
Abstract
Barley is one of the most important cereal crops cultivated over a wider environment in the diverse agro-ecologies in Ethiopia. Study on genotype by environment interaction and stability analysis using thirty barley genotypes was conducted across nine environments in randomized complete block design with three replications to study the magnitude of genotype by environment interaction and to evaluate the stability and adaptability of barley genotypes with high mean yield performance. Additive main effect and multiplicative interaction (AMMI), genotype main effect and genotypes by environment interaction (GGE) and Eberhart and Russell models were employed for stability and adaptability analysis. Thus, AMMI analysis of variance for grain yield showed highly significant (P ≤ 0.001) differences due to genotypes, environment and genotype by environment interaction. Accordingly, environment accounted for (54.61%) of the total variations followed by genotype (10.69%) and G × E interaction (34.70%). Moreover, a substantial percentage of the G × E interaction sum of squares was explained by IPCA1 (45.48%) followed by IPCA2 (24.65%) and IPCA3 (13.02%) while the first two IPCAs explained 70.13%. Moreover, AMMI and GGE were found to be efficient in grouping the barley growing environment in the central highland, whereas Debre Markos and Bekoji were good representative testing environments. Generally, the current study indicated that 3514-A, 24,990, and 17,148 were desirable genotypes for subsequent breeding line identification and variety development.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Barley is an ancient crop in Ethiopia which has been cultivated dating back as early as 5000 years (Gamst 1969; Zemede 2000). Ethiopia is the first important barley producer in Africa in terms of grain volume followed by Algeria and Morocco and the third in area allocated to barley production after Morocco and Algeria (FAOSTAT 2020). Barley is one of the major crops grown next to tef, maize, sorghum and wheat in Ethiopia with a total area of 926,106.9 hectares and total annual production of about 2.34 million tons in the main season (CSA 2021). It can be grown in the highland and mid-highland of Oromia, Amhara, Tigray and SNNP regional states in the altitude range of 1400 and 4000 mas (Zemede 2000), but predominantly cultivated between 2000 and 3000 masl in diverse agro-ecologies (Berhane et al. 1996; Sintayehu et al. 2010). These regional states contribute 48%, 34%, 9%, 9% of barley area and 53%, 32%, 7% and 8% of barley grain production, respectively (CSA 2021). Although the country is recognized as a center of diversity for barley, most of the farmers still obtain very low yield and the current national average barley yield is 2526 kg ha−1 which is far below attainable yield potential (Wondimu et al. 2011; CSA 2021). This low productivity could be attributed to constraints imposed by various abiotic and biotic stress factors and limited availability of stable, high-yielding and adaptable improved varieties (Bayeh and Berhane 2011).
Crop performance is a function of genotype, environment, genotype by environment interaction (GEI) and thus, the importance of understanding crop management and its growing environment has, therefore, been suggested by several researchers to increase agricultural production (Yan and Kang 2003; Kaya et al. 2006; Dia et al. 2017). In this respect, variety development to address food security is one of the major global focuses in general and national crop research of the country in particular. Thus, to maintain high crop productivity, development of varieties with high yield potential performance with good level of stability and adaptability across environment is the ultimate goal of plant breeders in respective crop improvement program (Naroui et al. 2013; Frutos et al. 2014; Tamene et al. 2014). Particularly, barley production is subjected to various stress environments under diverse agro-ecologies in Ethiopia. Accordingly, barley breeding in the country has been focusing on development of varieties which are resistant/tolerant to major biotic and abiotic stress with improved grain yield and acceptable grain quality (Bayeh and Berhane 2011). Thus, besides breeding endeavors for yield improvement, strategic focus on stability and adaptability of the variety is indispensable under the scenario of current climate change. Therefore, information on genotype, environment and G × E interaction is crucial in barley breeding, given that phenotypic response to change in environment is different among genotypes (Sintayehu et al. 2010; Sintayehu and Tesfahun 2011; Muluken and Jemal 2011; Muluken 2013; Abtew et al. 2015).
Analysis of genotype by environment interaction would help in getting information about adaptability and stability performance of genotype (Bose et al. 2014). High genotype by environment interaction for quantitative traits such as grain yield can severely limit gain in selecting superior genotypes for improved variety development (Kang 1990; Flores et al. 1998; Kaya et al. 2006). When the varieties being selected are for a large group of environments, evaluating performance stability and range of adaptation has become increasingly important. Several stability parameters have been proposed to characterize yield stability when genotypes are tested across multiple environments, with each parameter giving different results (Naroui et al. 2013; Tamene et al. 2015). The two major groups of stability statistics are univariate and multivariate stability statistics (Lin et al. 1986). The most commonly used method of univariate analysis in G × E interaction is the linear regression model of Eberhart and Russell (1966) which made further improvement in partitioning the G × E interaction of each variety in to slope of regression coefficient (bi) and deviation from regression line (S2di) (Singh and Chaudhary 1985; Bose et al. 2014). Additive main effect and multiplicative interaction analysis (AMMI) as well as genotype and genotype by environment interaction (GGE) biplot are among the methods of multivariate statistical analysis of multi-environment trials (Naroui Rad et al. 2013). The AMMI model has been extensively applied in statistical analysis and it has a number of promising implications in plant breeding programs (Gauch and Zobel 1988; Crossa 1990). Likewise, GGE has been utilized for variety evaluation in multi environmental trials and it allows interactive visualization of the biplot from various perspectives and relative advantages (Yan 2001; Yan et al. 2007). There is limited information regarding barley genotypes with respect to nature and magnitude of genotype, environment and genotype by environment interaction in the central highland. Therefore, the objectives of the current study were (1) to determine the magnitude of genotype by environment interaction for grain yield and some yield components and (2) to evaluate the stability and adaptability of barley genotypes for their grain yield performance across environments.
Materials and methods
Description of study sites
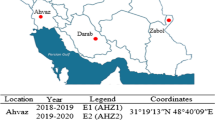
The experiments were conducted in the highland of Ethiopia at nine test environments of Rob Gebeya, Wetebecha Minjaro, Jeldu, Holeta, Midakegn, Bekoji, Werabe, Wolkite and Debre Markos locations during 2019 cropping season, while some of the experimental sites were categorized under acid soil affected barley growing areas of the central highland. These locations are among the major barley production areas which receive sufficient annual rainfall. Description of study sites of barley multi-environment evaluation is shown in Table 1 and geographic locations of testing districts and regional states were designated with different colors (Fig. 1).

