
Abstract
Effective monitoring requires clear questions and a well-designed sampling regime. However, objectives often evolve over time which can render the initial sampling design ineffective. Using a vegetation monitoring program employed in Newnes Plateau Shrub Swamps, Australia, as a case study, we examine a sampling design based on small numbers of 400 m2 plots to assess if it can meet the stated monitoring objectives of detecting significant changes in number and abundance of species per wetland. To determine this, we intensively sampled four monitored wetlands using randomly distributed 4 m2 plots to obtain representative estimates of species composition and abundance. The 400 m2 plots captured 91 % of the common species and a similar proportional distribution of life-forms as found in the 4 m2 plots, but missed 62 % of the sparse species found in 4 m2 plots. Insufficient replication of 400 m2 plots made detection of statistically significant changes at the swamp scale difficult or impossible. Our review showed the weak sampling design was contributed to by 1) an initial lack of clearly stated management triggers and 2) changes in monitoring objectives and triggers over time, without revising the sampling design. We highlight the need for an adaptive approach to monitoring.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Effective and robust monitoring requires an adaptive and data driven approach (Lindenmayer and Likens 2009). However, there is strong tendency to develop non-intensive and rapid-assessment methods at the expense of statistical rigour in monitoring ecological systems (Lee et al. 2005). Often in an attempt to increase efficiency and decrease expense, a smaller number of larger plots are used. This can lead to higher sampling error, e.g. not detecting a species which is present or miss-classifying the species abundance, and lower power to reject the null hypothesis. High (or unknown) sampling error in a monitoring program can lead to missing or miss-identifying impacts such as changes in presence or abundance of rare or weedy species, with flow on effects for implementation of management actions. Here, we review an existing wetland monitoring program, to examine if the low-intensity, rapid-assessment sampling method used can achieve the stated objectives of detecting a statistically significant change in species diversity and abundance due to anthropogenic impacts. We then discuss the limitations of the current monitoring program and suggest several modifications that could improve the existing sampling design.
Often, the overarching aim of a monitoring program is to detect adverse environmental impacts early, before catastrophic damage occurs. To achieve this aim, it is necessary to collect data for a set of relevant ecological indicators, with sufficient statistical power to detect changes of concern. The indicators chosen need to reflect the type and magnitude of predicted changes and impacts of concern, e.g. increased depth to water table would be predicted to reduce hydrophyte abundance, and hence hydrophyte abundance could be an indicator. Common ecological indicators include changes in soil nutrient levels and litter accumulation (Tongway and Hindley 2004) or changes in species density, or number of functional groups or life-form groups present (Philippi et al. 1998; Stevens et al. 2004; Ramsay et al. 2006). To detect whether environmental impacts have occurred, it is necessary to accurately and efficiently capture a change in a chosen indicator, such as species abundance, which requires careful consideration of sampling methods, e.g. plot size, replication and spatial arrangement (Gitay et al. 1991; Hellmann and Fowler 1999; Gotelli and Colwell 2001; Scheiner et al. 2011). For example increasing the size of a plot has been shown to decrease the accuracy of both the abundance estimates and detection of plant species (Jalonen et al. 1998; Klimes et al. 2001; Archaux et al. 2007). Modifying the sampling extent changes the probability of encountering rare species or ‘hot spots’ that support a high number of species (Palmer and White 1994). A change in sample size (replication) has a direct impact on statistical power (Downs et al. 2002; Legg and Nagy 2006). The sampling methodology employed should be focused on efficiently detecting unacceptable changes that are attributable to the perceived threats. Nevertheless, sampling methodologies are often based on (i) ‘rules-of-thumb’, (ii) funding availability or (iii) human convenience rather than statistical rigour and ecological principles.
