tables lower, and often snow covers meadow surfaces in winter (Loheide and Gorelick 2007). The timing of spring snowmelt, peak streamflow, snow cover and the relative seasonal snow water equivalent (SWE) in the SN are influenced by inter-annual and decadal-scale climate variability (Hamlet et al. 2005; Stewart et al. 2005). In general, over the past five decades, the timing of spring streamflow in western North America has been arriving earlier and with less precipitation falling as snow (Stewart et al. 2005; Lowry et al. 2011; Viers and Rheinheimer 2011). Previous studies of moist meadow carbon cycling have primarily focused on carbon stocks from biometric estimates (Norton et al. 2011; Reed et al. 2020) but much less is known about the rates of CO2 exchange, its seasonal evolution and its main environmental controls (Fites-Kaufman et al. 2007).
Mountain meadows are also sensitive to land use changes and many of these ecological processes have been altered as a result of anthropogenic activities (Ratliff 1982; Kattelmann and Embury 1996; Purdy and Moyle 2006; Loheide and Gorelick 2007). Historic and current land use (e.g., grazing, logging and mining) in the Sierra Nevada have contributed to increased stream channel incision in riparian meadows (Ratliff 1985; Kattelmann and Embury 1996; Purdy and Moyle 2006; Viers et al. 2013; Lowry et al. 2011; Weixelman et al. 2011). This channel incision can disconnect the stream channel from the meadow floodplain, lowering the water table and reducing soil water content in the root zone, which impacts the productivity and distribution of native vegetation (Kattelmann and Embury 1996; Loheide et al. 2009; Lowry et al. 2011). Lowering of the water table also allows oxygenation of organically rich soil, which enhances soil microbial respiration resulting in the accelerated loss of stored soil carbon (Scott et al. 2010; Knox et al. 2015).
The impact of degradation on vegetation patterns tends to be a succession from native hydric/mesic species to more xeric species commonly associated with dryland meadows (Allen-Diaz 1991; Loheide and Gorelick 2007; Loheide et al. 2009; Pope et al. 2015). Previous studies have linked water availability in meadows to fluctuations in species richness, vulnerability to invasive species encroachment and the capacity to sequester atmospheric CO2 for plant production and contribute to soil carbon storage (Dwire et al. 2006; Fites-Kaufman et al. 2007; Haugo and Halpern 2007; Blankinship and Hart 2014; Maher 2015). A comparison of existing measurements of CO2 fluxes in wetland, grassland and semi-arid ecosystems suggests that hydric/mesic species are much faster growing and absorb greater amounts of CO2 from the atmosphere (e.g. Ratliff 1985; Flanagan et al. 2002; Kayranli et al. 2010; Norton et al. 2011). Xeric species, on the other hand, tend to be weak sinks of carbon and can shift from a net sink to a source of carbon dioxide to the atmosphere during dry years (Lund et al. 2010; Scott et al. 2010). Because of the important ecological benefits and services that mountain meadows provide, such as water quality and flood control, carbon sequestration and storage, biodiversity enhancement, and culturally important food and weaving resources, there has been increased interest in restoring meadows that have been degraded due to anthropogenic activities (Loheide and Gorelick 2007; Pope et al. 2015). The goal of these “rewatering” projects is to mimic and restore natural processes that raise the volume of subsurface storage by providing a greater spatial opportunity for water to infiltrate (Hammersmark et al. 2008). One of the implications of restoring meadow hydrology is to increase plant productivity rates as well as suppress microbial respiration, leading to an increase in soil carbon storage.
Despite the significant attention to mountain meadow watershed assessment and restoration over the past two decades, relatively few studies have investigated carbon cycling in mountain meadows directly. The FLUXNET global network of eddy flux towers has so far produced hundreds of site-years reflecting most major biomes of the world, but has produced relatively few studies of detailed carbon fluxes in mountain meadows (Oliphant 2012). The objective of this study is to investigate the carbon and water cycles of a mountain meadow ecosystem throughout the growing season. The meadow selected (Loney Meadow) is in the South Yuba River watershed in the Northern Sierra Nevada, California, USA, at about 1800 m elevation. Specifically, the study aims to use high-frequency, meadow-scale eddy covariance observations over a 5 month period of the growing season to (a) investigate meadow ecosystem functioning over timescales from diurnal to multi-day to seasonal, (b) assess the key environmental controls on ecosystem functioning, particularly the ability of the ecosystem to sequester atmospheric CO2 and (c) approximate the annual total meadow CO2 exchange. The results from this study are compared to the limited results published from other meadow ecosystems as well as wetlands and grasslands for context, and implications for the role of climate and land use changes on meadow carbon sequestration are discussed.
Biophysical Setting and Methods
Biophysical Setting
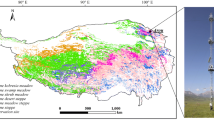
Loney Meadow is located on the upper western flank of the Northern Sierra Nevada in the headwaters of the South Yuba River watershed, which drains west to the Sacramento River (Fig. 1). The meadow has a surface area of approximately 138,000 m2 and is located at 39.421°N, −120.655°W, with an elevation near the center of 1822 m MSL. The main Loney Meadow investigated in this research is a riparian low-gradient type in the Weixelman et al. (2011) classification. We expect that its hydrology is supported by surface streamflow inputs, hillslope runoff, and geologic groundwater sources; a likely input of non-gaged snowmelt-fed streams and groundwater is suggested by an 83% increase in discharge between the inflow and outflow gauge reported in 2016 from Hutchinson et al. (2020) while the contributing catchment area is only 20% greater (5.75 km2 at the outflow as compared with 4.77 km2 at the inflow).

Study site characteristics and eddy covariance flux measurement location including a visible image of the meadow extent (source: Esri, DigitalGlobe, GeoEye, Earthstar Geographics, CNES/Airbus DS, USDA, USGS, AeroGRID, IGN, and the GIS User Community) and a normalized difference vegetation index (NDVI = (NIR-red)/(NIR + red)) image of the meadow ecosystem within the main flux source area created from a MicaSense RedEdge radiometrically calibrated 5-band camera mounted on a 3D Robotics Solo drone flown at 80 m above ground level with 75% overlap on July 16, 2017, processed with Pix4D Mapper to produce 5-cm pixels of reflectance in blue (20-nm band centered at 475 nm), green (20-nm band centered at 560 nm), red (10-nm band centered at 668 nm), near IR (40-nm band centered at 840 nm), and red edge (10-nm band centered at 717 nm) (Davis et al. 2020)
Loney Meadow experiences mountain Mediterranean climate conditions with warm dry summers and cold wet winters. Following a three-year period of severe drought conditions in California, the 2015–16 water year experienced closer to average levels of precipitation, snowfall and temperature. Total precipitation for the 2015–16 water year of 194 cm was recorded at the Bowman Dam monitoring station located approximately 2.4 km from Loney Meadow, which was 15% higher than the long-term average between 1896 and 2020. Snowfall (454 cm) however, was 24% lower than the long-term average and temperatures were slightly above average. Daily average temperatures at Bowman Dam range between −3 °C during the winter and 26 °C in the summer with snow typically covering the ground from October to May and peak snow depth occurring in March (WRCC 2020). In a comparison of 13 northern California montane meadows studied by Reed et al. (2020), Loney was among the wettest and most productive. In 2016, the average height of groundwater (relative to a ground surface datum) using an array of 10 piezometers in Loney Meadow during the summer recession (June – September) was +0.1 m, with a minimum of −1.83 m and a maximum of +1.47 m (Hutchinson et al. 2020; Reed et al. 2020).
The vegetation of Loney Meadow is dominated by mixed graminoids and forbs, consistent with hydric to mesic meadow communities elsewhere in the Sierra Nevada. Vegetation sampling conducted across Loney Meadow in 2016 identified more than 120 species (Reed et al. 2020). Five of the 10 most abundant species observed were graminoids, four sedges (Carex utriculata (10.1% cover), C. senta (6.6%), C. spectabilis (5.1%), C. nebrascensis (4.9%) and the California Brome (Bromus carinatus - 4.7%). Other abundant species included perennial grasses and herbs such as Symphyotrichum spathulatum (9.8%), Juncus balticus (5.8%), Potentilla gracilis (5%), Perideridia lemmonii (3.3%), and Achillea millefolium (2.8%). Reed et al. (2020) assessed that the species observed in Loney Meadow in 2016 were approximately 20% obligate wetland species and 54% facultative wetland species. Furthermore, they reported biomass estimates obtained for 2015–16 in Loney Meadow, which averaged 150 g m−2 for above ground biomass and root carbon of 2.2 kg m−2. These observations were made in a grid pattern across the meadow surrounding the tower location, with all corners of the grid within 120 m of the flux tower location for this study, so they provide a representative sampling of the species that the flux tower ‘observes’ (Section 2.2).