Study area of barley multi-environment evaluation and different colors designate districts and regional states of the experimental sites in Ethiopia
Experimental materials
A total of thirty barley genotypes comprising twenty-three promising genotypes which were selected in the preliminary screening trials for acid soil stress along with seven standard check varieties which showed good performance under acid soil condition. These barley genotypes were originally received from Ethiopian Biodiversity Institute (EBI, http://www.ebi.gov.et) and the check varieties were obtained from Holeta barley breeding project.
Experimental design and field management
The experiment was laid out in randomized complete block design (RCBD) with three replications. Plot size was 2.5 m × 1.2 m with 6 rows at 20 cm spacing between rows and 40 cm between plots while spacing between blocks was 1 m. Fertilizer rate and type for each testing site was used based on the recommended rate and time of application for respective areas of the experimental site. Seed rate of 85 kg ha−1 and planting was done by drilling. Pertinent agronomic and weed management practices were implemented as per the standard recommendation. Data were collected from the four middle rows of the experimental plot.
Data collection
Pertinent morpho-agronomic data were collected at some of the testing sites based on standard procedure on plot and plant basis. Data on phenological and agro-morphological traits of the genotypes on plot basis were days to 50% flowering, days to maturity at the stage when 75% of the plants in a plot reached maturity and grain filling period was calculated as a number of days between days to flowering and physiological maturity. Likewise, thousand kernel weight (g), hectoliter weight (kg/hl) and adjusted grain yield (t ha−1) were recorded. Plant height (cm) and spike length (cm) data were recorded on plant basis using ten randomly tagged plants from each plot. However, this paper mainly focuses on grain yield (t ha−1) adjusted at 12% moisture level.
Data analysis
Assumptions of analysis of variance were checked for a valid test of the hypothesis based on the standard analysis procedure. Based on the assumption’s individual location and combined analysis of variance was carried out using different software packages (SAS and GEA-R).
Analysis of variance
Separate analysis of variance for each environment and combined over environments was carried out. Combined analysis of variance was calculated using the model: Yij = μ + gi + ej + geij + eij, where Yij = is the corresponding variable of the ith genotype in jth environment; μ = is the total mean; gi = is the main effect of ith genotype; ej= is the effect of jth environment; geij = is the effect of genotype × environment interaction; eijk= random error.
Stability analysis
The Eberhart and Russell model is one of the regression coefficient models in stability analysis among the several univariate stability parameters proposed by different authors. Eberhart and Russell (1966) made further improvement in stability analysis by partitioning the genotype × environment interaction of each variety into slope of regression coefficient (bi) and deviation from regression line (S2di).
This model is given as: \({X}_{ij}=m+{b}_{i }{I}_{i }+{{\delta }^{2 }}_{ij}\),
where m = the grand mean, \({X}_{ij}\)= the mean of the ith genotype, \({b}_{i}\) =the linear regression coefficient of the ith genotype on environment index, \({I}_{i}\)= the environmental index, i.e., \({I}_{i}\) = (Xj−m), \({{\delta }^{2 }}_{ij}\)= deviation from regression.
Additive main effects and multiplicative interaction (AMMI) analysis
Additive main effects and multiplicative interaction (AMMI) model is one of the multivariate methods which combines the analysis of variance of genotype and environment main effects with principal component analysis of genotype by environment interaction (GEI) into unified approach (Gauch 1988; Zobel et al. 1988). It has proven useful in understanding complex GEI. The data of each trial were analyzed using this model because this model partitions the GEI sum of squares into interaction principal component (IPCA) axes. The AMMI analysis of variance summarizes most of the magnitude of genotype by environment interaction into one or few interactions principal component analysis (IPCA). The AMMI statistical model is given as:
where Yijk is yield of genotypes i in environment j for the kth replication, µ is the grand mean, gi is the genotype i mean deviation (genotype mean minus grand mean), ej is the environment j mean deviation, N is the number of singular value decomposition (SVD) axes retained in the model, λn is singular value for SVD axes n, αin is the genotype i eigenvector value for SVD axes n, βjn is the environment j eigenvector value for SVD axes n, θij ~ N(0, σ2ge) is the genotype by environment interaction residual, εijk ~ N (0,σ2ε) is the error term and θij and εijk are independent that is to say cov (θij, εijk) = 0.
Analysis of both additive main effects and multiplicative interactions (AMMI), and genotype plus genotype by environment (GGE) were performed using the Genotype by Environment Analysis with R (GEA-R) version 4.0 (Pacheco et al. 2016).
AMMI stability value (ASV) as described by Purchase et al. (2000) is used to further investigate the stability of the genotypes. Generally, the AMMI model does not make provision for a quantitative stability measure, such a measure is essential to quantify and rank genotypes according to their yield stability. In effect the ASV is the distance from zero in a two-dimensional scatter of IPCA1 scores against IPCA2 score. Since the IPCA1 score contributes more to the G × E sum of squares, it has to be weighted by the proportional difference between IPCA-2 score to compensate for the relative contribution of IPCA1 and IPCA-2 to the total G × E sum of squares.
where ASV AMMI stability value.
IPCA1 = interaction principal component analysis1 scores for the specific genotype; IPCA2 = interaction principal component analysis2 scores for the specific genotype; SSIPCA1 = sum of square of the interaction principal component 1; SSIPCA2 = sum of square of the interaction principal component 2.
Genotype and genotype by environment interaction (GGE) biplot analysis
The concept of GGE originates from analysis of multi-environment trials of crop varieties. Performance of a variety in an environment is a mixed effect of genotype main effect (G), Environment main effect (E), and genotype by environment (GE) interaction (Yan 2001; Kaya et al. 2006). Hence, G and GE are relevant for the purpose of variety evaluation and the term GGE was adopted (Yan et al. 2000). GGE biplot enables visual observation to understand which genotypes are best performed in which environment or which genotypes are stable and unstable. Moreover, GGE biplot is also used to visualize the discriminating ability and representativeness of the test environment (Yan 2001).