Here we review the Newnes Plateau wetland flora monitoring program where the principle of increased plot size and reduced replication has been applied in heterogeneous wetlands. The monitoring program is focused on the wetland vegetation type ‘Newnes Plateau Shrub Swamp’ (NPSS) (Benson and Baird 2012). The potential anthropogenic impacts on the hydrology of these wetland plant communities include landform deformation and/or cracking of aquitards due to subsidence from underground long-wall mines (Benson and Baird 2012); it is not known whether the hydrological impacts will be evident across an entire wetland or confined to portion(s) of a wetland. The stated objective of the monitoring is to determine whether plant species diversity, composition or abundance have been negatively impacted by changes in the water table due to underground mining. The stated triggers of the monitoring program are a) at the species level, a “significant decline in condition, health, or decline in population numbers compared to baseline”, b) “major dieback of flora compared to baseline” c) an “increase in weed species” and d) a “significant change in species diversity” within a wetland (Blick et al. 2013). The program has evolved from using visual assessments to determine ecologically significant changes to a requirement that a Before-After, Impact-Control approach (with associated statistical tests) be used to determine if statistically significant changes have occurred. In addition, the program has grown from monitoring eight wetlands to 26 individual NPSS wetlands. These 26 wetlands are monitored seasonally using between one and six 400 m2 plots per wetland, placed using expert opinion (53 plots in total), with wetlands ranging in size from less than 0.5 ha to more than 5 ha (Blick et al. 2013). A review of this program was initiated when the plot data failed to detect major vegetation die-back in a wetland. Here, we focus on reviewing the sampling design employed, and ask whether it has the potential to detect a significant change in species abundance and diversity (management triggers ‘a’ and ‘d’ above) within a wetland.
To address this question, we intensively sampled four wetlands in the current monitoring program using large numbers of randomly distributed 4 m2 plots to obtain a representative estimate of species composition and abundance. We compare the species diversity, life-form diversity, and abundance from the established sampling design (400 m2 plots) with those data obtained from the intensive survey and discuss how well data from the 400 m2 plots meet the requirements of the monitoring program. Specifically, we ask three questions:
-
1)
What proportion of the total species, common species and sparse species found in the 4 m2 plots are recorded in the 400 m2 plots in each wetland?
-
2)
Do the 400 m2 plots capture the same proportional distribution of life-form groups as the larger number of small plots distributed across each wetland?
-
3)
Are estimates of species abundance obtained in the established 400 m2 plots similar to those obtained using a larger number of small plots distributed across each wetland?
Methods
Study Site
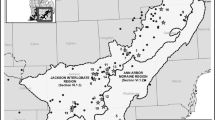
Newnes Plateau is a state forest reserve situated approximately 20 km northeast of Lithgow in the Blue Mountains, Australia (33° 23 S, 150° 12 E). It covers close to 400 km2, with elevations ranging between 950 and 1300 m ASL. The plateau is floristically diverse with approximately 600 native plant species (Royal Botanical Gardens and Domain Trust, http://www.rbgsyd.nsw.gov.au, accessed 14, March 2013) in a mosaic of forest, woodland, heath and shrub wetlands over Triassic sandstone. The endangered ecological communities ‘Newnes Plateau Shrub Swamp’ (NPSS) generally grow on a peat substrate overlaying sandstone along drainage lines and are characterised by a shrub canopy and sedge or rush (Cyperaceae, Juncaceae, Restionaceae) understorey, with occasional emergent eucalypt trees (Benson and Baird 2012).
The wetlands were selected a priori for this study because they contained at least one 400 m2 plot (i.e. were included in the current monitoring program), covered a range of sizes and hydrology types (from periodically to permanently wet), and were all located in different catchments (Table 1). Three wetlands (Clarence West, Bungleboori North, and Carne Central) each contained one 400 m2 plot, while West Wolgan contained six 400 m2 plots. The 400 m2 plots were located in the centre of the wetland in ‘representative areas’ using expert opinion (Blick et al. 2013). In some wetlands additional plots were placed at the wetland edge to capture changes occurring at higher elevations.