Based on regular site visits and daily images captured from a digital camera mounted at the site, the meadow ecosystem transitioned through four distinct phases over the observation period from May 17th to September 6th, 2016 (Table 1). The first period saw the emergence from melting snow cover of grasses, sedges and small forbs, with rapid growth. During the early spring season, ponding occurring on over 75% of the surface, and a significant storm event provided snow cover for several days over the emerging vegetation. By July, vegetation height and density appeared to reach a maximum, and this was followed by a long period of senescence. From early July to early August, this included a changing of vegetation color with limited decrease in height and density. By the end of August, a significant decline in plant density and height had occurred and negligible green vegetation was visible (Table 1).
Loney Meadow vegetation was grazed by a small herd of cattle between late June and September 2016. Approximately 50 cow and calf pairs were released into the meadow at the end of June and were left to graze a large area around and including Loney Meadow until the end of September. The main observable impact of cattle grazing was the reduction in above ground biomass, especially during the latter part of summer when new growth declined, and the addition of manure on the surface. Cattle density on the meadow remained fairly low, and we did not see the creation of bare patches or undue compaction, although channel scouring evident in several places was likely exacerbated by their presence.
The NDVI image of the meadow in the vicinity of the tower (Fig. 1) was captured in the peak of the growing season using a drone-mounted multispectral camera (Davis et al. 2020), illustrating the high values from the meadow vegetation (yellow to green), and the low values of rocks and water. The latter indicates both the location of meadow channels and the presence of standing water (particularly to the SW of the tower). This area contained emerging vegetation and would show high NDVI if re-imaged once the standing water subsided, as the summer progressed. The image was captured in 2017, following a wetter winter than in 2016, when the reported measurements were made. The standing water was evident in the same location in 2016, but approximately a month earlier in the melt season.
The soil organic matter (SOM) for Loney Meadow was found to average 234 g kg−1 with a standard deviation of 59 g kg−1, from 19 samples, which were collected in the 5-15 cm layer, randomly sampled from an area of about one hectare near the center of the meadow at different times during the summer of 2016. These values were obtained using the loss on ignition (LOI) technique, with soils subjected to 360 °C temperatures for 2 h, after 24 h of drying at 105 °C. Roche et al. (2014) found similar but slightly higher values for the same depths across nine ‘moderately wet’ and ‘wet’ montane meadows in the Sierra Nevada. These values are also similar to and slightly higher than those found by Maher (2015) for a restored montane meadow. Both of these previous studies also sampled dry or degraded meadows, which were universally lower in SOM than was found for Loney Meadow. Additionally, Hutchinson et al. (2020) reported values of approximately 7.4 kg m−2 soil carbon averaged over 2016 and 2017 for Loney Meadow, and these values were similar to those found elsewhere in Northern California mountain meadows (e.g. Plumas Corporation 2020). The U.S. Forest Service (USFS) and National Fish and Wildlife Foundation (NFWF) had identified Loney Meadow as ‘degraded’ at the time of this study, and low-impact restoration work to help reduce channel discharge in several locations was conducted in the following year. This appears to have resulted in wetter habitat conditions based on results of hydrologic, soil and vegetation monitoring over the period 2015–2019 (Hutchinson et al. 2020). Nevertheless, based on the abundant hydric plant species, and significant soil carbon stocks observed at the time of this study, we expect the meadow ecosystem was acting as a net carbon sink overall.
Experimental Design
A micrometeorological observation system with eddy covariance (EC) instruments (flux tower) was deployed in the meadow between May 17th and September 6th, 2016. The location of the flux tower slightly to the west of center, was selected in order to ensure reasonable measurement length across the meadow surface during both the dominant westerly up-valley winds observed during the day and down-valley drainage flows from the east at night, which produce a longer measurement footprint due to the stable nocturnal boundary layer (Fig. 1). The eddy covariance system was comprised of a 3-dimensional sonic anemometer-thermometer (CSAT3, Campbell Sci., Logan, Utah, USA) and an open path infrared gas analyser (Li7500, LiCor Inc., Lincoln Nebraska, USA), which were deployed at 2.44 m above ground level (approximately four times the maximum canopy height). These were sampled at 10 Hz by a CR3000 data logger (Campbell Sci., Logan, Utah, USA). In addition, 30-min average radiation fluxes were measured using a four-component radiometer (NR01, Hukseflux, Delft, The Netherlands) deployed at 1.2 m and air temperature and relative humidity were measured at 2.44 m using a HMP45C probe (Vaisala Corp., Helsinki, Finland). A tipping bucket rain gauge (TR-5251 Texas Electronics, Dallas, Texas, USA) was mounted at 0.5 m. The soil heat flux was determined from the average of direct measurements from a pair of heat flux plates (HF01, Hukseflux, Delft, The Netherlands) installed at a depth of 5 cm combined with heat storage change estimated for the soil layer above the plates from four spatial averaging thermocouples inserted into this layer. Mean soil temperature was also determined at 2 and 10 cm depths using CS107 probes (Campbell Sci. Inc.) and average soil moisture in the 0–15 cm depth range was determined using a CS616 TDR probe (Campbell Sci. Inc.). In addition, a Moultrie game camera was attached to the center pole of the tower at 2.2 m facing west and captured daily (midday) images of the meadow surface.
Data Processing, Rejection and Uncertainty
EC-derived fluxes of CO2, water vapor and heat were calculated from 30-min covariance blocks, after removal of spikes in the high frequency data. These fluxes were corrected for density fluctuations (WPL corrections) and planar-fit coordinate rotations (Lee et al. 2004). The distribution of the flux source area in the upwind direction was calculated for each 30-min period using the analytical footprint model of Hsieh et al. (2000).
Eddy covariance measurements have been shown to underestimate the flux under conditions of low turbulent energy (Massman and Lee 2002; Papale et al. 2006; Burba 2013). The friction velocity (u*) threshold for rejection varies based on the ecosystem being sampled and typically ranges between 0.05 and 0.2 m s−1 (Massman and Lee 2002). Using the method of Papale et al. (2006) to determine a site-specific u* threshold, the rejection threshold for this site was established at u* > =0.1 m s−1. This criterion caused the most frequent rejection of data, with a strong bias toward nocturnal hours. Loney Meadow also provides a challenge for EC measurements due to its relatively small size. The meadow boundary was defined from analysis of satellite imagery (Fig. 1) and the radial distance from the tower to the meadow boundary was evaluated for 21 directions. For each 30-min flux, data were defined as acceptable if the 90th percentile of the source area distance in the upwind direction was less than the distance to the meadow boundary. This ensured accepted flux data were representative of the meadow plant communities. Over the entire study period, the 90th percentile of the flux source in the upwind direction ranged from a few meters to more than 600 m, though averaged 90 m, with a standard deviation of 49 m. Thus, the eddy covariance CO2 and H2O flux observations mostly represent the same area in which plant sampling was conducted, and therefore represents biophysical processes driven by the plant community described in Section 2.1.
Understanding uncertainty in EC observations can be additionally challenging in complex terrain, particularly with heterogeneous vegetation and during calm conditions when local scale atmospheric circulations dominate (Castellví and Oliphant 2017). Though these are generally larger than the flux footprint scale, they potentially add a local scale signal to vertical transport that is unmeasured by EC. Since all components of the surface energy balance were directly measured (assuming heat storage changes were minimal in the short meadow vegetation), closure of the energy balance was used as an independent check on the quality of the EC measurements. This is normally assessed by the linear relationship between the combined EC-derived heat fluxes (sensible and latent heat flux) and available energy (difference between net radiation and ground heat flux) on a 30-min basis (e.g. Wilson et al. 2002). In this case, the slope of the linear model for all acceptable data was 0.67 (r2 = 0.88). These values did not change significantly when applied to four subsets of data representing the four different phenological stages identified in Table 1. This slope value is slightly lower than average but well within the distribution of closure estimates from synthesis studies comparing multiple sites (e.g. Wilson et al. 2002) and were similar to other studies in complex terrain (Stoy et al. 2013), especially a similar meadow in the Sierra Nevada (Castellví and Oliphant 2017). The coefficient of determination as well as the slope value suggests that the EC fluxes presented here are strongly correlated with the actual turbulent fluxes but consistently underestimate them.
