
Abstract
Caffeine is the world’s most popular psychoactive drug and is also an active adulterant found in many drugs of abuse, including seized cocaine samples. Despite several studies which examine the effects of caffeine or cocaine administered as single agents, little data are available for these agents when given in combination. The purpose of the present study was to determine if combined intake of both psychostimulants can lead to maladaptive changes in striatal function. Mice were injected with a binge regimen (intermittent treatment for 13 days) of caffeine (3 × 5 mg/kg), cocaine (3 × 10 mg/kg), or combined administration. We found that chronic caffeine potentiated locomotion induced by cocaine and that both caffeine-treated groups showed sensitization. Striatal tissue was obtained 24 h and 7 days after last injection (withdrawal) for immunohistochemistry and mRNA expression. Our results show that combined intake of both psychostimulants can increase GFAP immunoreactivity in the striatum at both times post treatment. Gene expression analysis, targeted at dopamine, adenosine, and glutamate receptor subunit genes, revealed significant transcript down-regulation in the dorsal striatum of AMPA, NMDA, D1 and D2 receptor subunit mRNA expression in the group that received combined treatment, but not after individual administration. At withdrawal, we found increased D1 receptor mRNA expression along with increased A1, AMPA, NMDA, and metabotropic subunit expression. A2A mRNA showed decreased expression after both times in all experimental groups. Our study provides evidence that there are striatal alterations mediated by combined caffeine and cocaine administration, and highlights negative outcomes of chronic intake of both psychostimulants.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The methylxanthine caffeine is the world’s most popular psychoactive drug. In recreational drug use settings, caffeine is found in a range of commercially available products such as energy drinks that may be consumed with other drugs (Reissig et al. 2009). The primary effect of caffeine is to relieve fatigue and enhance mental performance, but excessive ingestion might lead to intoxication and withdrawal symptoms if intake is discontinued (Dews et al. 2002). DSM-IV did not include caffeine dependence as a substance use disorder but caffeine withdrawal was included as a research diagnosis to encourage investigation (Hasin et al. 2013).
Apart from analyzing the potential clinical importance of caffeine by itself, another aspect that deserves consideration is its interactions with other addictive substances (Ferré 2013). It needs to be noted that forensic analyses of seized illicit drug samples have reported quantities of caffeine mixed with other stimulants, including cocaine and amphetamines (Cole et al. 2011). As an example relevant to the objectives of the present study, cocaine paste which is an intermediate product of the cocaine alkaloid extraction process from coca leaves has a high content of cocaine base mixed with other chemical substances and it is typically adulterated with caffeine (López-Hill et al. 2011).
Several preclinical and clinical studies have suggested that caffeine can augment the reinforcing (Schenk et al. 1994; Kuzmin et al. 1999), motor stimulant (Gasior et al. 2000), and subjective effects (Jones and Griffiths 2003, Liguori et al. 1997) of drugs of abuse, with potential implications for human drug users. It is also likely that concomitant consumption of caffeine with other stimulant drugs such as cocaine can profoundly alter the drug response and induce adverse interactive effects, both on the SNC and on the other organs. Our group has recently described that caffeine and cocaine administration can produce testicular toxicity and that caffeine can potentiate some of the detrimental changes induced by cocaine on testicular tissue (Gonzalez et al. 2015). Previous reports in animal models showed that high doses of caffeine are required to amplify the toxicity of d-amphetamine and cocaine as reported by Derlet et al. (1992), while lower doses of caffeine are sufficient to promote toxicity and lethality when combined with MDMA or MDA (McNamara et al. 2006; Vanattou-Saïfoudine et al. 2012). Also, caffeine potentiated the toxic effect of methamphetamine in the striatum (Sinchai et al. 2011), a brain structure that is implicated in psychostimulant-induced neuroplasticity (Kalivas and O’Brien 2008). Furthermore, glial fibrillary acidic protein (GFAP), a cytoskeletal intermediate filament protein exclusively expressed in astrocytes (Bignami et al. 1972), has been previously demonstrated to be responsive to cocaine administration (Freeman et al. 2010; Blanco-Calvo et al. 2014). Little is known about glial responses evoked by chronic caffeine treatment.
The striatum receives synaptic inputs from many different sources. Glutamatergic afferents arrive from areas of the cortex and the thalamus, whereas the nigrostriatal pathway and intrinsic circuits provide the striatum with acetylcholine, GABA, dopamine (DA), nitric oxide, and adenosine (A) (Centonze et al. 2003, Ferré et al. 2010). The A1 receptor (A1R) and A2A receptor (A2AR) are primarily responsible for the central effects of adenosine (Dunwiddie and Masino 2001). A2AR are more concentrated in the striatum than anywhere else in the brain, while A1R are more widespread distributed (Fredholm et al. 2001; Rosin et al. 2003). The same receptors are also the main target of moderate, nontoxic doses of caffeine that acts as A1R and A2AR antagonist (Fredholm et al. 1999). In addition, caffeine is a well-known antagonist of inositol trisphosphate (IP3) receptors and an agonist of ryanodine receptors (RyRs) (McPherson et al. 1991). At the neuronal level, caffeine is capable of changing intracellular [Ca2+] levels through ryanodine intracellular stores (Garaschuk et al. 1997). On the other hand, cocaine blocks all three neuronal membrane monoamine transporters with approximately equal affinity. By blocking the dopamine transporter (DAT), cocaine allows released DA to persist in the extracellular space, extending DA receptor activation leading to behavioral activation (Volkow et al. 2012).
Despite several studies which examined caffeine or cocaine as single agents, little data are available for these agents given in combination. Complex interactions have now been clearly shown for A1R and dopamine receptor 1 (D1R) and A2AR and dopamine receptor 2 (D2R) in the striatum (Ferré et al. 2010). In fact, there are indications of the involvement of A2AR and dopamine receptors in the locomotor and sensitizing effects of cocaine (Filip et al. 2006). Moreover, a recent study suggested that caffeine might enhance DA signaling in the striatum by increasing D2/D3R levels or their affinity in the human brain (Volkow et al. 2015).
Several studies reported detrimental effects of caffeine when combined with other drugs of abuse (i.e., co-administration of caffeine with methamphetamine or MDMA). However, information is lacking on the impact of caffeine combined with cocaine on striatal function. Therefore, the purpose of the present study was to determine if combined intake of both psychostimulants leads to maladaptive changes and/or additive effects in the striatum. Particularly, neuroinflammation of the central nervous system (CNS) seems to participate in neuroplasticity changes and sensitizing effects of psychostimulants (Cadet and Bisagno 2014). Thus, in the present study treated mice were behaviorally tested for acute locomotor effects and sensitization after chronic intermittent treatment with caffeine, cocaine, or the combination of both psychostimulants. As mentioned before, information is scarce regarding potential neuroplastic changes involving striatal neuronal and glial cells produced by combined chronic treatment of caffeine and cocaine. Additional information is needed since striatal neuroadaptations could underlie some of the behavioral changes associated with chronic drug use (Cadet and Bisagno 2016) affecting cortico-striatal network physiology. Thus, we investigated the effect of psychostimulant treatments on reactive astrogliosis in the striatum at two different time points: (a) 24 h after the last binge injection and (b) 7 days after the last binge injection (withdrawal). Additionally, we measured mRNA expression of several dopaminergic, adenosinergic, and glutamatergic receptor subunits associated to psychostimulant-induced neuroplasticity on striatal tissue (Ferré et al. 2010).