Data Collection: Species Composition and Abundance
In November 2012 (late spring) we sampled the current plots (400 m2 plots; n = 9) using the established method. Two researchers used the Braun-Blanquet method, with modified cover/abundance classes (1 =<5 % of plot, rare; 2 =<5 % of plot, uncommon; 3 =<5 % of plot, common; 4 =5–20 %; 5 =20–50 %; 6 =50–75 %; 7 = >75 %), with 45 min spent recording the presence and cover/abundance classes of all species found in each plot.
To obtain an estimate of species presence and abundance in each wetland we implemented a stratified random design and used species-area curves to estimate the number of species in each wetland. For the stratified random design, each wetland was divided into 25 m × 25 m grid cells and a single randomly located point was placed in each cell to locate the plot using Geospatial Modelling Environment (Beyer 2009) tools in ArcGIS 10.1 (ESRI, CA, USA). Each point located inside the mapped wetland boundary was used as the centre of a 4 m2 (2 m × 2 m) plot (Fig. 1). The plots were located in the field using a consumer-grade handheld GPS device (Juno SB, Trimble) with real-time positional accuracy of 2–5 m in good conditions. GPS accuracy varied throughout the day according to forest and cloud cover. To minimise positional bias, we considered the plot to be located when the GPS distance was less than 3 m from each waypoint, rather than attempting to take several readings over a short period which may replicate errors over time (see Dodd (2011)). As we were not trying to resample these waypoints we considered a consumer-grade GPS sufficient for locating randomly distributed plots. The number of plots in Clarence West Swamp was reduced to 12 to minimise impact on the dense fragile vegetation. All plots were divided into four contiguous 1 m2 quadrats, within which the cover of each species (defined as shoot presence) was visually estimated to the nearest 1 %. The plot size was chosen as we could be reasonably sure of detecting all species in the 1 m2 plots (Klimes et al. 2001) and of the accuracy of percentage cover estimates within plots.

Species density as modelled by kriging from the 4 m2 plot data in each wetland. Pixels = 25 m2, open squares = location of 400 m2 plot (symbol to scale); closed square = location of 4 m2 plots (symbol not to scale, plot size = 1/6 of a pixel). Examination of the variance raster associated with the kriging layer showed that the estimates were least reliable on the lobes of wetlands; where there were fewer 4 m2 plots to train the interpolation, and thus the kriged species density should be interpreted cautiously, especially at the northern and south-eastern tips of West Wolgan
Data Analysis
We used species-accumulation curves to compare the number of species recorded using 4 m2 plots with that of the 400 m2 plots. Species-accumulation curves were generated using the speccaccum function with the random algorithm from the vegan package in R (Oksanen et al. 2012; R Core Team 2012). Patterns of species density were then interpolated and mapped, using data derived from 4 m2 plots, by applying ordinary kriging (spherical model, with a 50 m search radius, and a minimum of four points) in ArcGIS 10.1.
We tested whether the proportional species recorded across six life-forms (tree, shrub, forb, grass, rush/sedge, exotic forb) within each wetland, differed according to sampling design, using chi-squared tests. All analyses were carried out using untransformed data and evaluated using the core stats package in R.
Results
Species Density
The total number of species found in each wetland using 4 m2 plots ranged between 41 (Clarence West) and 135 (Carne Central; Fig. 2). These numbers were 1.6 to 5 times greater than those found in the 400 m2 plots, with 400 m2 plots capturing a higher proportion of total species in the smaller wetlands than the larger wetlands (Fig. 2). To acquire a comparable number of species in the smaller plots as the larger plots, much less area needed to be surveyed. For example, in Carne Central, just two 4 m2 plots were required, a total sampling area of 8 m2. While in Bungleboori North Swamp, just four 4 m2 plots were required; a total of 16 m2 or 4 % of the area in a single 400 m2 plot.