Volumetric soil water content was measured using a Time Domain Reflectometry probe (TDR), with 30 cm probe lengths, which were inserted at a 30° angle relative to the surface, thus producing an average measurement in the layer 0–15 cm. The probes were carefully inserted into firm soil ensuring good contact between the soil and probes along their entire lengths. Well-sited TDR probes provide an excellent record of soil moisture change over time, although site calibration is recommended to control the absolute magnitude, particularly in more porous soils and those with high organic content (Zegelin et al. 1992). Gravimetric soil water content was determined during site visits from 16 soil samples collected in random locations within 100 m of the TDR probe/flux tower at a depth of 5–15 cm with a hand trowel and aluminum soil tins. To convert these to volumetric soil water content for comparison with the TDR, we used a bulk density of 0.54 Mg m−3 based on observations by Baccei et al. (2020) and Reed et al. (2020) in numerous Sierra Nevada meadows. Resulting volumetric water content ranged from 50% during the early wet phase when standing water was observed nearby to around 10% in early September and comparison with the TDR probe for the five soil sampling periods produced a linear fit with a zero offset, a slope of 0.98 and a coefficient of determination of 0.98.
Partitioning and Gap Filling CO2 Exchanges
In carbon budget terms, the 30-min EC-derived CO2 flux equates to the net ecosystem exchange of CO2 (NEE, mgC m−2 s−1). NEE is the net result of two much larger and directionally opposing CO2 exchange processes; gross primary production by photosynthesis (GPP) which causes an atmosphere-to-ecosystem flux of carbon, and ecosystem respiration (RE) which causes an ecosystem-to-atmosphere flux,
Eq. 1 produces the meteorological sign convention for NEE where positive values indicate a net ecosystem source of CO2 to the atmosphere and negative values indicate a net ecosystem sink. The relative contributions of GPP and RE to an eddy covariance measurement of NEE are not directly observed by EC. However, since these plant species require light for photosynthesis, it can be assumed RE ≈ NEE (observed) during nocturnal hours when PAR is zero. Many studies have shown that soil temperature has a strong correlation with respiration, particularly in grasslands, but that the relationship varies with soil moisture as well as seasonal changes in plant phenology (Gilmanov et al. 2005; Papale et al. 2006; Reichstein et al. 2005). The relationship between nocturnal NEE values and bin averaged soil temperatures (Ts) was evaluated, and best described by a linear fit;
In order to account for the impact of soil moisture, which declined throughout the period as well as changes in phenology and biomass, the model was determined for four seasonal periods independently (as defined in Table 1). The slope of the model increased significantly throughout the summer from 0.01 to 0.019. Using the period-dependent models, 30-min RE values were calculated for the entire study. These derived RE values were used to gap-fill daylight periods and to replace data rejected using the QC criteria outlined above.
For daylight hours, observations of NEE were combined with modeled RE, to calculate GPP by residual using Eq. 1. A commonly used empirical light use efficiency model based on a rectangular hyperbola (Eq. 3) was then fit to the observed data using PAR and accepted daylight GPP estimates at the 30-min timescale (after Xu and Baldocchi 2004; Gilmanov et al. 2007; Oliphant et al. 2011), such that
where the coefficient α is the initial slope of the light-use efficiency (LUE) curve and Amax is the point of maximum carbon assimilation. Since LUE is expected to change through the growing season due to changes in leaf area density and chlorophyll concentration, independent models were generated and applied to the four distinct phases of the growing season identified in Table 1.
Results
Diurnal Patterns and Drivers of Ecosystem CO2 Exchanges and Evapotranspiration
Based on ensemble averages over the observed growing season, Fig. 2 shows that the ecosystem was a strong and dynamic net sink of CO2 from the atmosphere during the day, and a more consistent and weaker source of CO2 to the atmosphere at night. Daylight NEE values were closely anti-correlated with PAR, with an immediate response (decreasing NEE) following sunrise. The morning switch from source to sink occurred on average 45 min later, and peak sequestration rates of approximately −0.6 mgC m−2 s−1 on average occurred by 11:30 PDT, approximately 90 min before solar noon. The net ecosystem exchange switched from a sink to a source approximately 75 min before sunset, reaching its peak soon after sunset. Though clearly driven by the symmetrical cycle of PAR, the diurnal pattern of NEE indicates a stronger response (sink) in the morning than the afternoon for the same light levels. Nocturnal NEE values remained relatively steady, declining from a little over 0.2 mgC m−2 s−1 to a little under by the end of the night (Fig. 2). The inter-diel variability of NEE is greater during the mid-day hours of peak uptake than through the night, suggesting more day-to-day control on CO2 flux variability in photosynthesis than respiration over the growing season. The sum of the diurnal ensemble 30-min averages for the observational period was −6.1 gC m−2 d−1, similar to mature forests in the peak of their growing season (e.g. Baldocchi 2008).

Diurnal time series of 30-min ensemble averages ± one standard deviation (error bars) of (a) PAR, (b) NEE and (c) ET observed at Loney Meadow for the entire observation period
Evapotranspiration (ET) rates began to climb significantly about an hour after sunrise, with the lag closely following the switch in net radiation from negative to positive (not shown). However, ET thereafter followed a more symmetrical correlation with PAR than NEE, so the loss of water relative to carbon uptake was less during the morning than afternoon hours. The relatively lower ET in the morning is probably related to greater atmospheric demand for water in the warmer and drier afternoon hours relative to the morning hours for the same levels of PAR. There was also a secondary small peak in mean ET around sunrise as well as a much larger inter-diel variability. Assessment of the raw data reveal that this resulted from small spikes on some mornings at this time, suggesting evaporation of dewfall, which was also visually observed occasionally during site visits. Although close to zero at night on average, the standard deviation bars for ET descending below zero suggests that water deposition on the leaves occurred on some nights.
Decomposition of the diurnal cycle into partitioned CO2 fluxes, as well as key environmental drivers for four different seasonal periods are shown in Fig. 3, with daily magnitudes provided in Tables 1 and 2. The diurnal pattern of NEE was clearly driven most strongly by GPP, which was closely correlated with PAR. The diurnal pattern of GPP remained mostly symmetrical in each of the four seasonal periods, though at very different magnitudes throughout the growing season. Relative to the pattern of PAR, there is a flattening during the mid-day hours, suggesting a lower light use efficiency during those hours. In the senescent period, this flattening occurred from about 1100 to 1700 PDT and peaked at only 0.5 mgC m−2 s−1, compared with a diurnal peak of 1.26 mgC m−2 s−1 during the period of strongest growth (Period 2). Differences within the growing season can be less easily explained by PAR. The emergent phase had the 2nd highest GPP but the lowest PAR, and the senescence onset period (Period 3) had the highest available PAR but produced a 33% decline in diurnal peak GPP. This shift coincided with a significant reduction in volumetric water content (VWC), and an increase in vapor pressure deficit (VPD).

Diurnal ensemble 30-min averages of GPP (a), RE (b), PAR (c), soil temperature (TS) (d), vapor pressure deficit (VPD) (e), and volumetric water content (VWC) (f) according to seasonal period, Loney Meadow, 2016
RE followed the somewhat asymmetrical diurnal cycle governed by soil temperature but also showed distinct seasonal differences (Fig. 3c). The diurnal range in RE is only about one quarter of the range in GPP. On a seasonal basis, the emergent phase exhibited the lowest RE rates. This was also the period with the lowest soil temperature (Fig. 4e) and highest soil water content (Fig. 4f). The high water table during this period (water flowed across majority of the meadow) would likely suppress root and soil respiration as with other wetlands (e.g. Knox et al. 2015). The three following seasonal periods had similar daily average RE rates to each other, and about 20% higher than the wet emergent phase (Table 2). However, the peak value of RE consistently increased as the season progressed, reaching a maximum of about 0.38 mgC m−2 s−1 during the senescent phase (Fig. 3b).