Materials and Methods
Animals
Male C57BL/6 mice (20–25 g) from the School of Exact and Natural Sciences of the University of Buenos Aires were housed in a light- and temperature-controlled room. Mice had free access to food and water. Principles of animal care were followed in accordance with “Guidelines for the Care and Use of Mammals in Neuroscience and Behavioral Research” (National Research Council, 2003), and approved by Universidad de Buenos Aires authorities (Protocol Number: A5801-01) using OLAW and ARENA directives (NIH, Bethesda, USA).
Pharmacological and Physiological Procedures
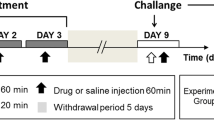
Cocaine hydrochloride and caffeine (Sigma-Aldrich, St Louis, MO) were administered in a binge-like regimen of three i.p. injections per day, 1 h apart. Animals were assigned to four different groups: Coc (Cocaine 3 × 10 mg/kg), Caf (Caffeine 3 × 5 mg/kg), Caf–Coc (3× Cocaine + Caffeine combined solution: Coc 10 mg/kg +Caf 5 mg/kg, both dissolved in the same sterile saline solution and co-administered in a single injection), or Veh (3× sterile saline), see Fig. 1.

Schematic representation of the experimental protocol used in this study. Male C57BL/6 mice were subjected to an intermittent chronic treatment (one day on/one day off for 13 days) of caffeine 5 mg/Kg (Caf), cocaine 10 mg/kg (Coc), and their combination (5 mg/kg Caf + 10 mg/kg Coc) in a binge protocol: 3 i.p. injections per day, 1 h apart. Locomotor activity was evaluated at Day 1 and Day 13 in an open field with 30 min of habituation before the first injection. Tissue samples were taken 1 day and 7 days after the last binge injection (Days 14 and 21, respectively)
The induction of locomotor sensitization depends on the temporal pattern of drug exposure. Repeated intermittent treatments with moderate doses of drugs are more effective for inducing locomotor sensitization than continuous exposure to high or escalating drug doses (Robinson and Berridge 1993; Vanderschuren and Kalivas 2000). Accordingly, we selected an intermittent protocol of psychostimulant administration, i.e., mice received binge injections 1 day on/1 day off during 13 days for a total of 7 binges, see Fig. 1. Striatal tissue for gene and protein expression was obtained 24 h after the last injection (Experiment #1), to investigate the effect of chronic treatment using both psychostimulants. It should be noted that striatal tissue obtained 24 h after the last binge injection likely reflects the consequences of chronic administration of both caffeine and cocaine. Since the mice were subjected to an intermittent chronic treatment (one day on/1 day off for 13 days), it is unlikely that the mice would perceive this time as “withdrawal” (since mice were already used to intermittent injections).
For Experiment #2, we also used the same protocol as Experiment #1, but in this case mice were sacrificed 7 days after last binge injection. At that time, striatal tissue was also collected to investigate the effect of withdrawal from chronic treatment with both psychostimulants.
Behavioral Studies
Locomotor Activity
Mice were housed in plastic cages for at least 1 week prior to the initiation of locomotor experiments. Mice were moved to an experimental room 12 h before drug treatment. Food and water were provided ad libitum in a temperature-controlled room, with a 14/10-h light/dark cycle. Locomotor activity (total distance in cm) was recorded using a CCD camera (Sony, U.S.A.) in custom-designed open-field boxes located in a sound-attenuated room. For acquisition and analysis, we used Ethovision XT 7.0 software (Noldus, The Netherlands). Each box consisted of an open-field compartment (19 × 40 × 40 cm). On injection Days 1 and 13, animals were placed in open-field boxes for 30 min (recorded as baseline), and later injected with drugs or saline. Total distance traveled (in cm) was quantified in 5-min bins for a total of 180 min after the first injection (i.e., 60 min were recorded after each of the three drug injections). Behavioral recordings were made simultaneously in four open-field arenas using the Ethovision XT multiple arena feature from 9 AM to 4 PM of the light period of the light/dark cycle, similar to a previously published study (Bisagno et al. 2010). Injection time and arenas (right and left) were counterbalanced across experimental groups.
Immunohistochemistry for GFAP
Animals were deeply anesthetized 24 h or 7 days after treatments with ketamine–xylazine (0.5 ml/kg i.p.; 1.4 ml/kg i.p., respectively) and then transcardially perfused with 30 ml of phosphate buffer saline (PBS) 10 mM (pH 7.4) followed by 80 ml of ice-cold paraformaldehyde 4 % (Sigma, USA) diluted in 0.1 M (pH 7.4) PBS. Brains were dissected and placed in the same fixative solution for 16 h at 4 °C, and put in PBS/sucrose 30 % for cryoprotection until coronal sections were cut throughout the striatum.
Immunostaining for GFAP was performed on 25-µm free-floating coronal serial slices from striatal sections. During all staining procedures 0.1 M PBS containing 0.3 % Triton X-100 (PBS-T) was used for diluting all immunoreagents and for washing after incubating with antibodies. Sections were incubated in 2 % H2O2 followed by 2 % normal goat serum (NGS) in PBS-T and exposed to a rabbit anti-GFAP antibody (1:500, Sigma, USA) overnight at 4° C. After washing, sections were incubated for 2 h in biotinylated goat antirabbit IgG antiserum (1:250, Sigma, USA) followed by the avidin–biotin peroxidase complex (1:125, Vectastain, ELITE ABC kit, Vector Laboratories). Chromogenic reactions were induced with 0.5 mg/ml DAB and 0.015 % H2O2. Sections were then rinsed, mounted on gelatin-coated slides, air-dried, dehydrated, cleared, and cover slipped.
GFAP-immunoreactive area was quantified as follows: light microscopic examination of microtome sections revealed GFAP-positive cell staining throughout the striatum, with scattered distribution. Data were collected with the help of mapping software (Mercator Pro; Explora Nova, France), coupled to a Nikon microscope. This software has an embedded navigation system that allowed us to draw maps of the entire dorsal striatum (left and right hemispheres) at low magnification (×4), identify a GFAP-stained cell, and then use a higher magnification (×60) to confirm GFAP-positive astrocytes and mark them. At the end of each map quantification, Mercator software displays an excel file with the # of GFAP-positive astrocytes. Cells were quantified on 3 different striatal sections, bilaterally. Data were averaged in order to have a unique estimative count per animal.
