
Abstract
Early brain injury (EBI), delayed cerebral vasospasm (DCVS), and delayed cerebral ischemia (DCI) are common complications of subarachnoid hemorrhage (SAH). Inflammatory processes in the cerebrospinal fluid (CSF) are one of the causes for such complications. Our aim to study the effects of an IL-6 receptor antagonist (Tocilizumab) examines the occurrence of DCVS, neuronal cell death, and microclot formation in an acute SAH rabbit model. Twenty-nine New Zealand white rabbits were randomized into one of three groups as the SAH, SAH + Tocilizumab, and sham groups. In SAH groups, hemorrhage was induced by extracranial-intracranial arterial blood shunting from the subclavian artery into the cisterna magna under intracranial pressure (ICP) monitoring. In the second group, Tocilizumab was given once intravenously 1 h after SAH induction. Digital subtraction angiography was performed, and CSF and blood were sampled before and after (day 3) SAH induction. IL-6 plasma and CSF levels were measured. TUNEL, FJB, NeuN, and caspase-3 immunostaining were used to assess cell apoptosis, neurodegeneration, and neuronal cell death, respectively. Microclot formation was detected by fibrinogen immunostaining. Between baseline and follow-up, there was a significant reduction of angiographic DCVS (p < 0.0001) in the Tocilizumab compared with the SAH group. Tocilizumab treatment resulted in decreased neuronal cell death in the hippocampus (p = 0.006), basal cortex (p = 0.001), and decreased microclot formation (p = 0.02). Tocilizumab reduced DCVS, neuronal cell death, and microclot formation in a rabbit SAH model, and could be a potential treatment to prevent DCVS and DCI in SAH patients.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Despite recent advances in the treatment of subarachnoid hemorrhage (SAH), neurological outcomes after SAH remain poor. Early brain injury (EBI), delayed cerebral vasospasm (DCVS), and delayed cerebral ischemia (DCI) are common complications of SAH along with high morbidity and mortality [1,2,3]. Compartmental inflammatory response in the subarachnoid space is seemingly involved in the pathogenesis of DCVS and neuronal injury [1, 4,5,6]. In this context, interleukin (IL)-6, which has been shown to play a key role in the inflammation cascade after experimental and clinical SAH, has been related to the development of DCVS and worse clinical outcomes [5,6,7]. We recently reported a positive correlation between IL-6 and the vasoconstrictor endothelin 1 (ET-1) and DCVS induction after SAH [5]. Other studies have described elevated IL-6 to activate the coagulation pathway and vascular endothelial cells in various clinical conditions [8], thus possibly initiating microclot formation. Therefore, the presence of microcirculatory dysfunction is a suspected key factor of early brain injury after SAH [9,10,11]. On the basis of the reported preclinical, experimental, and clinical relevance of IL-6 in the setting of DCVS and neuronal injury after SAH, our study aims to analyze the effectiveness of the IL-6 receptor antagonist Tocilizumab in diminishing DCVS, neuronal injury, and microclot formation in an acute hemorrhage rabbit model.
Material and Methods
Animals, Study Design, and Anesthesia
The authors declare that all supporting data are available within the article and its online resource files, including the detailed protocols for anesthesia, surgery, and angiography previously described [5, 12,13,14,15,16]. A total of 29 female New Zealand white rabbits (3.3–4.2 kg body weight, Charles River, Sulzfeld, Germany) were randomly allocated to one of three groups as the SAH (n = 12), SAH + Tocilizumab (n = 12), or sham (n = 5) groups. Due to animal welfare regulations, the study was performed with adult female rabbits. General anesthesia was induced in all rabbits under continuous monitoring of vital parameters. Neurological status was assessed at 6, 12, 24, 48, and 72 h post-SAH using a four-point grading system previously described (Online Resource Fig. 1) [17].

Flowchart of the study design. Three animals were prematurely excluded (one per group) because of death during anesthesia induction (a). Two rabbits (one in the SAH and SAH + Tocilizumab group each) died during SAH induction, and a third rabbit, from the SAH + Tocilizumab group, was prematurely euthanized because of a severe postoperative neurological deficit after SAH induction (b). Illustration demonstrates the experimental set-up of subarachnoid hemorrhage induction previously described by Marbacher et al. (c) [16]. The extra-/intracranial blood shunt allows blood release under arterial pressure into the subarachnoid [16]. BL, baseline; FU, follow-up; CSF, cerebrospinal fluid; ICP, intracranial pressure; iv, intravenous
Digital Subtraction Angiography, SAH Induction, and Tocilizumab Infusion
Digital subtraction angiography (DSA) was performed on day 0 before SAH and at follow-up on day 3 (Fig. 1). The rabbit subclavian artery was dissected and cannulated with a 5.5-french catheter (Silicone Catheter STH-C040, Connectors Verbindungstechnik AG, Switzerland). The catheter tip was advanced to the origin of the vertebral artery followed by an intra-arterial bolus injection of contrast dye (0.6 ml/kg Iopamidol, Iopamiro, Bracoo Suisse, Switzerland). The diameter of the basilar artery (BA) was measured as previously described in a blinded manner using ImageJ (National Institutes of Health, Bethesda, MD, USA) [5, 12, 18,19,20]. Extent of DCVS was determined by comparing the diameters at baseline and follow-up. After baseline DSA on day 0, either induction of SAH or sham procedure was performed as described earlier [5, 12,13,14,15,16, 20].