Daily total ecosystem CO2 and H2O exchanges and environmental conditions at Loney Meadow during the 2016 growing season. At top is time sequence of approximately weekly images of meadow surface taken from 2.2 m a.g.l., west facing at noon, and illustration of the four seasonal periods distinguished
Seasonal and Weather Controls on Ecosystem CO2 Exchange
Daily total NEE fluctuated significantly both at seasonal and synoptic scales, but resulted in a net sink of CO2 on 82 of the 112 days measured (Fig. 4). The emergent period until May 19th (DOY 160) produced a decline in NEE (increasing sink of CO2) as leaf area index (LAI) rapidly increased, punctuated by sharp increases in NEE in response to a significant early summer storm. The impact of the storm resulted in the ecosystem switching from a net sink to source of CO2 on two of the days. During the storm, PAR decreased significantly (88%) due to deep cloud cover, which coincided with a similarly large decline in GPP (80%). Daily average air temperature dropped 12 °C and soil temperature dropped 8 °C, which coincided with a 42% decline in RE (Fig. 4). Since the meadow was nearly saturated at this time, the main hydrologic impact on NEE was the partial coverage of the meadow with snow, which would have shielded emerging vegetation from PAR and helped suppress soil and plant respiration. From Day 160 (May 19th) to Day 187 (July 5th) NEE reached its peak uptake and remained fairly constant, with weather disturbances having a smaller impact. Throughout this period PAR remained high, volumetric soil water content was above 40% and very high daily totals of carbon sequestration occurred (−20 and − 25 gC m−2 d−1). This period was also characterized by maximum vegetation height and density (Table 1).
From Day 187 (July 5th) a steady rise in NEE began, which continued to the end of the study, and resulted in the meadow ecosystem switching from a net sink to a source on a daily basis around Day 224 (August 11). Despite maintaining high values for PAR, this rise in NEE coincided with decreasing volumetric soil water content from 40% to 10%, and appeared to be directed by changes in GPP, which declined steadily throughout this period. Senescence is also evident in shift in vegetation color from daily surface images (Fig. 4). Respiration by comparison remained fairly constant throughout this period. A small rain event (< 1 mm) occurred on August 18 (Day 231), but had little to no impact on recorded soil moisture levels (Fig. 4c & d). At this point, most of the vegetation had become insensitive to light levels and productivity was constrained by water availability. Throughout the growing season, ET more closely followed GPP than soil moisture, suggesting the dominance of transpiration. However, during cloudy periods, as indicated by PAR in Fig. 4b, ET declined more significantly than GPP, raising the water-use efficiency on those days.
In comparison to other meadow ecosystems surveyed using the EC method, the daily CO2 flux magnitudes found in Loney Meadow (averaging −18.51 gC m−2 d−1) were significantly higher (larger negative value). A seasonal peak daily total CO2 flux of −3.9 gC m−2 d−1 was observed in an alpine meadow on the Tibetan Plateau (Kato et al. 2004), −6.3 gC m−2 d−1 for a meadow steppe ecosystem in Northeast China (Dong et al. 2011) and − 7 gC m−2 d−1 for a montane meadow at 1450 m MSL in the Sierra Nevada, California following a very dry winter (Maher 2015). Loney Meadow was also among the most productive ecosystems in comparison to grasslands and wetlands observed elsewhere. For example, the highest recorded daily CO2 exchange in a young wetland studied by Knox et al. (2015) was about −11 gC m−2 d−1 and a survey of wetlands by Lund et al. (2010) showed maximum daily total NEE values ranging between −1 and − 4 gC m−2 d−1. Grasslands exhibit a high degree of variability in peak daily total CO2 values ranging between −5 (Flanagan et al. 2002; Xu and Baldocchi 2004) and − 50 gC m−2 d−1 (Dugas et al. 1999). The latter noted that daily fluxes lower than −20 gC m−2 d−1 are typically rare and generally short-lived.
Light, Carbon and Water Relationships in the Meadow Ecosystem
The coefficients and statistics of rectangular hyperbola LUE curve fitting are provided in Table 3. Both the initial slope of the curve (α value) and the point of maximum CO2 assimilation (Amax) are high compared to grasslands, especially during the emergent and peak growth phases. In a comparison, LUE of twenty European grasslands, Gilmanov et al. (2007) reported α values ranging from 0.016 to 0.075 and Amax values ranging from 42.5 to 216 μmol m−2 s−1, with the higher values attributed to wetter grasslands. The light response parameters observed in Loney Meadow for the full observation period was close to the maximum of these grassland sites and exceeded them all during the peak growth phase. The beginning of senescence produced a decline in both the magnitude and consistency in LUE and the late senescence showed heavily suppressed photosynthesis and a largely disconnected relationship with PAR.
The relationships between GPP and both PAR and ET at the daily timescale are presented in Fig. 5. Although the magnitude of GPP was generally much higher during the emergent and peak growth phases than the senescent phases (by 12–18 gC m−2 d−1), the slope of the relationships remained fairly similar. This suggests the ecosystem maintained fairly consistent light use and water use efficiencies throughout the growing season, despite operating at very different levels of productivity. By comparison with similarly defined values for WUE reported elsewhere, the fairly consistent value of about 5.5 g kg−1 found for Loney Meadow is quite high. In a summary of WUE from 43 different ecosystems, similar values were found for deciduous broadleaf and mixed forests, but grasslands were on average significantly lower (Beer et al. 2009). In this comparison, Loney Meadow would be ranked 2nd of 43. However, a steppe meadow ecosystem in Northeast China produced similar mean WUE values (Dong et al. 2011), with highest WUE occurring during the peak of the warm wet growing season. This site also produced evidence of a drought-induced lowering of WUE. Ponton et al. (2006) similarly found a negative correlation between daily WUE and maximum daily vapor pressure deficit for both forests and grasslands. In the current study, since soil moisture was driven by runoff more than precipitation during the summer months, the largest inter-diel differences in either LUE or WUE were caused by cloudiness. Cloudy days reduced PAR, GPP and ET, but increased both LUE and WUE. The former is likely to be due to the higher use efficiency of scattered light than direct beam, and the latter due to the lower vapor pressure deficit found on those days.

Relationships between daily total GPP and (a) PAR and (b) ET for different seasonal periods. Linear models and coefficient of determination correspond with adjacent lines, which are derived from a combination of the two sub-periods
Estimate of 2016 Annual NEE at Loney Meadow
A direct estimate of the annual total net ecosystem exchange of CO2 was not possible because observations did not span the complete annual cycle. However, since most of the growing season was captured by the eddy covariance record and the meadow is covered by snow for about 5 months of the year, an approximation is possible, with some reasonable assumptions. First, we assumed ecosystem photosynthesis began around April 25th 2016, when it was observed that approximately 75% of the meadow surface was still covered by snow but that it was melting quickly and new shoots were beginning to emerge across the meadow. The second assumption was that the rates of change in daily GPP, RE and NEE trended similarly at the beginning of the emergent phase as during the latter portion that was observed. The rates of change in observed daily GPP, RE and NEE during the growing season were found to follow a linear trend, which was extrapolated backward from the beginning of observations (May 17th) to the beginning of the snow-free phase (April 25th) to estimate daily totals for each day in between. Daily values from the final senescent phase were similarly extrapolated forward from the end of the observational record to the beginning of snow cover, which was determined to be November 24th 2016, using snow depth data from a nearby climate station (SNOTEL: Robinson Cow Camp, Elevation: 1975 m MSL). The final assumption was that ecosystem photosynthesis was zero but that soil respiration continued beneath the snowpack. It has been found that respiration from organic soils through winter snowpacks tends to be small but not negligible. This is because microbial activity can be significant in organic soils in near and even below freezing conditions but that the snowpack limits oxygen supply to the soil, increasing concentrations of CO2 in the snowpack (Sommerfeld et al. 1993). Previous estimates of soil respiration beneath snowpacks include approximately 0.52 gC m−2 d−1 found both for a wet meadow (Knowles et al. 2016) and a deciduous forest floor (Hirano 2005), 1 gC m−2 d−1, for mountain grasslands in Switzerland (Wohlfahrt et al. 2008), and 0.7 to 2 gC m−2 d−1 for alpine and subalpine meadows respectively (Sommerfeld et al. 1993). Given the organic soils and large growing season productivity at Loney Meadow, here we made the assumption that Loney meadow winter respiration rates beneath a snowpack was at the higher end of this range (~2 gC m−2 d−1). Daily NEE totals were calculated for the observation period using the high quality observations of NEE and gap-filled data when observational data did not pass quality control tests. We then combined these with the extrapolated values found for the unmeasured beginning and end of the snow-free period, and the estimated values from the literature for the period with snowpack to produce an annual record. This resulted in an estimated annual carbon sink of −313 gC m−2 a−1. Hirano (2005) showed that CO2 efflux more than doubled in the approximately 2 week period following snowmelt, due to both built-up CO2 in the snow and soil air parcels that is released, and invigorated microbial respiration. If we factor a two-fold increase in CO2 efflux for 2 weeks following snowmelt, the annual sink reduces to −285 gC m−2 a−1. It is also possible that soil respiration under the snowpack followed a more modest level previously observed (~1 gC m−2 a−1), in which case the annual sink would be as high as −450 gC m−2 a−1. These annual estimates are fairly crude but suggest Loney Meadow is a significant sink of carbon on an annual basis. Reed et al. (2020) used soil carbon and biomass measurements to estimate net soil carbon fluxes from 13 montane meadows of the Sierra Nevada and found Loney Meadow to be the largest sink at 847.7 gC m−2 y−1.