Real-Time PCR
Striatal tissue was extracted 24 h or 7 days after treatments: mouse brains were rapidly removed, and striatal tissues were dissected, placed on dry ice, and then stored at −70 °C in RNAlater (Qiagen) until further assays. Total RNA was isolated using TRIZOL reagent (Invitrogen) according to the manufacturer’s protocol. Five hundred nanograms of RNA were treated with DNAseI (Invitrogen) and reverse-transcribed in a 20 μL reaction using M-MLV reverse transcriptase (Promega) and random hexamers (Biodynamics). For quantitative real-time PCR, primer sets were designed for the specific amplification of murine Drd1a, Drd2, Adora1, Adora2a, Gria1, Gria2, Grin1, Grin2a, Grin2b, Grm1, Grm5, and Gapdh as a housekeeping control gene (sequences listed in Table 1). Each sample was assayed in duplicate using 4 pmol of each primer, 1X SYBR Green Master Mix (Applied Biosystems), and 2–20 ng of cDNA in a total volume of 13 μL. Amplification was carried out in an ABI PRISM 7500 Sequence Detection System (Applied Biosystems).
Statistical Analysis
InfoStat 2010 software (www.infostat.com.ar) was used for statistical analysis. Statistics were performed using one-way ANOVA followed by Fisher’s LSD post hoc tests. Data were transformed when required using logarithmic transformations in order to comply with ANOVA assumptions. For the analysis of locomotor sensitization, two-way repeated measures ANOVA followed by Fisher’s LSD was performed. Data are expressed as the mean ± SEM. Differences were considered significant if p < 0.05.
Results
Acute and Chronic Effects of Caffeine, Cocaine, and Their Combination on Locomotor Activity and Sensitization
Locomotor activity was measured after acute (Day 1) and chronic (Day 13) administration of caffeine, cocaine, and the combination of both psychostimulants (administered in binge-like form, i.e., three injections, see protocol on Fig. 1). On experimental Days 1 and 13, locomotor activity was measured in all treated groups for 30 min before the first injection (baseline locomotion), and 60 min after each one of the three injections (for a total of 180 min), see Fig. 2.

Effect of chronic treatment with caffeine, cocaine, and their combination on locomotor activity. a. Acute effect. Left panel locomotor activity induced by the 3 injections of caffeine (Caf), cocaine (Coc), and their combination (Caf–Coc) during binge treatment on Day 1. Right panel cumulative distance traveled (expressed as % baseline) during binge treatment on Day 1 (180 min). b. Chronic effect. Left panel locomotor activity induced by the 3 injections of Caf, Coc, and their combination (Caf–Coc) during binge treatment on Day 13. Right panel cumulative distance traveled (expressed as % baseline) on Day 13 (180 min). c. Basal locomotor activity. Distance traveled during 30-min habituation to the locomotor arena prior to binge treatment on Day 1 and Day 13. Data are expressed as mean ± SEM (n = 14–20). The arrows indicate the time of injection. The values indicate mean ± SEM (N = 14–20). One-way ANOVA–Fisher’s LSD: *p < 0.05, **p < 0.01, ***p < 0.001 different from Veh; $ p < 0.05, $$ p < 0.01, $$$ p < 0.001 different from Coc
Acute effects
As shown in Fig. 2a, caffeine, cocaine, and their combination induced increases in total locomotion after acute administration compared to Veh (Fig. 2a left). Figure 2a right shows accumulated distance traveled throughout Day 1 binge (for a total time of 180 min after the first injection). ANOVA–Fisher’s LSD showed significant differences among treatments (F (3,59) = 136.20, p < 0.0001), where all groups injected with psychostimulants showed increased total locomotion compared to Veh values (p < 0.01). Also, the Coc group showed higher locomotion than the Caf group (p < 0.001), and was not different from the Caf–Coc group (Fig. 2a right).
Chronic effects
As shown in Fig. 2b, after chronic psychostimulant administration, caffeine, cocaine, and their combination induced increases in total locomotion after acute administration compared to Veh (Fig. 2b left). Figure 2B right shows accumulated distance traveled throughout Day 13 binge. ANOVA–Fisher’s LSD also showed significant differences across treatments (F (3,64) = 205.84, p < 0.0001), where all groups injected with psychostimulants showed increased total locomotion compared to Veh values (p < 0.001). Also, the Coc group showed higher locomotion than the Caf group (p < 0.001), and interestingly the group that received the combination of Caf–Coc showed significantly higher values compared to the Coc group (p < 0.05).
We also quantified and analyzed baseline levels of locomotor activity on Days 1 and 13 (measured 30 min before the first injection, Fig. 2c). As expected, baseline locomotion on Day 1 showed no significant differences across treatments. However, at Day 13 ANOVA–Fisher’s LSD showed significant differences across treatments (F (3,60) = 3.95, p < 0.05). Coc and Caf–Coc groups showed higher values of basal locomotion compared to Veh (p < 0.05). Repetitive treatment was administered in the animal’s cages during the protocol with the exception of Days 1 and 13 when injections were done in the open-field cages in order to record behavior. Significant differences on Day 13 observed at baseline might be related to the fact that vehicle-treated animals showed lower baseline values on Day 13 (compared to the first exposure to open-field cages on Day 1). Open-field arenas and procedures on Day 13 might have not been seen as “novelty” for vehicle-treated animals and thus less locomotion was observed in that group.
Sensitization
Locomotor sensitization to psychostimulants was defined as a greater total distance traveled after drug injections on the last administration (Day 13) compared to the first drug exposure (Day 1). Repeated measures two-way ANOVA followed by Fisher’s LSD showed significant treatment (F (3,60) = 71.77, p < 0.0001), and day (F (1,60) = 8.07, p < 0.01) effects, with no significant interaction effects (F (2,60) = 2.05, p > 0.05). As shown in Fig. 3, both the Caf- and Caf–Coc-treated groups showed increased locomotor activity on Day 13 compared to the values observed after the injections on Day 1, suggesting locomotor sensitization.

Effect of chronic treatment with caffeine, cocaine, and their combination on locomotor sensitization. Cumulative distance traveled (expressed as % baseline) on Day 1 and Day 13. The values indicate mean ± SEM (N = 14–20). Two-way repeated measures ANOVA–Fisher’s LSD: # p < 0.05 different from Day 1 of the respective treatment
It should to be noted that our treatment involved intermittent administration of low doses of caffeine in a binge form that was able to show locomotor sensitization. However, no incidence of seizures or death was observed in our study in any experimental group, unlike previous studies involving higher doses of caffeine (100 mg/kg) (Derlet et al. 1992).
Chronic effects of caffeine and cocaine on GFAP immunoreactivity in the striatum
Astrocytes have the ability to change their molecular expression and morphology in response to different CNS insults (Sofroniew 2009). These changes on astrocytes are called reactive astrogliosis and are a hallmark of all CNS pathologies (Sofroniew 2009). Immuno-detection of GFAP, a standard marker for astrogliosis, was analyzed following chronic administration with both psychostimulants. GFAP levels have been previously demonstrated to respond to cocaine exposure (Freeman et al. 2010). However, no reports are available showing cocaine treatment increasing reactive astrogliosis in the dorsal striatum. Moreover, other conditions where other psychostimulants were co-administered with caffeine reported that caffeine can potentiate astrogliosis in different SNC areas (Khairnar et al. 2010; Frau et al. 2015). We have found increased GFAP-positive astrocytes in the dorsal striatum after chronic psychostimulant treatment and after 7 days of withdrawal. Figure 4a shows the effect of chronic treatment with both psychostimulants and their combination. ANOVA–Fisher’s LSD showed significant differences across treatments (F (3,15) = 32.81, p < 0.0001), all psychostimulant-treated groups showed significant increase in GFAP expression (p < 0.05), with the group that received the combination of caffeine and cocaine showing the highest GFAP-positive astrocyte values (p < 0.001). We also measured GFAP expression after withdrawal (Fig. 4b). ANOVA–Fisher’s LSD showed significant differences across treatments (F (3,15) = 9.69, p < 0.01); at this time, all psychostimulant-treated groups also showed increased GFAP immunoreactivity, (p < 0.05), but following drug washout possible additive effects of combined treatment with both psychostimulants were no longer statistically different.

