
Abstract
The bacteriocin-producing Lactiplantibacillus plantarum SL47 was isolated from conventional fermented sausages, and the bacteriocin SL47 was purified using ethyl acetate, Sephadex G-25 gel chromatography, and reversed-phase high-performance liquid chromatography (RP-HPLC). Bacteriocin SL47 was identified by HPLC–MS/MS combined with whole-genome sequencing, and the results showed it consisted of plantaricin A, J, K, and N. Further characterization analysis showed that the bacteriocin SL47 was highly thermostable (30 min, 121 °C), pH stable (2–10), sensitive to protease and exhibited broad-spectrum antibacterial ability against Gram-positive and Gram-negative bacteria. The mechanism of action showed that the bacteriocin SL47 increased cell membrane permeability, and 2 × minimum inhibitory concentration (MIC) treatment for 40 min caused apoptosis of Staphylococcus aureus F2. The count of S. aureus in the sausage that was inoculated with L. plantarum SL47 and bacteriocin SL47 decreased by about 64% and 53% of that in the initial stage, respectively. These results indicated the potential of L. plantarum SL47 and bacteriocin SL47 as a bio-preservative in meat products.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Fermentation technology was one of the earliest food technology applications. Although fermented sausages are made and eaten worldwide, they are most widely consumed in Europe. For instance, Germany has at least 350 varieties of fermented sausage, whereas Spain has at least 50 [1, 2]. Fermented sausages are popular with consumers because of their unique flavor and rich nutrition, and they ripen under specified temperatures and humidity conditions and usually have no health risks [3]. However, the contaminated raw meat and the propensity for some pathogenic microbes to adapt to their surroundings may produce unsafe sausages [4]. As a common strain of contaminated fermented sausages, Staphylococcus aureus causes sausage corruption and reduces the shelf life of this food. If it is multiplied in food, Staphylococcus aureus enterotoxin can be produced. Moreover, it is highly heat-resistant and cannot be destroyed by ordinary cooking processes. Foodborne diseases can be started by consuming 1 µg of Staphylococcus aureus enterotoxin. Common poisoning symptoms include vomiting, diarrhea, and abdominal cramps [5,6,7,8].
Lactic acid bacteria (LAB) play important roles in fermented sausages. As a sausage starter, they can control the whole process of fermentation, shorten the ripening time, control the growth of pathogenic bacteria and total colonies, inhibit the formation of nitrite, and extend the shelf life of sausages. As a probiotic, they can provide more functions for the development of new products [9, 10]. Bacteriocins produced by some lactic acid bacteria are secondary metabolites with antibacterial activity, and they have the characteristics of safety and efficiency [11, 12]. At present, the abuse of antibiotics enhances the resistance of pathogenic bacteria, so bacteriocin may be a choice to substitute for antibiotics [13,14,15,16,17,18,19,20]. Meanwhile, bacteriocin can improve the safety of fermented meat products which is beneficial to human health [21, 22].
So far, more than one hundred kinds of bacteriocins produced by LAB have been found, but only nisin and pediocin PA-1 are used for commercialization, and the rest are limited to laboratory research [23, 24]. This is mainly because of their properties (narrow antibacterial spectrum, pH sensitivity, and poor thermal stability), time consuming separation and purification methods, and many other factors limit the acquisition of bacteriocins and their industrial production [25, 26]. Bacteriocin has a weak inhibitory effect on S. aureus in foods with high fat and protein content. Lin et al. reported that nisin has no significant inhibitory effect on S. aureus in pork sausages [27]. Therefore, screening lactic acid bacteria bacteriocins for broad antibiotic activity, good pH stability, good thermal stability, and even effective inhibition of spoilage bacteria in foods with high-fat content is still the primary task at present [28, 29].
In this study, a LAB that produces bacteriocin was screened from conventional fermented sausages. The bacteriocin was then purified and identified. The stability, antibacterial spectrum, and mode of action of the bacteriocin SL47 were investigated. L. plantarum SL47 and bacteriocin SL47 as potential contributors to upgrading product quality and safety were also investigated by the production of fermented sausages.
Materials and Methods
Screening and Identification of Lactic Acid Bacteria-Producing Bacteriocins
Manufacturing and Sampling
A total of 24 fermented sausages were randomly selected from different provinces in China. According to the geographical location of different provinces, 4 samples were selected from Xinjiang (Northwest China), 6 from Shandong (East China), 3 from Anhui (East China), 5 from Beijing (North China), 3 from Wuhan (Central China), and 3 from Guangdong (South China).
These fermented sausages were made using pork or mutton as the base, and then, other ingredients such as salt, paprika, sugar, and other spices were added. The products were then packed into natural or artificial casings to begin the fermentation process, followed by the maturation stage. Without using starter cultures, all fermented sausages were made with traditional methods.
Isolation of Strains
All 24 samples were used for the screening of lactic acid bacteria. The 25 g sausages were crushed, and it was immersed in 225 mL of sterile 0.9% NaCl (normal saline) and beaten in a beating machine (Shanghai Jingxin Industrial Development Co., Ltd., Shanghai, China) for 10 min. The homogenate was serially diluted 10-fold with 0.9% NaCl, and 100 µL dilutions were inoculated on an MRS agar (Huankai Microbial Co., Ltd., Guangdong, China) plate containing 3% calcium carbonate [30], and incubated at 37 °C for 48 h. The colonies with dissolved calcium circles were randomly isolated from the plate.
Lactic acid bacteria were cultured in MRS broth at 37 °C for 48 h, and the cell-free supernatant (CFS) was obtained by removing the bacterial with centrifugation at 10000 × g for 15 min at 4 °C. To remove any residual bacteria, the CFS was filtered by a 0.22-µm filter membrane. The pH of the CFS was adjusted to 6 using 1 M NaOH, and then, 2 mg/mL catalase was added to CFS (pH = 6) at a ratio of 1:1 (v/v) and incubated at 37 °C for 2 h. The antibacterial activity was detected by the agar well diffusion method [31]. S. aureus F2 isolated from spoiled fermented sausage was the indicator bacteria species and was incubated in LB broth (Huankai Microbial Co., Ltd., Guangdong, China) at 37 °C for 12 h. Then, 1 mL of indicator bacteria was added to 100 mL LB agar (1%), and then, the culture was poured into a sterile plate with a diameter of 10 cm and dried. After drying, 50 µL of treated CFS was added to the well, and the plate was placed at 4 °C for 2 h and incubated at 37 °C for 6 h. The size of the inhibitory zone was measured to determine the antibacterial activity. Meanwhile, untreated CFS was used as a control test [32].