Despite having a similar seasonal pattern of NEE and potentially reaching large daily magnitudes of CO2 uptake during the peak of the growing season, most grasslands observed in related climates sequester significantly less CO2 from the atmosphere annually than Loney Meadow appears to. Annual values tend to range from −50 to −160 gC m−2 a−1 during years with sufficient precipitation (e.g., Ma et al. 2007; Dong et al. 2011) to annual sources from 50 to 100 gC m−2 y−1 under drier conditions (Flanagan et al. 2002; Ma et al. 2007; Scott et al. 2010). Water availability and its timing is consistently identified as a driving environmental control on annual NEE of grasslands and several studies show this can cause a switch from a net carbon sink to source between years (Flanagan et al. 2002; Wohlfahrt et al. 2008; Scott et al. 2010; Dong et al. 2011). A synthesis study of wetland ecosystems consisting of peatlands and tundra by Lund et al. (2010) reported average annual net CO2 uptake of −103 + 103 gC m−2 a−1. The two most similar sites, with annual carbon sinks of about −200 gC m−2 a−1, were fen type wetlands with high vegetation density. Not far from Loney Meadow but near sea level, Knox et al. (2015) observed an annual NEE of −397 gC m−2 a−1 in a dense mature wetland in the Sacramento/San Joaquin Delta of California. Loney Meadow is likely to be a significantly larger sink than the global average of −156 gC m−2 a−1 based on 1459 site-years of data collected from flux towers covering terrestrial ecosystems around the planet (Baldocchi 2014).
Discussion
Implications for the Carbon Cycle in Mountain Meadows
The results from this study indicate that mountain meadows such as Loney Meadow can be large sinks for atmospheric CO2, with highly dynamic seasonal variability. This appears to be mostly due to the unique hydrology of mountain meadows which can provide moisture to the root zone of meadow ecosystems during summer despite a lack of precipitation, producing a unique seasonal signal for the eco-climate region. Furthermore, meadow soils have been found to be high in organic content to significant depths, suggesting net annual carbon uptake and longer term storage of carbon in the substrate. Snow cover in winter and water saturation in spring likely helps limits the loss of organic matter by heterotrophic respiration. There may be some loss of dissolved carbon from the meadow via the stream network, and additional losses of carbon to the atmosphere in the form of methane emissions produced during saturated soil conditions, and also emitted from grazing cattle. A fuller discussion of the likely role of cattle on the meadow carbon cycle is discussed in the following section.
The Loney Meadow ecosystem sequestered CO2 at rates similar to mature wetlands and productive forests. The net CO2 flux observed in this study was more than double that measured during an above average precipitation year in a California grassland. Yearly precipitation totals that impacted this study were close to (slightly higher) than average and followed a multi-year severe drought. Despite this, the meadow produced abundant vegetation and acted as a significant sink over the annual cycle. This was also found by Reed et al. (2020) using assessment of carbon stocks in soil and vegetation. They found Loney to be the largest annual sink of 13 SN meadows, although their estimate of more than 800 gC m−2 a−1 is 2–3 times higher than the estimate we produced using eddy covariance. Their value is also extremely high in comparison to annual estimates of a wide range of global ecosystems (e.g. Baldocchi 2014). The two methods for estimating the annual carbon flux are very different, and have different assumptions and weaknesses. For example, eddy covariance has a tendency to under-estimate the total flux, though this error is typically about 10–20% (e.g. Stoy et al. 2013). In addition, we were only able to measure over the growing season, and had to make many assumptions about winter and early spring fluxes. The soils approach, on the other hand, has coarse temporal coverage, which can describe only general seasonal changes in CO2 exchanges, but not the impacts of meteorology on day-to-day variability or diurnal variability in soil temperature and plant functioning. We find significant variability in CO2 fluxes over these timescales. On the other hand, the spatial sampling should be similarly representative of the meadow ecosystem for both techniques. This disparity in the annual carbon budget points to the need for further comparisons between carbon fluxes obtained from the two different techniques for a range of meadows. In addition, the information provided by the two techniques is highly complementary and could be combined. The eddy covariance method provides a direct measure of the CO2 exchange rate with excellent temporal resolution but cannot directly ascertain the individual components and drivers of the gas flux, which is a strength of the approach of Reed et al. (2020).
Though the number of site years of mountain meadow observations are few, the comparison between this and other studies in China, Europe and elsewhere in the Sierra Nevada suggests mountain meadow ecosystems are highly complex and show a great deal of variability compared to other ecosystems. Factors that may contribute to this variability are elevation, latitude, watershed characteristics, precipitation, temperature and meadow hydrogeomorphology. The results presented here agree with other studies that show soil moisture levels in the root zone appear to be the main environmental driver (along with temperature) that controls the larger seasonal trends as they relate to plant functioning and the resulting carbon cycling. This means that mountain meadows have great potential to help sequester atmospheric carbon, but that their capacity to provide this role is highly dependent on soil moisture levels throughout the growing season, making them susceptible to changes in both climate and land use.
Implications for Meadow Carbon Cycling of Land-Use and Climate Change
This study and previous research conducted in SN meadows have indicated that water availability shows a strong positive relationship to ecosystem productivity. Channel incision, resulting from degradation, effectively lowers the water table and reduces water available in the root zone. Any decrease in soil moisture availability in the meadow due to water table lowering will likely reduce net ecosystem uptake of CO2, and potentially cause a switch from a sink to a source of CO2. On a seasonal basis, this switch occurred when volumetric soil moisture content dropped below about 12%. Temperature and PAR are also likely to play a role in the senescent phase although, in this case, temperatures and solar radiation remained seasonally high while both soil moisture and vegetation productivity declined. If restoration of degraded meadows to raise the water is successfully implemented, there is potential to increase carbon uptake and to retain higher levels of photosynthesis later in the growing season.
Above-ground biomass consumption, animal respiration, digestive release of methane, soil compaction, and excretion deposits are all likely impacts from grazing cattle on meadow carbon cycles (Jerome et al. 2014; Roche et al. 2014). Cattle grazing in Loney Meadow during the study period was low density, yet the wet meadow vegetation clearly provided excellent forage and evidence of reduced biomass from grazing and feces deposition was widespread. Sousanna et al. (Soussana et al. 2007) found that 25–40% of cattle forage intake is returned to the soil as non-digestible carbon (e.g. feces), while a larger portion is removed from the ecosystem as live weight (LW) gained during grazing. Some of the net carbon sink we have recorded for Loney Meadow was therefore lost from the meadow as LW gain. Tofastrud et al. (2020) found the weight gain of a range of early maturing beef cows grazing at low stocking density averaged 24 kgC per growing season. If we use this value and assume all of the weight gain was obtained from Loney Meadow vegetation, the carbon loss from the meadow by cattle weight gain for the Lonwy 2016 grazing season would be 17 gC m−2, which represents a small but significant reduction of our estimated net carbon sink.
The cattle also directly impact the meadow carbon budget through autotrophic respiration of CO2 and the digestive release of CH4. Some of the respiration of CO2 from grazing cattle would have been recorded by the instruments when cows were grazing in the meadow upwind of the flux tower. We can approximate the magnitude of the annual flux independently based on the number of animals, number of days on site and using the daily estimate of cattle CO2 emissions of 2.6 kgC d−1 per livestock unit (Jerome et al. 2014). If we assume the cattle only grazed Loney, we get an upper estimate of cattle respiration of approximately 150 gC m−2 a−1, which is small compared to the estimated annual meadow respiration of over 4 kg m−2 a−1. Some portion of this would be contained in the estimate provided by the eddy covariance measurements, although this portion is impossible to estimate due to the low grazing density and lack of positional data for the cattle. From both visual observations during site visits and images captured by the wildlife camera, it is clear that cattle respiration would be recorded by the instruments, at least part of the time. We did not measure the flux of CH4, though Sousanna et al. (Soussana et al. 2007) estimated that non-lactating cattle release between 0.33 and 0.45 gCH4 kg−1 living weight per year. If we assume the larger of these values, and multiply by the total stock estimated living weight, and again assume that cattle only consumed Loney Meadow vegetation, we estimate this to be about 78 mgCH4 m−2. In meadow carbon budget terms this is very small, although it represents a more significant impact to atmospheric greenhouse gasses due to the relatively high radiative forcing of CH4. Although CH4 fluxes were not observed in this study, Reed et al. (2018) observed a diurnally-consistent soil-atmosphere CH4 flux of −0.65 nmol m−2 s−1 for Loney Meadow soils on a single day in the 2015 growing season. This equates to an uptake of nearly 1 mgCH4 m−2 d−1, which is opposite in sign but similar magnitude to the estimated total cattle emissions on a daily basis.