Effect of caffeine, cocaine, and their combination on striatal GFAP immunoreactivity after chronic treatment or withdrawal. The values (number of GFAP-positive cells) indicate mean ± SEM (N = 4) from striatal tissue extracted after chronic treatment a or after a 7-day withdrawal period b. One-way ANOVA–Fisher’s LSD: *p < 0.05, **p < 0.01, ***p < 0.001 different from Veh; $$ p < 0.01 different from Coc. Representative images of striatal sections from animals treated with either Veh, Caf, Coc, or Caf + Coc; scale bar 500 µm, and their respective higher magnification photomicrographs, scale bar 150 µm
It should to be noted that the pattern of GFAP-positive astroglia found in this study was different than the massive GFAP reaction that is usually seen after toxic doses of methamphetamine (Raineri et al. 2012, 2015). This difference in the astrogliosis produced by toxic methamphetamine and cocaine might be related to the different doses assayed in both studies (methamphetamine toxic regime involves repetitive high-dose administration) or to the toxic profile that cocaine or methamphetamine can display. GFAP-positive cells using our protocol of intermittent caffeine and cocaine administration followed a scattered distribution through the striatal tissue, with more activation found in dorsal/medial striatal areas, adjacent to ventricles.
In addition, since we found increased GFAP expression after chronic treatment and withdrawal, we also evaluated the pro-apoptotic marker Bax that was previously shown to be increased in toxic psychostimulant protocols (Jayanthi et al. 2001; Raineri et al. 2012). We did not find a significant difference in Bax expression across treatments after chronic treatment or after withdrawal (data not shown).
Effects of Chronic Caffeine and Cocaine Treatments on Striatal Gene Expression: Dopamine, Adenosine, and Glutamate Receptor Subunit mRNA Expression
Considering the central role of striatal circuitry in behaviors related to reward processing, motivation, emotion, and motor activity (Volkow et al. 2012), we characterized relevant gene expression in the dorsal striatum. We were interested in investigating whether dopamine, adenosine, and glutamate receptor expression would be altered after chronic psychostimulant treatments. Thus, we analyzed the effect of chronic combined treatment with caffeine and cocaine on mRNA expression of i) dopamine receptor subunits D1 (Drd1a) and D2 (Drd2), ii) adenosine receptor subunits A1 (Adora1) and A2AR (Adora2a), and iii) glutamate receptor subunits AMPA-type (Gria1 and Gria2), NMDA-type (Grin1, Grin2a, and Grin2b), and metabotropic group I (Grm1 and Grm5) in the striatum.
Chronic treatment
We found that mice receiving the combined Caf–Coc treatment showed specific changes in DA and glutamate receptor subunit expression that were not seen in the Veh, Caf, or Coc groups (Fig. 5). Specifically, ANOVA–Fisher’s LSD showed significant differences across treatments for Drd1a (F (3,19) = 4.19, p < 0.05), Drd2 (F (3,19) = 8.03, p < 0.01), AMPA-type Gria1 (F (3,19) = 8.02, p < 0.01), and NMDA-type Grin2b (F (3,19) = 7.83, p < 0.01). We found reduced expression of these markers in the Caf–Coc group compared to Veh values (p < 0.05). Interestingly, all the psychostimulant-treated mice showed reduced expression of Adora2a after chronic treatment (F (3,19) = 11.74, p < 0.0001). Additionally, we detected a specific effect of chronic caffeine on Grin2a expression (F (3,19) = 7.23, p < 0.01). We found no significant changes in metabotropic group I glutamate receptors after any treatment.

Effect of chronic treatment with caffeine, cocaine, and their combination on striatal dopamine, adenosine, and glutamate receptor subunit mRNA expression. a Dopamine receptor subunits Drd1a (D1R) and Drd2 (D2r). b Adenosine receptor subunits Adora1 (A1R) and Adora2a (A2aR). c Glutamate receptors subunits AMPA-type Gria1 (GluA1) and Gria2 (GluA2), NMDA-type Grin1 (GluN1), Grin2a (GluN2B), and Grin2b (GluN2B), and metabotropic group I Grm1 (mGluR1) and Grm5 (mGluR5). The values indicate mean ± SEM (N = 5). One-way ANOVA–Fisher’s LSD: *p < 0.05, **p < 0.01, ***p < 0.001 different from Veh; $ p < 0.05, $$ p < 0.01, $$$ p < 0.001 different from Coc
Withdrawal
Following a withdrawal period of 7 days, several changes in gene expression in the striatum were observed. Mice treated with the combination of caffeine and cocaine manifested specific changes in DA and glutamate receptor subunit expression that were not seen in the Veh, Caf, or Coc groups (Fig. 6), but with a different profile of gene expression changes in comparison to the one obtained 24 h after the last injections. Specifically, ANOVA–Fisher’s LSD showed significant differences across treatments for Drd1a (F (3,21) = 3.28, p < 0.05), Adora1 (F (3,21) = 6.67, p < 0.01), AMPA-type Gria1 and Gria2 (F (3,21) = 8.02 and 7.15, respectively, p < 0.01), metabotropic Grm1 (F (3,21) = 4.8, p < 0.05), and NMDA-type Grin1 (F (3,21) = 6.72, p < 0.01), where we found increased expression of these markers in the Caf–Coc group compared to Veh values (p < 0.05). Interestingly, psychostimulant-treated mice showed reduced expression of Adora2a (F (3,21) = 4.56, p < 0.05), in a similar manner to the effects found 24 h after chronic treatment. Additionally, we observed an effect of chronic cocaine withdrawal, alone or when combined with caffeine, since both groups that received cocaine showed decreased Grin2a expression (F (3,21) = 8.64, p < 0.001).

Effect of 7-day withdrawal from chronic treatment with caffeine, cocaine, and their combination on striatal dopamine, adenosine, and glutamate receptor subunit mRNA expression. a Dopamine receptor subunits Drd1a (D1R) and Drd2 (D2r). b Adenosine receptor subunits Adora1 (A1R) and Adora2a (A2aR). c Glutamate receptor subunits AMPA-type Gria1 (GluA1) and Gria2 (GluA2), NMDA-type Grin1 (GluN1), Grin2a (GluN2B), and Grin2b (GluN2B), and metabotropic group I Grm1 (mGluR1) and Grm5 (mGluR5). The values indicate mean ± SEM (N = 6). One-way ANOVA–Fisher’s LSD: *p < 0.05, **p < 0.01, ***p < 0.001 different from Veh; $ p < 0.05, $$ p < 0.01, $$$ p < 0.001 different from Coc
Discussion
The results from this study provide evidence that there are significant striatal alterations mediated by the combined caffeine and cocaine administration and highlight the potential negative outcome of chronic intake of both psychostimulants. Combined treatment was associated with increased reactive astrogliosis in striatal tissue and altered gene expression involved in dopaminergic, adenosinergic, and glutamatergic transmission. We also show that some of these detrimental effects persist into withdrawal.