Identification of Strains
The strain SL47 was Gram-stained using a Gram-staining kit (Sangon Biotech Co., Ltd., China), and morphology was observed under a light microscope. Genomic DNA was extracted using the bacterial genomic DNA extraction kit GenElute (Sangon Biotech Co., Ltd., China), and the 16S rRNA gene was amplified with the forward primer 27F and a reverse primer 1492R [33]. Then, the purified PCR products were sequenced by Sangon Biotech Co., Ltd. The 16S rRNA gene sequence was aligned with related sequences from the NCBI database using the BLAST program [34].
Identification of Bacteriocin
Genome Sequencing and Annotation
The strain SL47 was incubated in MRS broth at 37 °C for 12 h. The cells were collected after centrifugation at 10,000 × g for 10 min at 4 °C, and the collected cells were sent to Shanghai Majorbio Bio-pharm Technology Co., Ltd. for genomic DNA extraction. The method of Li et al. was used to obtain genome sequencing of the strain SL47 [35]. The BAGEL4 and Uniport databases were used for genome mining to locate genes encoding antimicrobial compounds.
Purification of Bacteriocin
CFS and ethyl acetate (Sinopharm Chemical Reagent Co., Ltd., China) were mixed at a ratio of 1:1 for 3 times [36]. The ethyl acetate fraction was evaporated on a rotary evaporator (Shanghai Yarong Biochemical Instrument Co., Ltd., Shanghai, China). The bacteriocin was dissolved in 1 mL of ultrapure water, and the antibacterial activity was detected by the agar well diffusion method with S. aureus F2 as the indicator bacteria.
The bacteriocin obtained by the ethyl acetate extraction was filtered with a 0.22-µm filter membrane (Sinopharm, China). Sephadex G-25 (GE Healthcare, USA) was loaded into a glass column (16 mm × 500 mm), and 3 mL of the extracted sample was added to the chromatographic column and eluted with ultrapure water (Milli-Q) at a flow rate of 1 mL/min. The protein peaks were collected at 2 min/tube using an automatic collector at 280 nm. The antibacterial activity of each fraction on S. aureus F2 was detected by the agar well diffusion method.
Fractions with antimicrobial effect collected from the Sephadex G-25 gel chromatography column were filtered with 0.22-µm filter membrane and further purified by RP-HPLC (Waters 2998, Milford, MA, USA). The purification conditions were as follows: C18 column (Suzhou maikewangzhi Biotechnology Co., Ltd., Suzhou, China, 4.6 mm I.D. × 250 mm, 100 A), mobile phase A comprised ultrapure water (containing 0.1% trifluoroacetic acid), mobile phase B comprised 100% acetonitrile, gradient: 90% A and 10% B within 10 min, 80% A and 20% B within 10 min, 90% A and 10% B within 10 min, the flow rate at 5 mL/min, the volume of samples loaded into the system was 100 µL [37], protein peaks were collected using 280-nm wavelength UV detection, and the collected protein peaks were evaporated to remove acetonitrile. The antibacterial activity of each fraction for S. aureus F2 was determined by the agar well diffusion method.
Peptide Sequencing by HPLC–MS/MS Analysis
The active peaks obtained by RP-HPLC separation and purification were sent to Shanghai Fudan University for HPLC–MS/MS analysis. The bacteriocin solutions were hydrolyzed by trypsin and the resulting peptide fragments were analyzed by Easy-nLC 1200 (Thermo Scientic, P/N LC140) and Orbitrap Exploris 480 (Thermo Scientific, P/N BRE725533). The peptides were captured on a PepMap C18 column (100 µm × 2 cm) at a flow rate of 10 µL/min for 3 min, and then, the peptides were separated on a PepMap C18 column (75 µm × 25 cm) by gradient elution. The MS used Orbitrap for a first-order scan with a range of 350–1600 (m/z) and a resolution of 120,000 m/z. The maximum ion introduction time was 50 ms, and then, the top 20 parent ions that met the higher energy C-trap dissociation (HCD) were used to break the MS/MS fragmentation conditions, and the orbitrap was used to scan the samples with a resolution of 15,000 FWHM (full width at half maxima). The minimum scanning range was set at m/z = 110, and the scanning range was automatically regulated based on the mass charge ratio of the parent ion. In MS/MS, ions might be introduced for a maximum of 50 ms. The amino acid sequences were aligned using the Uniport database (http://www.uniprot.org/blast/).
Analysis of Bacteriocin Molecular Weight
The bacteriocin produced by L. plantarum SL47 was named bacteriocin SL47. The 6 mg/mL crude bacteriocin purified by Sephadex G-25 gel chromatography and freeze-drying were subjected to Tricine SDS-PAGE electrophoresis (4% in the stacking gel, and 16.5% in the running gel). A peptide marker (Sangon Biotech Co., Ltd., China) with a molecular weight (MW) of 4.1–66 kDa was used as the standard control. After electrophoresis at 120 V for 120 min, Coomassie brilliant blue G-250 was used for staining, and the results were recorded using a GenoSens 1850 gel imaging system (ClinX Science Instruments Co., Ltd., China). The bacteriocin SL47 purified by gel filtration chromatography was used for subsequent stability, antibacterial spectrum, and mode of action research.
Characterization of Bacteriocin
Bacteriocin of Stability and Antibacterial Spectrum
The bacteriocin SL47 was treated at − 20, 0, 20, 40, 60, 80, 100, and 121 °C for 30 min, respectively, and S. aureus F2 was used as an indicator bacteria to measure the diameter of the inhibition zone.
The pH of PBS (0.1 mol/L, pH = 7.0) was adjusted to 2, 3, 4, 5, 6, 7, 8, 9, and 10 by 1 mol/L HCl or 1 mol/ L NaOH. The 20 µL bacteriocin SL47 was added to 180 µL PBS with different pH and mixed for 20 min, and then, its pH was adjusted back to the initial pH of bacteriocin SL47 (pH = 6.5). S. aureus F2 was used as an indicator bacteria to measure the diameter of the inhibition zone.
The bacteriocin SL47 was treated with the following proteolytic enzymes: trypsin (pH = 7.4), pepsin (pH = 2.2), papain (pH = 9), α-chymotrypsin (pH = 7.5), and protease K (pH = 7) were dissolved in 20 mmol/L PBS at the optimal pH of each protease. The concentration of each enzyme was 1.0 g/L. Finally, the treated bacteriocin SL47 was adjusted back to its original pH (pH = 6.5) and the agar well diffusion method tested their antimicrobial activity.
Spoilage bacteria from different food sources were incubated in LB broth at 37 °C for 12 h. Then, 1 mL of bacteria was mixed with 100 mL of LB agar (1%) and poured into a sterile plate. 50 µL of bacteriocin was added to a well. The plate was then placed at 4 °C for 2 h and incubated at 37 °C for 6 h. The size of the inhibitory zone was measured to determine the antibacterial activity.