Species area curves for each wetland calculated using 4 m2 plots. The vertical bars indicate the 90 % confidence interval. The horizontal line shows the number of species recorded with the 400 m2 plots for each wetland except West Wolgan where the mean number of species per plot and the total found in all six plots are shown
The proportional distribution of species across life-forms in each wetland did not differ significantly between methods, though the total number of species representing each life-form varied between methods (Fig. 3). For each life-form, the greatest number of species were recorded in 4 m2 plots, except for the rush/sedge group in Clarence West, where two additional species were recorded in the 400 m2 plot (Fig. 3).

Total number of species recorded in each life-form group by each design per wetland. Note: the y-axes differ between graphs; “Forb” includes both annual and perennial forbs; “Rush” is the rush/sedge group and includes Cyperaceae, Juncaceae, Restionaceae species; “Exotic” includes all non-native species (which happen to be all forbs). See Table 1 for number of replicates per design for each wetland
Spatial interpolation of the 4 m2 plot data showed considerable variation in species density across the wetlands (mean species = 16.6 ± 4.8 SD per 4 m2), with larger wetlands tending to show greater variation than smaller wetlands. The highest species density was recorded on the eastern edge of Carne Central, where typical wetland and forest species were intermixed, and the lowest species density recorded in areas of dense rush or sedge, e.g. areas dominated by Empodisma minus, in Carne Central and West Wolgan (Fig. 1).
Species Abundance
All common species (mean cover >15 % in the 4 m2 plots) were recorded in the 400 m2 plots (Fig. 4). In comparison, the 400 m2 plots missed 232 sparse species (mean cover <15 % in the 4 m2 plots) found in the 4 m2 plots across all four wetlands. Species with higher Braun-Blanquet cover classes (5, 6 and 7) in the 400 m2 plots tended to have higher percentage cover across the wetland. For Braun-Blanquet cover classes 1 to 4 there was little correspondence between percentage cover across the wetland recorded in 4 m2 plots and Braun-Blanquet scores recorded in 400 m2 plots (Fig. 4).

Species Braun-Blanquet cover class in the 400 m2 plots across all wetlands versus mean percentage cover estimate in 4 m2 plots for that wetland; NA = not found in a 400 m2 plot, dashed boxes show the range for each cover class. Number of species in each cover class: NA =232; 1 = 23; 2 = 37; 3 = 38; 4 = 17; 5 = 9; 6 = 2; 7 = 1
Discussion
Here we examined a sampling method employed to monitor vegetation in Newnes Plateau Shrub Swamps to see whether it can achieve the stated objectives of detecting a change in species diversity, composition or abundance due to altered hydrology. We conducted detailed studies of four of the monitored wetlands to better understand whether the natural spatial variation across the wetlands is recorded by the established sampling method. These studies demonstrate that the established sampling method using 400 m2 plots does record both the common species and the life-forms present; though it lacks ability to detect statistically significant changes in vegetation over time at the wetland scale due to an absence of spatial replication within wetlands. Clearly the choices made in the design stage of a monitoring program will determine its effectiveness (Lindenmayer and Likens 2010). Sampling choices should depend on a) the reason for sampling, b) the predicted changes that might occur and c) the trigger values (i.e. the level of unacceptable change) set. Our study demonstrates a case where a lack of detail in the original objectives, subsequent changes in objectives, and introduction of trigger values defined as statistically significant changes in the indicator variables, has led to an inadequate sampling design. Below we discuss how the monitoring objectives and design could be improved based on our current knowledge of the wetland system.
It is well known that as area sampled increases, so too does the number of species that will be recorded. The spatial distributions of species, along with the size (grain) and distribution of plots (extent) all influence the species accumulation curve (Hurlbert 1990; Scheiner et al. 2011). Though the species accumulation curves presented here do not reach an asymptote, they provide the lower bound estimates of species richness for these four wetlands (Gotelli and Colwell 2001). The species accumulation curves clearly illustrate that a small number of 400 m2 purposively-located plots capture less than a third of species present in these heterogeneous wetlands. This may be because most plots are located in central, wetter, areas of the wetland which also tend to be less diverse. Most of the species missed by 400 m2 plots were sparse species. This bias towards the more abundant species is a common problem with plot-based methods (Gaston 1999). The implications here are that the current sampling method of a small number of 400 m2 plots will likely miss changes in the widely dispersed sparse species and the uncommon clumped species, such as herbaceous weeds, which are listed as one of the indicator groups (Blick et al. 2013).