Results and discussion
Analysis of variance
Highly significant (P ≤ 0.01) variation was detected among the genotype, environment and genotype by environment interaction for almost all the traits considered in this study (Table 2). The existence of significant variation among environments might be attributed to variation in climatic and soil variables across testing environments. The highly significant variation among genotypes revealed the existence of high inherent variability among the barley genotypes and significant G × E interactions suggest that agronomic traits of the genotypes varied across the tested environments. This result revealed that there was differential yield performance among barley genotypes across testing environments. Similarly, significant genotype by environment interaction and environmental effects on yield and agronomic traits of barley were reported by several researchers in various part of Ethiopia (Muluken et al. 2010; Sintayehu and Tesfahun 2011; Muluken and Jemal 2011; Zerihun 2011; Gebremedhin et al. 2014). According to Fox et al. (1996), significant G × E interaction indicates differential genotypic performance across environments and reduces the association between phenotypic and genotypic values, and thus genotypes that perform well in one environment may perform poorly in another. Generally, genotypes by environment interaction can be either exploited by selecting superior genotypes for specific target environments or avoided by selecting widely adapted stable genotypes across a wide range of environments (Ceccarelli 1989; Adugna and Labuschagne 2002).
Mean grain yield performance
The mean grain yield across the nine test environments ranged from 1.7 t ha−1 at Rob Gebeya to 4.7 t ha−1 at Bekoji with a grand mean of 3.10 t ha−1 (Table 3). Bakoji followed by Werabe were identified to be the best environments with mean grain yield greater than the grand mean. On the other hand, mean grain yield among thirty barley genotypes at nine test environments ranged from 4.01 t ha−1 for G3 to 2.35 t ha−1 for G20. Moreover, G27, G7, G30, G12, G16, G25 and G29 were among the highest-yielding genotypes. In the earlier multi-environment national barley variety trial, Bekoji was characterized as a high-yielding environment, while South Gonder was low-yielding and Holeta being intermediate yielding (Wondimu and Berahne 2016).
Analysis based on Eberhart and Russell regression model
Linear regression for average yield of single genotype on the average yield of all genotypes in each environment resulted in regression coefficient (bi) ranging from 0.42 to 1.69 and deviation from regression (S2di) ranging from − 0.01 to 1.91 (Table 3 and Fig. 2). This variation in regression coefficient indicates differential response of barley genotypes to environmental changes. According to Eberhart and Russell (1966) stable genotypes should have a regression coefficient (bi = 1) and a minimum deviation from the regression line (S2di = 0). When this combines with high yield, that genotype could be considered suitable for wider cultivation. In the present study, high-yielding genotypes G3, G27, G7, G30, G12, G16, G29, G21, and G25 had regression coefficient of 1.42, 1.52, 1.44, 1.26, 1.26, 0.83, 1.29, 0.81 and 1.69, respectively (Table 3 and Fig. 2B). The respective values of deviation from regression for these genotypes were 0.21, 0.47, 0.25, 0.39, 0.29, 1.14, 0.96, 0.12 and 0.09. According to this model, G30, G12, G21 and G16 were relatively stable genotypes as their regression coefficient was close to unity with high yield potential. On the other hand, G3, G7, G12, G21 and G25 had relatively small deviations from regression with high yield potential while G19 and G16 had high values of deviation from regression (Table 3 and Fig. 2A). However, G1, G4, G11, G14, G10 and G13 were among the stable genotypes in terms of both regression coefficient and deviation from regression but characterized with low-yield potential performance. Therefore, considering yield and stability, G12 and G21 were promising genotypes. Bose et al. (2014) elaborated that the commonly used in G × E interaction is the linear regression model of Eberhart and Russell (1966) in which regression value (bi) gives information about adaptability and deviation from regression(s2di) used as measure of stability of performance.

Plot of genotype mean grain yield versus deviation from regression (S2di) (A), and Plot of genotype mean grain yield versus regression coefficient (bi) (B); vertical broken line designates grand mean of barley genotypes
Furthermore, the result of this study was in agreement with similar works of stability assessment in fifteen barley genotypes tested at twelve environments in Ethiopia indicating regression slope and deviation from regression values within the range of 0.65–1.56 and 0.50–23.5, respectively (Muluken and Jemal 2011). Likewise, fifteen barley genotypes were evaluated in the national variety trial at ten environments and stability analysis showed regression slope and deviation from regression within the range of 0.60–1.5 and 0.11–0.55, respectively. Consequently, a stable and high-yielding variety was identified and approved for national release (Wondimu and Berhane 2016). Moreover, Bose et al. (2014) in rice and Dagnachew et al. (2014) finger millet genotypes stability and adaptability study, reported similar results indicating significance of the model in identifying genotypes for the target environments.
Generally, Eberhart and Russell (1966) noted that regression values above 1.0 describe genotypes with higher sensitivity to environmental change (below average stability) and greater specificity of adaptability to high-yielding environment, whereas regression coefficient less than 1.0 provides greater resistance to environmental change (above average stability), and thus showing tendency to specific adaptation at low-yielding environments.
Additive main effect and multiplicative interaction (AMMI) analysis
AMMI analysis depicted a highly significant (P ≤ 0.001) difference for grain yield (t ha−1) of thirty barley genotypes owing to genotypes, environment and genotype by environment interaction.
Accordingly, environment accounted for (54.61%) of the total variations followed by genotype (10.69%) and G × E interaction (34.70%) (Table 4). The magnitude G × E interaction sum of squares was three times larger than that of genotypes, indicating that there were sizable differences in genotypic response across the environment. Large yield variation explained by environment followed by GEI revealed that the environments were diverse, with large differences among environmental means causing most of the variation in grain yield. Moreover, significant variation of genotype also indicates differential grain yield performance among genotypes tested across environments.
Various studies in barley multi-environment trial disclosed that large proportion of the variations attributed to environment followed by G × E interaction (Muluken et al. 2010; Zerihun 2011; Gebremedhin et al. 2014; Verma et al. 2016). Similarly, several authors reported significant G × E interaction indicating larger proportion of variation attributed to environment followed by G × E interaction and genotype in the stability analysis of various crop genotypes such as; wheat (Kaya et al. 2002), rice (Samonte et al. 2005), cowpea (Kasaye et al. 2013), finger millet (Dagnachew et al. 2014) and faba bean (Tamene et al. 2014).
