
Abstract
Long-term marsh sustainability is threatened by environmental changes, especially accelerated rates of sea-level rise (SLR) and changing fluvial sediment supplies. Although direct observations of long-term marsh responses to these changes are challenging, this study examines shorter-term responses via bimonthly sedimentation measurements over 5 years in Dyke Marsh Preserve (DMP), a tidal freshwater marsh in the Potomac River (Chesapeake Bay tributary, USA). Observed patterns are evaluated with physical drivers (winds, river discharge, water-level changes, suspended-sediment concentrations (SSC)) that influence spatiotemporal variability within the marsh. Linear regression models reveal that water-level changes exert more influence on this variability than SSC in a system with plentiful sediment supply. Sedimentation rates increase linearly with inundation time, but a quadratic regression model explains more of the variability, especially for the inorganic fraction of sediment. This result indicates that fluvial sediment supply is key for marsh accretion and suggests an optimal response of sedimentation to increased inundation that non-linearly affects marsh vulnerability to future SLR. Ultimately, the SSC in adjacent waters is not available to nourish the marsh unless it is transported and retained on the platform. Self-sustaining feedbacks among inundation, sedimentation, elevation, and plants likely aid in maintaining equilibrium elevations over time and thus also marsh sustainability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Tidal marshes are among the world’s most valuable ecosystems (Feagin et al. 2010; Luisetti et al. 2013), enhancing coastal resilience to storms (Day et al. 2007; Barbier et al. 2013), filtering supply of terrestrial sediments and nutrients to adjacent waters (Bruland 2008; Loomis and Craft 2010; Ouyang and Lee 2014; Oczkowski et al. 2016), and providing habitat for rare and threatened species (Beck et al. 2003). Yet, more than half of US marshes have been altered or destroyed in the last century (Craft et al. 2009; Pendleton et al. 2012), and many existing marshes face an uncertain future due to accelerated rates of sea-level rise (SLR), changes in fluvial sediment supplies (Nicholls et al. 2011; Kirwan et al. 2011; Rahmstorf et al. 2012; Weston 2014), and wind-wave erosion (Marani et al. 2011; Fagherazzi et al. 2013; Leonardi et al. 2016).
Complex interactions among SLR, fluvial sediment supply, and the vegetation community generally control marsh elevation (Allen 2000; Friedrichs and Perry 2001; Torres et al. 2006), and most tidal marshes have kept pace with post-glacial SLR (Redfield 1972; Morris et al. 2002; Day et al. 2008; Engelhart et al. 2009). On the marsh platform, accretion (defined here as accumulation of inorganic and organic material (Lynch et al. 2015) and used interchangeably with sedimentation) occurs primarily through sediment settling at slack water (French and Stoddart 1992; Christiansen et al. 2000), particle capture by vegetation (Stumpf 1983; Leonard and Luther 1995), biogenic production, and direct organic-matter deposition from root growth and litter deposition (Nyman et al. 2006; Neubauer 2008). While organic sedimentation can dominate in slowly accreting marshes, limits to plant productivity, and thus organic sedimentation, allow mineral sediments to dominate in rapidly accreting marshes (Chmura and Hung 2004; Mudd et al. 2009). At rapid rates of SLR and/or when starved of sediments, vertical accretion of the marsh surface can fail to keep pace with SLR, resulting in marsh submergence (Reed 1995; Kirwan et al. 2010). Marshes in many regions are threatened by both accelerated SLR and declining fluvial sediment supply; e.g., in Chesapeake Bay, where local relative SLR is much greater than the global average (~3–5 mm/y; Boon et al. 2010; Sallenger et al. 2012) and many tributaries have declining sediment supplies in recent years (Weston 2014).
Most insights on marsh accretion have been gained from large-scale studies of salt marshes that often assume vegetation structure, mineral sedimentation, and associated processes are functions of elevation, so lower elevation sites that are flooded longer and more often (Cahoon and Reed 1995; Leonard and Luther 1995; Leonard 1997) have the highest plant production and mineral sedimentation rates (Morris et al. 2002; D’Alpaos et al. 2007; Day et al. 2008; Kirwan and Megonigal 2013). Moreover, sediment supply to the marsh surface is assumed to be limited by the SSC of adjacent waters (Kirwan et al. 2010). Although recent work has focused on evaluating assumed responses of both plant production (D’Alpaos et al. 2007; Marani et al. 2013; Belliard et al. 2015) and mineral sedimentation (Neubauer et al. 2002; Lagomasino et al. 2013; Palinkas and Engelhardt 2016) to inundation, much less attention has centered on understanding mechanisms of sediment supply to marshes. This study focuses on wind-driven subtidal (longer than a tidal cycle) changes in sea level and SSC, especially since SSC can vary over an order of magnitude and at multiple timescales in estuaries and coastal waters (Schoellhamer 2002; Moskalski and Torres 2012). The few observational studies examining the relative influence of SSC and inundation on marsh sedimentation focus on salt marshes and offer contrasting views. For example, Murphy and Voulgaris (2006) conclude that inundation is more important than SSC for a salt marsh in South Carolina, but Moskalski and Sommerfield (2012) find that SSC is more important for a salt marsh in Delaware. These differences highlight the considerable spatiotemporal variability in marsh sedimentation and its internal and external drivers, especially along a salinity gradient (Butzeck et al. 2015).
Tidal freshwater marshes (TFM) and salt marshes are both located within the tidal frame and subjected to the same physical drivers of accretion. However, the strength of drivers differs between them. TFM are located in upstream reaches of rivers and estuaries, where inputs of sediments and nutrients may be enhanced from anthropogenic activities and/or the presence of an estuarine turbidity maximum (region of salinity-induced flocculation and settling of fine muds in the upper reaches of an estuary; e.g. Ganju et al. 2013). TFM are more likely to experience smaller tidal ranges, increasing the importance of subtidal sea-level changes, and more localized fluvial sediment supply (Coulter et al. 2004; Brakebill et al. 2010).
This study focuses on a TFM (Dyke Marsh Preserve; DMP) in the Potomac River estuary (Chesapeake Bay watershed, USA) to evaluate how bimonthly to interannual wind-driven variations in SSC and water level are related to marsh sedimentation. (Note that the preceding discussion referred to “sea level” for consistency with the literature; “water level” is used hereafter to reflect specific conditions at DMP.) If all else were equal, higher SSC should result in higher sedimentation rates. However, we hypothesize that variations in riverine SSC are not directly related to marsh sedimentation in systems with plentiful sediment supply, like the Potomac. Instead, wind-driven changes in water levels, and correspondingly inundation, explain more of the variability in sedimentation by influencing sediment supply to the marsh. These relationships between sedimentation patterns in space (banks, interior) and time (bimonthly, seasonal, interannual) and variations in SSC and water level lend valuable insights into the response of TFM to anticipated future changes in longer-term phenomena, like SLR. The ultimate goal of this study is to better understand mechanisms that translate regional SLR and SSC into localized sedimentation rates that determine marsh elevations and potential vulnerability to future environmental changes.