In all animals, intracranial pressure (ICP), arterial blood pressure, and respiration rate (RR) were continuously monitored [18, 21]. Arterial blood gas analyses (ABGA) were performed prior to angiography. In prone position, a 27-G spinal access needle was inserted into the cisterna magna. In SAH animals, the needle was connected to the catheterized subclavian artery to induce a hemorrhage [5, 12,13,14,15,16]. In sham-operated animals, puncture of the cisterna magna, CSF sampling (1.5 ml), and CSF replacement with 1.5 ml artificial CSF (ACSF, Tocris Bioscience, Bristol, UK) were used. Directly after SAH induction in the postoperative phase, rabbits in the SAH + Tocilizumab group received Tocilizumab intravenously (8 mg/kg body weight) for a 1-h span.
Blood and CSF Sample Analysis
CSF and serum were sampled on days 0 and 3. Blood samples were collected using EDTA-coated tubes. For IL-6 and ET-1 measurements, a specific rabbit IL-6 and ET-1 enzyme-linked immunosorbent assay (ELISA) kit was used (Rabbit IL-6 ELISA Kit, CSB-E06903Rb and Rabbit ET-1 ELISA Kit, CSB-E06951Rb Cusabio Biotech Co. Ltd., USA). Euthanasia was induced on day 3.
Tissue Processing, Histology, and Histochemistry
After the brains were removed, the volume of blood in the subarachnoid space was analyzed with the bleeding sum score, a 4-point grading system: 0, no blood; 1, minimal blood; 2, moderate blood clot (basilar artery visible); 3, massive blood clot (basilar artery not visible) [14]. Terminal apoptosis identified by DNA fragmentation was assessed with TUNEL staining (In Situ Cell Death Detection Kit, Roche, Switzerland). Earlier stages of apoptosis, characterized by caspase-3 activation, were identified with an anti-cleaved caspase-3 antibody staining (Cell Signaling, USA). For neurodegeneration, Fluoro Jade B (Millipore AG, Zug, Switzerland) staining was used. To facilitate the identification of neurons, an anti-neuronal nuclei staining (NeuN, Millipore, Switzerland) was performed. Cerebellar Purkinje cells were stained with antibodies against calbindin (Swant, Marly, Switzerland). Quantitative analysis of apoptosis in the hippocampus (CA1 and CA3), basal cortex, and cerebellum was performed in a blinded way within predefined regions of interest (ROI). Extent of microthrombosis was detected by fibrinogen immunohistochemistry using the Leica Bond III IHC staining system and analyzed in a blinded manner as previously described [10].
Statistical Analysis
Data were analyzed and visualized using GraphPad Prism statistical software Version 8.0 (GraphPad Software, Inc. California, USA). Continuous values were given as mean ± standard deviation (SD) if not otherwise indicated. Non-parametric tests were used given the relatively small sample size; baseline and follow-up values were compared using the Wilcoxon signed rank test (non-parametric, repeated measures). Differences between the normally distributed data of two groups were analyzed by Student’s t test, Kruskal-Wallis test (non-parametric ANOVA) among three or more groups, and the Scheffé post hoc analysis. A p value < 0.05 was regarded as statistically significant.
Ethic Approval
The project has been performed according to the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines [22] and was performed in accordance with the National Institutes of Health Guidelines for the care and use of experimental animals and with the approval of the Animal Care Committee of the Canton Bern, Switzerland (Approval Nr. BE58/17).
Results
Physiological Parameters, ICP and CPP Time Course, SAH, and Neuronal Scores
Among 29 rabbits, 23 rabbits reached the primary endpoint, 10 in the SAH, 9 in the SAH + Tocilizumab, and 4 in the sham groups (Fig. 1). All animals in the SAH and SAH + Tocilizumab groups showed significantly elevated ICPs during SAH induction and corresponding significantly decreased CPPs from baseline to peak (Online Resource Fig.1Panel A).
All rabbits in the SAH and SAH + Tocilizumab groups demonstrated extensive, coagulated, diffuse subarachnoid blood (Online Resource Fig.2) that resulted in moderate grades of SAH. Neurological scores (Online Resource Fig.1, Panel B) at 6, 12, 24, 48, and 72 h after the procedure were comparable among groups.
Angiographic Delayed Cerebrovascular Vasospasm
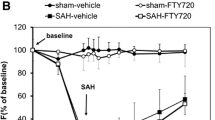
On angiographic findings, BA diameter averaged 310 ± 50 μm at baseline and decreased to 230 ± 60 μm at day 3 in the SAH group (p < 0.00001) (Fig. 2). BA diameter averaged 320 ± 60 μm at baseline and decreased to 300 ± 50 μm (p = 0.16) at follow-up in the SAH + Tocilizumab group; the occurrence of DCVS was significantly decreased in this group compared with the SAH group (p < 0.00001).