In the present study, the presence of genotype by environment interaction was evidently demonstrated by the AMMI model. When the interaction was partitioned among the first four interaction principal component axis (IPCA) which cumulatively captured 89.75% of the total GEI, while the remaining IPCAs contributed 10.25% (Table 4). Thus, a sufficient percentage of G × E interaction was significantly explained by IPCA1 (45.48%) followed by IPCA2 (24.65%), IPCA3 (13.02%) and IPCA4 (6.60%). Furthermore, AMMI analysis depicted that the first two principal components contributed maximum proportion (70.13%), hence IPCA1 and IPCA2 were used to describe two-dimensional AMMI biplot for further analysis of genotype stability. This result disclosed that there was differential yield performance among barley genotypes across testing environments owing to presence of G × E interaction. In similar studies of barley genotypes stability analysis. Proportion of G × E interaction of explained by IPCA1 and IPCA2, respectively, were 52.0% and 28.4% by Muluken et al. (2010), 44.4% and 21.3% by Abtew et al. (2015), 44.9% and 25.0% by Verma et al. (2016). Likewise, studies of other crop genotypes describing the magnitude G × E interaction explained by the first two principal components were 50.8% and 27.9% in wheat by Kaya et al. (2002), 40.9% and 27.0% in rice by Samonte et al. (2005), 62.6% and 31.2% in rice by Bose et al. (2014), 66.1% and 12.8% in finger millet by Dagnachew et al. (2014), 53.0% and 19.5% in tef by Habte et al. (2019).
AMMI stability value (ASV)
AMMI stability value aids selection of relatively stable high-yielding genotypes. Thus, an ideal genotype should have high mean grain yield and small ASV. In this study, ASV ranged from 0.19 to 1.91. Accordingly, G3, G7, G21 and G25 had the highest mean yield of 4.01, 3.74, 3.34, 3.33 t ha−1 with intermediate ASV of 0.74, 0.79, 0.55, 0.67, respectively. On the other hand, G6, G28 and G14 had the lowest ASV of 0.19, 0.20 and 0.20 with yield performance lower than grand mean. Moreover, G16, G17, and G27 genotypes had better grain yield performance of 3.46, 3.22, 3.78 t ha−1 but high ASV of 1.39, 1.15, 1.05, respectively signifying their potential for specific adaptation (Table 3). In a similar study of multi environmental evaluation of fifteen barley genotypes, having ASV ranging from 0.27 to 2.14 was reported that the highest-yielding genotypes were characterized with a score of an intermediate ASV (Wondimu and Berhane 2016). In AMMI analysis of grain yield stability in rice ASV of 0.13–1.48 observed in which top yielding genotypes had intermediate values (Bose et al. 2014). ASV in wheat also reported within the range of 0.17–5.119 (Naroui Rad et al. 2013). Moreover, a similar study of finger millet varieties by Dagnachew et al. (2014) described a wide range of ASV (0.27–7.42), while the result of two finger millet accessions showed relatively better stability with small to intermediate ASV of the other high-yielding genotypes.
AMMI1 biplot
AMMI biplot is a glimpse for displaying genotype main effect and interaction effect of the genotype and environment simultaneously. AMMI biplot analysis is considered to be an effective tool to diagnose interaction patterns graphically in which display of PCA scores plotted against each other provides visual inspection and interpretation of G × E interaction (Kaya et al. 2002). The relation between the pair of environments or genotypes in the biplot is proportional to the response they show to genotype by environment interaction effect (Crossa et al. 1990). Generally, biplot aids the interpretation of interaction effect among genotype and environment in the study of genotype adaptability. Biplot of the first interaction principal component axis (IPCA1) of both genotype and environment versus the main effects gives information on mean yield and genotype stability. As shown in Fig. 2A, the result of AMMI biplot in which abscissa and ordinate were designated as main effects and the IPCA1 scores, respectively. The biplot depicted that relatively the environmental effect scores were more scattered than genotype effect scores indicating variability owing to the environment is greater than variability due to genotypes. Oliveira et al. (2014) studied stability of yellow passion fruit varieties at eight environments describing similar findings.
Accordingly, G1, G4, G6, G10, G2, G28 and G14 were among the genotypes located near the origin with low contribution to genotype by environment interaction. On the other hand, G19, G16 and G18 were those genotypes which highly contributed to the interaction effect implying specific adaptation of these genotypes. Moreover, Bekoji, Debre Markos and Holeta had positive IPCA1 score and positively interacted with G3, G7, G27, G12, G30, G29, G26, G25, G24 genotypes with good yield performance (Fig. 3A). Similar study of AMMI analysis in rice by Nasssir and Ariyo (2011), cowpea by Kasaye et al. (2013), rice by Bose et al. (2014) were reported.

AMMI1 biplot of IPCA-1 against grain yield (A) and AMMI-2 biplot of IPCA against IPAD-2 for grain yield of thirty barley genotypes tested across nine environments (B)
Generally, genotypes that are characterized by mean greater than grand mean and IPCA score close to zero were considered as adaptable to all environments, while genotypes with high mean grain yield and large value of IPCA score were considered as having specific adaptation. Genotypes and environments on the same parallel line, relative to the ordinate have similar yields, and those relative abscissae have similar interaction pattern while those on the right side of the midpoint of the axis have higher yields than those on the left side (Nassir and Ariyo 2011; Bose et al. 2014). Besides, genotypes or environments located on the right side of the midpoint of the perpendicular line had higher yield than those on the left side. Thus, G3, G7, G27, G12, G30, G29, G26, G25, G2, G21, G17 and G16 had higher yield compared to those genotypes located on the left side. Moreover, AMMI1 analysis positioned the genotypes in different environments indicating interaction trends of the genotypes. Thus, Bekoji and Midakegn were the most discriminating environments as indicated by long distance from the origin, while Watabacha Minjaro and Wolkite environments were characterized by short vector length signifying less discriminating power. Accordingly, Bekoji and Midakegn testing environments were located distant from the origin signifying that these locations had higher contribution to magnitude of G × E interaction, while Watabacha Minjaro and Wolkite were located near to origin indicating low contribution of these locations to the interaction (Fig. 3A).
AMMI2 biplot
AMMI2 is an exploratory technique by which the G × E relationship can be expressed in terms of interaction patterns derived in biplot (Bose et al. 2014). AMMI2 biplot presents the spatial pattern of the first two IPCAs of the interaction effects and helps in visualizing interpretation of G × E interaction pattern and identifies genotypes or locations that exhibit low, medium or high level of interaction effect. In the current study, AMMI2 biplot analysis based on grain yield of barley genotypes tested at nine environments is presented in Fig. 3B. Genotypes which are very close to the origin are more stable than those away from the origin. Thus, G11, G20, G14, G28 and G6 were stable genotypes though they had below average yield performance. The high-yielding genotypes such as G3, G7, G12, G25, G26, and G30 were moderately in a proximity to the origin. Unlike in the case of AMMI1 G2, G1, G10 and G15 become unstable under AMMI2 biplot. Thus, the stability information obtained using AMMI2 is more precise than AMMI1 biplot as AMMI2 contains information from both IPCA1 and IPCA2 (Oliveira et al. 2014). Moreover, based on AMMI2 analysis Midakegn, Bekoji and Debre Markos were located far from the origin while Wolkite and Jeldu were very close to the origin (Fig. 3B). Generally, the longer the vector projected from the origin indicates a discriminating environment among the genotypes evaluated whereas environments with a short length vector from the origin indicate low interaction of these locations with genotypes evaluated. Several studies disclosed that genotypes near the origin are non-sensitive environment interactive forces and those distant from the origin have large interactions (Samonte et al. 2005; Pacheo et al. 2016). If the genotypes have nearly zero IPCA1 score they are stable and show general adaptation across their testing environments (Flores et al. 1998). If genotypes are far from zero it is highly responsive or does not perform consistently across environments. On the other hand, if genotypes and environments have similar sign on the IPCA score in close proximity on the biplot graph, then the interaction between them is positive and they have positive association which facilitate formation of environments with similar agronomic performance (Kaya et al. 2006; Bose et al. 2014; Olivera et al. 2014).