In the current sampling design it is evident that replication is not related to the geographic extent of these wetland plant communities. For example, the current sampling design has one plot in Bungleboori North (area = 0.5 ha) and one plot Carne Central (area = 5 ha) which is an order of magnitude difference in area. The haphazard nature of the current sampling design is exemplified in West Wolgan (area = 5 ha) which has six plots, instead of one. The difference in sampling intensity between Carne Central (reference wetland, not undermined) and West Wolgan (undermined) is largely determined by compliance and regulatory frameworks rather than biological or statistical reasons. Due to the different sizes of the wetlands, the 400 m2 plot represents a different proportion of each wetland and hence capture a different proportion of species in each wetland; making comparisons of changes between ‘impact’ and ‘control’ wetlands difficult. As we found a greater variance in species density within larger wetlands (perhaps indicative of a wider range of habitats present), we recommend that the current monitoring design could be improved by distributing sampling effort across the wetland and be scalable with wetland size.
There are many ways the current sampling design could be made more effective. One such method is the stratified random method used here to characterise the wetlands but small randomly distributed plots are difficult to relocate accurately, potentially leading to reduced precision (i.e. repeatability) of cover estimates; in addition they are inefficient to sample due to the amount of time required to locate a large number of individual plots. In order to capture the range of habitats present in each wetland, it would be ideal to sample across the environmental gradients. Specifically across the hydrological gradient as the main impact from undermining is predicted to be changes in flow patterns of groundwater. Given that detailed hydrological information is not available for each wetland, nor are there clear hydrological zones present in all wetlands from which to sample [some wetlands range from dry to damp while others range from dry to standing surface water (Brownstein et al. unpublished data], transects could be an effective method of capturing the variation present in a wetland. We suggest a method that would use replicate transects in each wetland that would be more statistically robust in detecting ecologically relevant changes. The proposed method would use a sub-sampling regime with smaller plots or point intercept samples along transects spanning the width of the wetland (i.e. each transect spans the whole soil moisture/water depth gradient in each wetland), with the number of replicate transects scaled to wetland size, addressing the main sampling issues. Smaller plots within the transects would have the advantage that they can be searched quickly and accurately (Klimes et al. 2001). In addition placing small sub-plots along transects allows plots to be relocated with greater speed and accuracy, compared to relocating individual plots randomly distributed across the wetland (such as the sampling method used here to characterise the wetlands). A second, more novel approach, would be to use remote sensing (Chabot and Bird 2013). Aerial imagery with a spatial resolution of 0.15 m or less would provide detailed coverage over the whole of the wetland and allow quantification of factors such as vegetation die-back and changes in canopy species composition, canopy height and extent of disturbed or bare ground. Using aerial imagery would overcome problems associated with on-the-ground plot based sampling such as: restricted spatial scales (Booth and Cox 2008; Baldeck et al. 2014) and disturbance to fragile vegetation (Fletcher and Erskine 2012). Either a transect based or remote sensing based method would be an improvement on the current 400 m2 plot based method as they would both be more effective for detecting changes in vegetation within a wetland.
Due to the lack of replication here (14 of the 26 wetlands in current monitoring program contain only a single plot) many simple and common statistics, e.g. means and confidence intervals, cannot be calculated for a wetland. In addition, the plot locations were chosen subjectively, which invalidates a basic assumption of many statistical tests. A statistically sound sampling design is required for testing hypotheses and determining whether triggers have been exceeded (Barrett and Gray 2011). In all four wetlands, the 400 m2 plots were located in areas of relatively low species density (Fig. 1); this may be due in part to an unintentional bias toward choosing areas that are easy to access, or an intentional bias towards choosing the wettest areas (which are also the least diverse). Removing any unintentional bias in sampling locations and sampling across equivalent extents in each wetland will improve this sampling design.