a DSA at baseline (BA) and at day 3 follow-up (FU). White arrows: point of measurement of basilar artery diameter. Note decreased size of the basilar artery at FU. b Angiographic mean basilar artery diameter in μm (y axis) at baseline and follow-up among the different groups. Angiography in the SAH group showed a significant (*) decrease of diameter at FU compared to BL. *p < 0.00001
Brain Histology
In the SAH group, amounts of TUNEL/Hoechst double-positive cells in the CA1 and CA3 regions of the hippocampus numbered 56 ± 19 and 58 ± 22.3 cells, respectively (Fig. 3). In the SAH + Tocilizumab group, there were significantly fewer TUNEL/Hoechst-positive cells at 6.4 ± 3 cells in the CA1 (p = 0.006) and 11.6 ± 5.5 cells (p = 0.002) in the CA3 regions of the hippocampus (Online Resource Fig.3).

a Representative images of apoptotic double-positive TUNEL/Hoechst, FJB/Hoechst, and caspase-3/Hoechst cells (arrow heads) can be seen in the hippocampus of animals. b Apoptosis and neurodegeneration significantly decreased in the SAH + Tocilizumab group compared to SAH. Data are presented as mean ± SD: *p < 0.05. Scale bar = 100 μm
The FJB-staining cells in the CA1 and CA3 regions of hippocampus numbered 189.6 ± 46.6 and 214.2 ± 48, respectively, in the SAH group (Online Resource Fig.4) but were significantly fewer at 73.2 ± 18.8 (p = 0.002) and 92.2 ± 20.7 (p = 0.001), respectively, in the SAH + Tocilizumab group.
Caspase-3-positive cells in the CA1 and CA3 regions of hippocampus (Online Resource Fig.5) numbered 20.1 ± 8.4 and 28.6 ± 14.7, respectively, in the SAH group compared with 16.8 ± 4.8 and 21.6 ± 6.5 (p = 0.2), respectively, in the SAH + Tocilizumab group. In the basal cortex, caspase-3-positive cells were 66.2 ± 53.5 in the SAH group and fewer at 15 ± 5.3 (p = 0.06) in the SAH + Tocilizumab group.
NeuN-positive staining cells in the CA1 and CA3 regions of the hippocampus numbered 27.2 ± 10.2 and 45.6 ± 17.6, respectively, in the SAH group (Fig. 4) and were significantly higher at 197.4 ± 34.8 (p < 0.00001) and 174.4 ± 33.2 (p = 0.0005), respectively, in the SAH + Tocilizumab group.

a Cumulative number of NeuN-positive cells shown as bar graph (mean ± SD, *p < 0.05. b NeuN-positive staining neurons (open arrows) seen in higher amounts in the hippocampus and basal cortex of animals in the SAH + Tocilizumab and sham groups compared to NeuN negative neurons (arrowhead) in the SAH group. Scale bar = 100 μm
Calbindin-positive cells (Online Resource Fig.6), staining Purkinje cells in the cerebellum, were 2.9 ± 1.7 in the SAH group and significantly increased to 15.5 ± 9.3 in the SAH + Tocilizumab group (p = 0.0008). FJB-positive cells (Online Resource Fig.7) were 4.7 ± 1.5 in the SAH group compared to 2 ± 1.2 in the SAH + Tocilizumab group (p = 0.01). TUNEL/Hoechst staining-positive cells (Online Resource Fig.8) numbered 19.3 ± 14.2 in the SAH group compared to 6 ± 2.2 in the SAH + Tocilizumab group (p = 0.059).
TUNEL/Hoechst staining-positive cells (endothelial and smooth muscle cells) in the arterial wall of the basilar artery numbered 48.5 ± 41.8 in the SAH group compared to 8.5 ± 13.3 in the SAH + Tocilizumab group (p = 0.008, Online Resource Fig.9). Caspase-positive cells numbered 18.8 ± 10.5 in the SAH group and were significantly fewer at 3.7 ± 2.4 in the SAH + Tocilizumab group (p = 0.0004).
Microclot Formation
Fibrinogen staining showed distinct microclot formation in vessels of the hippocampus in the SAH group compared with the sham and SAH + Tocilizumab groups (Fig. 5). In the hippocampus CA1 region, microclot formation was significantly higher in the SAH than the SAH + Tocilizumab groups (6.8 ± 2.3 vs. 2.4 ± 1.3, respectively), (p = 0.02). In the CA3 region, there were 4 ± 1.8 microclots in the SAH group and 1.7 ± 1.1 in the SAH + Tocilizumab group (p = 0.52). In the basal cortex, there were 3.4 ± 1.3 microclots in the SAH group and 3.2 ± 1.5 in the SAH + Tocilizumab group (p = 0.7).

a Cumulative number of fibrinogen-positive microclots shown as bar graph presented as mean ± SD, *p < 0.05. b Significantly more fibrinogen-positive staining microclots (arrowheads) formed in the hippocampus and basal cortex in the SAH group than in the SAH + Tocilizumab and sham groups. Scale bar = 100 μm
ELISA IL-6 and ET-1 Quantification
CSF IL-6 levels increased from baseline to follow-up from 28.6 ± 21.4 pg/ml to 38.8 ± 17.9 pg/ml (p = 0.34) in the SAH group and from 29.9 ± 17.7 pg/ml to 58.9 ± 53.6 (p = 0.15) in the SAH + Tocilizumab group. At day 3, CSF IL-6 values were non-significantly higher in the SAH + Tocilizumab group. Overall, IL-6 values were significantly higher in the CSF than in the plasma (43.73 ± 33.1 pg/ml vs 0.31 ± 0.05 pg/ml, p < 0.00001), (Fig. 6).