Materials and Methods
Physical Setting
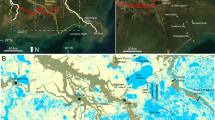
Dyke Marsh Preserve (DMP) is a TFM located near Alexandria, VA (USA), bounded by the Potomac River (east) and the George Washington Memorial Parkway (west) (Fig. 1). DMP is underlain by ~15-m thick alluvium deposits of sand and gravel interlayered with silt and clay (NPS 1977). Dredging these deposits reduced the marsh area from 0.75 km2 in 1937 to 0.34 km2 in 1976, when the National Park Service (NPS) assumed direct management (Litwin et al. 2011), and removed a promontory at the entrance to Hog Island Gut (HIG; DMP’s largest tidal channel) that had protected the shoreline from wave erosion (Litwin et al. 2013). Consequently, wave-induced lateral erosion currently dominates marsh loss, at rates comparable to regional marshes (Litwin et al. 2013). Accretion and vertical erosion on the marsh platform vary widely, likely dependent on elevation and geomorphic position, with accretion in many areas throughout the marsh (Palinkas et al. 2013; Cadol et al. 2014). Elevations range from below the datum (NAVD88) to slightly >0.6 m, with higher elevations on a remnant dike at the northeastern border of the central marsh, lower areas in the marsh interior, and gradually rising elevations towards western upland areas (Palinkas et al. 2013). Sediments generally are finer and less organic on the banks of the Potomac and HIG than in the marsh interior. Over decadal time scales, physical processes (fluvial discharge, tides) drive sedimentation on the banks, while complex interactions with the vegetation community are more active in the interior (Palinkas and Engelhardt 2016). Long-term sedimentation rates can support elevation gain in the face of relatively rapid relative SLR, though it is unclear how these rates will respond to future environmental changes (Palinkas and Engelhardt 2016).

Map of study sites in Dyke Marsh Preserve
DMP lies entirely within the floodplain of the Potomac and experiences salinities <0.5. River discharge near DMP averages 323 m3/s (Carter et al. 1994) and is typically highest in the spring (~560 m3/s; March/April, the “spring freshet”) from mountain snowmelt and groundwater saturation, and lowest in the summer (~100 m3/s) from evapotranspiration and groundwater depletion (Fisher et al. 1998). The spring freshet generally precedes establishment of most marsh vegetation. Tides near DMP are mixed semidiurnal, with a mean range of 0.5–0.9 m (Carter et al. 1994); currents during ebb discharges in HIG average 4.8 m3/s (Harper and Heliotis 1992). However, in Chesapeake Bay and its tributaries, subtidal sea-level changes from wind-driven water level setup on the Atlantic continental shelf (Chuang and Boicourt 1989) can be much larger than tides (up to ~1 m), and strong easterly winds coupled with an incoming tide can flood the entire marsh for several hours. Non-tidal currents in the Potomac respond to both local winds and non-local effects propagating from the Bay (Elliott 1978).
The growing season at DMP is relatively long (208 days; Johnston 2000) due to warm, humid summers and mild winters. The TFM at DMP comprises ~35% of the area and supports vegetation typical of Atlantic TFM (Simpson et al. 1983; Leck and Simpson 1995). Dominant species at DMP include Acorus calamus (L.), Impatiens capensis Meerb., Nuphar luteum (L.) Sm., Peltandra virginica (L.) Schott, Schoenoplectus fluviatilis (Torr.) M.T. Strong, and Typha angustifolia L. (Palinkas et al. 2013).
Field Methods
Sedimentation was studied at the same 27 sites as in Palinkas et al. (2013) and Palinkas and Engelhardt (2016), using the same rough geomorphic classification (banks of the Potomac or HIG versus the marsh interior). These sites were co-located with existing Surface Elevation Tables (SETs; n = 9) and plots established by previous studies (n = 9 from Hopfensperger et al. 2009; n = 3 from Litwin et al. 2011), as well as new sites to broaden representation across the vegetated elevation gradient (n = 6). Field activities generally occurred at low tide when the marsh was exposed. At each site, a ceramic tile (Pasternack and Brush 1998) was deployed in April 2010 and revisited bimonthly until October 2014; sediment deposited on each tile was placed into a pre-weighed glass jar. In spring (April/May) and late summer (August/September) of each year, push cores (20 cm long, 5 cm diameter) were collected to assess seasonal sediment character and rates, using 7Be (see below). The upper ~1–2 cm of cores collected in spring and summer represented “winter” and “growing season” conditions (preceding ~4–5 half-lives of 7Be (~200–250 days), the assumed limit of detectability).
Water-level data were collected with a vented pressure-sensor gauge, which eliminates the need to compensate for barometric-pressure changes. The gauge was installed on a pier at Belle Haven Marina (BHM) in May 2010. Data were logged at 15-min intervals, and gauge elevation was surveyed with RTK-GPS. Because the gauge was installed just after core collection began, and the 7Be calculations require water-level data preceding core collection, water levels at the National Oceanic and Atmospheric Administration (NOAA) tide gauge at Washington, DC, 12 km upstream of DMP, were used for 2010 (http://tidesandcurrents.noaa.gov/) correcting for a 4-h lag time due to position in the river. BHM water levels were used for 2011–2014.
Site elevations were surveyed annually from 2011 to 2014 using RTK-GPS, with an accuracy of ~1 cm horizontal and ~2 cm vertical. All surveys included a USGS vertical control benchmark located at BHM for additional validation of base-station elevations. All reported elevations use NAVD88 as the datum.
Flooding regime was characterized from tide-gauge and elevation data by calculating inundation duration (percentage of time a site was inundated during each deployment), frequency (percentage of high tides that flood a site), and average depth. Elevations were not available in 2010 and assumed to equal 2011 elevations, which were used for Deployments 1–11; 2012, 2013, and 2014 elevations were used for Deployments 12–18, 19–23, and 24–25, respectively.
Laboratory Analyses
Immediately after collection, glass jars and push cores were returned to the laboratory, where cores were sectioned into 1-cm increments; jar and core sediments were then dried at 60 °C until constant weights were reached. Dry sediment was analyzed for organic content via combustion in a muffle furnace at 450 °C for 4 h (Erftemeijer and Koch 2001).
Push-core samples were also analyzed for 7Be (half-life 53.3 d), using gamma spectroscopy of the 477.7 keV photopeak. 7Be is produced by atmospheric cosmic-ray spallation and attaches to organic and inorganic sediments during wet and dry deposition (Olsen et al. 1986; Dibb and Rice 1989). Because the entire marsh surface is exposed at least some of the time, marsh sediments are naturally labeled with 7Be, as are sediments supplied by external sources like the adjacent watershed and/or the Potomac. This atmospheric contribution must be subtracted from depth-integrated sediment inventories prior to calculating sediment deposition or erosion rates (inventories above and below atmospherically supported values, respectively; Neubauer et al. 2002; Palinkas et al. 2013) and was determined following Palinkas et al. (2013). Observed daily rainfall totals at nearby National Airport (http://www.nws.noaa.gov) were binned into biweekly intervals to apply Kim et al. (2000) relationship with 7Be flux in Chesapeake Bay. The atmospheric inventory at each site was then adjusted for interception by water when the marsh is flooded and by vegetation. For the former, site-specific elevations and the BHM tide-gauge record were used to calculate the exposure time of each site for 77 d (mean lifetime of 7Be) prior to core collection, assuming no 7Be reaches the surface when it is flooded. Vegetative interception of 7Be in TFM has not been studied but can be significant in other systems (Casey et al. 1986; Kaste et al. 2011). For this study, vegetation interception was assumed to equal the observed total vegetation cover at each site in summer and spring to account for potential interception by year-round litter (Elmore et al. 2016) at all sites except those dominated by Nuphar lutea and Peltandra virginica. No vegetative interception was assumed for these sites in spring, because leaf tissues of these species decompose rapidly, leaving the marsh surface bare in winter.
Sediment inventories of 7Be were calculated from the specific activity of each depth horizon with detectable 7Be. Specific activities were determined from ground, dry sediment placed into identical jars filled to the same height to ensure consistent counting geometry. A calibrated Canberra germanium detector (Larsen and Cutshall 1981) then counted gamma emissions for 24 h. Measured activities (dpm/g; disintegrations per minute per gram) were decay-corrected to the time of collection, and sedimentation rates were calculated following Palinkas et al. (2013). These rates are likely upper estimates of the actual sedimentation rates. Lower estimates are sedimentation rates calculated without vegetation interception. For simplicity, averages of upper and lower estimates for each site at each sampling time are used herein. Note positive and negative sedimentation rates reflect sediment deposition and erosion, respectively. Thus, while ceramic tiles record sediment deposition, 7Be integrates sediment deposition and erosion and is more reflective of marsh elevation change (direct comparisons to observed elevation change are possible by multiplying sedimentation rates and bulk densities).