The lack of replication also raises an issue of plant cover estimates; as others have found poor repeatability (between workers or time points) for plant cover estimates in a single quadrat, regardless of quadrat size (Jalonen et al. 1998; Archaux et al. 2007). In addition, there was little correspondence between estimates of percentage cover in the 4 m2 plots, especially for the species in cover classes 1 to 4 in the 400 m2 plot (the vast majority of the species recorded). Using multiple plots within a wetland allows i) internal variability and ii) the precision of the estimates of species abundances to be assessed (Jennings et al. 2009). One way to increase spatial replication without spending additional money is to reduce the frequency of wetland monitoring. The current program requires seasonal monitoring, though given that many of the species present are long-lived, it is possible that impacts to vegetation will occur over years, not months. In this case, resources may be better spent sampling only once per year (instead of the current four times) and increasing the replication per wetland. Visiting the sites less often would also reduce trampling damage to the fragile wetland vegetation, which could lead to a cumulative impact over time. Results from other studies suggest that fragile communities such as subalpine button grass can sustain prolonged damage with as few as 100 passes (Whinam and Chilcott 2003). Ultimately the effect of track damage could increase species richness and alter species composition (Scott and Kirkpatrick 1994; Gremmen et al. 2003), decreasing the sensitivity and reliability of the un-replicated sampling design.
Resource and time limitations often provide the main constraints in the design and implementation of monitoring programs. The choice of which scale to measure is often arbitrary and based on funding or convenience rather than knowledge (Harris 2003). The 400 m2 plot with Braun-Banquet cover/abundance scores is quick to sample and is a standard sampling protocol used for vegetation mapping in New South Wales (see Schultz et al. 2014) but we would argue that it does not suit the requirements here. This large plot size is difficult to accurately search in dense shrubby wetland vegetation and due to the small size of some wetlands there is limited space for replicate plots. In addition the cover/abundance scale with uneven classes makes setting trigger values more complex, e.g. a change in cover/abundance score from 1 to 2 is potentially not the same as a change in cover/abundance score from 3 to 4. While the single 400 m2 plot design is an informative way to record both the common species and calculate the proportion of species across life-forms in a wetland; this type of design seems better suited for broad landscape-scale vegetation mapping as it not effective for detecting change over time in vegetation.
In order to sample most efficiently, a clear statement of monitoring objectives and trigger values (i.e. level of unacceptable change) for management action is needed. Now that more is known about Newnes Plateau shrub swamp wetland systems, it is time to re-evaluate the monitoring objectives and develop detailed objectives and trigger values accordingly. For example, the current objective of detecting ‘a change in species diversity and abundance’ should be made specific to reflect the expected impact of reduced water availability, e.g. trigger values should be set related to a reduction in hydrophyte abundance or increases in terrestrial species abundance within a wetland. Using plant functional groups rather than individual species can make it easier to interpret trends in abundance (Casanova 2011; Campbell et al. 2014). In addition, trigger values for total live vegetation cover and bare ground would help determine whether vegetation dieback is occurring within a wetland, which when combined with the hydrological data will make it possible to relate negative changes in vegetation to undermining. Once these detailed hypotheses and triggers have been established, a rigorous sampling design could be developed with the statistical power to detect unacceptable changes in wetland vegetation.
When implementing monitoring programs, it seems that the basic tenets of the scientific method and sampling design are often over looked. There are many examples of monitoring programs with inefficient or ineffectual sampling designs (e.g. Legg and Nagy 2006; Martin et al. 2007), and here we present yet another example. We join other authors (e.g. Downs et al. 2002; Reid and Quinn 2004), to reiterate that the definition of clear monitoring objectives is an often overlooked step that has the potential to save time, money and reduce environmental impacts. Effective monitoring requires periodic review of objectives and revision of methods to ensure that the most efficient method for detecting unacceptable changes is being utilised.