a, b Mean baseline and follow-up IL-6 CSF and serum levels in pcg/ml. c, d Mean baseline and follow-up ET-1 CSF and serum levels in pcg/ml. Data are presented as mean ± SD, *p < 0.05
ET-1 CSF levels at baseline and at day 3 were 3.69 ± 1.83 pg/ml and 4.566 ± 2.33 pg/ml (p = 0.22), respectively, in the SAH group and 5.9 ± 1.9 pg/ml and 5.59 ± 1.93 pg/ml (p = 0.13), respectively, in the SAH + Tocilizumab group. Overall, ET-1 values were significantly higher at 5.6 ± 5.8 pg/ml in CSF compared to 1.0 ± 0.2 pg/ml in plasma (p < 0.00001). At day 3, levels of plasma ET-1 were significantly lower in the SAH + Tocilizumab group than either the SAH group (p = 0.0006) or sham group (p = 0.003).
Discussion
In this closed-cranium rabbit model, interleukin (IL)-6 receptor antagonist (Tocilizumab) prevented macro- and microcirculatory disturbances and was acutely neuroprotective after induced SAH. Additionally, Tocilizumab prevented smooth muscle and endothelial cell death in large cerebral arteries and diminished angiographic delayed cerebral vasospasm. On a microcirculatory level, Tocilizumab reduced microclot formation. Notably, Tocilizumab treatment resulted in neuroprotection by means of reduced neuronal cell death and neurodegeneration not only in regions more susceptible to ischemia (hippocampus and cerebellum) but also in the basal cortex.
Concordant with previous reports using the SAH rabbit model, we found a significant presence of DCVS in the SAH group [5, 12–16]. Interestingly, the Tocilizumab-treated rabbits presented a significant decrease of DCVS at follow-up compared with the untreated group. This might be explained by the increasing relevant role of the inflammation cascade, especially IL-6, in the induction of DCVS. Osuka et al. previously described that application of only IL-6 in the CSF of dogs caused DCVS [23]. Similar results were reported by our research group in which IL-6 application in the CSF of rabbits led to DCVS and significantly increased ET-1, which is a potent vasoconstrictor [5]. Moreover, Bowman et al. showed that the administration of a polyclonal antibody against IL-6 reduced vasospasm in a rat femoral artery vasospasm model [24]. Although IL-6 seems unlikely to directly induce DCVS, it more likely works indirectly as an important mediator by stimulating CSF-monocytes activation that in turn secrete ET-1, or indirectly with its role in impairing the cerebral autoregulation [6, 24, 25].
Correlating to this finding, our histological analysis showed a decrease of apoptotic cells in the endothelial and smooth muscle cell layers of the BA in the animals treated with Tocilizumab. Apoptotic damage to the endothelium and smooth muscle cells is known to be critical after SAH, especially in obstructing physiologic vasoregulation and breaking down the blood brain barrier (BBB), and has been reported in previous animal studies [26, 27]. Therefore, the reduction of cell apoptosis observed in the BA of the treatment group animals may have also contributed to the reduction of DCVS because the vessel wall was more intact.
Through the competitive inhibitory effect of Tocilizumab on the IL-6 receptor, the SAH + Tocilizumab group showed significant decreases in neuronal cell death and neurodegenerative cells in the CA1 and CA3 region of the hippocampus, cerebellum, and basal cortex. IL-6 acting as an intermediate induces the aggregation of inflammatory cells in a lesion area, increases the release of oxygen-free radicals from neutrophils, and collaborating with TNF-α promotes neuronal cell apoptosis through calcium overload in the cells [28]. Additionally, the impairment of cerebral autoregulation and DCVS related to IL-6 release may lead to hypoxia and neuronal cell death, which was prevented in the SAH + Tocilizumab group. In our previous study using the same model, we also noted that CSF application of IL-6 led to neuronal cell death [5].
The fact that Tocilizumab reduced neuronal cell death, especially in the hippocampus and cerebellum, is highly relevant from a clinical point of view. Specifically, the hippocampus and cerebellum are well known to be highly sensitive to neuro-inflammatory and hypoxic changes [29]. In clinical studies, it is widely accepted that damage to the hippocampus and cerebellum is implicated in loss of high cognitive functions, such as memory and learning. In fact, due to the global cerebral damage that occurs after SAH, even patients with supposed good outcomes (modified Rankin scale between 0 and 2) have severe cognitive impairment, with two thirds of the patients unable to return to work [30].
Differences in the time course and location of DCVS and DCI have called into question the cause and effect relationship: that is, clinically not all patients who develop DCVS have DCI, and not all patients with DCI have DCVS [10, 11, 31, 32]. Microclot formation in the brains of SAH patients found on autopsy studies has shown a correlation with location and severity of ischemia. Eventually, they were considered as possible contributing factor for DCI because of microvessel occlusion that leads to neuronal ischemia, degeneration, and apoptosis [10, 11, 31, 32]. Experimental studies with SAH models in mice and rabbits have also described microclot formations [10, 33]. Moreover, the application of ADAMTS-13, which cleaves the von Willebrand factor (vWF), resulted in fewer microclots, reduced apoptotic and degenerative neurons, and therefore achieved improvement of neurologic function in mice [34]. In our study, microclots formed in relevant amounts, especially in the SAH group. Interestingly, microclots were found in both hemispheres, which might have been consistent with the global insult and inflammation reaction after SAH.