Genotype and genotype by environment interaction (GGE) biplot analysis
Yield performance of the genotype in a given environment is a mixed effect of genotype main effect (G), environment main effect (E) and genotype by environment (GE) interaction (Kaya et al. 2006). GGE enables to classify the study genotypes and environments based on the performance of genotypes and response of the growing environments. GGE biplot often visualized on the basis of results explained for the first two principal components (Yan 2001). Thus, in the present study, the first and the second principal components of GGE biplot explained 45.06% and 25.19% sum of squares of GEI, respectively. Both principal components together accounted for 70.24% of the sum of squares of GEI. A similar study of GGE analysis of multi-environment yield trials of barley in Southeastern Ethiopia showed that PC1 and PC2 explained 52.2% and 19.6% of GGE sum of squares (Zerihun 2011). Likewise, study of GGE analysis of multi-environment yield trials in wheat by Kaya et al. (2006) stated that 46.2% and 15.8% of GGE sum of squares explained by PC1 and PC2, respectively. Habte et al. (2019) also reported in GEI and stability analysis in tef in which the first two principal components of GGE explained 49.8% and 23.0% of the total variation. According to Kaya et al., (2006) genotypic PC1 scores > 0 detect the adaptable and/or higher-yielding genotypes, while PC1 scores < 0 discriminate against the non-adaptable and/or lower-yielding ones. Unlike genotypic PC1 scores, near-zero PC2 scores identify stable genotypes, whereas absolute larger PC2 scores detect the unstable ones. Thus, favorable test environments should have large PC1 scores (more discriminating of the genotypes) and near-zero PC2 scores (more representative of an average environment) (Yan et al. 2001).
Ranking of genotypes and environments
Although an ideal genotype and ideal environment may not exist in reality, it can be used as a reference point for genotype evaluation and selection in multi-environment yield trials (Kaya et al. 2006). Genotypes and environments that fell in the central (concentric) circle are considered ideal genotypes and environments (Yan and Rajcan 2002). The genotypes are ranked in the direction of the tester axis and the perpendicular line separates genotypes that perform below average from those performing above average in a given environment (Yan 2001). Ideal genotype should have both high mean yield performance and high stability across environments (Kaya et al. 2006; Yan and Tinker 2006). Figure 4A illustrates ranking of genotype relative to the ideal genotype. The arrow indicates where an ideal genotype should be located and genotypes located closer to the ideal genotype are more desirable compared to others. Consequently, G3 and G27 which were located in the center of concentric circles were ideal genotypes in terms of high-yielding ability and stability compared with the rest of the genotypes. In addition, G7, G29, and G12 were genotypes located in the next concentric circles and can be regarded as desirable genotypes whereas G19, G20, G18, G5 and G23 were situated far from the concentric circle and less desirable genotypes. In a similar study of wheat GGE biplot analysis in which three genotypes were designated as ideal genotypes in terms of yielding ability and stability while five genotypes were considered desirable genotypes (Kaya et al. 2006).

Ranking genotype based on yield and stability (A); GGE biplot showing discriminative vs representativeness and relationship among test environment (B)
An environment is more desirable and discriminating when located closer to the center of a circle or to an ideal environment. An environment is more desirable if it is located closer to the ideal environment (Naroui et al. 2013). The ideal test environment should be most discriminating and also most representative of the target environment (Kaya et al. 2006). Figure 4A describes an ideal test environment which is the center of concentric. Accordingly, Debre Markos was closest to this point and therefore considered as a good environment for barley evaluation whereas Midakegn and Rog Gebeya were found to be the poorest for selecting genotypes adapted to central highland. Kaya et al. (2006) elaborated in GGE biplot analysis of multi-environment yield trials in wheat indicating similar findings. However, Yan and Tinker (2006) noted that for a specific test location to be considered as an ideal environment, the experiment needs to be evaluated over locations and years. Yan et al. (2001) and Kaya et al. (2006) indicated that a favorable test environment must have large PC1 scores (more discriminating genotypes) and small PC2 scores (more representative of an average environment).
Relation among the environments
In this study, mean grain yield of barley genotypes tested at nine test environments which was visualized by the line connecting each environment to biplot origin (Fig. 4B). Both the Environment PC1 and PC2 had negative and positive scores indicating that there was a difference in ranking of yield performance among genotypes across environments leading to possible cross over interaction. Similar study revealed a positive environmental PC1 score (non-crossover type), while both negative and positive PC2 scores (Kaya et al. 2006). However, in the study of barley stability analysis, Zerihun (2011) disclosed that environment PC1 scores were negative indicating crossover type and positive PC2 scores showing non-crossover type of G × E interaction.
Vector view of the GGE biplot provides a brief summary of the interrelationship among the environments. The cosine of the angle between two environments was used to calculate the correlation between them (Yan 2001; Dehghani et al. 2009; Kaya et al. 2006). When the angle between two locations or environments is < 90° (acute angle), the test environments are positively correlated whereas formation of angle > 90° (obtuse angle) between the test environments signifies negative association (Yan and Tinker 2006; Frutos et al. 2014). Accordingly, the angle between Holeta and Wolkite was close to 90° showing the absence of correlation which signifies independent genotypic performance in those environments. Moreover, Holeta, Rob Gebya and Watabacha Minjaro were positively correlated as the angels among their vectors were smaller than 90°. Similarly, Jeldu, Wolkite and Werabe were positively associated. When environments are correlated in their ranking of genotypes, these environments produce similar information regarding the tested genotypes and thus, some of the testing environments can be omitted without affecting validity of the information (Yan and Tinker 2006; Yan et al. 2007). The angle between Holeta and Midakegn as well as Bekoji and Midakegn was greater than 90°, indicating negative correlation. Obviously, the strong negative correlation between environments indicates high crossover interaction (Yan and Tinker 2006; Kaya et al. 2006). In general, similarity (covariance) between two environments is determined by both length of their vector and cosine of the angle between them (Yan and Tinker 2006).