References
Archaux F, Bergès L, Chevalier R (2007) Are plant censuses carried out on small quadrats more reliable than on larger ones? Plant Ecology 188:179–190. doi:10.1007/s11258-006-9155-y
Baldeck CA, Colgan MS, Féret J-B et al (2014) Landscape-scale variation in plant community composition of an African savanna from airborne species mapping. Ecological Applications 24:84–93. doi:10.1890/13-0307.1
Barrett TM, Gray AN (2011) Potential of a national monitoring program for forests to assess change in high-latitude ecosystems. Biological Conservation 144:1285–1294. doi:10.1016/j.biocon.2010.10.015
Benson D, Baird IRC (2012) Vegetation, fauna and groundwater interrelations in low nutrient temperate montane peat swamps in the upper Blue Mountains, New South Wales. Cunninghamia 12:267–307. doi:10.7751/cunninghamia.2012.12.021
Beyer HL (2009) Geospatial modelling environment
Blick RAJ, Brownstein G, Johns C, et al (2013) Spring and annual flora monitoring report for Centennial Coal operations. 72
Booth DT, Cox SE (2008) Image-based monitoring to measure ecological change in rangeland. Frontiers in Ecology and the Environment 6:185–190. doi:10.1890/070095
Campbell CJ, Johns CV, Nielsen DL (2014) The value of plant functional groups in demonstrating and communicating vegetation responses to environmental flows. Freshw Biol In press. doi:10.1111/fwb.12309
Casanova MT (2011) Using water plant functional groups to investigate environmental water requirements. Freshwater Biology 56:2637–2652. doi:10.1111/j.1365-2427.2011.02680.x
Chabot D, Bird DM (2013) Small unmanned aircraft: precise and convenient new tools for surveying wetlands. Journal of Unmanned Vehicle Systems 01:15–24. doi:10.1139/juvs-2013-0014
Dodd M (2011) Where are my quadrats? Positional accuracy in fieldwork. Methods in Ecology and Evolution 2:576–584. doi:10.1111/j.2041-210X.2011.00118.x
Downs BJ, Barmuta LA, Fairweather PG et al (2002) Assessing ecological impact. Applications in flowing waters. Cambridge University Press, Cambridge
Fletcher AT, Erskine PD (2012) Mapping of a rare plant species (Boronia deanei) using hyper-resolution remote sensing and concurrent ground observation. Ecological Management and Restoration 13:195–198. doi:10.1111/j.1442-8903.2012.00649.x
Gaston KJ (1999) Implications of interspecific and intraspecific abundance-occupancy relationships. Oikos 86:195–207
Gitay H, Roxburgh SH, Wilson JB (1991) Species-area relations in a New Zealand tussock grassland, with implications for nature reserve design and for community structure. Journal of Vegetation Science 2:113–118. doi:10.2307/3235903
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4:379–391
Gremmen N, Smith V, van Tongeren O (2003) Impact of trampling on the vegetation of subantarctic Marion Island. Arctic Antarctic and Alpine Research 35:442–446. doi:10.1657/1523-0430(2003)035[0442:IOTOTV]2.0.CO;2
Harris GP (2003) Ecological paradigms, change detection and prediction. Integr Model Biophys Soc Econ Syst Resour Manag Solut
Hellmann JJ, Fowler GW (1999) Bias, precision, and accuracy of four measures of species richness. Ecological Applications 9:824–834
Hurlbert SH (1990) Spatial distribution of the Montane Unicorn. Oikos 58:257–271
Jalonen J, Vanha-majamaa I, Tonteri T (1998) Optimal sample and plot size for inventory of field and ground layer vegetation in a mature Myrtillus- type boreal spruce forest. Annales Botanici Fennici 35:191–196
Jennings MD, Faber-Langendoen D, Loucks OL et al (2009) Standards for associations and alliances of the U.S. National Vegetation Classification. Ecological Monographs 79:173–199. doi:10.1890/07-1804.1