Ancillary Data
Several variables in statistical models were obtained directly from publicly available sources between 1 January 2010 and 31 December 2014. Daily average wind speed and direction at National Airport were obtained from NOAA (http://www.nws.noaa.gov). Daily average Potomac River discharge at Washington, DC, was acquired from the USGS (http://waterdata.usgs.gov/); data from the full record preceding the study (1930–2009) were used to calculate the flood threshold, defined as the 99th percentile of values (2378.6 m3/s) following Pasarić and Orlić (2001). Corresponding suspended-sediment concentrations (SSC) at Washington, DC, have been measured since 1977 (http://cbrim.er.usgs.gov). Generally, SSC data were collected bimonthly in the 1980s and 1990s and monthly in the 2000s, with some gaps in the record and more frequent samples during storms. Sediment rating curves that quantify the relationship between river discharge and SSC were developed with these data by Palinkas and Engelhardt (2016); the most recent curve (2008–2012) was used to calculate daily SSC values over the study period.
Calculations and Statistical Analyses
Daily SSC data were averaged over each deployment (tiles) or season (cores) and combined with corresponding average sedimentation rates to calculate theoretical particle settling velocities, following Temmerman et al. (2003a). This approach assumes settling only when the marsh is flooded (product of deployment/season length and average inundation duration) and calculates inorganic sedimentation rates by multiplying settling velocity and depth-averaged SSC above the marsh surface, then dividing by the dry bulk density. Potomac SSC data were used to represent sediment supply to the marsh; settling velocities were calculated for periods only with positive sedimentation rates. Dry bulk density was measured only for cores; seasonal averages were consistently ~0.2 g/cm3, which was used in all calculations. Expected sedimentation rates also were calculated, assuming a particle settling velocity of 0.01 cm/s (Temmerman et al. 2003a).
Because tile sediments were collected more frequently than cores, tile data were averaged between the dates of core collection when comparing results. Pearson Product Moment was used to identify correlations among physical drivers (wind speed and direction, SSC, water level). Linear regression models were used to evaluate the relationship of these data to average tile sedimentation rates. Multiple linear regression and polynomial regression models were used to explore relationships among multiple explanatory variables. Assumptions of linear regressions were tested using the gvmla function in R, transforming data when necessary. Differences among deployments were assessed using paired t-tests. For all tests, p < 0.05 was the preferred level for statistical significance; however, p ≤ 0.1 was used to identify trends that may be physically meaningful if not statistically robust. R statistical software was used for all analyses. All averages are reported as the mean ± 1 standard deviation.
Results
Physical Drivers and Elevation
Daily average river discharge ranged 12–5434 m3/s over the study period (Fig. 2a). Average annual river discharge was highest in 2011 (472 ± 633 m3/s) and lowest in 2012 (253 ± 308 m3/s), averaging 347 ± 80 m3/s. Seasonally averaged river discharge was highest in the late winter and early growing season and lowest during the growing season (Table 1). Floods occurred on 18 days of the study, mostly during spring freshets except spring 2012 and 2013. Other floods followed large snow events (26–27 Jan 2010 and 1 Feb 2013) and an intense storm event (31 Oct 2012; Hurricane Sandy) but not after Tropical Storm Lee (peak discharge 2081 m3/s; 9 Sept 2011), the other significant storm during the study period (Palinkas et al. 2014).

Time series of physical drivers from 1 January 2010 to 31 December 2014: a river discharge, b water level, c average wind speed, and d suspended-sediment concentrations. Dashed line in (a) represents the threshold for flood conditions (2378 m3/s)
Water levels ranged from −1.2 to +1.6 m (Fig. 2b), driven by semi-diurnal tides, river discharge, and winds. Seasonally averaged water levels were lowest in winter, with moderate river discharge and more westerly winds, and highest in the early and late growing season, with higher river discharge and more southerly winds. Daily average wind speeds ranged 0.3–6.7 m/s (Fig. 2c), with larger wind events more prevalent in winter; however, seasonal averages were similar. SSC (Fig. 2d) ranged 0.9–317 mg/L, averaging 22.1 ± 28.8 mg/L and following temporal patterns of river discharge.
At bimonthly scales coincident with tile measurements, water level was negatively related to both wind speed (p = 0.04, R2 = 0.13) and direction (p < 0.001, R2 = 0.56). DMP is oriented roughly north-south, bounded by land and the Potomac on the west and east, respectively. Thus, more westerly winds tended to transport water away from the marsh, whereas more southerly winds tended to transport water toward the marsh. Wind speed was positively correlated with both wind direction (p = 0.03, R2 = 0.16) and SSC (p = 0.006 R2 = 0.25), with storms (higher wind speeds) coming from a more westerly direction and resulting in higher SSC, likely from runoff and/or resuspension. At seasonal scales coincident with core measurements, wind speed was correlated with SSC (p = 0.02, R2 = 0.72), and wind direction was correlated with water level (p = 0.005, R2 = 0.87).
Elevation across the marsh was relatively stable for 2011–2013, averaging 0.42–0.43 m, then decreased significantly (p = 0.07) in 2014 to an average of 0.38 m. In all years, average elevation was similar on the banks and in the interior (p > 0.10). Average inundation time of tiles ranged from 12.4% (Dec 2010-Feb 2011) to 39.7% (Apr-June 2014). The increase in inundation time after Apr 2014 reflects the widespread elevation decrease between 2013 and 2014.
Sediment Characteristics and Deposition Rates
The organic content of sediments from tiles and in the uppermost 1 cm of cores showed much variability at single points of space and time (e.g. ranging 15.2–84.2% for cores), but average values were similar throughout the study. Differences among sampling times and geomorphic units were not statistically significant with either method, but some general trends emerged. For tiles, average organic content was highest in Aug-Oct 2011 and lowest in Apr-Jun 2013 (Fig. 3a). Higher values generally occurred in the summer and at sites on the banks (Fig. 3b). For cores, average organic content was highest in the 2012 growing season and lowest in 2010–2011 winter; higher values typically occurred on the banks and during winter. Differences between bank and interior organic content likely reflect differences in the plant community and its year-round presence on banks versus more seasonal presence in the interior. Differences between tile and core measurements were significant (p < 0.001 for all sites, banks only, interior only), with higher tile organic content, likely from preferential trapping of plant litter and leafy debris by tiles; core sediments presumably mix these materials with older and less organic material.

a Average organic content of tile and core (uppermost cm) sediments (gray diamonds and black boxes, respectively). Note that observations were more frequent for tiles (bimonthly) than cores (seasonal). b Boxplots of core (left) and tile (right) data for sites on the banks (dark gray) and the interior (light gray); error bars represent 1 standard deviation
Average bulk (organic and mineral) tile sedimentation rates were lowest in Dec 2010-Feb 2011 and highest in Aug-Oct 2011 (Fig. 4a). These rates declined during the study, especially from September 2011 to October 2014, for all sites (p < 0.001; R2 = 0.58), on the banks (p = 0.001, R2 = 0.48) and in the interior (p < 0.001, R2 = 0.50). Spatially, bank sedimentation rates were higher than those in the interior, although differences were not statistically significant. Bulk core sedimentation rates ranged from −4.14 g/cm2/y (net erosion) to 3.67 g/cm2/y (net deposition), with higher average rates usually in the interior (Fig. 4b). On the banks, average rates were highest in the 2011 growing season, immediately followed by net erosion in winter 2011–2012 until the 2013 growing season. In the interior, average rates decreased from the maximum in winter 2009–2010 until winter 2010–2011, then were seasonally variable (higher and lower during the growing season and winter, respectively), with minimum values in winter 2013–2014. Average sedimentation rates were higher on tiles than in cores (Fig. 4c). These differences were not surprising, since tiles only capture sediment deposition, whereas cores integrate deposition and erosion. Sedimentation rates determined by these methods were statistically different in paired t-tests including all sites (p = 0.001), as well as separate tests for sites on the banks (p = 0.04) and in the interior (p = 0.01).