Disturbances of the dorsolateral striatum glutamatergic system are well documented to influence stimulus response, habit learning, and compulsive behaviors (Vollstädt-Klein et al. 2010). Astrocytes can regulate synaptic transmission between neurons by modifying the concentration of extracellular potassium, controlling local blood flow, by releasing and/or taking up neurotransmitters or neuromodulators, by delivering nutrients to neurons, and by altering the geometry and volume of the brain extracellular space (Araque et al. 2014). Therefore, astrocytes are major contributors to synaptic plasticity induced by drugs of abuse (Cadet and Bisagno 2014), and can react to various neurodegenerative insults rapidly, leading to vigorous astrogliosis (Eng et al. 2000). Also, it has been shown that glutamate receptor activation can increase the expression of GFAP (Bowers and Kalivas 2003). It has been suggested that reactive astrogliosis might result following altered neuronal activity, particularly synaptic activity (Maragakis and Rothstein 2006). In our study, all psychostimulant-treated groups showed increased astrogliosis after chronic treatment. Even if many studies are not available reporting cocaine effects on astrocyte reactivity, GFAP has been previously demonstrated to be responsive to cocaine administration (Freeman et al. 2010; Blanco-Calvo et al. 2014). In addition, it was previously shown that caffeine enhanced astroglia and microglia reactivity induced by MDMA in the mouse striatum (Khairnar et al. 2010) and that caffeine can potentiate MDMA-induced neurotoxicity (Frau et al. 2015). Moreover, caffeine potentiated the neurotoxic effects of methamphetamine (Sinchai et al. 2011). Caffeine detrimental effects when given in combination with other psychostimulants might not be exclusive on CNS physiology. Recently, our group (using the same experimental protocol and drug doses) has shown that caffeine can potentiate cocaine toxicity in different cellular components of the mouse testicular tissue (Gonzalez et al. 2015) and that combined administration of both psychostimulants also led to increased lipid peroxidation and altered components of a local dopaminergic system in the testis.
Central effects elicited by caffeine at pharmacologically relevant doses are believed to be principally mediated by an interaction with A1 or A2 subtypes of adenosine receptors since the putative brain concentration is attained with moderate doses (doses not high enough to elicit blockade of adenosine cAMP-phosphodiesterase activity (Fredholm 1995). Adenosine, acting on type-2 receptors, is an effective endogenous anti-inflammatory agent that can modulate inflammation both in the periphery and the brain (Rosi et al. 2003). It is possible that continuous blockade of A2 receptors by chronic caffeine treatment may have disrupted inflammatory responses involving astrocyte reactivity in striatal tissue. In addition, A2AR was found in astrocytes, which co-localized with different glutamate transporters, suggesting the potential role of astrocytic A2AR in the control of pathophysiological processes known to be influenced by glial activity (Matos et al. 2012). It should be noted that chronic cocaine was found to increase reactive oxygen species production in the striatum (Dietrich et al. 2005). We cannot rule out that the cocaine doses used in the present study may have increased reactive oxygen species to some extent, or that when combined with caffeine some oxidative stress might have contributed to astrocyte activation in the striatum.
Repeated exposure to drugs of abuse results in a progressive and long-lasting enhancement of the locomotor response, a phenomenon termed psychomotor (or locomotor) sensitization (Robinson and Berridge 1993). In rodents, sensitization has been shown to correlate with enhanced predisposition to self-administer psychostimulants (Schenk and Partridge 1997). Of relevance to this study, cocaine intake was found to be increased by caffeine administration, and the combination of both drugs showed acutely increased c-Fos expression in the basal ganglia (Kuzmin et al. 2000b). Our study shows that chronic intermittent treatment with caffeine is able to induce locomotor sensitization, in agreement with previous reports showing that caffeine itself may induce sensitization and cross-sensitization with other psychostimulants (Cauli et al. 2003, Hsu et al. 2009). We also showed that chronic treatment with caffeine and cocaine induced locomotor sensitization, while treatment with cocaine alone did not show sensitization. The induction of locomotor sensitization depends on several factors such as the temporal pattern of drug exposure and dose regimen (discussed in Valjent et al. 2010). In addition, the cocaine dose used in this study was previously used to show that Disulfiram was able to facilitate cocaine sensitization, while no sensitization was observed for some variables (distance traveled) after a withdrawal period from chronic 10 mg/kg cocaine (Haile et al. 2003). Moreover, Eisener-Dorman et al. (2011) used a 20 mg/kg cocaine dose in order to investigate cocaine sensitization in several mouse strains suggesting that cocaine doses lower than 20 mg/kg might not be sufficient to induce locomotor sensitization. Thus, in our study, we intentionally decided to use a low dose of cocaine (10 mg/kg), a dose that would have low probability of inducing locomotor sensitization on its own, but that would allow us to search for possible additive effects while combined with caffeine.
Our results are in agreement with a recent study by Prieto et al. (2015) that demonstrated that caffeine can enhance and accelerate the expression of sensitization induced by coca paste in rats. It was previously suggested that caffeine and cocaine stimulate locomotion through different mechanisms (Kuzmin et al. 2000a). Caffeine-stimulated motor responses appear to be mediated by A2AR blockade and to involve dopamine-dependent as well as dopamine independent mechanisms (El Yacoubi et al. 2000). Caffeine could increase motor activity by blocking A2AR and reduce tonic activation of the cAMP/PKA pathway in striatopallidal neurons. This effect would result in reduced phosphorylation of downstream target proteins, thereby affecting the activity of striatopallidal neurons and ultimately enhancing locomotion (Fisone et al. 2007), possibly involving the D2R. Additionally, it has been shown that caffeine alters D2 expression in neurons (Stonehouse et al. 2003). Cocaine effects are linked to both D2 (Ujike et al. 1990; Collins and France 2015) and D1 receptor activation (Wang et al. 2014). Of related interest to this discussion is the fact that a critical aspect of the mechanisms underlying caffeine’s psychostimulant effects is the release of the pre- and postsynaptic brakes that adenosine imposes on striatal dopaminergic neurotransmission (Ferré et al. 2010). In addition, caffeine appears to prime reward-relevant dopaminergic circuits implicated in cocaine misuse, and has been found to reinstate extinguished cocaine-taking behavior in rats (Worley et al. 1994).