Klimes L, Dancak M, Hajek M et al (2001) Scale-dependent biases in species counts in a grassland. Journal of Vegetation Science 12:699–704
Lee WG, McGlone M, Wright E (2005) Biodiversity inventory and monitoring: a review of national and international systems and a proposed framework for future biodiversity monitoring by the Department of Conservation. 213
Legg CJ, Nagy L (2006) Why most conservation monitoring is, but need not be, a waste of time. Journal of Environmental Management 78:194–199. doi:10.1016/j.jenvman.2005.04.016
Lindenmayer DB, Likens GE (2009) Adaptive monitoring: a new paradigm for long-term research and monitoring. Trends in Ecology & Evolution 24:482–486. doi:10.1016/j.tree.2009.03.005
Lindenmayer DB, Likens GE (2010) Effective ecological monitoring. CSIRO Publishing
Martin J, Kitchens WM, Hines JE (2007) Importance of well-designed monitoring programs for the conservation of endangered species: case study of the snail kite. Conservation Biology 21:472–481. doi:10.1111/j.1523-1739.2006.00613.x
Oksanen J, Blanchet FG, Kindt R, et al (2012) vegan: community ecology package. R package version 2.0-4
Palmer MW, White PS (1994) Scale dependence and the species-area relationship. American Naturalist 144:717–740
Philippi TE, Dixon P, Taylor BA (1998) Detecting trends in species composition. Ecological Applications 8:300–308
R Core Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ramsay PM, Kent M, Reid CL, Duckworth JC (2006) Taxonomic, morphological and structural surrogates for the rapid assessment of vegetation. Journal of Vegetation Science 17:747–754
Reid MA, Quinn GP (2004) Hydrologic regime and macrophyte assemblages in temporary floodplain wetlands: Implications for detecting responses to environmental water allocations. Wetlands 24:586–599. doi:10.1672/0277-5212(2004)024[0586:HRAMAI]2.0.CO;2
Scheiner SM, Chiarucci A, Fox GA et al (2011) The inderpinning of the relationship of species richness with time and space. Ecological Monographs 81:195–213
Schultz NL, Reid N, Lodge G, Hunter JT (2014) Seasonal and interannual variation in vegetation composition: Implications for survey design and data interpretation. Austral Ecol n/a–n/a. doi:10.1111/aec.12141
Scott JJ, Kirkpatrick JB (1994) Effects of human trampling on the sub-Antarctic vegetation of Macquarie Island. Polar Record 30:207–220. doi:10.1017/S003224740002427X
Stevens CJ, Dise NB, Mountford JO, Gowing DJ (2004) Impact of nitrogen deposition on the species richness of grasslands. Science 303:1876–1879. doi:10.1126/science.1094678
Tongway DJ, Hindley NL (2004) Landscape function analysis manual: procedures for monitoring and assessing landscapes with special reference to minesites and rangelands, version 3.1. CSIRO Sustainable Ecosystems, Canberra
Whinam J, Chilcott NM (2003) Impacts after four years of experimental trampling on alpine/sub-alpine environments in western Tasmania. Journal of Environmental Management 67:339–351. doi:10.1016/S0301-4797(02)00218-9
Acknowledgments
Thank you to Cameron Kilgour and Vanessa Glenn for their assistance with fieldwork and plant identification. This work was supported by funds from Centennial Coal Ltd.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Brownstein, G., Blick, R., Johns, C. et al. Optimising a Sampling Design for Endangered Wetland Plant Communities: Another Call for Adaptive Management in Monitoring. Wetlands 35, 105–113 (2015). https://doi.org/10.1007/s13157-014-0599-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-014-0599-x