
Abstract
Soil organic components are important factors in the quality and productivity of forest ecosystems. Timber harvesting reduces plant cover and the amount of organic matter in forest floor layer, increases surface runoff, soil erosion and alters microclimatic conditions over large areas. These changes can have important implications for nutrient cycling dynamics and soil organic matter mineralization. Fueguian temperate forests of Nothofagus pumilio (lenga) have been intensely harvested for the last decades, mainly by shelterwood-cut silvicultural system. Harvesting removes nutrients contained in logs from the site, modifies light, temperature and soil humidity, constraining nutrient cycling process. In this study, we evaluate available copper (Cua), zinc (Zna), iron (Fea), and manganese (Mna) concentrations and reservoirs in stands that represent a chronosequence and their respective primary forests (controls): stands cut 1 year ago, stands cut 5–10 years ago, and stands harvested more than 50 years ago. Concentrations of Zna and Cua in primary forest were 39.9 and 2.6 mg/kg, and increased in harvested sites to 60.5 and 3.2 mg/kg, respectively. Fea and Mna concentrations showed similar ranges in both harvested and control sites. Recent harvested sites showed the highest Cua concentrations. Micronutrient reservoirs showed similar ranges in both harvested and primary forests. We concluded that micronutrient availability changes at short term after forest harvesting; thus, the inclusion of soil fertility assessment in forest management plans should be incorporated to preserve the fertility of lenga forests soils and ensure sustainability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The importance of micronutrients for the suitable development and growth of forest trees has been widely recognized (Stone 1968), but these elements have received little attention compared with macroelements. At plant level, micronutrients are required for normal growth and development as they are important for biosynthesis of proteins, nucleic acids, chlorophyll and secondary metabolites, growth substances and stress tolerance (Rengel 2007).
Available micronutrient levels in the medium term play a very important role in soil fertility of forests as they are associated with organic matter (Sahuquillo et al. 2003), as its progressive decomposition increases micronutrient availability. Micronutrients, as trace elements, are present in soil in a variety of forms (1) as free ions and complexes in soil solution, (2) as nonspecifically and specifically adsorbed ions, (3) as ions occluded in soil hydrous oxides and carbonates, (4) organically bound in microbial and plant biomass, detritus, and humic substances, (5) substituted in aluminum–silicate minerals, and (6) as precipitates (Martens and Lindsay 1990). The complex of biogeochemical processes that controls the distribution and sustainability of mineral-soil nutrients includes: recycling processes, such as litterfall, root turnover, canopy leaching, organic matter decomposition, or removal processes, such as plant-root uptake and harvesting (Rengel 2007; Li et al. 2008).
Each nutrient element is characterized by a unique biogeochemical cycle. Extractable zinc (Zn) and copper (Cu) are uniformly distributed in soil profile and both reflect little changes during forest growth (Boardman and McGuire 1990). These elements, as iron (Fe), are mainly distributed in small roots of tree biomass of deciduous species, while manganese (Mn) is mainly located in leaves (Fortescue and Marten 1979). Once the tree cover is removed, the nutrient cycling mechanisms which sustained the soil fertility are disrupted, dropping to levels unable to sustain even a marginal level of productivity. The information on micronutrients cycling in forest ecosystems is not enough compared with that available either for cultivated plants or for macronutrients in forest plants. This lack of information is noteworthy in harvested forests (Imbert et al. 2004).
Nothofagus pumilio, commonly called lenga, is a deciduous and cold-tolerant species with a relatively shallow rooting system. Lenga forest is found from sea level to high elevations (tree line approximately 600–700 m) in southern Patagonia (Arroyo et al. 1996), constituting part of the world’s most austral forests and reaching up to latitudes about 55°S. Lenga forests in Tierra del Fuego and southern Patagonia have been exploited since the beginning of European colonization (Gea-Izquierdo et al. 2004). Currently, shelterwood-cut silvicultural system is the most widespread method implemented in mature primary forests (Martínez Pastur et al. 2000). This method involves the gradual removal of the canopy by subsequent partial timber cuttings; after the first cut, 50 % of the basal area remains. This method results in regular and even-aged managed forests (Schmidt and Urzúa 1982), which are unable to be used for other uses, such as livestock. The state of the art related to the consequences of local forest management has been focused in species diversity (Deferrari et al. 2001), changes in microclimate patterns (Ibarra et al. 2011) and plant litter decomposition (Caldentey et al. 2001; Mansilla 2013; Oro 2014); to our knowledge there are no studies that address aspects related to soil erosion or change in water regime.