Effect of Bacteriocin SL47 on Cell Membrane Permeability
The MIC of bacteriocin SL47 against S. aureus F2 was determined by the double dilution method. LB medium with 512 µg/mL bacteriocin SL47 was 2-fold serially diluted. Then, 50 µL of different concentrations of diluents were added to a 96-well plate containing S. aureus F2 suspension (6 log CFU/mL) and cultured at 37 ℃ for 12 h to determine the MIC. The concentration corresponding to the diluent with the minimum inhibitory effect was the MIC [38].
The bacteria of S. aureus F2 with OD600 = 0.4 were collected, and 2 × MIC bacteriocin was added to S. aureus F2 at a ratio of 1:1 at 37 °C for 20 min, respectively. The blank control was treated with the same amount of sterile PBS. The cell morphology was fixed with 2.5% glutaraldehyde (Solarbio, Beijing, China) at 4 °C for 12 h. Then, the bacteria were washed three times with sterile PBS and dehydrated by a series of graded ethanol solutions (30, 50, 70, 80, 90, and 100%). Each gradient was washed for 10 min and centrifuged at 10000 × g for 5 min to remove the ethanol. Finally, after resuspension with 500 µL of anhydrous ethanol, 20 µL of resuspension was evenly coated on a clean coverslip. The samples were dried and gilded and imaged using a Sigma 300 scanning electron microscope (SEM) (ZEISS, Germany).
Following the same steps as scanning electron microscopy, 20 µL of suspension was dropped onto a mica plate. The results were observed using BioScope Resolve Atomic Force Microscope (AFM) (BRUKER, USA).
Membrane permeability and dynamic changes were assessed using the LIVE/DEAD™ BacLight™ Bacterial Viability Kit, which utilizes a vital-staining probe mixture (UElandy, Suzhou, China). S. aureus F2 with OD600 = 0.4 was treated with 1: 1 bacteriocin at 2 × MIC for 20 and 40 min at 37 °C, and the same amount of sterile PBS was added as the blank control. After being washed three times with PBS, the cells were mixed with NucGreen and EthD-III staining. The mixture was incubated in the dark at 37 ℃ for 15 min. Finally, fluorescence images were collected using the LSM-710 (Carl Zeiss, Germany) microscope. The excitation/emission wavelengths used for NucGreen and EthD-III were 503/530 nm and 530/620 nm, respectively. [34].
The Effect of Bacteriocin SL47 and L. plantarum SL47 on S. aureus in Fermented Sausages
L. plantarum SL47 and S. aureus F2 were grown at 37 °C for 12 h in MRS and Barder-Parker broth (Huankai Microbial Co., Ltd., Guangdong, China), respectively. The cells were washed, collected by centrifugation, and resuspended in 25 mL 0.05 M PBS (pH = 7). The meat was cut into pieces about 8 mm in diameter and mixed with the ingredients. Sausages were produced using lean pork (65%), backfat pork (20%), starch (4%), sugar (4%), garlic (4%), salt (2.5%) and black pepper (0.5%). The sausages were divided into 5 equal groups. Each of the groups was contaminated with S. aureus F2 at a concentration of 3 log CFU/g and treated with bacteriocin or strain as follows:
-
Group A: a control group, without added bacteriocin SL47 or L. plantarum SL47.
-
Group B: 6 log CFU/g L. plantarum SL47 was added.
-
Group C: 40 µg/g bacteriocin SL47 was added.
-
Group D: 6 log CFU/g L. plantarum SL47 and 40 µg/g bacteriocin SL47 were added.
-
Group E: 40 µg/g nisin was added.
All groups were filled into casings and placed in a VI180SA-1 incubation chamber (Vinicole Electrical Appliance Co. Ltd., China). The incubation was performed at 20 ℃ and 95% relative humidity (RH) for 14 days. The sausages of five groups were analyzed at 0, 3, 7, 10, and 14 days for S. aureus, lactic acid bacteria, total viable CFU count, and pH. For microbiological determinations, the same method as the isolation of strains was used. MRS agar (containing 2% calcium carbonate) was used for total lactic acid bacteria count, PCA agar (Huankai Microbial Co., Ltd., Guangdong, China) was used for total viable count, and Baird-Parker agar was used for S. aureus count. The pH was measured by a pH electrode connected to a pH meter PHS-3G (INESA scientific Instrument Co., Ltd., China).
Statistical Analysis
The data analysis was performed using GraphPad Prism 9. For the stability of bacteriocin, the inhibition zone size of bacteriocin at different temperatures and pH was analyzed. The effects of bacteriocin SL47 and L. plantarum SL47 in fermented sausages, the changes in S. aureus count, total viable CFU count, lactic acid bacteria count, and pH during fermentation were statistically analyzed. All data were the average representation of three experimental replicates, and the results were shown as mean ± SD. The analysis of variance (ANOVA) test was used and p < 0.05 was considered significant.
Accession Number
Lactiplantibacillus plantarum SL47 was identified by 16S rRNA gene sequencing (GenBank accession number: OQ799052). The whole-genome sequence of L. plantarum SL47 has been uploaded to the GenBank database with accession number JARQOB000000000. The GenBank accession numbers of plantaricin A, E, F, J, K, and N were OR068554, OR068555, OR068556, OR068557, OR068558, and OR068559, respectively.
Results
Screening and Identification of Strains
The 186 strains producing inhibition zones were obtained from 24 fermented sausages. After eliminating the possibility that the inhibitory potency of strain SL47 was due to the organic acid and hydrogen peroxide, strain SL47 exhibited the strongest antimicrobial activity with the largest diameter of the inhibition zone against S. aureus F2. The phylogenetic tree based on the 16S rRNA gene sequence of strain SL47 was shown in Fig. 1. According to Fig. 1, Lactiplantibacillus plantarum was the closest species to strain SL47, so strain SL47 was identified as Lactiplantibacillus plantarum.

Phylogenetic tree of strain SL47
Identification of Bacteriocin
Identification of Bacteriocin-Producing Genes
Bacteriocin-coding loci were found in the genome of L. plantarum SL47, as shown in Fig. 2. Six proteins (plantaricin A (PlnA), E (PlnE), F (PlnF), J (PlnJ), K (PlnK), and N (PlnN)) were identified as core proteins in areas of interest (AOI), and their complete amino acid sequences were aligned with plantaricin A, E, F, J, K, and N in Lactiplantibacillus plantarum (strain ATCC BAA-793/NCIMB 8826/WCFS1) by Uniport, the identities of plantaricin A, E, J, and N were the highest (100%), the identity of plantaricin F was 96.4%, and the identity of plantaricin K was 98.2%.