These estimates of the impact of cattle on the meadow carbon budget are approximations only and the impact of livestock on the carbon cycle has shown significant variability among cattle species, type of ingested forage, climate conditions, ecosystem health and management intensity (Soussana et al. 2007; Jerome et al. 2014; Roche et al. 2014). Nevertheless, these estimated direct impacts of cattle reflect a loss of carbon from the meadow, some of which was unmeasured, though the overall contribution is expected to be small compared to the observed vegetation CO2 exchanges. Further, the estimates of unmeasured components of meadow carbon fluxes suggest that our estimates of annual NEE are overestimated due to the unmeasured loss of carbon by cattle, although even a conservative accounting leaves the conclusion that the meadow is a strong net annual sink.
Depending on management intensity (e.g. stocking rate) and history, grazing can also alter plant community composition, soil characteristics, and the hydrologic regime (Soussana et al. 2007; Roche et al. 2014). Drought conditions reduce GPP, making the ecosystem more vulnerable to stress caused by grazing animals, while wet, healthy meadows exhibit greater resilience to disturbance (Roche et al. 2014). Many of these impacts are long-lasting, and meadow ecosystems have been found to be particularly vulnerable to any management impacts that cause a lowering of the water table. Considering the low stocking rate and large area available to graze, the presence of livestock during the study period likely had a small direct impact of lowering the net annual carbon sequestration. It is also likely that legacy impacts from activities such as grazing has degraded the natural hydrologic regime, creating secondary impacts on plant communities and their ability to sequester carbon. Given the relatively low plant productivity and soil carbon of degraded meadows with depleted water availability to plant roots, these indirect impacts on the carbon budget may be larger and are certainly longer lasting.
Similar to the effects of degradation, climate change has the potential to impact the net CO2 potential of SN mountain meadows by altering precipitation and seasonal hydrologic inputs. Research suggests that a warming climate will reduce the amount of precipitation that falls as snow and initiate snowmelt earlier in the season, which will contribute to a longer and drier growing season (Lowry et al. 2011). Using this study as an example, climate trends suggest that the peak growth period would shorten and start earlier and the senescence period would lengthen. Since the peak growth period accounts for nearly half of the entire net CO2 uptake measured in the 2016 growing season, a shorter peak growth period would have a significant negative impact on the overall strength of the sink on an annual basis. Similarly, a longer period of carbon release, stemming from earlier seasonal drying, will contribute to declines in annual carbon uptake. With a possible switch to a net annual loss of carbon, meadow soils would likely lose carbon to the atmosphere from stocks built up over time. Indeed, Roche et al. (2014) and Maher (2015) both found organic content in dry or degraded meadows to be significantly less than that of wet or restored meadows in the Sierra Nevada. Another potential effect of a warmer climate is increasing soil temperatures, which would likely stimulate higher respiration rates throughout the season. Furthermore, if a high water table is maintained through protection and restoration practices, it is likely that SN meadows will be more resilient to the effects of climate change and maintain their effectiveness at sequestering carbon from the atmosphere.
Conclusions
This study employed eddy covariance to investigate surface-atmosphere exchanges of CO2 in a mountain meadow in the northern Sierra Nevada from May to September 2016. Loney Meadow acted as a strong net sink of CO2 from the atmosphere over most of the growing season, averaging −7.71 gC m−2 d−1. Though clearly driven by PAR, the diurnal pattern of NEE showed a slightly stronger response (sink) in the morning than the afternoon for the same light levels. At night, ecosystem respiration produced a weak but consistent source of CO2 to the atmosphere and these rates (~0.1 < NEE < 0.3 mgC m−2 s−1) were similar throughout the measurement period.
Following snowmelt in early May, GPP increased rapidly and NEE declined so that the ecosystem became a strong sink of atmospheric CO2, peaking at −18.5 gC m−2 d−1. With daily total values ranging between about 10 and 50 gC m−2 d−1, GPP drove the variability in NEE throughout the growing season. GPP was governed principally by light at the diurnal and synoptic timescales and by soil water availability over the seasonal timescale. RE rates were much smaller and more consistent throughout the growing season than GPP, though were weakly positively correlated with temperature changes. Decline in soil moisture appeared to be the strongest control on the seasonal growth cycle and by August the ecosystem had switched from a net sink to source of CO2 peaking at 3 gC m−2 d−1. Although the complete annual cycle was not observed, approximations of the annual budget ranged from −285 to −450 gC m−2 a−1 depending on assumptions made. These values are closer to mature wetlands and forests, and represent a significantly higher carbon sink than most grasslands.
Using a rectangular hyperbola LUE model, the initial slope of the curve (α value) and the point of maximum CO2 assimilation (Amax) were high (0.86 and 246 respectively) compared to grasslands, though the relationship weakened considerably during senescence. WUE values for Loney Meadow (~ 5.5 g kg−1) were consistent and high compared with those determined equivalently from the eddy covariance record for other ecosystems. Similar to LUE, these values were closer to those found for mature wetlands and forests than grasslands. On a day-to-day basis, both LUE and WUE increased on cloudy days. This likely reflects both the higher use-efficiency of scattered light than direct beam, and the lower vapor pressure deficit experienced by the ecosystem on those days.
More long-term monitoring of the carbon exchange in SN meadows is needed to explore differences in space and time. Networks and collaborations are required to compare meadows of differing elevations, topography, level of degradation or restoration, and hydroclimate regimes. Long-term studies are required to understand inter-annual variability of carbon exchanges due to differences in snow and rainfall amounts and timing and future research should also include CH4 fluxes since they become saturated during the spring melt. Additional measurements using chamber approaches would be helpful for deciphering the role of different plant communities within and between meadows. These approaches, along with better collaboration with the soil science community, will help provide a more complete understanding of carbon dynamics in mountain meadows. This is required to inform policymakers, land managers, and stakeholders of the likely impact of both future climates and land-use management decisions.
Data Availability
All data used in this study are freely available by contacting the corresponding author.