Soils of N. pumilio primary forests of Tierra del Fuego have a preferential accumulation of medium-term available Fe and Cu in A horizons, whereas medium-term available Mn and Zn were mainly stored in O horizons (Peña-Rodríguez et al. 2013). Therefore, it is expected that tree removal changes micronutrients availability in these horizons. Previous work has shown that Fe, Mn, Zn, and B concentrations in litter layer of forest floor varied in shelterwood-cut N. pumilio forests (Caldentey et al. 2001). However, to our knowledge soil micronutrient response to shelterwood-cutting of N. pumilio forest has not been reported. The role of nutrients gains special relevance for ecosystem function and stability in those environments where conditions limit forest vegetation development. Thus, this study was conducted to quantify available micronutrients (Fe, Mn, Zn, and Cu) present in the uppermost 10 cm of soil of N. pumilio forest after shelterwood-cutting. In addition, the effects of time lapsed after logging on soil micronutrient concentrations and reservoirs were also assessed.
Materials and methods
Study sites
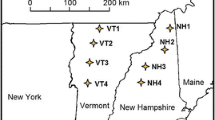
The study was conducted in monospecific N. pumilio forests located in the central part of Tierra del Fuego island, Argentina (54°51′S, 67°35′W) (Fig. 1). In 2009, nine stands with different years after intervention (YAI) by shelterwood-cut (here in after harvested forest or HF) were selected. Thus, there were three stands for each period lapsed after intervention (1, 5–10 and more than 50 years). Likewise, nine unharvested old growth forests (OGF) were selected nearby each harvested forest. These old growth forests are composed of trees with similar diameters at breast height and dominant height, corresponding to sites of quality II according to Martínez Pastur et al. (1997). Therefore, the experimental design consisted of a 2 × 3 factorial experiment where two treatments were considered: forest practices (HF and OGF), and years after intervention (1, 5–10 and >50 years). We took three replicates for each situation, totalizing N = 18 sites.

Location of the study sites in Southern Patagonia. Different colors indicate different years after intervention (YAI)
The climate of the region is characterized by short and cool summers, whereas winters are long showing frequently snow and frost. The average monthly temperature varies between −3 and −4 °C in winter and 9 °C in summer. Precipitation ranges from 400 to 500 mm/year, evenly distributed throughout the year (Iturraspe et al. 1989; Tuhkanen et al. 1989–1990). The forest soils in the study area are characterized by a surface layer of litter almost 2 cm thick (O horizon), followed by a root zone of less than 40 cm (mostly A horizon), with a large proportion of stony material (Contreras et al. 1975). The forest soils in this area are usually slightly acid (pH ranging from 5.5 to 6.0 in top horizons), and notably enriched in organic C especially in the O horizons where normally ranges from 10 to 38 % (Nóvoa-Muñoz et al. 2008).
Soil sampling and processing
In autumn 2010, five composite samples (each after comprising four subsamples) of the top 10 cm of the forest soil were collected in each selected stands. After collection, composite soil samples were stored in plastic bags and transported to the laboratory where they were air-dried and then sieved by 2 mm mesh to obtain the fine-earth soil fraction. Soil samples (<2 mm) were used to determine available Fe (Fea), Mn (Mna), Zn (Zna), and Cu (Cua) which were extracted with a solution of 0.02 M Na2-EDTA + 0.5 M NH4OAc at pH 4.6 (Lakenen and Ervio 1971) using a soil:solution ratio 1:5 and a shaking time of 30 min. The extracts were centrifuged 15 min at 2800 g and filtered with a 0.45 µm pore size fiberglass filters to obtain a clear supernatant. Finally, available concentrations of micronutrients in the centrifuged and filtered extracts were determined by flame atomic absorption spectrometry, expressing the results as amount of micronutrient (mg) per soil mass (kg).