Representativeness and discriminating ability of the environment
GGE biplot is useful to assess how much of a test environment is capable of generating unique information about the differences among genotypes and representativeness of the mega environment. Therefore, among the nine environments studied, Bekoji, Midakegn and Debre Markos were most discriminating (informative), whereas Rob Gebeya and Werabe were among the least discriminating (Fig. 4B).
The vector length, that is the absolute distance between the marker of an environment and the plot origin, is a measure of the discriminating ability; thus, the longer the vector, the higher discriminative power of the environment (Frutos et al. 2014). When the test environment has a very short vector, it means that all genotypes performed similarly thus provide little information about the genotypes difference (Yan et al. 2007; Yan and Tinker 2006). The average environment (denoted by the small circle at the end of the arrow) has the average coordinate of all the test environments.
The average environment axis (AEA) is the line that pass through the average environment and biplot origin. Accordingly, Debre Markos was more representative followed by Bekoji, whereas Midakegn was least representative (Fig. 4B). Test environments that have smaller angle with the AEA are more representative of other environments, whereas large deviations from AEA indicate least representative once (Yan and Tinker 2006; Yan et al. 2007). In general, from GGE analysis of barley genotypes revealed that, Debre Markos and Bekoji were found to be both discriminating and representative. Environments that are both discriminating and representative are good test environments for selecting adapted genotypes (Yan and Tinker 2006).
Genotype performance and mega environment determination
GGE biplot enables to classify the study genotypes and environments based on the performance of genotypes and the response of the growing environments. One of the most attractive features of GGE biplot is its ability to show the which-won-where pattern of genotype by environment (Yan 2001; Yan and Tinker 2006). A polygon view was created by vertex genotypes G30, G27, G3, G16, G19, G18, G20 and G5 which were found away from the biplot origin such that all other genotypes were contained within the polygon. The lines coming from the origin which are perpendicular to the line connecting the furthest genotypes divided the test environments and genotypes into different groups. Based on this grouping, the nine test environments were grouped into three mega environments, whereas the thirty test genotypes were grouped into five genotypic groups (Fig. 5A). Bekoji, Debre Markos, Wolkite and Watabacha Minjaro testing environments fell into one of the sectors, while vertex genotypes for this sector were G3 and G27 suggesting that these genotypes were higher yielding for these environments. Similarly, Holeta and Rob Gebeya were found to be in the second sector and the vertex genotype for this sector was G25. Midakegn was also the other single environment in the other sector while G16 and G19 were the vertex genotypes, suggesting that the higher-yielding genotypes for this environment were G16 and G19. According to Yan and Tinker (2006), genotypes located on the vertices of the polygon perform either the best or the poorest in one or more environments. The other vertex genotypes G5 and G20 had no corresponding environment implying that these genotypes were not the best in any of the environments, which means they were the poorest genotypes in some or all of the environments.

Polygon view of the GGE showing ‘which won- where’ (A) and mean versus stability view of GGE (B)
Yan and Tinker (2006) also reported similar observations. Generally, it is pertinent to conduct the same experiment over years and locations to determine mega environments of barley production in the central highland. Likewise, Kaya et al. (2006) noted that mega-environment patterns need to be verified through multi-year and environment trials conducted in the target region.
Assessment of genotypes yield performance and their stability
Within a single mega-environment, genotype should be evaluated for both mean performance and stability across the environment. Average environment is defined by the average PC1 and PC2 scores of all environments represented by small circles (Kaya et al. 2006). The arrow on the axis of average environment coordinate (AEC) abscissa (AEA) points in the direction of higher mean performance of genotypes and consequently ranks the genotypes with respect to mean performance. The axis of average environment coordinate ordinate is the line that passes through the biplot origin and perpendicular to AEC abscissa (Yan 2001; Kaya et al. 2006).
In the present study, analysis of the GGE biplot view is shown in (Fig. 5B). Therefore, G3 had the highest mean yield followed by G27 and G7, whereas G20, G23 and G15 were among the low-yielding genotypes. Thus, genotypes with above average yield potential could be selected for future variety evaluation or subsequent breeding works. According to Kaya et al. (2006) and Yan et al. (2007) AEC ordinate separates genotypes with above average means from below average means. Moreover, AEC ordinate points to the greater variability (poor stability) in either direction (Naroui et al. 2013). Thus G3, G7 and G27 were close to AEA indicating relative stability, while G19 and G16 were among the highly unstable genotypes. On the other, one-third of the genotypes with above average means were from G3 to G24 whereas genotypes below average were from G2 to G20 (Fig. 5B). From the perspective of stability and mean performance; G20, G23 and G15 were among the stable but the lowest yielding genotypes. This result is in conformity with the report of Zerihun (2011). Generally, the concept of high stability is desirable only when evidently associated with good mean yield performance which is meaningful in economic terms (Yan and Tinker 2006).
Summary and conclusion
Promising barley varieties need to be adapted to a broad range of environmental factors to ensure food security and address the industrial demand of barley in Ethiopia. Farmers are more interested in varieties which produce consistent yield under their growing circumstances. Likewise, barley breeders are envisioned to develop promising varieties which are consistently high yielding with relatively broad adaptation and tolerant/ resistant to biotic and abiotic stresses. Thus, information on genotype by environment interaction and stability is of paramount importance for barley breeders and farmers engaged in barley production in the country.
Accordingly, multi-environment evaluation of barley genotypes across nine locations in the central highland was initiated to study magnitude of genotype by environment interaction and determine yield performance and stability across test locations. In this study, AMMI, GGE and Eberhart and Russell models were employed in the evaluation of thirty genotypes, whereas both AMMI and GGE models better elaborated to quantify the magnitude of genotype, environment and genotype by environment interaction to determine extent of stability and adaptability. Accordingly, combined analysis of variance depicted highly significant variation for grain yield and some agronomic traits among genotypes, environments and G × E interaction. The result of AMMI analysis of variance in this study indicated that barley grain yield performances were highly influenced by environmental effects followed by G × E interaction and genotypes. The magnitude of the G × E interaction effect was about three times higher than that of genotype. Moreover, AMMI analysis further partitioned the G × E variance into interaction principal component axes (IPCA) to investigate stability of genotypes. Accordingly, a significant percentage of G × E interaction was explained by IPCA1 (45.48%) followed by IPCA2 (24.65%), IPCA3 (13.02%) and IPCA4 (6.60%). The significant IPCAs together accounted for 89.8%, whereas the first two IPCAs contributed a maximum proportion of 70.13%.