Among other various factors from endothelial damage and clotting disorders known to lead to microclot formation, inflammation is particularly regarded as a possible mechanism [35]. IL-6 has been described to have prothrombogenic actions and to participate in the activation of the coagulation cascade [34,35,36]. Moreover, IL-6 together with IL-1 and TNF-α are known to induce tissue factor upregulation, which in turn promotes a procoagulant state among endothelial cells [8]. In acute coronary syndrome, IL-6 has been associated with endothelial dysfunction and enhanced expression of markers of endothelial cell activation, such as cellular adhesion molecules and thrombogenic vWF [37].
Bernardo et al. described that inhibition of the cleavage of ultralarge vWF by IL-6 resulted in platelet aggregation and adhesion of the vascular endothelium, thus possibly causing thrombosis in the microvessels [38]. Vergouwen et al. suggested that endothelium dysfunction and the hemostatic system are involved in the development of DCI; they noted lower amounts of ADAMTS-13 and higher levels of vWF antigen and propeptide in patients who suffered DCI after SAH. However, in contrast with the results of Bernardo et al., no clear correlation with IL-6 could be found [39].
Our findings of reduction in microclot formation in the SAH + Tocilizumab group, especially in the hippocampus, might have been caused by the inhibitory effect of Tocilizumab. Therefore, this reduction might have directly or indirectly contributed to the lower amounts of neuronal cell death and ischemia-induced neurodegeneration in the treatment group because of the intact microvessels without microclots. Another important factor to consider, with an eye on future studies, is that the coagulation cascade in rabbits appears very similar to that seen in humans [40]. Therefore, these results might also be reflected in humans.
Overall, we found that IL-6 and ET-1 levels were much higher in CSF than plasma levels. This finding further confirmed the compartmentalization of the inflammatory response in CSF after SAH, similar to other previously reported experimental and clinical studies [5, 6], and the importance of inflammation in the development of DCVS and DCI. Despite that, ELISA measurement of IL-6 and ET-1 was somewhat inconclusive. Interestingly, in the SAH + Tocilizumab group, levels of CSF IL-6 at day 3 were higher than baseline, but not significantly so.
Study Limitations and Clinical Implications
This study is subject to the various limitations in the executions of such work. SAH was effectively induced in both groups. Concordant with our previous studies using this model [5, 12,13,14,15,16], ABGA parameters were stable during hemorrhage induction and, between baseline and peak, ICP significantly increased and CPP decreased. As known predominant factors in the pathophysiology of SAH, changes in ICP and CPP are relevant in the abnormal autoregulation after intracranial circulatory arrest, which leads to rebound hyperemia and early global cerebral edema [41, 42]. From a clinical point of view, loss of consciousness due to acute SAH, which may reflect ictal cerebral circulatory arrest, has been described as a risk factor for global cerebral edema and as a predictor of death and poor functional outcome in SAH patients [41, 42]. Eventually, the model used reflected the pathophysiology of SAH in humans and seemed appropriate for the research question [43].
Tocilizumab is a humanized monoclonal antibody with a heavy molar mass of 145.0 kg/mol. Therefore, the question arises if it went through the blood brain barrier. Although we did not directly measure Tocilizumab in the CSF or brain, we measured the amounts of IL-6 in the CSF and plasma. As described above, we found higher amounts of IL-6 in CSF at day 3 in the SAH + Tocilizumab group than baseline, which seems an indirect indication that Tocilizumab (a competitive antagonist of the IL-6 receptor) went through the blood brain barrier. Additionally, several case reports demonstrated the successful use of Tocilizumab in the treatment of central nervous system diseases (therefore passing the blood brain barrier), such as neuromyelitis optica [44, 45]. Moreover, several studies have demonstrated the disruption of this barrier after SAH [46, 47], eventually facilitating the passage of Tocilizumab. However, no clear conclusion of the presence of Tocilizumab in CSF and parenchyma could be drawn. Therefore, further studies should aim to better clarify this issue and especially quantify the amount of Tocilizumab in the CSF and parenchyma to confirm the results described in this study. Tocilizumab was given immediately after SAH induction without knowing the therapeutic window, in addition the dosage of Tocilizumab was weight adapted according to the standard use of this medication in the clinical setting. Furthermore, a specific rabbit-related dose-finding study could not be performed because of the lack of specific studies and our lack of knowledge if rabbits respond differently with high or low dosages. Outcomes such as neurological scores and histological evaluation were measured only until 3 days after SAH. Considering that Tocilizumab is a relatively long-lasting drug, results might lead to different results at longer follow-up. Despite performing a power analysis, the n of the study is still low, especially for evaluation of CSF and plasma IL-6 and ET-1 levels. Other inflammatory cytokines were not measured, and ELISA quantitative analysis of IL-6 and ET-1 was only performed at day 0 and day 3. Possibly, frequent regular measurements during follow-up might have given different results.
Conclusions
The inhibition of the inflammation cascade with the IL-6 receptor antagonist Tocilizumab reduced DCVS, neuronal cell death, and microclot formation in a well-established rabbit acute SAH model. Considering the wide use of Tocilizumab in the treatment of rheumatoid arthritis, the results of this study might easily open potential new doors for future clinical studies to evaluate such treatment in the prevention of DCVS and neuroprotection in patients affected by SAH. Future animal studies should aim to confirm our results, especially analyzing the dose-response relationship of Tocilizumab.