Medium-term reservoirs of soil micronutrients were calculated using the concentrations of available Fe, Mn, Cu, and Zn, soil bulk density and horizon thickness, being expressed in mass unit (Mg) per unit area (ha).
Data analysis
To evaluate the effect of the treatments on soil Fea, Mna, Zna, and Cua concentrations and reservoirs, we used two-way ANOVA, with forest practices and years after intervention (temporal variation) as main factors. To evaluate spatial variation we tested micronutrient differences between old growth forests using one-way ANOVA. Tukey tests were performed to test differences among factors or sites when F values were significant (P < 0.05). All these analyses were carried out with InfoStat v2014 (Di Rienzo et al. 2014).
Results and discussion
The concentration of available micronutrients in forest soils follows the sequence Fea > Mna > > Zna > > Cua (Fig. 2a–d). The same sequence had been previously reported for N. pumilio forest, both the soil (Peña-Rodríguez et al. 2013) (horizons O and A) and the litter layer of forest floor (Caldentey et al. 2001).

Mean values (±standard error) of available concentrations micronutrients (Fea, Mna, Zna, and Cua) in the uppermost 10 cm of soils from N. pumilio forests intervened by shelterwood-cut (harvested forest—HF) and old growth forests (OGF) with different years after intervention
Available concentrations of Fea and Mna showed the greatest variability, with values between 751–1015 mg/kg and 403–1016 mg/kg, respectively (Fig. 2a, b), while Zna and Cua showed less variation (Zn: 28–62 mg/kg; Cu: 2.3–4.4 mg/kg) (Fig. 2c, d). According to Kabata-Pendias (2011), micronutrient values found in this study are indicative of their high availability in forest soils. Similar values were reported by Peña-Rodríguez et al. (2013) and Gerding and Thiers (2002) for forests without intervention in Tierra del Fuego island. These high concentrations may be the result of soil acidity (pH 5.6–6), high organic carbon content (10–38 %), and predominantly fine-textured soils characteristic of the study area (Romanyá et al. 2005, Gerding and Thiers 2002, Nóvoa-Muñoz et al. 2008).
Micronutrient concentrations showed high spatial variability (P < 0.05) when comparing old growth forests (Table 1), which did not differ in their forest structure (Mansilla 2013). Fe and Mn presented the greatest variability (Fe: F = 5.60, P = 0.0014; Mn: F = 5.13, P = 0.0023). This variability may be related to the natural heterogeneity of soils in the area, which is given by their glacial origin (Rabassa and Coronato 2007; Panigatti 2010). Despite this heterogeneity, Fea presented an increased gradient in the EW direction. Furthermore, there are other variables that could control the availability of nutrients in the soils, such as texture, mineral composition, temperature and water regime, among others (Fageria et al. 2002, Kabata-Pendias 2004).
Available Fe and Mn concentrations of the forest floor were not affected by the forest practices or by years after intervention (P > 0.05, Table 2). However, significant differences were found in Mn concentrations between years after intervention (P = 0.069). In the present study, a clear difference was observed particularly for Mn in sites with 5–10 years after intervention, being the availability of this nutrient at harvested forests higher than controls (Fig. 2a, b; Table 2). Although the solubility of Mn always increases with the increase of soil acidity (Rengel 2007), high values of Mn found along with higher pH values in harvested sites compared to controls (Mansilla 2013) could be related to the ability of Mn to form anionic complexes and to complex with organic ligands. This situation could have been contributed to the increment of Mn solubility in soils with alkaline pH range (Kabata-Pendias 2011). In this sense, Caldentey et al. (2001) reported that shelterwood-cut silvicultural systems produce an increment in Mn levels and a decrease in Fe concentrations in L layer of the forest floor. The difference in micronutrient pools reflects how these nutrients are recycling in harvested vs control sites. Additionally it indicates the demand for these elements in relation to return to soil and uptake by young lenga trees at both sites. This also would imply that those differences could be manifested in F and H layers of the forest floor through the course of the time.