a Average sedimentation rates measured from tiles and cores (gray diamonds and black boxes, respectively). b Average sedimentation rates from cores for sites on the banks (black triangles) and in the interior (gray circles). c Boxplots of core (left) and tile (right) data for sites on the banks (dark gray) and the interior (light gray); error bars represent 1 standard deviation
For tiles, organic content and sedimentation rates were negatively correlated for all sites (p = 0.03, R2 = 0.15) and for sites in the interior (p = 0.01, R2 = 0.21). For cores, lower sedimentation rates generally corresponded with lower organic content, though the relationship was statistically significant only in winter 2011 (p = 0.08).
Relationships among Physical Drivers and Sedimentation Rates
Tile sedimentation rates were positively correlated with water levels across the marsh, on the banks, and in the interior (Table 2); the correlation was more robust when organic content was included for interior sites (p = 0.008, R2 = 0.30). Water levels were positively correlated with core sedimentation rates across the marsh and in the interior but not on the banks. Neither tile nor core sedimentation rates were related to wind speed or direction, or SSC.
Calculated average settling velocities for tile deployments ranged 0.002–0.07 cm/s, averaging 0.02 ± 0.02 cm/s. Expected inorganic tile sedimentation rates (assumed average settling velocity 0.01 cm/s), ranged 0.02–0.51 g/cm2/y, averaging 0.14 ± 0.12 g/cm2/y. The corresponding observed rates ranged 0.03–0.26 g/cm2/y, averaging 0.14 ± 0.07 g/cm2/y. Expected and observed rates were statistically similar (p = 0.74) and linearly correlated (p = 0.01, R2 = 0.23). For cores, average settling velocities for seasons with net deposition ranged 0.0004–0.01 cm/s, averaging 0.007 ± 0.004 cm/s. Expected inorganic core sedimentation rates ranged 0.09–0.22 g/cm2/y, averaging 0.15 ± 0.05 g/cm2/y. Corresponding observed rates ranged −0.17-0.21 g/cm2/y, averaging 0.04 ± 0.13 g/cm2/y. Expected and observed core rates were statistically distinct (p = 0.03) and not correlated (p = 0.23).
Tile and core sedimentation rates were averaged between the annual elevation surveys for comparison with observed elevation changes. There are three periods (year 1 = February 2011–2012; year 2 = February 2012–2013; year 3 = February 2013–2014) with coincident elevation and sedimentation measurements, but only data from years 1 and 2 were considered since nearly every site lost elevation in year 3 (Table 2). Observed elevation changes were not correlated with average tile sedimentation rates or organic content but were correlated with average core sedimentation rates (p = 0.01, R2 = 0.16). Observed elevation changes were smaller (p = 0.01) than those calculated by dividing tile sedimentation rates by corresponding bulk densities; elevation changes were similar to corresponding calculations with core data (p > 0.1) (Fig. 5).

Box plots of elevation changes in years 1 (February 2011–2012) and 2 (February 2012–2013) from annual surveys (left to right) – observed, calculated with core sedimentation rates, calculated with tile sedimentation rates. Error bars represent 1 standard deviation; open circles denote outliers
During years 1 and 2, inundation duration, frequency, and depth were positively linearly correlated with tile sedimentation rates (Fig. 6; Table 2) across the marsh, on the banks, and in the interior, except for inundation frequency and bank rates. No linear regression models were significant for core sedimentation rates; however, quadratic regressions were significant across the marsh and in the interior. Organic content was negatively correlated with inundation duration, frequency, and depth, for both tiles and cores across the marsh, on the banks, and in the interior.

Average sedimentation rate versus average inundation time from tiles (gray diamonds) and cores (black boxes) for years 1 and 2. Best-fit regression lines are shown; linear for tiles, quadratic for cores (see Table 2 for statistics). Note all tile rates are positive, whereas core rates are both positive and negative
Discussion
The relationship between sediment supply and delivery to marshes is complex, with feedbacks between water level and SSC ultimately driving long-term marsh stability. Tidal marshes must accumulate sediment at approximately the rate of RSLR to maintain long-term equilibrium, which depends on both available SSC in adjacent waters and net sediment transport into the marsh. In this study, the observed temporal variability in tile- and core-measured sedimentation across 5 years could be explained by subtidal water level, not SSC, suggesting that any amount of SSC in adjacent waters will not affect marsh accretion unless it is transported into the marsh through directional transport processes. These processes are complex, because they are influenced by locally and regionally interacting drivers of water level (Salas-Monreal and Valle-Levinson 2008; Ganju et al. 2013).
Separating the influences of SSC and water level on sedimentation was hindered in this study by lack of spatially variable SSC data throughout the marsh. Expected sedimentation rates, calculated with SSC, water level, and estimated particle-settling velocities, were statistically similar to observed rates on tiles, highlighting the importance of fluvial sediment for inorganic marsh sedimentation. However, the linear regression models of observed versus expected rates and observed rates versus water level had similar statistics (p < 0.05; R2 = 0.23 and R2 = 0.24, respectively), suggesting water level as the main driver, at least averaged over individual deployments. In contrast, expected and observed rates in cores were statistically distinct, likely because calculations capture only deposition whereas cores integrate both deposition and erosion. This result, and the correspondence of core sedimentation rates to observed elevation changes, shows the importance of erosion in determining net sedimentation and ultimately elevation change.
The dominance of water level as a driver of marsh sedimentation in this study is interesting in light of recent work emphasizing the role of SSC in driving spatiotemporal sedimentation patterns and morphological responses to environmental change (D’Alpaos et al. 2011; Stralberg et al. 2011; Butzeck et al. 2015). Observational studies also highlight the role of SSC in determining spatial sedimentation patterns (Moskalski and Sommerfield 2012). However, the link between SSC and sedimentation can be relatively weak (French 2006), and even if SSC and sedimentation are positively linked, a marsh with higher SSC is not necessarily more stable over time (Ganju et al. 2015). SSC reflects the quantity of sediment available for transport; the net direction of transport ultimately determines its fate. Sediment supply clearly limits accretion rates in systems with low SSC in adjacent waters, and a lag may exist between changes in SSC and delivery to the marsh platform (D’Alpaos et al. 2011). However, variability in supply is relatively unimportant for systems like DMP, with plentiful fluvial sediment supply (Palinkas and Engelhardt 2016), because there is “enough” sediment to nourish the marsh. Instead, the mechanism of supply is critical.
Marsh inundation provides insight into one possible transport mechanism – water-level changes influenced by tides, local and remote weather (temperature, winds, atmospheric pressure), Chesapeake Bay circulation patterns, and propagation of Atlantic Ocean water-level changes. Generally, sedimentation rates and the duration and frequency of inundation are positively correlated (Cahoon and Reed 1995; Leonard and Luther 1995; Leonard 1997, among others). At DMP, inundation frequency was indeed positively correlated with tile sedimentation rates (deposition was directly related to sediment supply) but not linearly related to core sedimentation rates. Instead, a quadratic regression fit was significant. In this model, rates increase with inundation frequency to ~30–40%, then decrease, suggesting a local optimum of sedimentation. Cadol et al. (2014) observed a similar localized maximum of elevation gain at mid-elevations in DMP; together these results indicate that increased inundation could increase marsh vulnerability to rapid RSLR. The quadratic model was significant only for sites in the interior, suggesting that other processes are more dominant on the banks (storm deposition, flow routing by vegetation; Cahoon and Reed 1995; Temmerman et al. 2005; Thorne et al. 2014; Vandenbruwaene et al. 2015; Nardin et al. 2016).