An interesting feature of the molecular markers studied in the striatum was that the combined treatment showed a biphasic, time-dependent effect on dopaminergic and glutamatergic markers, with decreased gene expression after chronic treatment and increased expression at withdrawal, suggesting distinct and compensatory changes in chronic administration and following withdrawal. Combined treatment with both psychostimulants showed reduced AMPA and NMDA glutamate receptor subunit expression, along with reduced D1 and D2 mRNA expression. On the contrary, at withdrawal, we found increased D1 mRNA expression along with increased A1, AMPA, NMDA, and metabotropic subunit expression. The exception to this trend was Adora2a mRNA that showed decreased expression after both chronic treatment and withdrawal in all experimental groups. Similarly to the observations in this study, down-regulation of synaptic plasticity-related genes (e.g., Gria1, Drd2, and other molecules that regulate glutamate receptor function) has been reported in the dorsal striatum of relapse-vulnerable rats following chronic cocaine self-administration (Brown et al. 2011). While the precise nature of the changes that occur at the protein level remains to be fully elucidated, our data imply the presence of altered transcriptional control that may contribute to abnormal synaptic AMPA/NMDA subunit composition following chronic treatment and withdrawal. Interestingly, concomitant administration of cocaine and A2AR antagonists reduced glutamatergic synaptic transmission in striatal spiny neurons, while these drugs failed to produce the effect when given in isolation (Tozzi et al. 2012). The contrasting effects on gene expression that we found in this study could reflect changes in glutamate signaling in cortico-striatal synapses. Of interest to this discussion, both D1 and D2, the most abundant subtypes of DA receptors in vivo, are critically involved in drug addiction-related neuroplastic events in the striatum (Kalivas and O’Brien 2008). These results only indicate alterations at the transcriptional level and should then be interpreted with caution since protein levels may not follow the same trend.
There is evidence that patients who are addicted to other drugs use more caffeine than general psychiatric patients (Hays et al. 1998). Our study shows that chronic treatment with caffeine can have deleterious effects on striatal function, and that caffeine intake should be monitored and taken into account in clinical settings aimed to treat cocaine-dependent patients. Caffeine is not only the most consumed psychoactive drug in the world but also a common additive of cocaine (Cole et al. 2011). Also, it was previously reported that cocaine and caffeine are the main components of cocaine paste base and responsible for the CP-induced stimulant action (López-Hill et al. 2011). Cocaine-dependent subjects commonly abuse multiple substances and many of them drink coffee before and after cocaine use. The high frequency of simultaneous exposure to both the drugs may influence the progression to cocaine addiction and the consequences of cocaine exposure. Also, caffeine and cocaine combined exposure may be behind the profound neuropathological alterations found in the brains of cocaine addicts (Cadet et al. 2014). Even after long periods of abstinence, addicts remain vulnerable to drug craving and/or relapses that can be triggered by stimuli previously associated with drugs (Volkow et al. 2012). These features of addiction suggest that drugs might cause a form of persistent neuroplasticity that is acutely responsive to environmental stimuli, with consequent compulsive drug-seeking and drug-taking behaviors. Our report reveals some of the effects that might influence behavioral responses to cocaine, but also highlights interactive effects of the combination of both drugs on neuroplastic changes in the striatum. These effects on striatal gene expression on withdrawal were different compared to the ones obtained 24 h after the last injection suggesting that some of the effects on dopaminergic and glutamatergic synapses in the striatum may actually persist after a washout period.
In conclusion, our study provides novel evidence of detrimental mechanisms activated by combined exposure to caffeine and cocaine. Understanding the underlying mechanisms will guide appropriate intervention strategies for the management of severe reactions and drug-related adverse effects resulting from concomitant caffeine consumption.
References
Araque A, Carmignoto G, Haydon PG, Oliet SH, Robitaille R, Volterra A (2014) Gliotransmitters travel in time and space. Neuron 81:728–739. doi:10.1016/j.neuron.2014.02.007
Bignami A, Eng LF, Dahl D, Uyeda CT (1972) Localization of the glial fibrillary acidic protein in astrocytes by immunofluorescence. Brain Res 43:429–435
Bisagno V, Raineri M, Peskin V, Wikinski SI, Uchitel OD, Llinás RR, Urbano FJ (2010) Effects of T-type calcium channel blockers on cocaine-induced hyperlocomotion and thalamocortical GABAergic abnormalities in mice. Psychopharmacology 212:205–214. doi:10.1007/s00213-010-1947-z
Blanco-Calvo E, Rivera P, Arrabal S, Vargas A, Pavón FJ, Serrano A, Castilla-Ortega E, Galeano P, Rubio L, Suárez J, Rodriguez de Fonseca F (2014) Pharmacological blockade of either cannabinoid CB1 or CB2 receptors prevents both cocaine-induced conditioned locomotion and cocaine-induced reduction of cell proliferation in the hippocampus of adult male rat. Front Integr Neurosci 8(7):106. doi:10.3389/fnint.2013.00106
Bowers MS, Kalivas PW (2003) Forebrain astroglial plasticity is induced following withdrawal from repeated cocaine administration. Eur J Neurosci 17:1273–1278
Brown AL, Flynn JR, Smith DW, Dayas CV (2011) Down-regulated striatal gene expression for synaptic plasticity-associated proteins in addiction and relapse vulnerable animals. Int J Neuropsychopharmacol 14:1099–1110. doi:10.1017/S1461145710001367
Cadet JL, Bisagno V (2014) Glial-neuronal ensembles: partners in drug addiction-associated synaptic plasticity. Front Pharmacol 2(5):204. doi:10.3389/fphar.2014.00204
Cadet JL, Bisagno V (2016) Neuropsychological consequences of chronic drug use: relevance to treatment approaches. Front Psychiatry. doi:10.3389/fpsyt.2015.00189
Cadet JL, Bisagno V, Milroy CM (2014) Neuropathology of substance use disorders. Acta Neuropathol 127:91–107. doi:10.1007/s00401-013-1221-7
Cauli O, Pinna A, Valentini V, Morelli M (2003) Subchronic caffeine exposure induces sensitization to caffeine and cross-sensitization to amphetamine ipsilateral turning behavior independent from dopamine release. Neuropsychopharmacology 28:1752–1759