Available Zn concentration in harvested forests was increased by over 65 % compared to control forests (F = 6.39, P = 0.014). No differences were observed when considering years after intervention (Fig. 2c; Table 2). Zn is a micronutrient with relatively high mobility in the Earth’s surface and its cycling may be increased by plant and organic debris accumulation, and agricultural practices (Huang and Jin 2008). This characteristic could explain the higher Zn concentration found in harvested sites. These results agree with those found by Caldentey et al. (2001).
Available Cu concentration was the only micronutrient that showed significant interaction (F = 4.43, P = 0.017) between treatments (Fig. 2d; Table 2). In contrast to Caldentey et al. (2001) that found no differences between control and harvested forests, in this study available Cu content showed an increase of 69 % in soils harvested 1 year ago compared to the control (F = 9.60, P = 0.0069) and a rise up to 48 % in the other sites with more years after intervention (F = 9.74, P = 0.0008). High affinity between Cu and organic matter indicates that most of the available Cu is complexed with the soil organic matter (Kabata-Pendias 2011). It is known that this high affinity reduces the rate of mineralization (Parat et al. 2002) favoring humidification and subsequent accumulation in surface soil levels. The highest values of C in soils recorded for sites of 1 year after intervention are consistent with these results (Mansilla 2013).
Similar to available micronutrient concentrations, medium-term reservoirs of soil followed the sequence Fe > Mn > > Zn > Cu, for both harvested and control forests (Fig. 3a–d). Available Fe reservoirs showed a mean value of 0.33 Mg/ha, with a minimum of 0.02 Mg/ha in controls of 5–10 years, and a maximum of 0.73 Mg/ha in controls of forest with more than 50 years after intervention (Fig. 3a). The average value of available Mn reservoir was 0.22 Mg/ha; with a range between 0.02 and 0.95 Mg/ha, both values recorded for harvested forests (Fig. 3b). In the case of Zn, available reservoir averaged 0.018 Mg/ha, with values between 0.0017 and 0.070 Mg/ha; both registered in harvested sites with more than 50 years after intervention (Fig. 3c). Available Cu showed an average value of 0.0010 Mg/ha ranging between 0.0001 and 0.0022 Mg/ha (Fig. 3d). Similar values of Cu were recorded by Peña-Rodríguez et al. (2013) for O horizons; however, reservoirs of the other micronutrients were significantly lower. The great variability shown in the case of Zn agrees with Peña Rodríguez et al. (2013).

Mean values (±standard error) of micronutrients reservoirs (Fe, Mn, Zn, and Cu) in the uppermost 10 cm of soils from N. pumilio forests intervened by shelterwood-cut (harvested forest—HF) and old growth forests (OGF) with different years after intervention
None of the studied reservoirs was affected by considered treatments (Table 3), with the exception of Zn. Harvested sites showed higher Zn values respect to the control (F = 4.16, P = 0.047), according to the reported concentration. Similar results were obtained for the reservoir layer of the forest floor litter in forests of N. pumilio recently managed (Caldentey et al. 2001). The contribution of litter with different quality received by each site could play a major role in micronutrients recycling (Mansilla et al. 2009). In deciduous forest ecosystems, Gallardo et al. (1998) maintained that greater proportion of Zn returned to the soil through canopy leaching.
Although the differences between years after intervention for most micronutrients were not relevant, it is remarkable that the differences of Fe and Mn concentrations between harvested and controls sites were greater at sites of 5–10 years after intervention. However, the differences observed for Zn and Cu were in the treatment with 1 year since intervention. Data dispersion may have masked the differences between treatments, so it would be important to consider for future studies a larger number of samples because of the great natural variability of fueguian soils. Because control sites are nearby harvested sites, found differences between them are due to forest management. This causes a series of changes in forest biotic (Martínez Pastur et al. 2000; Sparagino et al. 2001; Lencinas et al. 2009) and abiotic factors (Promis et al. 2010; Martínez Pastur et al. 2011), which could affect the dynamics of soil microorganisms involved in micronutrients recycling, water availability, soil compaction, etc.