We offer two explanations for the observed localized sedimentation maximum, which are not mutually exclusive or the only possible processes but rather suggest foci for future study. The first explanation relates to feedbacks with the vegetation community; productivity in species-rich TFM (Hopfensperger and Engelhardt 2008; Elmore et al. 2016) and Mediterranean salt marshes (Day et al. 1999; Silvestri et al. 2005; D’Alpaos et al. 2007; Marani et al. 2010) increases with elevation. At DMP, stem counts and above-ground biomass of erect but senesced plants at tile sites were higher at higher elevations (Elmore et al. 2016). Moreover, sites inundated <35% of the time, the midpoint of “optimal window” suggested above, had significantly higher stem counts than those inundated more often (p < 0.001). The reduction in near-bed shear stresses associated with dense vegetation communities (Mudd et al. 2010; Elmore et al. 2016; Nardin et al. 2016) could facilitate sediment settling onto the marsh surface at mid to high elevations; this process should be weaker in sparser communities common at low elevations.
Another explanation for the optimal sedimentation response relates to the “settling and scour lag” concept well established for supporting stable tidal landforms (Postma 1967; Pejrup 1988) and observed in field studies (Reed et al. 1999; Temmerman et al. 2003b). This concept is that tidal waters carry suspended sediments into the marsh during flood tide, with some sediment settling out of suspension and onto the surface during the following slack tide. Waters receding during ebb tide carry less sediment (Wang et al. 2017) due to enhanced shear stresses required for sediment resuspension. Water inundating the marsh on the next flood tide can be “recharged” with sediment via exchange with adjacent waters, resupplying sediment to the marsh and building elevation over time. We hypothesize that long flood durations limit the opportunity for “recharge”, delivering less sediment to the marsh over time. While fully testing this hypothesis requires measuring SSC in flood and ebb tides across a range of inundation times and frequencies, core and tile sedimentation rates provide preliminary support. Core and tile data show that sedimentation increases with longer inundation times, higher average water depth, and more frequent inundation (Fig. 6, Table 2). However, core rates decrease for the longest inundation times, water depth, and inundation frequency, suggesting a potential lack of complete recharge that will build less elevation over time and enhance marsh vulnerability to RSLR.
Feedbacks among inundation, sedimentation, and elevation could enhance marsh resiliency to environmental changes. Maintenance of equilibrium elevations requires adjusting inorganic and/or organic sedimentation to perturbations in elevation (Marani et al. 2013). When elevation is lowered in both salt marshes and TFM, inorganic sedimentation is enhanced, because marsh surfaces are inundated longer, deeper, and/or more frequently, increasing supply from adjacent waters and resulting in elevation gain. In contrast, organic-matter deposition from high above-ground production increases at higher elevations in TFM and Mediterranean salt marshes (but not some North American salt marshes; Morris et al. 2002; Kirwan and Murray 2007), allowing organic sedimentation to dominate and potentially offset lower inorganic sedimentation at higher elevations (Elmore et al. 2016). Repetition of these patterns over time allows the marsh to maintain an equilibrium elevation. Our results from 2011 to 2012 and 2012–2013 support these ideas for the study TFM, at least conceptually, as does the maintenance of sites as low versus high marsh over the 5-years study. However, the widespread elevation loss in 2013–2014 is striking in that feedbacks in low and high marshes appear similar. Inorganic and organic sedimentation rates that year were much lower than the previous 2 years; however, average organic content was slightly higher (42% compared to 39% for 2011–2012 and 2012–2013), and plant litter production was consistent across years (Elmore et al. 2016). Thus, loss of inorganic input appears to drive the sedimentation deficit. Water levels were much lower in 2013–2014, particularly from November–January when they were significantly lower than previous years (p < 0.001), and the corresponding decrease in inundation offers an explanation for reduced inorganic sediment supply.
The hypotheses of an optimal response versus self-sustaining feedbacks are not mutually exclusive, and there are many subtleties that are beyond the scope of this study. For example, the influence of water level is weaker in the interior than on the banks, likely because flooding waters must overtop the banks before they can reach the interior (Lagomasino et al. (2013) but see Ensign and Currin (2017)). While further research is needed to fully explore these hypotheses, this study provides evidence that the mechanism of sediment supply (inundation) can be more important than the quantity of sediment available for transport (SSC), which is critical insight for improving models of marsh response to anticipated environmental changes. This study also highlights the need for measuring the SSC that is actually delivered to the marsh platform to assess potential marsh vulnerability to environmental change.
Summary
This study presented results from 5 years of bimonthly sedimentation observations at DMP with coincident time-series of physical drivers (winds, river discharge, water level, and SSC). The main objective was to understand the influence of these physical drivers on the spatiotemporal variability of marsh sedimentation, so that potential marsh responses to environmental changes can be better predicted.
Local winds were correlated with both water level and SSC. However, these correlations did not explain the majority of the variability in water level or SSC, highlighting other local and remote influences as physical drivers. Sediment organic content was relatively stable over space and time, while sedimentation rates varied widely. The only single physical driver that was significantly related to sedimentation rates in linear regression models was water level and, correspondingly, inundation time. Inundation time and sedimentation rates were positively linearly related for tile observations (deposition only), as expected, but a quadratic regression model was required for core observations that integrate sediment deposition and erosion and correspond to observed elevation changes.
The dominance of water level, not SSC, as a driver of marsh sedimentation in a system with plentiful sediment supply is a key result of this study and has important implications for marsh resiliency to future environmental changes. Results support two, non-mutually exclusive hypotheses for marsh response to RSLR. The first proposes an optimal response of sedimentation to increasing inundation, such that sedimentation increases to some critical value then decreases. This response highlights the importance of water-level oscillations: at longer inundation times, flooding waters have limited opportunities to “recharge” with sediment from adjacent waters, delivering less sediment over time and enhancing marsh vulnerability to SLR. Another hypothesis posits that self-sustaining, cyclical feedbacks among inundation, sedimentation, and elevation maintain equilibrium elevations over time. Fully testing these hypotheses in future work is critical for improving models of marsh response to anticipated environmental changes.