Data Availability
The authors declare that all supporting data are available within the article and its online supplementary files, including the detailed protocols for anesthesia, surgery, and angiography previously described [5, 12–16].
Abbreviations
- ABGA:
-
Arterial blood gas analysis
- BA:
-
Basilar artery
- BBB:
-
Blood brain barrier
- CPP:
-
Cerebral perfusion pressure
- CSF:
-
Cerebrospinal fluid
- DCI:
-
Delayed cerebral ischemia
- DCVS:
-
Delayed cerebral vasospasm
- DSA:
-
Digital subtraction angiography
- EBI:
-
Early brain injury
- ELISA:
-
Enzyme-linked immunosorbent assay
- ET-1:
-
Endothelin-1
- FJB:
-
Fluoro Jade B
- HE:
-
Hematoxylin and eosin
- ICP:
-
Intracranial pressure
- IL:
-
Interleukin
- MAP:
-
Mean arterial pressure
- ROI:
-
Regions of interest
- RR:
-
Respiratory rate
- SAH:
-
Subarachnoid hemorrhage
- SD:
-
Standard deviation
- vWF:
-
von Willebrand factor
References
Niwa A, Osuka K, Nakura T, Matsuo N, Watabe T, Takayasu M. Interleukin-6, MCP-1, IP-10, and MIG are sequentially expressed in cerebrospinal fluid after subarachnoid hemorrhage. Journal of Neuroinflammation. Journal of Neuroinflammation; 2017;:1–6.
Kassell NF, Sasaki T, Colohan AR, Nazar G. Cerebral vasospasm following aneurysmal subarachnoid hemorrhage. Stroke. 1985;16:562–72.
Konczalla J, Kashefiolasl S, Brawanski N, Bruder M, Gessler F, Senft C, et al. Cerebral vasospasm-dependent and cerebral vasospasm-independent cerebral infarctions predict outcome after nonaneurysmal subarachnoid hemorrhage: a single-center series with 250 patients. World Neurosurg. 2017;106:861–4.
Al-Tamimi YZ, Bhargava D, Orsi NM, Teraifi A, Cummings M, Ekbote UV, et al. Compartmentalisation of the inflammatory response following aneurysmal subarachnoid haemorrhage. Cytokine. 2019;123:154778.
Croci D, Nevzati E, Danura H, Schöpf S, Fandino J, Marbacher S, et al. The relationship between IL-6, ET-1 and cerebral vasospasm, in experimental rabbit subarachnoid hemorrhage. J Neurosurg Sci. 2019;63(3):245–50.
Fassbender K, Hodapp B, Rossol S, Bertsch T, Schmeck J, Schütt S, et al. Inflammatory cytokines in subarachnoid haemorrhage: association with abnormal blood flow velocities in basal cerebral arteries. J Neurol Neurosurg Psychiatry BMJ Publishing Group. 2001;70:534–7.
Muroi C, Seule M, Sikorski C, Dent W, Keller E. Systemic interleukin-6 levels reflect illness course and prognosis of patients with spontaneous nonaneurysmal subarachnoid hemorrhage. Acta Neurochir Suppl Vienna: Springer Vienna. 2013;115:77–80.
Grignani G, Maiolo A. Cytokines and hemostasis. Haematologica. 2000;85:967–72.
Sehba FA, Hou J, Pluta RM, Zhang JH. The importance of early brain injury after subarachnoid hemorrhage. Prog Neurobiol. 2012;97:14–37.
Andereggen L, Neuschmelting V, Gunten von M, Widmer HR, Fandino J, Marbacher S. The role of microclot formation in an acute subarachnoid hemorrhage model in the rabbit. Biomed Res Int Hindawi. 2014;2014:161702–10.
Stein SC, Browne KD, Chen X-H, Smith DH, Graham DI. Thromboembolism and delayed cerebral ischemia after subarachnoid hemorrhage: an autopsy study. Neurosurgery. 2006;59:781–7 –discussion787–8.
Andereggen L, Neuschmelting V, Gunten von M, Widmer HR, Takala J, Jakob SM, et al. The rabbit blood-shunt model for the study of acute and late sequelae of subarachnoid hemorrhage: technical aspects. J Vis Exp. 2014;:e52132.
Marbacher S, Nevzati E, Croci D, Erhardt S, Muroi C, Jakob SM, et al. The rabbit shunt model of subarachnoid haemorrhage. Transl Stroke Res. Springer US. 2014;5:669–80.
Marbacher S, Neuschmelting V, Andereggen L, Widmer HR, Gunten von M, Takala J, et al. Early brain injury linearly correlates with reduction in cerebral perfusion pressure during the hyperacute phase of subarachnoid hemorrhage. Intensive Care Med Exp. Springer International Publishing; 2014;2:30.
Marbacher S, Fathi A-R, Muroi C, Coluccia D, Andereggen L, Neuschmelting V, et al. The rabbit blood shunt subarachnoid haemorrhage model. Acta Neurochir Suppl Cham: Springer International Publishing. 2015;120:337–42.