Trace elements are defined as elements that are present at low concentrations (mg/Kg or less) in most soils, plants, and living organisms (Phipps 1981). Therefore, higher nutrient concentrations observed in harvested sites may cause toxicity problems in plants as well as in other components of the forest ecosystem. In contrast to agroecosystems, where if more trace elements are introduced than those naturally present, they are accumulated in landscapes over long periods and may eventually be released to surface and ground waters, affecting both aquifers and aquatic ecosystems (Zhenli et al. 2005), the effect of intervention on micronutrient concentration is poorly known. For this reason, future research needs to focus on the balance of micronutrients in forest ecosystems, as well as on quantification micronutrients transport from forest ecosystems to the whole environment. Since these elements are essential for plants, it is necessary to quantify them because they are required for the regeneration of the southernmost forests of the world.
Conclusions
These results are the first work about the dynamics of soil micronutrients, in terms of concentration and reservoir, present in N. pumilio harvested forests of Tierra del Fuego. It concludes that micronutrient availability changes at short term after forest harvesting; thus, the inclusion of soil fertility assessment in forest management plans should be incorporated to preserve the fertility of N. pumilio forest soils, assuring in this way forest sustainability.
References
Arroyo MTK, Donoso C, Murúa RE, Pisano EE, Schlatter RP, Serey IA (1996) Toward an ecologically sustainable forestry project. Concepts, analysis and recommendations. Departamento de Investigación y Desarrollo, Universidad de Chile, Santiago
Boardman R, McGuire DO (1990) The role of zinc in forestry. I. Zinc in forest environments, ecosystems and tree nutrition. For Ecol Manage. doi:10.1016/0378-1127(90)90054-F
Caldentey J, Ibarra M, Hernández J (2001) Litter fluxes and decomposition in Nothofagus pumilio stands in the region of Magallanes Chile. For Ecol Manage. doi:10.1016/S0378-1127(00)00532-6
Contreras H, Borgel R, Quezada M, García de Cortázar V, Rojas M, Bitterlich W (1975) Informe de la primera etapa del proyecto sobre reforestación de la Precordillera Patagónica (Cuadrángulos Skyring y Rubens). Facultad de Ciencias Forestales, Universidad de Chile, Santiago p 76
Deferrari G, Camilion C, Martínez Pastur G, Peri P (2001) Changes in Nothofagus pumilio forest biodiversity during the forest management cycle: 2. Biodiv Conserv, Birds. doi:10.1023/A:1013154824917
Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW (2014) InfoStat versión 2014. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. http://www.infostat.com.ar
Fageria NK, Baligar VC, Clark RB (2002) Micronutrients in crop production. Adv Agron. doi:10.1016/S0065-2113(02)77015-6
Fortescue JAC, Marten GG (1979) Micronutrients: forest ecology and systems analysis. In: Reichle DE (ed). Analysis of temperate forest ecosystems 173–198. Springer-Verlag, New York p 304
Gallardo JF, Martin A, Santa Regina I (1998) Nutrient cycling in deciduous forest ecosystems of the Sierra de Gata Mountains: nutrient supplies to soil through both litter and throughfall. Ann Sci For 55:771–784. doi:10.1051/forest:19980701
Gea-Izquierdo G, Martínez Pastur G, Cellini JM, Lencinas MV (2004) Forty years of silvicultural management in southern Nothofagus pumilio (Poepp. Et Endl.) Krasser primary forests. For Ecol Manage. doi:10.1016/j.foreco.2004.07.015
Gerding V, Thiers O (2002) Caracterización de suelos bajo bosques de Nothofagus betuloides (Mirb) Blume, en Tierra del Fuego (Chile). Rev Chil Hist Nat. doi:10.4067/S0716-078X2002000400015