References
Allen BH (1987) Forest and meadow ecosystems in California. Rangelands 9:125–128 http://www.jstor.org/stable/3901046
Allen-Diaz BH (1991) Water table and plant species relationships in Sierra Nevada meadows. American Midland Naturalist 126:30–43. https://doi.org/10.2307/2426147
Baccei JS, McClaran MP, Kuhn TJ et al (2020) Multi-scale drivers of soil resistance predict vulnerability of seasonally wet meadows to trampling by pack stock animals in the Sierra Nevada, USA. Ecol Process 9. https://doi.org/10.1186/s13717-020-00236-7
Baldocchi DD (2008) ‘Breathing’of the terrestrial biosphere: lessons learned from a global network of carbon dioxide flux measurement systems. Australian Journal of Botany 56:1–26. https://doi.org/10.1071/BT07151
Baldocchi DD (2014) Measuring fluxes of trace gases and energy between ecosystems and the atmosphere – the state and future of the eddy covariance method. Global Change Biology 20:3600–3609. https://doi.org/10.1111/gcb.12649
Beer C, Ciais P, Reichstein M, Baldocchi DD, Law BE, Papale D, Soussana JF, Ammann C, Buchmann N, Frank D, Gianelle D, Janssens IA, Knohl A, Köstner B, Moors E, Roupsard O, Verbeeck H, Vesala T, Williams CA, Wohlfahrt G (2009) Temporal and among-site variability of inherent water use efficiency at the ecosystem level. Global Biogeochemical Cycles 23:GB2018. https://doi.org/10.1029/2008GB003233
Blankinship JC, Hart SC (2014) Hydrological control of greenhouse gas fluxes in a sierra Nevada subalpine meadow. Arctic Antarctic Alpine Res 46:355–364. https://doi.org/10.1657/1938-4246-46.2.355
Burba G (2013) Eddy covariance method for scientific, industrial, agricultural and regulatory applications: a field book on measuring ecosystem gas exchange and areal emission rates. LI-Cor Biosciences, Lincoln, USA, p 331
Castellví F, Oliphant AJ (2017) Daytime sensible and latent heat flux estimates for a mountain meadow using in-situ slow-response measurements. Agric Forest Meteorol 236:135–144. https://doi.org/10.1016/j.agrformet.2017.01.003
Davis J, Blesius L, Slocombe M, Maher S, Vasey M, Christian P, Lynch P (2020) Unpiloted aerial system (UAS)-supported biogeomorphic analysis of restored Sierra Nevada montane meadows. Remote Sensing 12:1828. https://doi.org/10.3390/rs12111828
Dong G, Guo J, Chen J, Sun G, Gao S, Hu L, Wang Y (2011) Effects of spring drought on carbon sequestration, evapotranspiration and water use efficiency in the Songnen meadow steppe in Northeast China. Ecohydrology 4:211–224. https://doi.org/10.1002/eco.200
Dugas WA, Heuer ML, Mayeux HS (1999) Carbon dioxide fluxes over bermudagrass, native prairie, and sorghum. Agric Forest Meteorol 93:121–139. https://doi.org/10.1016/S0168-1923(98)00118-X
Dwire KA, Kauffman JB, Baham JE (2006) Plant species distribution in relation to water-table depth and soil redox potential in montane riparian meadows. Wetlands 26:131–146. https://doi.org/10.1672/0277-5212(2006)26[131:PSDIRT]2.0.CO;2
Fites-Kaufman JA, Rundel P, Stephenson N, Weixelman DA (2007) Montane and subalpine vegetation of the Sierra Nevada and Cascade ranges. Terrestrial Vegetation of California, pp 456–501
Flanagan LB, Wever LA, Carlson PJ (2002) Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland. Global Change Biology 8:599–615. https://doi.org/10.1046/j.1365-2486.2002.00491.x
Gilmanov TG, Soussana JF, Aires L, Allard V, Ammann C, Balzarolo M, Barcza Z, Bernhofer C, Campbell CL, Cernusca A, Cescatti A (2007) Partitioning European grassland net ecosystem CO2 exchange into gross primary productivity and ecosystem respiration using light response function analysis. Agric Ecosystems Environ 121:93–120. https://doi.org/10.1016/j.agee.2006.12.008
Gilmanov TG, Tieszen LL, Wylie BK, Flanagan LB, Frank AB, Haferkamp MR, Meyers TP, Morgan JA (2005) Integration of CO2 flux and remotely-sensed data for primary production and ecosystem respiration analyses in the northern Great Plains: potential for quantitative spatial extrapolation. Glob Ecol Biogeography 14:271–292. https://doi.org/10.1111/j.1466-822X.2005.00151.x
Hamlet AF, Mote PW, Clark MP, Lettenmaier DP (2005) Effects of temperature and precipitation variability on snowpack trends in the western United States. J Climatol 18:4545–4561. https://doi.org/10.1175/JCLI3538.1
Hammersmark CT, Rains MC, Mount JF (2008) Quantifying the hydrological effects of stream restoration in a montane meadow, northern California, USA. River Res Appl 24:735–753. https://doi.org/10.1002/rra.1077
Haugo RD, Halpern CB (2007) Vegetation responses to conifer encroachment in a western Cascade meadow: a chronosequence approach. Bot. 85:285–298. https://doi.org/10.1139/B07-024
Hirano T (2005) Seasonal and diurnal variations in topsoil and subsoil respiration under snowpack in a temperate deciduous forest, global Biogeochem. Cycles 19:GB2011. https://doi.org/10.1029/2004GB002259
Hsieh CI, Katul G, Chi TW (2000) An approximate analytical model for footprint estimation of scalar fluxes in thermally stratified atmospheric flows. Adv Water Resour 23:765–772. https://doi.org/10.1016/S0309-1708(99)00042-1
Hutchinson R, Weisman A, Ronning K (2020) Yuba headwaters meadow restoration monitoring report. South Yuba River citizens league. California Department of Fish & wildlife ecosystem restoration program #P1496009, p 50
Jerome E, Beckers Y, Bodson B, Heinesch C, Moureaux B, Aubinet M (2014) Impact of grazing on carbon dioxide exchanges in an intensively managed Belgian grassland. Agric Ecosyst Environ 194:7–16
Kato T, Tang Y, Gu S, Cui X, Hirota M, Du M, Li Y, Zhao X, Oikawa T (2004) Carbon dioxide exchange between the atmosphere and an alpine meadow ecosystem on the Qinghai–Tibetan plateau, China. Agric Forest Meteorol 124:121–134. https://doi.org/10.1016/j.agrformet.2003.12.008
Kattelmann, R, Embury M (1996) Riparian areas and wetlands. Sierra Nevada Ecosystem Project, Final Report to Congress (Vol. 3)
Kayranli B, Scholz M, Mustafa A, Hedmark Å (2010) Carbon storage and fluxes within freshwater wetlands: a critical review. Wetlands 30:111–124. https://doi.org/10.1007/s13157-009-0003-4
Knowles JF, Blanken PD, Williams MW (2016) Wet meadow ecosystems contribute the majority of overwinter soil respiration from snow-scoured alpine tundra. J Geophys Res Biogeosci 121:1118–1130. https://doi.org/10.1002/2015JG003081
Knox SH, Sturtevant C, Matthes JH, Koteen L, Verfaillie J, Baldocchi D (2015) Agricultural peatland restoration: effects of land-use change on greenhouse gas (CO2 and CH4) fluxes in the Sacramento-san Joaquin Delta. Glob Change Biol 21:750–765. https://doi.org/10.1007/s13157-009-0003-4
Kondolf GM, Kattelmann R, Embury M and Erman DC (1996) Status of riparian habitat. Sierra Nevada ecosystem project: final report to congress. Center for Water Wildlands Research. University of California, Davis. (Vol. 2, 1009-1030)
Lee X, Massman W and Law B (2004) Handbook of micrometeorology: a guide for surface flux measurement and analysis (Vol. 29). Springer Science & Business Media
Loheide SP, Deitchman RS, Cooper DJ, Wolf EC, Hammersmark CT, Lundquist JD (2009) A framework for understanding the hydroecology of impacted wet meadowsin the Sierra Nevada and Cascade ranges, California, USA. Hydrogeol J 17:229–246. https://doi.org/10.1007/s10040-008-0380-4
Loheide SP, Gorelick SM (2007) Riparian hydroecology: a coupled model of the observed interactions between groundwater flow and meadow vegetation patterning. Water Resour Res 43. https://doi.org/10.1029/2006WR005233
Lowry CS, Loheide SP, Moore CE, Lundquist JD (2011) Groundwater controls on vegetation composition and patterning in mountain meadows. Water Resour Res 47. https://doi.org/10.1029/2010WR010086
Lund M, Lafleur PM, Roulet NT, Lindroth A, Christensen TR, Aurela M, Chojnicki BH, Flanagan LB, Humphreys ER, Laurila T, Oechel WC (2010) Variability in exchange of CO2 across 12 northern peatland and tundra sites. Glob Change Biol 16:2436–2448. https://doi.org/10.1111/j.1365-2486.2009.02104.x
Ma S, Baldocchi DD, Xu L, Hehn T (2007) Inter-annual variability in carbon dioxide exchange of an oak/grass savanna and open grassland in California. Agric Forest Meteorol 147:157–171. https://doi.org/10.1016/j.agrformet.2007.07.008
Maher SC (2015) Bio-micrometeorology of a Sierra Nevada Montane Meadow. Masters Thesis, San Francisco State University
Massman WJ, Lee X (2002) Eddy covariance flux corrections and uncertainties in long-term studies of carbon and energy exchanges. Agric Forest Meteorol 113(1–4):121–144. https://doi.org/10.1016/S0168-1923(02)00105-3