Centonze D, Grande C, Usiello A, Gubellini P, Erbs E, Martin AB, Pisani A, Tognazzi N, Bernardi G, Moratalla R, Borrelli E, Calabresi P (2003) Receptor subtypes involved in the presynaptic and postsynaptic actions of dopamine on striatal interneurons. J Neurosci 23:6245–6254
Cole C, Jones L, McVeigh J, Kicman A, Syed Q, Bellis M (2011) Adulterants in illicit drugs: a review of empirical evidence. Drug Test Anal 3:89–96. doi:10.1002/dta.220
Collins GT, France CP (2015) Determinants of conditioned reinforcing effectiveness: dopamine D2-like receptor agonist-stimulated responding for cocaine-associated stimuli. Eur J Pharmacol 769:242–249. doi:10.1016/j.ejphar.2015.11.024
Derlet RW, Tseng JC, Albertson TE (1992) Potentiation of cocaine and d-amphetamine toxicity with caffeine. Am J Emerg Med 10:211–216
Dews PB, O’Brien CP, Bergman J (2002) Caffeine: behavioral effects of withdrawal and related issues. Food ChemToxicol 40:1257–1261
Dietrich JB, Mangeol A, Revel MO, Burgun C, Aunis D, Zwiller J (2005) Acute or repeated cocaine administration generates reactive oxygen species and induces antioxidant enzyme activity in dopaminergic rat brain structures. Neuropharmacology 48:965–974
Dunwiddie TV, Masino SA (2001) The role and regulation of adenosine in the central nervous system. Annu Rev Neurosci 24:31–55
Eisener-Dorman AF, Grabowski-Boase L, Tarantino LM (2011) Cocaine locomotor activation, sensitization and place preference in six inbred strains of mice. Behav Brain Funct 1(7):29. doi:10.1186/1744-9081-7-29
El Yacoubi M, Ledent C, Ménard JF, Parmentier M, Costentin J, Vaugeois JM (2000) The stimulant effects of caffeine on locomotor behaviour in mice are mediated through its blockade of adenosine A(2A) receptors. Br J Pharmacol 129:1465–1473
Eng LF, Ghirnikar RS, Lee YL (2000) Glial fibrillary acidic protein: GFAP-thirty-one years (1969-2000). Neurochem Res 25:1439–1451
Ferré S (2013) Caffeine and substance use disorders. J Caffeine Res 3:57–58
Ferré S, Lluís C, Justinova Z, Quiroz C, Orru M, Navarro G, Canela EI, Franco R, Goldberg SR (2010) Adenosine-cannabinoid receptor interactions. Implications for striatal function. Br J Pharmacol 160:443–453. doi:10.1111/j.1476-5381.2010.00723.x
Filip M, Frankowska M, Zaniewska M, Przegaliński E, Muller CE, Agnati L, Franco R, Roberts DC, Fuxe K (2006) Involvement of adenosine A2A and dopamine receptors in the locomotor and sensitizing effects of cocaine. Brain Res 1077:67–80
Fisone G, Håkansson K, Borgkvist A, Santini E (2007) Signaling in the basal ganglia: postsynaptic and presynaptic mechanisms. Physiol Behav 92:8–14
Frau L, Costa G, Porceddu PF, Khairnar A, Castelli MP, Ennas MG, Madeddu C, Wardas J, Morelli M (2015) Influence of caffeine on3,4-methylenedioxymethamphetamine-induced dopaminergic neuron degeneration and neuroinflammation is age-dependent. J Neurochem. doi:10.1111/jnc.13377
Fredholm BB (1995) Astra Award Lecture. Adenosine, adenosine receptors and the actions of caffeine. Pharmacol Toxicol 76:93–101
Fredholm BB, Bättig K, Holmén J, Nehlig A, Zvartau EE (1999) Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol Rev 51:83–133
Fredholm BB, IJzerman AP, Jacobson KA, Klotz KN, Linden J (2001) International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol Rev 53:527–552
Freeman WM, Lull ME, Patel KM, Brucklacher RM, Morgan D, Roberts DC, Vrana KE (2010) Gene expression changes in the medial prefrontal cortex and nucleus accumbens following abstinence from cocaine self-administration. BMC Neurosci 11:29. doi:10.1186/1471-2202-11-29
Garaschuk O, Yaari Y, Konnerth A (1997) Release and sequestration of calcium by ryanodine-sensitive stores in rat hippocampal neurones. J Physiol (Lond) 502(Pt 1):13–30
Gasior M, Jaszyna M, Peters J, Goldberg SR (2000) Changes in the ambulatory activity and discriminative stimulus effects of psychostimulant drugs in rats chronically exposed to caffeine: effect of caffeine dose. J Pharmacol Exp Ther 295:1101–1111
Gonzalez CR, Gonzalez B, Matzkin ME, Muñiz JA, Cadet JL, Garcia-Rill E, Urbano FJ, Vitullo AD, Bisagno V (2015) Psychostimulant-induced testicular toxicity in mice: evidence of cocaine and caffeine effects on the local dopaminergic system. PLoS One 10(11):e0142713. doi:10.1371/journal.pone.0142713
Haile CN, During MJ, Jatlow PI, Kosten TR, Kosten TA (2003) Disulfiram facilitates the development and expression of locomotor sensitization to cocaine in rats. Biol Psychiatry 54:915–921
Hasin DS, O’Brien CP, Auriacombe M, Borges G, Bucholz K, Budney A, Compton WM, Crowley T, Ling W, Petry NM, Schuckit M, Grant BF (2013) DSM-5 criteria for substance use disorders: recommendations and rationale. Am J Psychiatry 170:834–851. doi:10.1176/appi.ajp.2013.12060782
Hays LR, Farabee D, Miller W (1998) Caffeine and nicotine use in an addicted population. J Addict Dis 17:47–54
Hsu CW, Chen CY, Wang CS, Chiu TH (2009) Caffeine and a selective adenosine A2A receptor antagonist induce reward and sensitization behavior associated with increased phospho-Thr75-DARPP-32 in mice. Psychopharmacology (Berl) 204:313–325. doi:10.1007/s00213-009-1461-3
Jayanthi S, Deng X, Bordelon M, McCoy MT, Cadet JL (2001) Methamphetamine causes differential regulation of pro-death and anti-death Bcl-2 genes in the mouse neocortex. FASEB J 15:1745–1752
Jones HE, Griffiths RR (2003) Oral caffeine maintenance potentiates the reinforcing and stimulant subjective effects of intravenous nicotine in cigarette smokers. Psychopharmacology 165:280–290
Kalivas PW, O’Brien C (2008) Drug addiction as a pathology of staged neuroplasticity. Neuropsychopharmacology 33(1):166–180
Khairnar A, PlumitalloA Frau L, Schintu N, Morelli M (2010) Caffeine enhances astroglia and microglia reactivity induced by 3,4-methylenedioxymethamphetamine (‘ecstasy’) in mouse brain. Neurotox Res 17:435–439. doi:10.1007/s12640-009-9125-y
Kuzmin A, Johansson B, Zvartau EE, Fredholm BB (1999) Caffeine, acting on adenosine A(1) receptors, prevents the extinction of cocaine-seeking behavior in mice. J PharmacolExpTher. 290:535–542
Kuzmin A, Johansson B, Fredholm BB, Ögren SO (2000a) Genetic evidence that cocaine and caffeine stimulate locomotion in mice via different mechanisms. Life sciences 66(8):PL113–PL118
Kuzmin A, Johansson B, Semenova S, Fredholm BB (2000b) Differences in the effect of chronic and acute caffeine on self-administration of cocaine in mice. Eur J Neurosci 12:3026–3032