Bacteriocin synthesis gene cluster (green represents the core genes, which are PlnE, PlnF, PlnA, PlnN, PlnJ, and PlnK from left to right)
Purification of Bacteriocin
The bacteriocin extracted by ethyl acetate was purified using Sephadex G-25 gel chromatography. As shown in Fig. 3, there are two elution peaks in the purification of Sephadex G-25. The results showed that only A-2 had antibacterial activity against S. aureus F2. At the active peak, only 2 mL of the fraction was collected at the peak tip, and it was concentrated by freeze-drying after repeated collection.

Sephadex G-25 gel chromatography of the ethyl acetate fraction (the antibacterial activity of each peak was displayed on a plate in the top right corner)
There were 4 absorption peaks after purification by RP-HPLC (Fig. 4). B-2 and B-4 had antibacterial activity. Therefore, the eluent of the B-2 and B-4 were collected for further analysis. Data related to the purification process were summarized in Table 1.

Reversed-phase high-performance liquid chromatography (RP-HPLC) of the A-2 fraction (the antibacterial activity of each peak was displayed on a plate in the top right corner)
Sequence and Molecular Weight Analysis of Bacteriocin
The amino acid sequence of purified B-2 was QLSNKEMQKIVGGK by HPLC–MS/MS, and its MW was 1559.84 Da. The amino acid sequence of purified B-4 was MTVNKMIKDLDVVDAFAPISNNK, NISGGR, and MKSLDKIAGLGIEMAEK by HPLC–MS/MS, and their MW were 2564.01, 602.65, and 1834.22 Da, respectively. The peptides obtained by HPLC–MS/MS (Fig. 5) were aligned with the sequence predicted by the genome of L. plantarum SL47. The peptide fragment in B-2 was identified as PlnA. B-4 contained three peptide fragments identified as PlnJ, K, and N. Therefore, L. plantarum SL47 produced four bacteriocins, namely, PlnA, J, K, and N, and there were silent genes encoding PlnE and F. The parameters of PlnA, PlnJ, PlnK, and PlnN were calculated by the antimicrobial peptide database (https://aps.unmc.edu/AP) and it found that their MW was similar, about 6.0 KDa (Table 2) and Tricine SDS-PAGE results also showed a single band at 4.1–6.5 kDa in the stained gel (Fig. 6).

Identification of plantaricin by HPLC–MS/MS. A Plantaricin A, B Plantaricin J, C Plantaricin K, and D Plantaricin N (A was from B-2 in RP-HPLC and B–D were from B-4 in RP-HPLC. “ + ” indicates that the charge of the ion is 1)

Determination of molecular weight of the bacteriocin SL47 by Tricine SDS-PAGE (M was protein marker; lane 1 was bacteriocin SL47 obtained by Sephadex G-25 gel chromatography)
Characterization of Bacteriocin
Stability and Antibacterial Spectrum of Bacteriocin
Figure 7A showed the stability of bacteriocin SL47 after heat treatment at −20 to 121 °C for 30 min. Figure 7B showed that the antibacterial activity of SL47 decreased under alkaline conditions, while the acidic environment had little effect on the activity of SL47. It should be noted that the activity of SL47 decreased significantly at pH 10. The antimicrobial activity of bacteriocin SL47 decreased after enzymatic treatment with pepsin and trypsin. After treatment with papain, α-chymotrypsin, and proteinase K, the antibacterial activity was lost. The bacteriocin SL47 exhibited a good antibacterial spectrum (Table 3) and significantly inhibited growth of Gram-positive bacteria such as S. aureus, Listeria monocytogenes, and Bacillus cereus. In addition, growth of Gram-negative bacteria such as Escherichia coli, Aeromonas hydrophila, Vibrio parahaemolyticus, P. aeruginosa, and Salmonella were inhibited.

Thermal and pH stability of bacteriocin SL47. A Thermal stability of bacteriocin SL47. B pH stability of bacteriocin SL47 (the errors bars are standard deviation). Different superscript lowercase letters represent significant differences between groups (p < 0.05)
Effect of Bacteriocin SL47 on Cell Membrane Permeability
The MIC of bacteriocin SL47 was 16 µg/mL by the double dilution method. Using SEM and AFM, the surface of S. aureus F2 cells without bacteriocin SL47 treatment was smooth. After 2 × MIC treatment for 20 min, the cell membrane began to rupture, the intracellular substances began to leak, and some cells were completely deformed (Figs. 8 and 9).

The cell morphology of S. aureus F2 observed by SEM (A, B control and C, D S. aureus cells treated with bacteriocin SL47 at 2 × MIC for 20 min)

The cell morphology of S. aureus F2 observed by AFM (A control and B, C S. aureus cells treated with bacteriocin SL47 at 2 × MIC for 20 min)
Living S. aureus F2 cells with intact membranes were green using NucGreen, while cells with damaged membranes appeared red due to the entry of EthD-III after membrane integrity was impaired. With the increase in treatment time, the transformation of bacteria from green fluorescence to red fluorescence was observed. The 2 × MIC bacteriocin SL47 killed all the S. aureus F2 cells within 40 min (Fig. 10).

The LIVE/DEAD of S. aureus F2 observed by confocal laser scanning microscopy (A control and B S. aureus cells treated with bacteriocin SL47 at 2 × MIC for 20 min; C, S. aureus cells treated with bacteriocin SL47 at 2 × MIC for 40 min). A control batch, B batch with 6 log CFU/g L. plantarum SL47, C batch with 40 µg/g bacteriocins, D batch with 6 log CFU/g L. plantarum SL47 and 40 µg/g bacteriocins, and (E) batch with 40 µg/g Nisin
The Effect of Bacteriocin SL47 and L. plantarum SL47 on S. aureus in Fermented Sausages
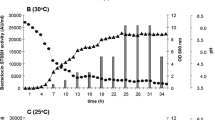
The results of microbiological and pH investigations of sausages during the fermentation process inoculated with L. plantarum SL47 or bacteriocin SL47 were shown in Fig. 11. During the whole maturation stage, the count of S. aureus in groups B, C, D and E decreased by 1.92, 1.6, 1.98, and 1.2 log CFU/g, respectively (Fig. 11a). The lactic acid bacteria and total viable count in all groups increased compared with the initial stage (Fig. 11b, c). In the group (B and D) inoculated with L. plantarum SL47, the lactic acid bacteria increased rapidly, and the total viable cell count also increased. In sausages (C and E) inoculated with bacteriocin SL47 and nisin, lactic acid bacteria increased rapidly in the early stage of ripening and decreased in the later stage. The results showed that bacteriocin SL47 and nisin did not sufficiently inhibit the growth of lactic acid bacteria, while nisin had a stronger inhibitory effect on lactic acid bacteria than bacteriocin SL47. The pH of all groups decreased compared with the initial stage. It is worth noting that the pH of the groups (B and D) inoculated with L. plantarum SL47 was lower than that of the bacteriocin SL47 during the whole maturation stage, especially in the first 3 days of fermentation (Fig. 11d).