Marbacher S, Sherif C, Neuschmelting V, Schläppi J-A, Takala J, Jakob SM, et al. Extra-intracranial blood shunt mimicking aneurysm rupture: intracranial-pressure-controlled rabbit subarachnoid hemorrhage model. J Neurosci Methods. 2010;191:227–33.
Endo S, Branson PJ, Alksne JF. Experimental model of symptomatic vasospasm in rabbits. Stroke. 1988;19:1420–5.
Marbacher S, Andereggen L, Neuschmelting V, Widmer HR, Gunten von M, Takala J, et al. A new rabbit model for the study of early brain injury after subarachnoid hemorrhage. J Neurosci Methods. 2012;208:138–45.
Croci DM, Wanderer S, Strange F, Grüter BE, Casoni D, Sivanrupan S, et al. Systemic and CSF interleukin-1α expression in a rabbit closed cranium subarachnoid hemorrhage model: an exploratory study. Brain Sci Multidisciplinary Digital Publishing Institute. 2019;9:249.
Zhang Z-W, Yanamoto H, Nagata I, Miyamoto S, Nakajo Y, Xue J-H, et al. Platelet-derived growth factor-induced severe and chronic vasoconstriction of cerebral arteries: proposed growth factor explanation of cerebral vasospasm. Neurosurgery. 2010;66:728–35 –discussion735.
Marbacher S, Milavec H, Neuschmelting V, Andereggen L, Erhardt S, Fandino J. Outer skull landmark-based coordinates for measurement of cerebral blood flow and intracranial pressure in rabbits. J Neurosci Methods. 2011;201:322–6.
NC3Rs Reporting Guidelines Working Group. Animal research: reporting in vivo experiments: the ARRIVE guidelines. J. Physiol. (Lond.). Blackwell Publishing Ltd; 2010. pp. 2519–21.
Osuka K, Suzuki Y, Tanazawa T, Hattori K, Yamamoto N, Takayasu M, et al. Interleukin-6 and development of vasospasm after subarachnoid haemorrhage. Acta Neurochir. 1998;140:943–51.
Bowman G, Dixit S, Bonneau RH, Chinchilli VM, Cockroft KM, et al. Neurosurgery. 2004;54:719–25 –discussion725–6.
Fassbender K, Hodapp B, Rossol S, Bertsch T, Schmeck J, Schütt S, et al. Endothelin-1 in subarachnoid hemorrhage: an acute-phase reactant produced by cerebrospinal fluid leukocytes. Stroke. Lippincott Williams & Wilkins. 2000;31:2971–5.
Zhou C, Yamaguchi M, Kusaka G, Schonholz C, Nanda A, Zhang JH. Caspase inhibitors prevent endothelial apoptosis and cerebral vasospasm in dog model of experimental subarachnoid hemorrhage. J. Cereb. Blood Flow Metab. SAGE Publications Sage UK: London, England. 2004;24:419–31.
Zhou C, Yamaguchi M, Colohan ART, Zhang JH. Role of p53 and apoptosis in cerebral vasospasm after experimental subarachnoid hemorrhage. J. Cereb. Blood Flow Metab. SAGE Publications Sage UK: London, England. 2005;25:572–82.
Li S-J, Liu W, Wang J-L, Zhang Y, Zhao D-J, Wang T-J, et al. The role of TNF-α, IL-6, IL-10, and GDNF in neuronal apoptosis in neonatal rat with hypoxic-ischemic encephalopathy. Eur Rev Med Pharmacol Sci. 2014;18:905–9.
Ribeiro MC, Bezerra TDS, Soares AC, Boechat-Ramos R, Carneiro FP, Vianna LM d S, et al. Hippocampal and cerebellar histological changes and their behavioural repercussions caused by brain ischaemic hypoxia experimentally induced by sodium nitrite. Behav Brain Res. 2017;332:223–32.
Powell J, Kitchen N, Heslin J, Greenwood R. Psychosocial outcomes at 18 months after good neurological recovery from aneurysmal subarachnoid haemorrhage. J Neurol Neurosurg Psychiatry. 2004;75:1119–24.
Treggiari MM, Walder B, Suter PM, Romand J-A. Systematic review of the prevention of delayed ischemic neurological deficits with hypertension, hypervolemia, and hemodilution therapy following subarachnoid hemorrhage. J Neurosurg Journal of Neurosurgery Publishing Group. 2003;98:978–84.
Macdonald RL, Pluta RM, Zhang JH. Cerebral vasospasm after subarachnoid hemorrhage: the emerging revolution. Nat Clin Pract Neurol Nature Publishing Group. 2007;3:256–63.
Pisapia JM, Xu X, Kelly J, Yeung J, Carrion G, Tong H, et al. Microthrombosis after experimental subarachnoid hemorrhage: time course and effect of red blood cell-bound thrombin-activated pro-urokinase and clazosentan. Exp Neurol. 2012;233:357–63.
Muroi C, Fujioka M, Mishima K, Irie K, Fujimura Y, Nakano T, et al. Effect of ADAMTS-13 on cerebrovascular microthrombosis and neuronal injury after experimental subarachnoid hemorrhage. J Thromb Haemost. John Wiley & Sons, Ltd. 2014;12:505–14.
Senchenkova EY, Komoto S, Russell J, Almeida-Paula LD, Yan L-S, Zhang S, et al. Interleukin-6 mediates the platelet abnormalities and thrombogenesis associated with experimental colitis. Am J Pathol. 2013;183:173–81.