Polygon view of the GGE biplot revealed that there exist three possible mega environments; whereas, the thirty test genotypes were grouped into five genotypic groups. Among the genotypes tested there were desirable genotypes in terms of high mean yield and stability. 3514-A(G3) and the earlier released popular variety HB-1307(G27) which located into the center of concentric circles were ideal genotypes in terms of higher-yielding ability and stability. Likewise, 17,148 (G7), (Ibon174/03 (G29), and 24,990 (G12) were located next to concentric circles may be regarded as desirable genotypes. It is pertinent to note particularly, 3514-A was one of the top five genotypes in all the test environments. Therefore, 3514-A(G3), 24,990 (G12), and 17,148 (G7) are stable and high-yielding promising barley genotypes for subsequent breeding line identification and variety development while others can be used as potential breeding materials.
Ranking of environment based on analysis of discriminating ability and representativeness results disclosed that Debre Markos was in a close proximity to the reference point of ideal environment, and therefore considered as favorable barley growing environments. Environment PC1 and PC2 had both negative and positive scores indicating differential ranking of yield performance among genotypes across environments leading to possible crossover type G × E interaction which might necessitate breeding for specific adaptation as breeding strategy. However, additional barley yield trials over locations and over years may be initiated to clearly identify and decide possible mega environments and breeding approaches.
References
Abtew WG, Lakew B, Haussmann BIG, Schmid KJ (2015) Ethiopia barley landraces show yield stability and comparable yield to improved varieties in multi environment trial. J Plant Breed Crop Sci 7(8):275–291
Adugna W, Labuschagne MT (2002) Genotype-environment interaction and phenotypic stability analyses of linseed in Ethiopia. Plant Breed 121:66–71
Bayeh M, Berhane L (2011) Barley research and development in Ethiopia- an overview.pp1–16. In: Mulatu B, Grando S (eds) Barley Research and Development in Ethiopia. Proceedings of the 2nd National Barley Research and Development Review Workshop. 28–30 November 2006, HARC, Holetta, Ethiopia. ICARDA, PO Box 5466, Aleppo, Syria. pp xiv + 391
Berhane L, Hailu G, Fekadu A (1996) Barley production and research. pp 1–8. In: Hailu G, Joob VL (eds) Barley research in ethiopia: past work and future prospects. In: Proceedings of the first barley research review workshop 16–19, October. 1993. Addis Ababa, IAR/ICARDA. Addis Ababa, Ethiopia
Bose LK, Jambhulkar NN, Singh ON (2014) Additive main effect and multiplicative interaction analysis of grain yield stability in early duration rice. J Anim Plant Sci 24(6):1885–1897
Cecceralli S (1989) Wide adaptation: how wide. Euphytica 40:197–205
Crossa J (1990) Statistical analysis of multi-location trials. Adv Agron 44:55–85
CSA (2021) Central statistical agency. Agricultural sample survey 2020/21/(2013 E.C.) Area and Production of Major Crops (Private Peasant Holdings, Meher Season). V1. Statistical Bulletin No.590. Addis Ababa, Ethiopia
Dagnachew L, Masresha F, de Santie V, Kassahun T (2014) Additive main effects and multiplicative interactions (AMMI) and genotype by environment interaction (GGE) biplot analyses aid selection of high yielding and adapted finger millet varieties. J Appl Biosci 76:6291–6303
Dehghani H, Sabaghnia N, Moghaddam M (2009) Interpretation of genotype-by-environment interaction for late maize hybrids grain yield using a biplot method. Turk J Agric For 33:139–148
Dia M, Wehner TC, Arellano C (2017) RG×E: an r program for genotype × environment interaction analysis. Am J Plant Sci 8:1672–1698. https://doi.org/10.4236/ajps.2017.87116
Eberhart SA, Russel WA (1966) Stability parameters for comparing varieties. Crop Sci 17:36–40
FAOSTAT (2020) Food and agriculture organization of the united nations (FAO). Crops and livestock products. https://www.fao.org/faostat/en/#data/QCL. Accessed March 18 2022.
Flores F, Moreno MT, Cubero JJ (1998) Comparison of univariate and multivariate methods to analyze GE interaction. Field Crop Res 56:271–286
Fox PN, Skovm B, Thomson BK, Braun HJ, Comier R (1996) Yield and adaptation of hexaploid spring triticale. Euphytica 47:57–64
Frutos E, Galindo MP, Levia V (2014) An interactive biplot implementation in R for modeling genotype by environment interaction. Stoch Environ Res Risk Assess 28:1629–1641. https://doi.org/10.1007/s00477-013-0821
Gamst FC (1969) The Qemant: a pagan-hebraic peasantry of ethiopia. Waveland Press, Homewood
Gauch HG (1988) Model selection and validation for yield trials with interaction. Biometrics 44:705–715
Gauch HG, Zobel RW (1988) Predictive and postdictive success of statistical analyses of yield trials. Theor Appl Genet 76:1–10
Gebremedhin W, M Firew, B Tesfaye (2014) Stability analysis of food barley genotypes in northern Ethiopia
Habte J, Kebebew A, Kasahun T, Kifile D, Zerihun T (2019) Genotype-by-environment interaction and stability analysis in grain yield of improved tef (Eragrostis tef) varieties evaluated in Ethiopia. JEAI 35(5):1–13
Kang MS (1990) Understanding and utilization of genotype-by-environment interaction in plant breeding, pp. 52–68. In: Kang MS (ed) Genotype-by-environment, interaction and plant breeding, Louisiana State University, Department of Agronomy, Baton Rouge
Kasaye N, Kidane T, Setegn G, Berhanu A (2013) Multi environment evaluation of cowpea (Viginia Unguiculata L.) Varieties in the drought prone areas of Ethiopia. Ethiop J Crop Sci 3:77–92
Kaya Y, Palta C, Taner S (2002) Additive main effect and multiplicative interactions analysis of yield performance in bread wheat across environments. Turk J Agric For 26:275–279
Kaya Y, Akcra M, Taner S (2006) GGE- biplot analysis of multi-Environment yield trial in bread wheat. Turk J Agric For. 30:325–337
Lin CS, Binns MR, Lefkovitch LP (1986) Stability analysis, where do we stand? Crop Sci 26:894–900
Muluken B (2013) Study on malting barley genotypes under diverse agro ecologies of north western Ethiopia. Afr J Plant Sci 7(11):548–557. https://doi.org/10.5897/AJPS10.068
Muluken B, Jemal E (2011) Achievements in food barley breeding research in the early production systems of North West Ethiopia. pp. 83–89. In: Ceccarelli S, Grando S (eds) Proceedings of the 10th International Barley Genetics Symposium, 5–10 April 2008, Alexandria, Egypt. ICARDA, PO Box 5466, Aleppo, Syria. pp xii + 794.