Liguori A, Hughes JR, Goldberg K, Callas P (1997) Subjective effects of oral caffeine in formerly cocaine-dependent humans. Drug Alcohol Depend 49:17–24
López-Hill X, Prieto JP, Meikle MN, Urbanavicius J, Abin-Carriquiry JA, Prunell G, Umpiérrez E, Scorza MC (2011) Coca-paste seized samples characterization: chemical analysis, stimulating effect in rats and relevance of caffeine as a major adulterant. Behav Brain Res 221:134–141. doi:10.1016/j.bbr.2011.03.005
Maragakis NJ, Rothstein JD (2006) Mechanisms of Disease: astrocytes in neurodegenerative disease. Nat ClinPractNeurol 2:679–689
Matos M, Augusto E, Santos-Rodrigues AD, Schwarzschild MA, Chen JF, Cunha RA, Agostinho P (2012) Adenosine A2A receptors modulate glutamate uptake in cultured astrocytes and gliosomes. Glia 60:702–716. doi:10.1002/glia.22290
McNamara R, Kerans A, O’Neill B, Harkin A (2006) Caffeine promotes hyperthermia and serotonergic loss following co-administration of the substituted amphetamines, MDMA (“Ecstasy”) and MDA (“Love”). Neuropharmacology 50:69–80
McPherson PS, Kim YK, Valdivia H, Knudson CM, Takekura H, Franzini-Armstrong C, Coronado R, Campbell KP (1991) The brain ryanodine receptor: a caffeine-sensitive calcium release channel. Neuron 7:17–25
Prieto JP, Galvalisi M, López-Hill X, Meikle MN, Abin-Carriquiry JA, Scorza C (2015) Caffeine enhances and accelerates the expression of sensitization induced by coca paste indicating its relevance as a main adulterant. Am J Addict 24:475–481
Raineri M, Gonzalez B, Goitia B, Garcia-Rill E, Krasnova IN, Cadet JL, Urbano FJ, Bisagno V (2012) Modafinil abrogates methamphetamine-induced neuroinflammation and apoptotic effects in the mouse striatum. PLoS One 7:e46599. doi:10.1371/journal.pone.0046599
Reissig CJ, Strain EC, Griffiths RR (2009) Caffeinated energy drinks–a growing problem. Drug Alcohol Depend 99:1–10. doi:10.1016/j.drugalcdep.2008.08.001
Robinson TE, Berridge KC (1993) The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res Brain Res Rev 18:247–291
Rosi S, McGann K, Hauss-Wegrzyniak B, Wenk GL (2003) The influence of brain inflammation upon neuronal adenosine A2B receptors. J Neurochem 86:220–227
Rosin DL, Hettinger BD, Lee A, Linden J (2003) Anatomy of adenosine A2A receptors in brain: morphological substrates for integration of striatal function. Neurology 61(11 Suppl 6):S12–S18
Schenk S, Partridge B (1997) Sensitization and tolerance in psychostimulant self-administration. Pharmacol Biochem Behav 57:543–550
Schenk S, Valadez A, Horger BA, Snow S, Wellman PJ (1994) Interactions between caffeine and cocaine in tests of self-administration. Behav Pharmacol 5:153–158
Sinchai T, Plasen S, Sanvarinda Y, Jaisin Y, Govitrapong P, Morales NP, Ratanachamnong P, Plasen D (2011) Caffeine potentiates methamphetamine-induced toxicity both in vitro and in vivo. Neurosci Lett 502:65–69. doi:10.1016/j.neulet.2011.07.026
Sofroniew MV (2009) Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci 32:638–647. doi:10.1016/j.tins.2009.08.002
Stonehouse AH, Adachi M, Walcott EC, Jones FS (2003) Caffeine regulates neuronal expression of the dopamine 2 receptor gene. Mol Pharmacol 64:1463–1473
Tozzi A, de Iure A, Marsili V, Romano R, Tantucci M, Di Filippo M, Costa C, Napolitano F, Mercuri NB, Borsini F, Giampà C, Fusco FR, Picconi B, Usiello A, Calabresi P (2012) A2A adenosine receptor antagonism enhances synaptic and motor effects of cocaine via CB1 cannabinoid receptor activation. PLoS One 7(6):e38312. doi:10.1371/journal.pone.0038312
Ujike H, Akiyama K, Otsuki S (1990) D-2 but not D-1 dopamine agonists produce augmented behavioral response in rats after subchronic treatment with methamphetamine or cocaine. Psychopharmacology 102:459–464
Valjent E, Bertran-Gonzalez J, Aubier B, Greengard P, Hervé D, Girault JA (2010) Mechanisms of locomotor sensitization to drugs of abuse in a two-injection protocol. Neuropsychopharmacology 35:401–415. doi:10.1038/npp.2009.143
Vanattou-Saïfoudine N, McNamara R, Harkin A (2012) Caffeine provokes adverse interactions with 3,4-methylenedioxymethamphetamine (MDMA, ‘ecstasy’) and related psychostimulants: mechanisms and mediators. Br J Pharmacol 167:946–959. doi:10.1111/j.1476-5381.2012.02065.x
Vanderschuren LJ, Kalivas PW (2000) Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: a critical review of preclinical studies. Psychopharmacology (Berl) 151:99–120
Volkow ND, Wang GJ, Fowler JS, Tomasi D (2012) Addiction circuitry in the human brain. Annu Rev PharmacolToxicol 52:321–336. doi:10.1146/annurev-pharmtox-010611-134625
Volkow ND, Wang GJ, Logan J, Alexoff D, Fowler JS, Thanos PK, Wong C, Casado V, Ferre S, Tomasi D (2015) Caffeine increases striatal dopamine D2/D3 receptor availability in the human brain. Transl Psychiatry v 5:e549. doi:10.1038/tp.2015.46
Vollstädt-Klein S, Wichert S, Rabinstein J, Bühler M, Klein O, Ende G, Hermann D, Mann K (2010) Initial, habitual and compulsive alcohol use is characterized by a shift of cue processing from ventral to dorsal striatum. Addiction 105:1741–1749. doi:10.1111/j.1360-0443.2010.03022.x
Wang N, Su P, Zhang Y, Lu J, Xing B, Kang K, Li W, Wang Y (2014) Protein kinase D1-dependent phosphorylation of dopamine D1 receptor regulates cocaine-induced behavioral responses. Neuropsychopharmacology 39:1290–1301. doi:10.1038/npp.2013.341
Worley CM, Valadez A, Schenk S (1994) Reinstatement of extinguished cocaine-taking behavior by cocaine and caffeine. Pharmacol Biochem Behav 48:217–221
Acknowledgements
Dr. Bisagno has been authorized to study drug abuse substances in animal models by A.N.M.A.T. (National Board of Medicine Food and Medical Technology, Ministerio de Salud, Argentina). This work was supported by FONCYT-Agencia Nacional de Promoción Científica y Tecnológica; BID 1728 OC.AR. T 2012-0924 Argentina (to Dr. Bisagno) and FONCYT-Agencia Nacional de Promoción Científica y Tecnológica; BID 1728 OC.AR. PICT-2012-1769, Argentina, and UBACYT 2014-2017 #20120130101305BA (to Dr. Urbano). In addition, this work was supported by core facilities of the Center for Translational Neuroscience, UAMS, USA, supported by NIH award P20 GM110702.
Author information
Authors and Affiliations
Corresponding author
Additional information
Javier A. Muñiz and Gimena Gomez have contributed equally to this work.
Rights and permissions
About this article

Cite this article
Muñiz, J.A., Gomez, G., González, B. et al. Combined Effects of Simultaneous Exposure to Caffeine and Cocaine in the Mouse Striatum. Neurotox Res 29, 525–538 (2016). https://doi.org/10.1007/s12640-016-9601-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12640-016-9601-0