Microbial and pH succession (log CFU/g ± SD) during the ripening of sausages (a S. aureus count, b lactic acid bacteria count, c total viable count, and d pH value of sausages). Different superscript lowercase letters represent significant differences between groups (p < 0.05)
Discussion
Staphylococcus aureus is one of the main pathogens that can form biofilms and contaminate food. Plantaricin, a class IIa LAB bacteriocin, was found to kill S. aureus and prevent biofilm formation [39]. L. plantarum C11 was reported to have genes encoding six bacteriocins (PlnA, PlnE, PlnF, PlnJ, PlnK, and PlnN). Among the genes encoding these six bacteriocins, PlnA encodes PlnA, which is located on the same operon as the two-component regulatory system (Pln-BCD). The operons PlnEFI and PlnJKLR encode 2 bacteriocin peptides (PlnEF and PlnJK) and hydrophobic proteins like bacteriocin immune proteins. The operon PlnMNOP encodes a similar bacteriocin-like peptide (PlnN) and a putative immune protein [40]. Among them, PlnA is mainly a peptide pheromone that can induce strains to produce bacteriocins. It also has membrane permeability and an antibacterial effect. PlnEF and PlnJK had antibacterial activity, while PlnN had no antibacterial activity [41]. Therefore, PlnA and PlnJK are bacteriocins that play a bacteriostatic role in bacteriocin SL47. PlnA belongs to class IIc bacteriocin and PlnJK belongs to class IIb bacteriocin, like class II bacteriocin purified by other studies [42,43,44]. Their mode of action is to lyse the cell membrane of bacteria, increase the permeability of the cell membrane, and achieve the bactericidal effect. In general, the MICs of plantaricin against S. aureus were low. Then, 220 µg/mL plantaricin LD4 caused S. aureus cell membrane shrinkage and content leakage [45]. When S. aureus was treated with 64 µg/mL plantaricin 827, a few cells began to wrinkle, and with the increase of plantaricin 827, the cell membrane began to distort significantly in more cells [46]. Then, 256 µg/mL plantaricin FB-2 caused S. aureus cell membrane rupture [47]. Compared with the above research, only 32 µg/mL of bacteriocin SL47 can destroy S. aureus, showing a good killing effect on S. aureus.
Bacteriocin SL47 was identified as PlnA, PlnJ, PlnK, and PlnN by HPLC–MS/MS and had a MW of about 6.0 KDa by Tricine SDS-PAGE. Similarly, the molecular weight of plantaricin JY22 and Bac23 were estimated as 4.1 kDa and 5.1 kDa using SDS-PAGE, respectively [48, 49]. Due to the band being composed of two or more proteins that migrate at the same apparent molecular weight on the gel, the presence of a single band using Tricine SDS-PAGE did not necessarily mean that there was only one protein [50], and due to their similar properties, separating them in the purification process was difficult. It was also difficult to identify four bacteriocins by conventional mass spectrometry and automatic Edman degradation. According to Moyer et al. only the mass spectrometry combined with the genomic information of L. plantarum SL47 could confirm the type of bacteriocin expressed [51]. This may be a new method for identifying bacteriocins in the future.
The bacteriocin SL47 had strong thermal and pH stability. Plantaricin W3-2 [52], plantaricin MG [53], plantaricin GZ1-27 [34], plantaricin LPL-1 [43], plantaricin ASM1 [54], and plantaricin SLG1 [55] have similar properties. The reason may be that the hydrophobic protein is small, has almost no tertiary structure, and has a strong hydrophobic region, making the cross-linked structure stable. Both bacteriocin SL47 and plantaricin 163 [30] were sensitive to protease, which indicated that they had little negative effects on intestinal microflora. In addition, the bacteriocins nisin and pediocin PA-1, both currently approved by the FDA as food preservatives, show a narrow antibacterial spectrum and most bacteriocins can only inhibit Gram-positive bacteria [24]. It is worth noting that the bacteriocin SL47 has activity to inhibit Gram-negative bacteria and has a broad antibacterial spectrum. According to the theory of Klaenhammer [56], bacteriocins produced by lactic acid bacteria are active against closely related bacteria. The structure and composition of the outer membrane of Gram-negative bacteria hinder the site for bacteriocin action, e.g., plantaricin S, T [57], C [58], and W [59] can only inhibit Gram-positive bacteria. However, a few plantaricins can not only inhibit Gram-positive bacteria but also Gram-negative bacteria. The interaction of plantaricin MG with the cytoplasmic membrane of energized cells due to the presence of proton dynamics led to the formation of pores, which allowed the efflux of ions leading to the death of Salmonella typhimurium [60]. Plantaricin BM-1 can inhibit the formation of biofilm, leading to the rupture and shrunk of E. coli cells [61]. Plantaricin J/K effectively inhibited E. coli due to changes in bacterial fatty acid composition and membrane fluidity at low temperatures [62]. Bacteriocin SL47 contains plantaricin J/K, and the Gram-negative bacteria reacted with bacteriocin SL47 at 4 ℃ for 2 h in the early stage of the agar well diffusion process. Like the above research, the bacteriocin SL47 promoted the inhibition of Gram-negative bacteria at low temperatures.
The bacteriocin SL47 not only had a bacteriostatic effect on the agar plate but also had a good bacteriostatic effect in the process of sausage fermentation. The bacteriostatic effect of bacteriocin SL47 was better than that of nisin, which may be because bacteriocin SL47 was composed of multiple bacteriocins, which could achieve a better bacteriostatic effect. However, the effect of direct addition of bacteriocin-producing strains in sausages was better than that of the direct addition of bacteriocin. This was because during the growth of lactic acid bacteria, not only bacteriocins were produced, but also antibacterial substances such as organic acids were produced [63]. The organic acid produced by lactic acid bacteria caused the pH in the system to decrease rapidly and then increase slightly or remain basically stable, which was consistent with the results of Kozacinski et al. [64]. The antibacterial mode, stability, and effectiveness in the sausage shows that bacteriocin SL47 has excellent properties to be used in meat products.