Norton JB, Jungst LJ, Norton U, Olsen HR, Tate KW, Horwath WR (2011) Soil carbon and nitrogen storage in upper montane riparian meadows. Ecosystems 14:1217–1231. https://doi.org/10.1007/s10021-011-9477-z
Oliphant AJ (2012) Terrestrial ecosystem-atmosphere exchange of CO2, water and energy from FLUXNET; review and meta-analysis of a global in-situ observatory. Geography Compass 6:689–705. https://doi.org/10.1111/gec3.12009
Oliphant AJ, Dragoni DD, Deng B, Grimmond CSB, Schmid HP, Scott SL (2011) The role of sky conditions on gross primary production in a mixed deciduous forest. Agric Forest Meteorol 151:781–791. https://doi.org/10.1016/j.agrformet.2011.01.005
Papale D, Reichstein M, Aubinet M, Canfora E, Bernhofer C, Kutsch W, Longdoz B, Rambal S, Valentini R, Vesala T, Yakir D (2006) Towards a standardized processing of net ecosystem exchange measured with eddy covariance technique: algorithms and uncertainty estimation. Biogeosciences 3:571–583 https://hal.archives-ouvertes.fr/hal-00330317
Plumas Corporation (2020) Mountain meadows restoration project at Greenville Creek and upper Goodrich and effects on greenhouse gases, wetlands restoration for greenhouse gas reduction, final report CDFW Grant agreement P1496002 01. Center for Watershed Sciences, University of California, Davis
Ponton S, Flanagan LB, Alstad KP, Johnson BG, Morgenstern K, Kljun N, Black TA, Barr AG (2006) Comparison of ecosystem water-use efficiency among Douglas-fir forest, aspen forest and grassland using eddy covariance and carbon isotope techniques. Glob Change Biol 12:294–310. https://doi.org/10.1111/j.1365-2486.2005.01103.x
Pope KL, Montoya DS, Brownlee JN, Dierks J, Lisle TE (2015) Habitat conditions of montane meadows associated with restored and unrestored stream channels of California. Ecol Restoration 33(1):61–73. https://doi.org/10.3368/er.33.1.61
Purdy SE, Moyle PB (2006) Mountain meadows of the Sierra Nevada. Center for Watershed Sciences, University of California, Davis
Ratliff RD (1982) A meadow site classification for the Sierra Nevada, California. USDA Forest Service General Technical Report PSW-60:16. https://doi.org/10.2737/PSW-GTR-60
Ratliff RD (1985) Meadows in the Sierra Nevada of California: state of knowledge. USDA Forest Service general technical report PSW-GTR-84, 52. https://doi.org/10.2737/PSW-GTR-84
Reed CC, Merrill AG, Drew WM, Christman B, Hutchinson RA, Keszey L, Odell M, Swanson S, Verburg PSJ, Wilcox J, Hart SC, Sullivan BW (2020) Montane meadows: a soil carbon sink or source? Ecosystems. https://doi.org/10.1007/s10021-020-00572-x
Reed CC, Winters JM, Hart SC, Hutchinson R, Chandler M, Venicz G, Sullivan BW (2018) Building flux capacity: citizen scientists increase resolution of soil greenhouse gas fluxes. PLoS One 13:e0198997. https://doi.org/10.1371/journal.pone.0198997
Reichstein M, Falge E, Baldocchi D, Papale D, Aubinet M, Berbigier P, Bernhofer C, Buchmann N, Gilmanov T, Granier A, Grünwald T (2005) On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm. Glob Change Biol 11:1424–1439. https://doi.org/10.1111/j.1365-2486.2005.001002.x
Roche LM, O’Green AT, Latimer AM, Eastburn DJ (2014) Montane meadow hydropedology, plant community, and herbivore dynamics. Ecosphere 5:1–16. https://doi.org/10.1890/ES14-00173.1
Scott RL, Hamerlynck EP, Jenerette GD, Moran MS, Barron-Gafford GA (2010) Carbon dioxide exchange in a semidesert grassland through drought-induced vegetation change. J Geophys Res 115(G3). https://doi.org/10.1029/2010JG001348
Sommerfeld R, Mosier A, Musselman R (1993) CO2, CH4 and N2O flux through a Wyoming snowpack and implications for global budgets. Nature 361:140–142. https://doi.org/10.1038/361140a0
Soussana JF, Allard V, Pilegaard K, Ambus P, Amman C, Campbell C, Ceschia E, Clifton-Brown J, Czóbel SZ, Domingues R, Flechard C (2007) Full accounting of the greenhouse gas (CO2, N2O, CH4) budget of nine European grassland sites. Agric Ecosyst Environ 121:121–134. https://doi.org/10.1016/j.agee.2006.12.022
Stewart IT, Cayan DR, Dettinger MD (2005) Changes toward earlier streamflow timing across western North America. J Climatol 18:1136–1155. https://doi.org/10.1175/JCLI3321.1
Stoy PC, Mauder M, Foken T, Marcolla B, Boegh E, Ibrom A, Arain MA, Arneth A, Aurela M, Bernhofer C, Cescatti A (2013) A data-driven analysis of energy balance closure across FLUXNET research sites: the role of landscape scale heterogeneity. Agric Forest Meteorol 171:137–152. https://doi.org/10.1016/j.agrformet.2012.11.004
Tofastrud M, Hessle A, Rekdal Y, Zimmermann B (2020) Weight gain of free-ranging beef cattle grazing in the boreal forest of South-Eastern Norway. Livestock Sci 233:103955. https://doi.org/10.1016/j.livsci.2020.103955
Viers JH, Purdy SE, Peek RA, Fryjoff-Hung A, Santos NR, Katz JV, Emmons JD, Dolan DV, Yarnell SM (2013) Montane Meadows In The Sierra Nevada: Changing Hydroclimatic Conditions and Concepts for Vulnerability Assessment. Centerfor Watershed Sciences Technical Report (CWS-2013-01), University of California, Davis, p 63
Viers JH, Rheinheimer DE (2011) Freshwater conservation options for a changing climate in California’s Sierra Nevada. Marine Freshwater Res 62:266–278. https://doi.org/10.1071/MF09286
Weixelman DA, Hill B, Cooper DJ, Berlow EL, Viers JH, Purdy SE, Merrill AG, Gross SE (2011) A field key to meadow hydrogeomorphic types for the Sierra Nevada and southern Cascade ranges in California. US Forest Service, Pacific Southwest Region, Vallejo, California, USA
Wilson K, Goldstein A, Falge E, Aubinet M, Baldocchi D, Berbigier P, Bernhofer C, Ceulemans R, Dolman H, Field C, Grelle A (2002) Energy balance closure at FLUXNET sites. Agric Forest Meteorol 113:223–243. https://doi.org/10.1016/S0168-1923(02)00109-0
Wohlfahrt G, Hammerle A, Haslwanter A, Bahn M, Tappeiner U, Cernusca A (2008) Seasonal and inter-annual variability of the net ecosystem CO2 exchange of a temperate mountain grassland: effects of weather and management. J Geophys Res 113(D8). https://doi.org/10.1029/2007JD009286
WRCC (Western Regional Climate Center) (2020) Data portal Accessed 20 Nov 2020. Retrieved from https://wrcc.dri.edu/cgi-bin/cliMAIN.pl?ca1018
Xu L, Baldocchi DD (2004) Seasonal variation in carbon dioxide exchange over a Mediterranean annual grassland in California. Agric Forest Meteorol 123(1–2):79–96. https://doi.org/10.1016/j.agrformet.2003.10.004
Zegelin SJ, White I, Russell G (1992) In: Topp GC, Reynolds WD, Green RE (eds) A critique of the time domain Reflectometry technique for determining Field soil-water content. In advances in measurement of soil physical properties: bringing theory into practice. https://doi.org/10.2136/sssaspecpub30.c10
Acknowledgements
The authors would like to acknowledge the US Forest Service for access to the meadow and South Yuba River Citizens League for field support and coordination. In particular we are grateful to Rachel Hutchinson for providing useful additional data and advice on this manuscript. The authors would also like to thank Quentin Clark for invaluable field support, the SFSU Department of Geography & Environment for providing transportation to the field site and some field sampling equipment, and the SFSU College of Science & Engineering for funds to acquire the multispectral camera and drone platform. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Code Availability
Not applicable.
Funding
External funding was not available for this study. Transportation for fieldwork was provided by San Francisco State University, Dept. of Geography & Environment.
Author information
Authors and Affiliations
Contributions
Study conception, design and most fieldwork was conducted by DB and AO. JD conducted field collection and analysis of drone imagery. Data analysis was principally conducted by DB and AO and the initial manuscript was drafted by DB. All authors worked on all drafts of the manuscript thereafter, with particular focus on meadow hydrogeomorphology by JD and biometeorological aspects by DB and AO. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics Approval
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Conflicts of Interest/Competing Interests
The authors have no conflicts of interest or competing interests related to material in this study.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Wetlands and Global change
Rights and permissions
About this article

Cite this article
Blackburn, D.A., Oliphant, A.J. & Davis, J.D. Carbon and Water Exchanges in a Mountain Meadow Ecosystem, Sierra Nevada, California. Wetlands 41, 39 (2021). https://doi.org/10.1007/s13157-021-01437-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13157-021-01437-2