References
Allen J (2000) Morphodynamics of Holocene salt marshes: a review sketch from the Atlantic and southern North Sea coasts of Europe. Quaternary Science Reviews 19:1155–1231. https://doi.org/10.1016/S0277-3791(99)00034-7
Barbier EB, Georgiou IY, Enchelmeyer B, Reed DJ (2013) The value of wetlands in protecting Southeast Louisiana from hurricane storm surges. PLoS One 8:e58715. https://doi.org/10.1371/journal.pone.0058715
Beck MW, Heck KL, Able KW et al (2003) The role of nearshore ecosystems as fish and shellfish nurseries. Issues in Ecology 11:1–14
Belliard J-P, Toffolon M, Carniello L, D’Alpaos A (2015) An ecogeomorphic model of tidal channel initiation and elaboration in progressive marsh accretional contexts. Journal of Geophysical Research - Earth Surface 120:1040–1064. https://doi.org/10.1002/2015JF003445
Boon JD, Brubaker JM, Forrest DR (2010) Chesapeake Bay land subsidence and sea level change: an evaluation of past and present trends and future Outlook. Report No. 425 in Applied Marine Science and Ocean Engineering. Virginia Institute of Marine Science Special, Gloucester Point, 81
Brakebill JW, Ator SW, Schwarz GE (2010) Sources of suspended-sediment flux in streams of the Chesapeake Bay watershed: a regional application of the SPARROW model. Journal of the American Water Resources Association 46:757–776. https://doi.org/10.1111/j.1752-1688.2010.00450.x
Bruland GL (2008) Coastal wetlands: function and role in reducing impact of land-based management. Coastal Watershed Management 13:85
Butzeck C, Eschenbach A, Gröngröft A, Hansen K, Nolte S, Jensen K (2015) Sediment deposition and accretion rates in tidal marshes are highly variable along estuarine salinity and flooding gradients. Estuaries and Coasts 38:434–450. https://doi.org/10.1007/s12237-014-9848-8
Cadol D, Engelhardt K, Elmore A, Sanders G (2014) Elevation-dependent surface elevation gain in a tidal freshwater marsh and implications for marsh persistence. Limnology and Oceanography 59:1065–1080. https://doi.org/10.4319/lo.2014.59.3.1065
Cahoon DR, Reed DJ (1995) Relationships among marsh surface-topography, hydroperiod, and soil accretion in a deteriorating Lousiana salt-marsh. Journal of Coastal Research 11:357–369
Carter V, Rybicki NB, Landwehr JM, Turtora M (1994) Role of weather and water quality in population dynamics of submersed macrophytes in the tidal Potomac River. Estuaries 17:417–426
Casey WH, Larsen IL, Olsen CR (1986) The distribution of cosmogenic 7Be in salt marsh sediments. Geophysical Research Letters 13:322–325. https://doi.org/10.1029/GL013i004p00322
Chmura GL, Hung GA (2004) Controls on salt marsh accretion: a test in salt marshes of eastern Canada. Estuaries 27:70–81. https://doi.org/10.1007/BF02803561
Christiansen T, Wiberg PL, Milligan TG (2000) Flow and sediment transport on a tidal salt marsh surface. Estuarine, Coastal and Shelf Science 50:315–331. https://doi.org/10.1006/ecss.2000.0548
Chuang W-S, Boicourt WC (1989) Resonant seiche motion in the Chesapeake Bay. Journal of Geophysical Research 94:2105. https://doi.org/10.1029/JC094iC02p02105
Coulter CB, Kolka RK, Thompson JA (2004) Water quality in agricultural, urban, and mixed land use watersheds. Journal of the American Water Resources Association 40:1593–1601. https://doi.org/10.1111/j.1752-1688.2004.tb01608.x
Craft C, Clough J, Ehman J, Joye S, Park R, Pennings S, Guo H, Machmuller M (2009) Forecasting the effects of accelerated sea-level rise on tidal marsh ecosystem services. Frontiers in Ecology and the Environment 7:73–78. https://doi.org/10.1890/070219
D’Alpaos A, Lanzoni S, Marani M, Rinaldo A (2007) Landscape evolution in tidal embayments: modeling the interplay of erosion, sedimentation, and vegetation dynamics. Journal of Geophysical Research. https://doi.org/10.1029/2006JF000537
D’Alpaos A, Mudd SM, Carniello L (2011) Dynamic response of marshes to perturbations in suspended sediment concentrations and rates of relative sea level rise. Journal of Geophysical Research. https://doi.org/10.1029/2011JF002093
Day J, Rybczyk J, Scarton F et al (1999) Soil accretionary dynamics, sea-level rise and the survival of wetlands in Venice lagoon: a field and modelling approach. Estuarine, Coastal and Shelf Science 49:607–628. https://doi.org/10.1006/ecss.1999.0522
Day JW, Boesch DF, Clairain EJ, Kemp GP, Laska SB, Mitsch WJ, Orth K, Mashriqui H, Reed DJ, Shabman L, Simenstad CA, Streever BJ, Twilley RR, Watson CC, Wells JT, Whigham DF (2007) Restoration of the Mississippi Delta: lessons from hurricanes Katrina and Rita. Science 315:1679–1684. https://doi.org/10.1126/science.1137030
Day JW, Christian RR, Boesch DM, Yáñez-Arancibia A, Morris J, Twilley RR, Naylor L, Schaffner L, Stevenson C (2008) Consequences of climate change on the ecogeomorphology of coastal wetlands. Estuaries and Coasts 31:477–491. https://doi.org/10.1007/s12237-008-9047-6
Dibb JE, Rice DL (1989) The geochemistry of beryllium-7 in Chesapeake Bay. Estuarine, Coastal and Shelf Science 28:379–394. https://doi.org/10.1016/0272-7714(89)90086-3
Elliott AJ (1978) Observations of the meteorologically induced circulation in the Potomac estuary. Estuarine and Coastal Marine Science 6:285–299. https://doi.org/10.1016/0302-3524(78)90017-8
Elmore AJ, Engelhardt KAM, Cadol D, Palinkas CM (2016) Spatial patterns of plant litter in a tidal freshwater marsh and implications for marsh persistence. Ecological Applications 26:846–860. https://doi.org/10.1890/14-1970.1
Engelhart SE, Horton BP, Douglas BC, Peltier WR, Tornqvist TE (2009) Spatial variability of late Holocene and 20th century sea-level rise along the Atlantic coast of the United States. Geology 37:1115–1118. https://doi.org/10.1130/G30360A.1
Ensign SH, Currin C (2017) Geomorphic implications of particle movement by water surface tension in a salt marsh. Wetlands 37:245–256. https://doi.org/10.1007/s13157-016-0862-4
Erftemeijer PLA, Koch EW (2001) Sediment geology methods for seagrass habitat. Global seagrass research methods. Elsevier Science B.V, Amsterdam, pp 345–367
Fagherazzi S, Mariotti G, Wiberg P, McGlathery K (2013) Marsh collapse does not require sea level rise. Oceanography 26:70–77. https://doi.org/10.5670/oceanog.2013.47
Feagin RA, Martinez ML, Mendoza-Gonzalez G, Costanza R (2010) Salt marsh zonal migration and ecosystem service change in response to global sea level rise: a case study from an urban region. Ecology and Society 15(4):14
Fisher TR, Lee KY, Berndt H, Benitez JA, Norton MM (1998) Hydrology and chemistry of the Choptank River basin. Water, Air, and Soil Pollution 105:387–397
French J (2006) Tidal marsh sedimentation and resilience to environmental change: exploratory modelling of tidal, sea-level and sediment supply forcing in predominantly allochthonous systems. Marine Geology 235:119–136. https://doi.org/10.1016/j.margeo.2006.10.009
French JR, Stoddart DR (1992) Hydrodynamics of salt marsh creek systems: implications for marsh morphological development and material exchange. Earth Surface Processes and Landforms 17:235–252. https://doi.org/10.1002/esp.3290170304
Friedrichs CT, Perry JE (2001) Tidal salt marsh morphodynamics: a synthesis. Journal of Coastal Research Special Issue 27:7–37
Ganju NK, Nidzieko NJ, Kirwan ML (2013) Inferring tidal wetland stability from channel sediment fluxes: observations and a conceptual model. Journal of Geophysical Research - Earth Surface 118:2045–2058. https://doi.org/10.1002/jgrf.20143