Conclusion
In conclusion, L. plantarum SL47 was screened from naturally fermented sausages, and it made the bacteriocins were PlnA, J, K, and N. Furthermore, several experiments showed that bacteriocin SL47 had broad spectrum antibacterial ability, good thermal, and pH stability. Bacteriocin SL47 eventually led to bacterial cell death by increasing the permeability of the S. aureus F2 cell membrane. Both bacteriocin SL47 and L. plantarum SL47 were inoculated into fermented sausages, and the result showed they could inhibit S. aureus to improve the safety and quality of fermented sausages.
Data Availability
All research data in this study are available from the corresponding author with a reasonable request.
References
Kolozyn-Krajewska D, Dolatowski Z, Zielinska D (2012) Probiotic use in meat products. Fleischwirtschaft 92(4):101–108
Rouhi M, Sohrabvandi S, Mortazavian AM (2013) Probiotic fermented sausage: viability of probiotic microorganisms and sensory characteristics. Crit Rev Food Sci Nutr 53(4):331–348. https://doi.org/10.1080/10408398.2010.531407
Villani F, Casaburi A, Pennacchia C, Filosa L, Russo F, Ercolini D (2007) Microbial ecology of the soppressata of Vallo di Diano, a traditional dry fermented sausage from southern Italy, and in vitro and in situ selection of autochthonous starter cultures. Appl Environ Microbiol 73(17):5453–5463. https://doi.org/10.1128/aem.01072-07
Casaburi A, Di Martino V, Ferranti P, Picariello L, Villani F (2016) Technological properties and bacteriocins production by Lactobacillus curvatus 54M16 and its use as starter culture for fermented sausage manufacture. Food Control 59:31–45. https://doi.org/10.1016/j.foodcont.2015.05.016
Schubert J, Krakowiak S, Bania J (2018) Production of staphylococcal enterotoxins in food. Med Weter-Vet Med-Sci Pract 74(1):16–22. https://doi.org/10.21521/mw.5837
Sameshima T, Magome C, Takeshita K, Arihara K, Itoh M, Kondo Y (1998) Effect of intestinal Lactobacillus starter cultures on the behaviour of Staphylococcus aureus in fermented sausage. Int J Food Microbiol 41(1):1–7. https://doi.org/10.1016/s0168-1605(98)00038-5
Lebert I, Leroy S, Giammarinaro P, Lebert A, Chacornac JP, Bover-Cid S et al (2007) Diversity of microorganisms in the environment and dry fermented sausages of small traditional French processing units. Meat Sci 76(1):112–122. https://doi.org/10.1016/j.meatsci.2006.10.019
Ferreira V, Barbosa J, Vendeiro S, Mota A, Silva F, Monteiro MJ et al (2006) Chemical and microbiological characterization of alheira: a typical Portuguese fermented sausage with particular reference to factors relating to food safety. Meat Sci 73(4):570–575. https://doi.org/10.1016/j.meatsci.2006.02.011
Wang ZL, Wang ZX, Ji LL, Zhang JM, Zhao ZP, Zhang R et al (2021) A review: microbial diversity and function of fermented meat products in China. Front Microbiol. https://doi.org/10.3389/fmicb.2021.645435
Minj J, Chandra P, Paul C, Sharma RK (2021) Bio-functional properties of probiotic Lactobacillus: current applications and research perspectives. Crit Rev Food Sci Nutr 61(13):2207–2224. https://doi.org/10.1080/10408398.2020.1774496
Shafique B, Ranjha MMAN, Murtaza MA, Walayat N, Nawaz A, Khalid W et al (2023) Recent trends and applications of nanoencapsulated bacteriocins against microbes in food quality and safety. Microorganisms 11(1). https://doi.org/10.3390/microorganisms11010085
Fischer SW, Titgemeyer F (2023) Protective cultures in food products: from science to market. Foods 12(7). https://doi.org/10.3390/foods12071541
Reis JA, Paula AT, Casarotti SN, Penna ALB (2012) Lactic acid bacteria antimicrobial compounds: Characteristics and applications. Food Eng Rev 4(2):124–140. https://doi.org/10.1007/s12393-012-9051-2
Montalban-Lopez M, Sanchez-Hidalgo M, Valdivia E, Martinez-Bueno M, Maqueda M (2011) Are bacteriocins underexploited? Novel applications for old antimicrobials. Curr Pharm Biotechnol 12(8):1205–1220. https://doi.org/10.2174/138920111796117364
Galvez A, Abriouel H, Benomar N, Lucas R (2010) Microbial antagonists to food-borne pathogens and biocontrol. Curr Opin Biotechnol 21(2):142–148. https://doi.org/10.1016/j.copbio.2010.01.005
Bhattacharya D, Nanda PK, Pateiro M, Lorenzo JM, Dhar P, Das AK (2022) Lactic acid bacteria and bacteriocins: novel biotechnological approach for biopreservation of meat and meat products. Microorganisms 10(10). https://doi.org/10.3390/microorganisms10102058
Bangar SP, Chaudhary VP, Singh TP, Ozogul F (2022) Retrospecting the concept and industrial significance of LAB bacteriocins. Food Biosci. https://doi.org/10.1016/j.fbio.2022.101607
Balciunas EM, Castillo Martinez FA, Todorov SD, de Melo Gombossy, Franco BD, Converti A, de Souza Oliveira RP (2013) Novel biotechnological applications of bacteriocins: a review. Food Control 32(1):134–142. https://doi.org/10.1016/j.foodcont.2012.11.025
Kaskoniene V, Stankevicius M, Bimbiraite-Surviliene K, Naujokaityte G, Serniene L, Mulkyte K et al (2017) Current state of purification, isolation and analysis of bacteriocins produced by lactic acid bacteria. Appl Microbiol Biotechnol 101(4):1323–1335. https://doi.org/10.1007/s00253-017-8088-9
Simons A, Alhanout K, Duval RE (2020) Bacteriocins, antimicrobial peptides from bacterial origin: overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 8(5). https://doi.org/10.3390/microorganisms8050639
Swetwiwathana A, Visessanguan W (2015) Potential of bacteriocin-producing lactic acid bacteria for safety improvements of traditional Thai fermented meat and human health. Meat Sci 109:101–105. https://doi.org/10.1016/j.meatsci.2015.05.030
Rivas FP, Castro MP, Vallejo M, Marguet E, Campos CA (2014) Sakacin Q produced by Lactobacillus curvatus ACU-1: functionality characterization and antilisterial activity on cooked meat surface. Meat Sci 97(4):475–479. https://doi.org/10.1016/j.meatsci.2014.03.003