Cahill J, Cahill WJ, Calvert JW, Calvert JH, Zhang JH. Mechanisms of early brain injury after subarachnoid hemorrhage. J. Cereb. Blood Flow Metab. SAGE Publications Sage UK: London, England. 2006;26:1341–53.
Interleukin-6 Receptor Mendelian Randomisation Analysis (IL6R MR) Consortium, Swerdlow DI, Holmes MV, Kuchenbaecker KB, Engmann JEL, Shah T, et al. The interleukin-6 receptor as a target for prevention of coronary heart disease: a Mendelian randomisation analysis. Lancet. 2012;379:1214–24.
Bernardo A, Ball C, Nolasco L, Moake JF, Dong J-F. Effects of inflammatory cytokines on the release and cleavage of the endothelial cell-derived ultralarge von Willebrand factor multimers under flow. Blood. 2004;104:100–6.
Vergouwen MDI, Bakhtiari K, van Geloven N, Vermeulen M, Roos YBWEM, Meijers JCM. Reduced ADAMTS13 activity in delayed cerebral ischemia after aneurysmal subarachnoid hemorrhage. J Cereb Blood Flow Metab. 2009;29:1734–41.
Yamashita A, Asada Y. A rabbit model of thrombosis on atherosclerotic lesions. J Biomed Biotechnol Hindawi. 2011;2011:424929–15.
Claassen J, Carhuapoma JR, Kreiter KT, Du EY, Connolly ES, Mayer SA. Global cerebral edema after subarachnoid hemorrhage: frequency, predictors, and impact on outcome. Stroke Lippincott Williams & Wilkins. 2002;33:1225–32.
Suwatcharangkoon S, Meyers E, Falo C, Schmidt JM, Agarwal S, Claassen J, et al. Loss of consciousness at onset of subarachnoid hemorrhage as an important marker of early brain injury. JAMA Neurol. 2016;73:28–35.
Marbacher S, Grüter B, Schöpf S, Croci D, Nevzati E, D'Alonzo D, et al. Systematic review of in vivo animal models of subarachnoid hemorrhage: species, standard parameters, and outcomes. Transl Stroke Res Springer US. 2018;10:250–8.
Breu M, Glatter S, Höftberger R, Freilinger M, Kircher K, Kasprian G, et al. Two cases of pediatric AQP4-antibody positive neuromyelitis optica spectrum disorder successfully treated with tocilizumab. Neuropediatrics Georg Thieme Verlag KG. 2019;50:193–6.
Ringelstein M, Ayzenberg I, Harmel J, Lauenstein A-S, Lensch E, Stögbauer F, et al. Long-term therapy with interleukin 6 receptor blockade in highly active neuromyelitis optica spectrum disorder. JAMA Neurol. 2015;72:756–8.
Toyota Y, Wei J, Xi G, Keep RF, Hua Y. White matter T2 hyperintensities and blood-brain barrier disruption in the hyperacute stage of subarachnoid hemorrhage in male mice: the role of lipocalin-2. CNS Neurosci Ther. 2019;25:1207–14.
Kanamaru H, Suzuki H. Potential therapeutic molecular targets for blood-brain barrier disruption after subarachnoid hemorrhage. Neural Regen Res Medknow Publications. 2019;14:1138–43.
Acknowledgments
We are deeply grateful to: the team of Prof. Hans-Ruedi Widmer, PhD, at the Neurosurgical Research Institute, Department of Neurosurgery, University and University Hospital of Bern, Switzerland, for their assistance in histological staining; Mary Kemper for editing and proofreading and the team of the Experimental Surgical Facility and Central Animal Facility, Department of Biomedical Research, University of Bern, for animal care, anesthesia, and perioperative assistance.
Funding
The current project has been financially supported by the European Association of Neurological Surgeons (EANS) research grant; the research fund of the department of Neurosurgery Kantonsspital Aarau, Switzerland; the HANELA Foundation, Switzerland; and the research fund of the department of Neurosurgery University Hospital Basel.
Author information
Authors and Affiliations
Contributions
Conception and design: Croci, Marbacher. Experimental procedures: Croci, Marbacher, Grueter, Strange; Histological sample preparation and analysis: Croci, Widmer, Di Santo, von Gunten, Wanderer, Andereggen, Sivanrupan. Drafting the article: Croci, Wanderer, Marbacher; Statistical analysis and interpretation of data: Croci, Wanderer, Andereggen, Marbacher; Critically revising the article: Fandino, Widmer, Marbacher, Mariani; Administrative support: Fandino, Mariani.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Ethics Approval
The project has been performed according to the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines [22] and was performed in accordance with the National Institutes of Health Guidelines for the care and use of experimental animals and with the approval of the Animal Care Committee of the Canton Bern, Switzerland (Approval Nr. BE58/17).
Consent for Publication
All the authors agree for the publication of the manuscript.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(PDF 16290 kb)
Rights and permissions
About this article

Cite this article
Croci, D.M., Wanderer, S., Strange, F. et al. Tocilizumab Reduces Vasospasms, Neuronal Cell Death, and Microclot Formation in a Rabbit Model of Subarachnoid Hemorrhage. Transl. Stroke Res. 12, 894–904 (2021). https://doi.org/10.1007/s12975-020-00880-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12975-020-00880-3