Ganju NK, Kirwan ML, Dickhudt PJ, Guntenspergen GR, Cahoon DR, Kroeger KD (2015) Sediment transport-based metrics of wetland stability. Geophysical Research Letters 42:7992–8000. https://doi.org/10.1002/2015GL065980
Harper JD, Heliotis FD (1992) Dyke marsh hydrologic model. George Mason University
Hopfensperger KN, Engelhardt KAM (2008) Annual species abundance in a tidal freshwater marsh: germination and survival across an elevational gradient. Wetlands 28:521–526. https://doi.org/10.1672/07-117.1
Hopfensperger KN, Engelhardt KAM, Lookingbill TR (2009) Vegetation and seed bank dynamics in a tidal freshwater marsh. J Veg Sci 20:767–778
Johnston DW (2000) The dyke marsh preserve ecosystem. Virginia Journal of Science 51:223–272
Kaste JM, Elmore AJ, Vest KR, Okin GS (2011) Beryllium-7 in soils and vegetation along an arid precipitation gradient in Owens Valley, California. Geophysical Research Letters 38. https://doi.org/10.1029/2011GL047242
Kim G, Hussain N, Scudlark JR, Church TM (2000) Factors influencing the atmospheric depositional fluxes of stable Pb, 210Pb, and 7Be into Chesapeake Bay. Journal of Atmospheric Chemistry 36:65–79
Kirwan ML, Megonigal JP (2013) Tidal wetland stability in the face of human impacts and sea-level rise. Nature 504:53–60. https://doi.org/10.1038/nature12856
Kirwan ML, Murray AB (2007) A coupled geomorphic and ecological model of tidal marsh evolution. Proceedings of the National Academy of Sciences 104:6118–6122. https://doi.org/10.1073/pnas.0700958104
Kirwan ML, Guntenspergen GR, D’Alpaos A, et al (2010) Limits on the adaptability of coastal marshes to rising sea level. Geophysical Research Letters 37:n/a-n/a. https://doi.org/10.1029/2010GL045489
Kirwan ML, Murray AB, Donnelly JP, Corbett DR (2011) Rapid wetland expansion during European settlement and its implication for marsh survival under modern sediment delivery rates. Geology 39:507–510. https://doi.org/10.1130/G31789.1
Lagomasino D, Corbett DR, Walsh JP (2013) Influence of wind-driven inundation and coastal geomorphology on sedimentation in two microtidal marshes, Pamlico River estuary, NC. Estuaries and Coasts 36:1165–1180. https://doi.org/10.1007/s12237-013-9625-0
Larsen I, Cutshall N (1981) Direct determination of 7Be in sediments. Earth and Planetary Science Letters 54:379–384. https://doi.org/10.1016/0012-821X(81)90053-4
Leck MA, Simpson RL (1995) Ten-years seed bank and vegetation dynamics of a tidal freshwater marsh. American Journal of Botany 82:1547. https://doi.org/10.2307/2446183
Leonard LA (1997) Controls of sediment transport and deposition in an incised mainland marsh basin, southeastern North Carolina. Wetlands 17:263–274. https://doi.org/10.1007/BF03161414
Leonard LA, Luther ME (1995) Flow hydrodynamics in tidal marsh canopies. Limnology and Oceanography 40:1474–1484
Leonardi N, Ganju NK, Fagherazzi S (2016) A linear relationship between wave power and erosion determines salt-marsh resilience to violent storms and hurricanes. Proceedings of the National Academy of Sciences 113:64–68. https://doi.org/10.1073/pnas.1510095112
Litwin RJ, Smoot JP, Pavich MJ, et al (2011) Analysis of the deconstruction of dyke marsh, George Washington Memorial Parkway, Virginia: progression, geologic and manmade causes, and effective restoration scenarios. USGS Open File Report 2010–1269. 92 pp
Litwin RJ, Smoot JP, Pavich MJ, Oberg E, Steury B, Helwig B, Markewich HW, Santucci VL, Sanders G (2013) Rates and probable causes of freshwater tidal marsh failure, Potomac River estuary, northern Virginia, USA. Wetlands 33:1037–1061. https://doi.org/10.1007/s13157-013-0461-6
Loomis MJ, Craft CB (2010) Carbon sequestration and nutrient (nitrogen, phosphorus) accumulation in river-dominated tidal marshes, Georgia, USA. Soil Science Society of America Journal 74:1028. https://doi.org/10.2136/sssaj2009.0171
Luisetti T, Jackson EL, Turner RK (2013) Valuing the European ‘coastal blue carbon’ storage benefit. Marine Pollution Bulletin 71:101–106. https://doi.org/10.1016/j.marpolbul.2013.03.029
Lynch JC, Hensel P, Cahoon DR (2015) The surface elevation table and marker horizon technique: a protocol for monitoring wetland elevation dynamics. National Park Service Report NPS/NCBN/NRR–2015/1078
Marani M, D’Alpaos A, Lanzoni S et al (2010) The importance of being coupled: stable states and catastrophic shifts in tidal biomorphodynamics. Journal of Geophysical Research. https://doi.org/10.1029/2009JF001600
Marani M, D’Alpaos A, Lanzoni S, Santalucia M (2011) Understanding and predicting wave erosion of marsh edges. Geophysical Research Letters 38:n/a-n/a. https://doi.org/10.1029/2011GL048995
Marani M, Da Lio C, D’Alpaos A (2013) Vegetation engineers marsh morphology through multiple competing stable states. Proceedings of the National Academy of Sciences 110:3259–3263. https://doi.org/10.1073/pnas.1218327110
Morris JT, Sundareshwar PV, Nietch CT, Kjerfve B, Cahoon DR (2002) Responses of coastal wetlands to rising sea level. Ecology 83:2869–2877. https://doi.org/10.2307/3072022
Moskalski SM, Sommerfield CK (2012) Suspended sediment deposition and trapping efficiency in a Delaware salt marsh. Geomorphology 139–140:195–204. https://doi.org/10.1016/j.geomorph.2011.10.018
Moskalski S, Torres R (2012) Influences of tides, weather, and discharge on suspended sediment concentration. Continental Shelf Research 37:36–45. https://doi.org/10.1016/j.csr.2012.01.015
Mudd SM, Howell SM, Morris JT (2009) Impact of dynamic feedbacks between sedimentation, sea-level rise, and biomass production on near-surface marsh stratigraphy and carbon accumulation. Estuarine, Coastal and Shelf Science 82:377–389. https://doi.org/10.1016/j.ecss.2009.01.028
Mudd SM, D’Alpaos A, Morris JT (2010) How does vegetation affect sedimentation on tidal marshes? Investigating particle capture and hydrodynamic controls on biologically mediated sedimentation. Journal of Geophysical Research. https://doi.org/10.1029/2009JF001566
Murphy S, Voulgaris G (2006) Identifying the role of tides, rainfall and seasonality in marsh sedimentation using long-term suspended sediment concentration data. Marine Geology 227:31–50. https://doi.org/10.1016/j.margeo.2005.10.006
Nardin W, Edmonds DA, Fagherazzi S (2016) Influence of vegetation on spatial patterns of sediment deposition in deltaic islands during flood. Advances in Water Resources 93:236–248. https://doi.org/10.1016/j.advwatres.2016.01.001
Neubauer SC (2008) Contributions of mineral and organic components to tidal freshwater marsh accretion. Estuarine, Coastal and Shelf Science 78:78–88. https://doi.org/10.1016/j.ecss.2007.11.011
Neubauer SC, Anderson IC, Constantine JA, Kuehl SA (2002) Sediment deposition and accretion in a mid-Atlantic (U.S.a.) tidal freshwater marsh. Estuarine, Coastal and Shelf Science 54:713–727. https://doi.org/10.1006/ecss.2001.0854
Nicholls RJ, Marinova N, Lowe JA, Brown S, Vellinga P, de Gusmao D, Hinkel J, Tol RSJ (2011) Sea-level rise and its possible impacts given a “beyond 4 C world” in the twenty-first century. Philosophical Transactions of the Royal Society A - Mathematical Physical and Engineering Sciences 369:161–181. https://doi.org/10.1098/rsta.2010.0291
NPS (National Park Service) (1977) Environmental assessment of dyke marsh. National Capital Region, National Park Service, US Department of the Interior, Washington, DC