He ZL, Yang XE, Stoffella PJ (2005) Trace elements in agroecosystems and impacts on the environment. J Trace Elem Med Biol. doi:10.1016/j.jtemb.2005.02.010
Huang SW, Jin JY (2008) Status of heavy metals in agricultural soils as affected by different patterns of land use. Environ Monit Assess 139:317–327
Ibarra M, Caldentey J, Promis A (2011) Descomposición de hojarasca en rodales de Nothofagus pumilio de la región de Magallanes. Bosque. doi:10.4067/S0717-92002011000300004
Imbert JB, Blanco JA, Castillo FJ (2004) Gestión forestal y ciclos de nutrientes en el marco del cambio global. In: Valladares F (ed) Ecología del bosque mediterráneo en un mundo cambiante. Ministerio de Medio Ambiente, Madrid, pp 479–506
Iturraspe R, Sottini R, Schroeder C, Escobar J (1989) Generación de información hidroclimática en Tierra del Fuego. Hidrología y Variables Climáticas del Territorio de Tierra del Fuego, Información básica. CONICET-CADIC, Ushuaia, Argentina. Contr Cient 7:1–196
Kabata-Pendias A (2004) Soil-plant transfer of trace elements-an environmental issue. Geoderma. doi:10.1016/j.geoderma.2004.01.004
Kabata-Pendias A (2011) Trace elements in soils and plants, 4th edn. CRC, Boca Raton, p 520
Lakenen E, Ervio RA (1971) A comparison of eight extractants for the determination of plant-available micronutrients in soils. Acta Agralia Fenn 123:223–232
Lencinas MV, Martínez Pastur G, Gallo E, Cellini JM (2009) Alternative silvicultural practices with variable retention improve bird conservation in managed South Patagonian forests. Ecol Manage, For. doi:10.1016/j.foreco.2009.01.012
Li J, Richter DD, Mendoza A, Heine P (2008) Four-decade responses of soil trace elements to an aggrading old-field forest: B, Mn, Zn, Cu, and Fe. Ecology. doi:10.1890/07-1381.1
Mansilla PR (2013) Evaluación de la alteración y restauración de los ciclos biogeoquímicos en bosques de Nothofagus pumilio de Tierra del Fuego afectados por prácticas de manejo forestal. Ph.D. thesis in Biological sciences, Universidad de Buenos Aires, Argentina, p 150
Mansilla R, Moretto A, Pancotto V, Lencinas V, Escobar J, Vrsalovic J, Oro N (2009) Producción de hojarasca en los bosques intervenidos de Nothofagus pumilio de Tierra del Fuego. ¿Es posible su recuperación? Boletín de la Sociedad Argentina de Botánica, XXXII Jornadas Argentinas de Botánica, Resumem p 92
Martens DC, Lindsay WL (1990) Testing soils for copper, iron, manganese, and zinc. In: RL. Westerman (ed). Soil testing and plant analysis, 3rd edn. Soil Science of Society of America, Madison, p 229–264
Martínez Pastur G, Peri P, Vukasovic R, Vaccaro S, Piriz Carrillo V (1997) Site index equation for Nothofagus pumilio Patagonian forests. Phyton 6(1/2):55–60
Martínez Pastur G, Cellini JM, Peri P, Vukasovic R, Fernández MC (2000) Timber production of Nothofagus pumilio forests by a shelterwood system in Tierra del Fuego (Argentina). Ecol Manage, For. doi:10.1016/S0378-1127(99)00253-4
Martínez Pastur G, Cellini JM, Lencinas MV, Barrera M, Peri PL (2011) Environmental variables influencing regeneration of Nothofagus pumilio in a system with combined aggregated and dispersed retention. Ecol Manage, For. doi:10.1016/j.foreco.2010.10.002
Nóvoa-Muñoz JC, Pontevedra-Pombal X, Moretto A, Peña S, Escobar J, García-Rodeja Gayoso E (2008) Caracterización geoquímica de suelos forestales de lenga en el ecotono de Tierra del Fuego (Argentina). Resúmenes del XXI Congreso Argentino de la Ciencia del Suelo. Barbosa OA. (ed). Asociación Argentina de la Ciencia del Suelo. Buenos Aires, p 648
Oro N (2014) ¿Cómo varían los ciclos biogeoquímicos debido al aprovechamiento forestal en bosques de Nothofagus pumilio de Tierra del Fuego? Ph.D. thesis in Biology, Universidad Nacional del Sur, Argentina, p 167