Lu X, Hu P, Dang Y, Liu B (2014) Purification and partial characterization of a novel bacteriocin produced by Lactobacillus casei TN-2 isolated from fermented camel milk (Shubat) of Xinjiang Uygur Autonomous region. China. Food Control 43:276–283. https://doi.org/10.1016/j.foodcont.2014.03.020
Aljohani AB, Al-Hejin AM, Shori AB (2023) Bacteriocins as promising antimicrobial peptides, definition, classification, and their potential applications in cheeses. Food Sci Technol 43:e1181021. https://doi.org/10.1590/fst.118021
Deegan LH, Cotter PD, Hill C, Ross P (2006) Bacterlocins: biological tools for bio-preservation and shelf-life extension. Int Dairy J 16(9):1058–1071. https://doi.org/10.1016/j.idairyj.2005.10.026
Sharafi H, Maleki H, Ahmadian G, Zahiri HS, Sajedinejad N, Houshmand B et al (2013) Antibacterial activity and probiotic potential of Lactobacillus plantarum HKN01: a new insight into the morphological changes of antibacterial compound-treated Escherichia coli by electron microscopy. J Microbiol Biotechnol 23(2):225–236. https://doi.org/10.4014/jmb.1208.08005
Lin L, Hu JY, Wu Y, Chen M, Ou J, Yan WL (2018) Assessment of the inhibitory effects of sodium nitrite, nisin, potassium sorbate, and sodium lactate on Staphylococcus aureus growth and staphylococcal enterotoxin A production in cooked pork sausage using a predictive growth model. Food Sci Hum Wellness 7(1):83–90. https://doi.org/10.1016/j.fshw.2017.12.003
Woraprayote W, Janyaphisan T, Adunphatcharaphon S, Sonhom N, Showpanish K, Rumjuankiat K et al (2023) Bacteriocinogenic lactic acid bacteria from Thai fermented foods: potential food applications. Food Biosci 52:102385. https://doi.org/10.1016/j.fbio.2023.102385
Zhang T, Zhang Y, Li L, Jiang X, Chen Z, Zhao F et al (2022) Biosynthesis and production of class II bacteriocins of food-associated lactic acid bacteria. Fermentation-Basel 8(5). https://doi.org/10.3390/fermentation8050217
Hu MZ, Zhao HZ, Zhang C, Yu JS, Lu ZX (2013) Purification and characterization of plantaricin 163, a novel bacteriocin produced by Lactobacillus plantarum 163 isolated from traditional Chinese fermented vegetables. J Agric Food Chem 61(47):11676–11682. https://doi.org/10.1021/jf403370y
Ben Ayed H, Maalej H, Hmidet N, Nasri M (2015) Isolation and biochemical characterisation of a bacteriocinlike substance produced by Bacillus amyloliquefaciens An6. J Glob Antimicrob Resist 3(4):255–261. https://doi.org/10.1016/j.jgar.2015.07.001
Barbosa MS, Todorov SD, Belguesmia Y, Choiset Y, Rabesona H, Ivanova IV et al (2014) Purification and characterization of the bacteriocin produced by Lactobacillus sakei MBSa1 isolated from Brazilian salami. J Appl Microbiol 116(5):1195–1208. https://doi.org/10.1111/jam.12438
Lv XR, Lin Y, Jie Y, Sun MT, Zhang BL, Bai FL et al (2018) Purification, characterization, and action mechanism of plantaricin DL3, a novel bacteriocin against Pseudomonas aeruginosa produced by Lactobacillus plantarum DL3 from Chinese Suan-Tsai. Eur Food Res Technol 244(2):323–331. https://doi.org/10.1007/s00217-017-2958-3
Du H, Yang J, Lu X, Lu Z, Bie X, Zhao H et al (2018) Purification, characterization, and mode of action of plantaricin GZ1-27, a novel bacteriocin against Bacillus cereus. J Agric Food Chem 66(18):4716–4724. https://doi.org/10.1021/acs.jafc.8b01124
Li H, Liu T, Zhang X, Xiong Z, Hong Q, Jia S et al (2023) Whole-genome sequencing and bacteriocin purification of Lactiplantibacillus plantarum HY41 confirms bactericidal and probiotic potential. Int Biodeterior Biodegrad 185:105685. https://doi.org/10.1016/j.ibiod.2023.105685
Ruixiang Z, Yanqing L, Junjian R, Gang L, Shuang L, Yang Z et al (2020) Purification and characterization of bacteriocin produced by Lactobacillus rhamnosus zrx01. Food Biosci 38:100754-. https://doi.org/10.1016/j.fbio.2020.100754
Li H, Guo L, Zhang X, Mu H, Sha S, Lin Y et al (2022) Whole-genome sequencing combined with mass spectrometry to identify bacteriocin and mine silent genes. Lwt-Food Sci Technol. https://doi.org/10.1016/j.lwt.2022.113975
Du RP, Ping WX, Ge JP (2022) Purification, characterization and mechanism of action of enterocin HDX-2, a novel class IIa bacteriocin produced by Enterococcus faecium HDX-2. Lwt-Food Sci Technol. https://doi.org/10.1016/j.lwt.2021.112451
Pei JJ, Huang YG, Ren T, Guo YD, Dang J, Tao YD et al (2022) The antibacterial activity mode of action of plantaricin YKX against Staphylococcus aureus. Molecules 27(13). https://doi.org/10.3390/molecules27134280
Dzung Bao D, Havarstein LS, Nes IF (1996) Characterization of the locus responsible for the bacteriocin production in Lactobacillus plantarum C11. J Bacteriol 178(15):4472–4483
Anderssen EL, Diep DB, Nes IF, Eijsink VGH, Nissen-Meyer J (1998) Antagonistic activity of Lactobacillus plantarum C11: two new two-peptide bacteriocins, plantaricins EF and JK, and the induction factor plantaricin A. Appl Environ Microbiol 64(6):2269–2272
Arief II, Budiman C, Jenie BSL, Andreas E, Yuneni A (2015) Plantaricin IIA-1A5 from Lactobacillus plantarum IIA-1A5 displays bactericidal activity against Staphylococcus aureus. Benef Microbes 6(4):603–613. https://doi.org/10.3920/bm2014.0064
Wang Y, Qin Y, Xie Q, Zhang Y, Hu J, Li P (2018) Purification and characterization of plantaricin LPL-1, a novel class IIa bacteriocin produced by Lactobacillus plantarum LPL-1 isolated from fermented fish. Front Microbiol. https://doi.org/10.3389/fmicb.2018.02276
Jinjin P, Xinsheng L, Hao H, Yanduo T (2018) Purification and characterization of plantaricin SLG1, a novel bacteriocin produced by Lb. plantarum isolated from yak cheese. Food Control 84:111–117. https://doi.org/10.1016/j.foodcont.2017.07.034