Muluken B, Habtamu Z, Alemayehu A (2010) Genotype by environment interactions (G × E) and stability analyses of malting barley (Hordeum Distichon L.) genotypes across northwestern Ethiopia. Ethiop J Agric Sci 20:168–178
Naroui Rad MR, Abdul Kadir M, RafiiHawa MY, Jaafar Naghavi MR, Ahmadi F (2013) Genotype× environment interaction by AMMI and GGE biplot analysis in three consecutive generations of wheat (Triticum aestivum) under normal and drought stress conditions. Aust J Crop Sci 7(7):956–961
Nassir AL, Ariyo OJ (2011) Genotype × environment interaction and yield stability analysis of rice grown in tropical inland swamp. Not Bot Hort Agrobot Cluj 39(1):220–225
Oliveira EJ, de Freitas JPX, de Jesus ON (2014) AMMI analysis of the adaptability and yield stability of yellow passion fruit varieties. Sci Agricola 71(2):139–145
Pacheo AM, Vargas G, Alvarado F, Rodriguez M, Lopez J, Crossa JB (2016) GEA-R (genotype x environment analysis with R for windows.) Version 4.0. International Maize and Wheat Improvement Center (CIMMYT). http://hdl.handle.net/11529/10203
Purchase JL, Hatting H, Vandeventer CS (2000) Genotype × environment Interaction of winter wheat (Triticum aestivum L.) in South Africa: II. Stability analysis of yield performance. South Afr J Plant Soil 17:101–107
Samonte SOPB, Wilson LT, McClung AM, Med-ley JC (2005) Targeting cultivars onto rice growing environments using AMMI and SREG GGE biplot analysis. Crop Sci 45:2414–2424
Singh RK, Chaudhary BD (1985) Biometrical methods in quantitative genetic analysis. Kalyani Publishers, New Delhi-Ludhiana
Sintayehu D, Tesfahun A (2011) Response of exotic barley germplasm to low-moisture stressed environments. pp 57–64. In: Mulatu B, Grando S (eds) Barley Research and Development in Ethiopia. Proceedings of the 2nd National Barley Research and Development Review Workshop. 28–30 November 2006, HARC, Holetta, Ethiopia. ICARDA, PO Box 5466, Aleppo, Syria. pp xiv + 391
Sintayehu D, Tesfahun A, Bedada G (2010) Stability analysis and factors contributing to genotype by environment interactions in barley for low-moisture stress areas. Pp 250–259. In: Ceccarelli S, Grando S (eds) Proceedings of the 10th International Barley Genetics Symposium, 5–10 April 2008, Alexandria, Egypt. ICARDA, PO Box 5466, Aleppo, Syria. pp xii + 794
Tamene T, Gemechu K, Tadese S, Mussa J (2014) Yield stability and relationships among stability parameters in faba bean (Vicia faba L.) genotypes. Crop J 3:258–268
Verma RPS, Kharab AS, Kumar V, Verma A (2016) G×E evaluation of salinity tolerant barley genotype by AMMI model. Agric Sci Digest 36(3):191–196
Wondimu F, Berhane L (2016) Multi-environment evaluation of malt barley (Hordeum vulgare L.) genotypes in the central highlands of Ethiopia. Critical reviews in crop science. Wudpecker J 1(1):1–9
Wondimu F, Habtamu Z, Amsalu A (2011) Genetic improvement in grain yield potential and associated traits of food barley (Hordeum vulgare L.) in Ethiopia. Appl Sci Techno 2(2):43–60
Yan W (2001) GGE biplot- a window application for graphical analysis of multi environment trial data and other types of two-way data. Agron J 93:1111–1118
Yan W, Kang MS (2003) GGE Biplot analysis: a graphical tool for breeders, geneticists and agronomists, 1st edn. CRC Press LLC, Boca Raton
Yan W, Rajcan I (2002) Biplots analysis of the test sites and trait relations of soybean. Crop Sci 42:11–20
Yan W, Tinker NA (2006) Biplot analysis of multi environment trial data: principles and application. Can J Plant Sci 86:623–645
Yan W, Hunt LA, Sheng Q, Szlavnics Z (2000) Cultivar evaluation and mega environment investigation based on the GGE biplot. Crop Sci 40:597–605
Yan W, Cornelius PL, Cross J, Hunt LA (2001) Biplot analysis of multi environment trial data. Crop Sci 41:656–663
Yan W, Kang MS, Ma M, Woods S, Cornelious PL (2007) GGE Biplot vs AMMI analysis of genotype-by-environment data. Crop Sci 47:641–653
Zemede A (2000) The barley of Ethiopia. pp 77–108. In: Stephen BB (ed). Genes in the Field: On Farm Conservation of Crop Diversity. IDRC/IPGRI
Zerihun J (2011) GGE biplot analysis of multi environment yield trial of barley genotypes in Southeastern Ethiopian highlands. Int J Plant Breed Genet 5(1):59–75
Zobel RM, Wright J, Gauch HG (1988) Statistical analysis of a yield trial. Agron J 80:388–393
Acknowledgements
The authors are grateful for the financial and technical support provided by Ethiopian Institute of Agricultural Research through AGP-II, Haramaya University and Hohenheim University. Our special gratitude also goes to Ethiopian Biodiversity Institute and Holeta Agricultural Research Center for technical and material support. Moreover, breeding staff of respective testing locations are duly acknowledged for assisting the implementation the experiment and data collection.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Fekadu, W., Mekbib, F., Lakew, B. et al. Genotype × environment interaction and yield stability in barley (Hordeum vulgare L.) genotypes in the central highland of Ethiopia. J. Crop Sci. Biotechnol. 26, 119–133 (2023). https://doi.org/10.1007/s12892-022-00166-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12892-022-00166-0