Panigatti JL (2010) Argentina 200 años, 200 suelos. Ed. INTA Buenos Aires, p 345
Parat C, Chaussod R, Léveque J, Dousset S, Andreux F (2002) The relationship between copper accumulated in vineyard calcareous soils and soil organic matter and iron. Eur J Soil Sci 53:663–669
Peña-Rodríguez S, Moretto M, Pontevedra-Pombal X, Oro N, García-Rodeja Gayoso E, Rodríguez-Salgado I, Rodríguez-Racedo J, Escobar J, Nóvoa-Muñoz JC (2013) Trends in nutrient reservoirs stored in uppermost soil horizons of subantarctic forests differing in their structure. Syst, Agrofor. doi:10.1007/s10457-013-9635-8)
Phipps DA (1981) Chemistry and biochemistry of trace metals in biological systems. In: Lepp NW (ed) Effect of heavy metal pollution on plants: effects of trace metals on plant function, vol I. Applied Sci Publ, London, pp 1–54
Promis A, Caldentey J, Ibarra M (2010) Microclima en el interior de un bosque de Nothofagus pumilio y el efecto de una corta de regeneración. Bosque. doi:10.4067/S0717-92002010000200006
Rabassa J, Coronato A (2007) Glaciaciones del Cenozoico tardío en los Andes Patagónico–Fueguinos. En Patagonia total Antártida e Islas Malvinas. Ed. Alfa Centro Literario–Patagonia Argentina, p 1088
Rengel Z (2007) Cycling of Micronutrients in Terrestrial Ecosystems. In: Marschner P, Rengel Z (Eds) Soil Biology, Nutrient Cycling in Terrestrial Ecosystems, Vol 10. Springer-Verlag, Berlin, p 93–121
Romanyà J, Fons J, Sauras-Yera T, Gutiérrez E, Vallejo VR (2005) Soil–plant relationships and tree distribution in old growth Nothofagus betuloides and Nothofagus pumilio forests of Tierra del Fuego. Geoderma 124:169–180
Sahuquillo A, Rigol A, Rauret G (2003) Overview of the use of leaching/extraction tests for the risk assessment of trace metals in contamined soils and sediments. Chem, Trends Anal. doi:10.1016/S0165-9936(03)00303-0
Schmidt H, Urzúa A (1982) Transformación y manejo de los bosques de Lenga en Magallanes. Univ de Chile Cienc Agríc 11:62
Spagarino C, Martínez Pastur G, Peri PL (2001) Changes in Nothofagus pumilio forest biodiversity during the forest management cycle: insects. Biodivers Conserv 10(12):2077–2092
Stone EL (1968) Boron deficiency and excess in forest trees: a review. Ecol Manage, For. doi:10.1016/0378-1127(90)90046-E
Tuhkanen S, Ikuokka I, Hyvonen N, Stenroons S, Niemela J (1989–1990) Tierra del Fuego as a target for biogeographical research in the past and present. Anales del Instituto de la Patagonia 19(2):107
Acknowledgments
This research was funded by Agencia Nacional de Promoción Científica y Técnica Project (PICTO FORESTAL 36861) and by CIA3 which was granted by FEDER founds through the Program of Consolidation and Arrangement of Research Units from Consellería de Educación (Xunta de Galicia). The authors gratefully thank to the Centro Austral de Investigaciones Científicas, Ea. Los Cerros, Ea. Ushuaia, Ea. Ewan, Ea. María Cristina and Dirección de Bosques of Tierra del Fuego for their support during the realization of this work. We also thank to Soledad Diodato for their helpful comments.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is part of a Topical Collection in Environmental Earth Sciences on “3RAGSU”, guest edited by Daniel Emilio Martinez.
Rights and permissions
About this article

Cite this article
Mansilla, R., Nóvoa-Muñoz, J.C., Pontevedra-Pombal, X. et al. Temporal and spatial changes in soil micronutrients in managed Nothofagus pumilio forest of Tierra del Fuego, Argentina. Environ Earth Sci 75, 738 (2016). https://doi.org/10.1007/s12665-016-5470-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12665-016-5470-3