Sheoran P, Tiwari SK (2019) Anti-staphylococcal activity of bacteriocins of food isolates Enterococcus hirae LD3 and Lactobacillus plantarum LD4 in pasteurized milk. 3 Biotech 9(1). https://doi.org/10.1007/s13205-018-1546-y
Zhao D, Wang Q, Lu F, Bie X, Zhao H, Lu Z et al (2022) A novel plantaricin 827 effectively inhibits Staphylococcus aureus and extends shelf life of skim milk. Lwt-Food Sci Technol. https://doi.org/10.1016/j.lwt.2021.112849
Li Y, Yu S, Weng P, Wu Z, Liu Y (2023) Purification and antimicrobial mechanism of a novel bacteriocin produced by Lactiplantibacillus plantarum FB-2. Lwt-Food Sci Technol. https://doi.org/10.1016/j.lwt.2023.115123
Sidhu PK, Nehra K (2021) Purification and characterization of bacteriocin Bac23 extracted from Lactobacillus plantarum PKLP5 and its interaction with silver nanoparticles for enhanced antimicrobial spectrum against food-borne pathogens. Lwt-Food Sci Technol. https://doi.org/10.1016/j.lwt.2020.110546
Lv X, Miao L, Ma H, Bai F, Lin Y, Sun M et al (2018) Purification, characterization and action mechanism of plantaricin JY22, a novel bacteriocin against Bacillus cereus produced by Lactobacillus plantarum JY22 from golden carp intestine. Food Sci Biotechnol 27(3):695–703. https://doi.org/10.1007/s10068-017-0280-2
Brookes SJ, Gabe CM (2019) Purification of developing enamel matrix proteins using preparative SDS-PAGE. Methods Mol Biol (Clifton, NJ) 1922:251–265. https://doi.org/10.1007/978-1-4939-9012-2_25
Moyer TB, Parsley NC, Sadecki PW, Schug WJ, Hicks LM (2021) Leveraging orthogonal mass spectrometry based strategies for comprehensive sequencing and characterization of ribosomal antimicrobial peptide natural products. Nat Prod Rep 38(3):489–509. https://doi.org/10.1039/d0np00046a
Wang Z, Zhang Y, Chen C, Fan S, Deng F, Zhao L (2023) A novel bacteriocin isolated from Lactobacillus plantarum W3–2 and its biological characteristics. Front Nutr. https://doi.org/10.3389/fnut.2022.1111880
Gong HS, Meng XC, Wang H (2010) Plantaricin MG active against Gram-negative bacteria produced by Lactobacillus plantarum KLDS1.0391 isolated from “Jiaoke”, a traditional fermented cream from China. Food Control 21(1):89–96. https://doi.org/10.1016/j.foodcont.2009.04.005
Hata T, Tanaka R, Ohmomo S (2010) Isolation and characterization of plantaricin ASM1: a new bacteriocin produced by Lactobacillus plantarum A-1. Int J Food Microbiol 137(1):94–99. https://doi.org/10.1016/j.ijfoodmicro.2009.10.021
Pei J, Li X, Han H, Tao Y (2018) Purification and characterization of plantaricin SLG1, a novel bacteriocin produced by Lb. plantarum isolated from yak cheese. Food Control 84:111–117. https://doi.org/10.1016/j.foodcont.2017.07.034
Stevens KA, Sheldon BW, Klapes NA, Klaenhammer TR (1991) Nisin treatment for inactivation of Salmonella species and other gram-negative bacteria. Appl Environ Microbiol 57(12):3613–3615. https://doi.org/10.1128/aem.57.12.3613-3615.1991
Jimenez-Diaz R, Rios-Sanchez RM, Desmazeaud M, Ruiz-Barba JL, Piard JC (1993) Plantaricins S and T, two new bacteriocins produced by Lactobacillus plantarum LPCO10 isolated from a green olive fermentation. Appl Environ Microbiol 59(5):1416–1424
Gonzalez B, Arca P, Mayo B, Suarez JE (1994) Detection, purification, and partial characterization of plantaricin C, a bacteriocin produced by a Lactobacillus plantarum strain of dairy origin. Appl Environ Microbiol 60(6):2158–2163
Holo H, Jeknic Z, Daeschel M, Stevanovic S, Nes IF (2001) Plantaricin W from Lactobacillus plantarum belongs to a new family of two-peptide lantibiotics. Microbiology-Sgm 147:643–651. https://doi.org/10.1099/00221287-147-3-643
Gong H-S, Meng X-C, Wang H (2010) Mode of action of plantaricin MG, a bacteriocin active against Salmonella typhimurium. J Basic Microbiol 50:S37–S45. https://doi.org/10.1002/jobm.201000130
Wang H, Zhang H, Zhang H, Jin J, Xie Y (2021) Outer membrane channel protein TolC regulates Escherichia coli K12 Sensitivity to plantaricin BM-1 via the CpxR/CpxA two-component regulatory system. Probiotics Aniicrob Proteins 13(1):238–248. https://doi.org/10.1007/s12602-020-09671-6
Pal G, Srivastava S (2014) Inhibitory effect of plantaricin peptides (Pln E/F and J/K) against Escherichia coli. World J Microbiol Biotechnol 30(11):2829–2837. https://doi.org/10.1007/s11274-014-1708-y
Zdolec N, Hadziosmanovic M, Kozacinski L, Cvrtila Z, Filipovic I, Skrivanko M et al (2008) Microbial and physicochemical succession in fermented sausages produced with bacteriocinogenic culture of Lactobacillus sakei and semi-purified bacteriocin mesenterocin Y. Meat Sci 80(2):480–487. https://doi.org/10.1016/j.meatsci.2008.01.012
Kozacinski L, Zdolec N, Hadziosmanovic M, Cvrtila Z, Filipovic I, Majic T (2006) Microbial flora of the Croatian traditionally fermented sausage. Archiv Fur Lebensmittelhygiene 57(5):141–147
Funding
This work was supported by the National Key R&D Program of China (2020YFD0900905) and the cooperation collaborative education project on the Integration of Industry, Education, and Research of Higher Education Department of the Ministry of Education (201902187007).
Author information
Authors and Affiliations
Contributions
Hongbiao Li: writing—original draft, writing—review and editing, and data curation. Yongqi Yang: methodology, investigation, and data curation. Lanxin Li: methodology, investigation, and data curation Huojian Zheng: methodology and investigation. Zhiguo Xiong: investigation and data curation. Junjie Hou: investigation and formal analysis. Liping Wang: resources, project administration, and funding acquisition.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Li, H., Yang, Y., Li, L. et al. Genome-Based Identification and Characterization of Bacteriocins Selectively Inhibiting Staphylococcus aureus in Fermented Sausages. Probiotics & Antimicro. Prot. (2024). https://doi.org/10.1007/s12602-024-10240-4
Accepted:
Published:
DOI: https://doi.org/10.1007/s12602-024-10240-4