Nyman JA, Walters RJ, Delaune RD, Patrick WH (2006) Marsh vertical accretion via vegetative growth. Estuarine, Coastal and Shelf Science 69:370–380. https://doi.org/10.1016/j.ecss.2006.05.041
Oczkowski A, Wigand C, Hanson A, Markham E, Miller KM, Johnson R (2016) Nitrogen retention in salt marsh systems across nutrient-enrichment, elevation, and precipitation regimes: a multiple-stressor experiment. Estuaries and Coasts 39:68–81. https://doi.org/10.1007/s12237-015-9975-x
Olsen CR, Larsen IL, Lowry PD, Cutshall NH, Nichols MM (1986) Geochemistry and deposition of 7Be in river-estuarine and coastal waters. Journal of Geophysical Research 91:896–908. https://doi.org/10.1029/JC091iC01p00896
Ouyang X, Lee SY (2014) Updated estimates of carbon accumulation rates in coastal marsh sediments. Biogeosciences 11:5057–5071. https://doi.org/10.5194/bg-11-5057-2014
Palinkas CM, Engelhardt KAM (2016) Spatial and temporal patterns of modern (~100 y) sedimentation in a tidal freshwater marsh: implications for future sustainability. Limnology and Oceanography 61:132–148
Palinkas CM, Engelhardt KAM, Cadol D (2013) Evaluating physical and biological influences on sedimentation in a tidal freshwater marsh with 7Be. Estuarine, Coastal and Shelf Science 129:152–161. https://doi.org/10.1016/j.ecss.2013.05.022
Palinkas CM, Halka JP, Li M, Sanford LP, Cheng P (2014) Sediment deposition from tropical storms in the upper Chesapeake Bay: field observations and model simulations. Continental Shelf Research 86:6–16. https://doi.org/10.1016/j.csr.2013.09.012
Pasarić M, Orlić M (2001) Long-term meteorological preconditioning of the north Adriatic coastal floods. Continental Shelf Research 21:263–278
Pasternack GB, Brush GS (1998) Sedimentation cycles in a river-mouth tidal freshwater marsh. Estuaries 21:407–415. https://doi.org/10.2307/1352839
Pejrup M (1988) Suspended sediment transport across a tidal flat. Marine Geology 82:187–198. https://doi.org/10.1016/0025-3227(88)90140-5
Pendleton L, Donato DC, Murray BC, Crooks S, Jenkins WA, Sifleet S, Craft C, Fourqurean JW, Kauffman JB, Marbà N, Megonigal P, Pidgeon E, Herr D, Gordon D, Baldera A (2012) Estimating global “blue carbon” emissions from conversion and degradation of vegetated coastal ecosystems. PLoS One 7:e43542. https://doi.org/10.1371/journal.pone.0043542
Postma H (1967) Sediment transport and sedimentation in the marine environment. Estuaries. Washington, DC, pp 158–179
Rahmstorf S, Foster G, Cazenave A (2012) Comparing climate projections to observations up to 2011. Environmental Research Letters 7:044035. https://doi.org/10.1088/1748-9326/7/4/044035
Redfield AC (1972) Development of a New England salt marsh. Ecological Monographs 42:201–237. https://doi.org/10.2307/1942263
Reed DJ (1995) The response of coastal marshes to sea-level rise: survival or submergence? Earth Surface Processes and Landforms 20:39–48. https://doi.org/10.1002/esp.3290200105
Reed DJ, Spencer T, Murray AL, French JR, Leonard L (1999) Marsh surface sediment deposition and the role of tidal creeks: implications for created and managed coastal marshes. Journal of Coastal Conservation 5:81–90
Salas-Monreal D, Valle-Levinson A (2008) Sea-level slopes and volume fluxes produced by atmospheric forcing in estuaries: Chesapeake Bay case study. Journal of Coastal Research 2:208–217. https://doi.org/10.2112/06-0632.1
Sallenger AH, Doran KS, Howd PA (2012) Hotspot of accelerated sea-level rise on the Atlantic coast of North America. Nature Climate Change 2:884–888. https://doi.org/10.1038/nclimate1597
Schoellhamer DH (2002) Variability of suspended-sediment concentration at tidal to annual time scales in San Francisco Bay, USA. Continental Shelf Research 22:1857–1866. https://doi.org/10.1016/S0278-4343(02)00042-0
Silvestri S, Defina A, Marani M (2005) Tidal regime, salinity and salt marsh plant zonation. Estuarine, Coastal and Shelf Science 62:119–130. https://doi.org/10.1016/j.ecss.2004.08.010
Simpson RL, Good RE, Leck MA, Whigham DF (1983) The ecology of freshwater tidal wetlands. Bioscience 33:255–259
Stralberg D, Brennan M, Callaway JC, Wood JK, Schile LM, Jongsomjit D, Kelly M, Parker VT, Crooks S (2011) Evaluating tidal marsh sustainability in the face of sea-level rise: a hybrid modeling approach applied to San Francisco Bay. PLoS One 6:e27388. https://doi.org/10.1371/journal.pone.0027388
Stumpf RP (1983) The process of sedimentation on the surface of a salt marsh. Estuarine, Coastal and Shelf Science 17:495–508. https://doi.org/10.1016/0272-7714(83)90002-1
Temmerman S, Govers G, Meire P, Wartel S (2003a) Modelling long-term tidal marsh growth under changing tidal conditions and suspended sediment concentrations, Scheldt estuary, Belgium. Marine Geology 193:151–169
Temmerman S, Govers G, Wartel S, Meire P (2003b) Spatial and temporal factors controlling short-term sedimentation in a salt and freshwater tidal marsh, Scheldt estuary, Belgium, SW Netherlands. Earth Surface Processes and Landforms 28:739–755. https://doi.org/10.1002/esp.495
Temmerman S, Bouma TJ, Govers G, Wang ZB, de Vries MB, Herman PMJ (2005) Impact of vegetation on flow routing and sedimentation patterns: three-dimensional modeling for a tidal marsh. Journal of Geophysical Research 110. https://doi.org/10.1029/2005JF000301
Thorne KM, Elliott-Fisk DL, Wylie GD, Perry WM, Takekawa JY (2014) Importance of biogeomorphic and spatial properties in assessing a tidal salt marsh vulnerability to sea-level rise. Estuaries and Coasts 37:941–951. https://doi.org/10.1007/s12237-013-9725-x
Torres R, Fagherazzi S, vanProosdij D, Hopkinson C (2006) Salt marsh geomorphology: physical and ecological effects on landform. Estuarine, Coastal and Shelf Science 69:309–310. https://doi.org/10.1016/j.ecss.2006.05.001
Vandenbruwaene W, Schwarz C, Bouma TJ, Meire P, Temmerman S (2015) Landscape-scale flow patterns over a vegetated tidal marsh and an unvegetated tidal flat: implications for the landform properties of the intertidal floodplain. Geomorphology 231:40–52. https://doi.org/10.1016/j.geomorph.2014.11.020
Wang A, Ye X, Du Y, Yin X (2017) Hydrodynamic and biological mechanisms for variations in near-bed suspended sediment concentrations in a Spartina alterniflora marsh—a case study of Luoyuan Bay, China. Estuaries and Coasts 40:1540–1550. https://doi.org/10.1007/s12237-017-0253-y
Weston NB (2014) Declining sediments and rising seas: an unfortunate convergence for tidal wetlands. Estuaries and Coasts 37:1–23. https://doi.org/10.1007/s12237-013-9654-8
Acknowledgements
The authors thank many students and volunteers for field and laboratory assistance throughout this study, especially Debbie Hinkle, Lisa Wu, and Dan Cadol. Erik Oberg and Brent Steury of the National Park Service provided critical access and institutional knowledge of DMP. We thank several anonymous reviewers for constructive comments on previous drafts. The datasets generated during and/or analyzed during the current study are available from the corresponding author by request. This project was supported by the National Science Foundation grant DEB-0841394 and the Grayce B. Kerr Fund. This is UMCES contribution #5517.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Palinkas, C.M., Engelhardt, K.A.M. Influence of Inundation and Suspended-Sediment Concentrations on Spatiotemporal Sedimentation Patterns in a Tidal Freshwater Marsh. Wetlands 39, 507–520 (2019). https://doi.org/10.1007/s13157-018-1097-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-018-1097-3